Orphan Nuclear Receptor ERRγ Is a Novel Transcriptional Regulator of IL-6 Mediated Hepatic BMP6 Gene Expression in Mice

 ,
, 
 , ,
, ,  , and
, and 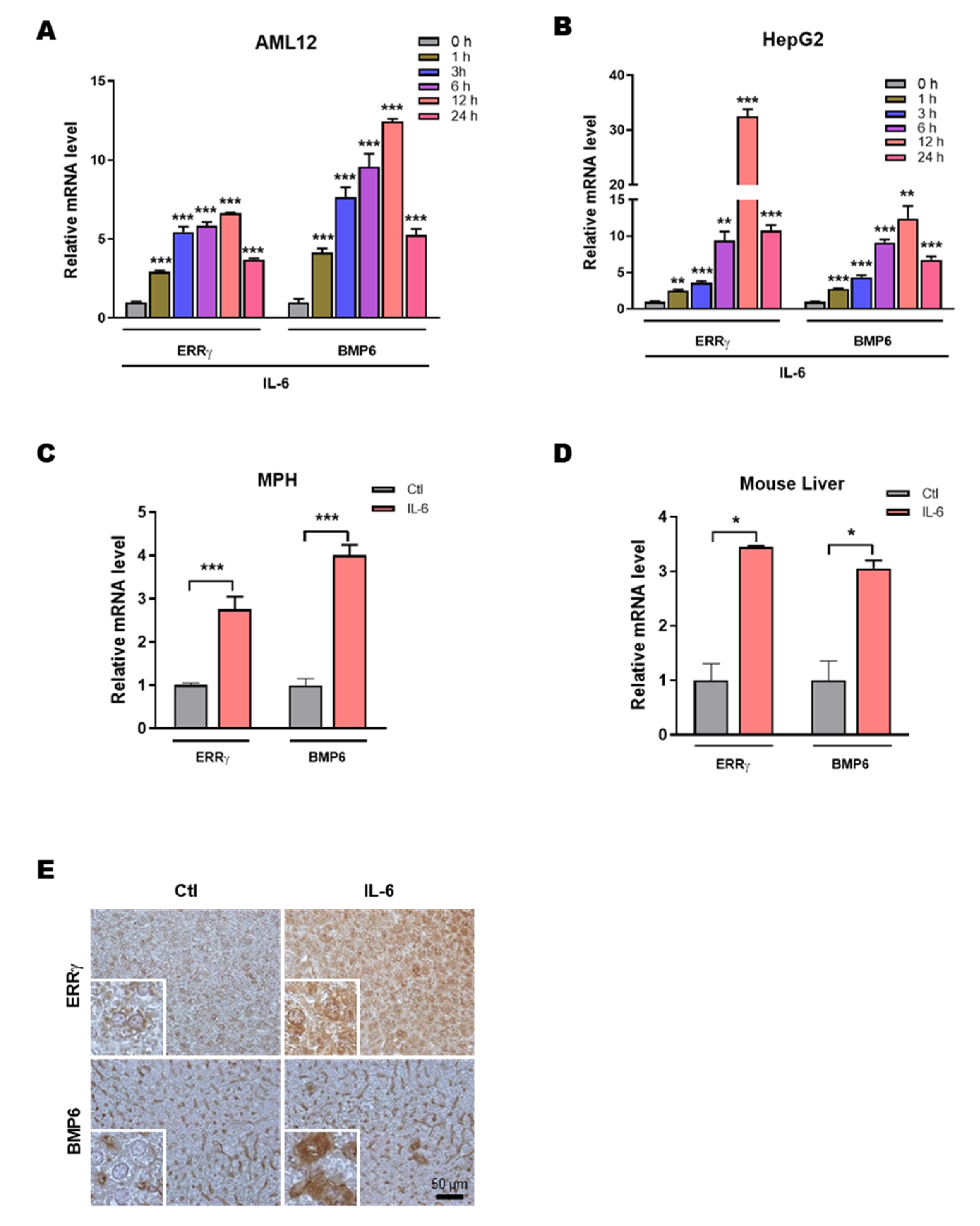{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. IL-6 Induces Hepatic ERRγ and BMP6 Gene Expression
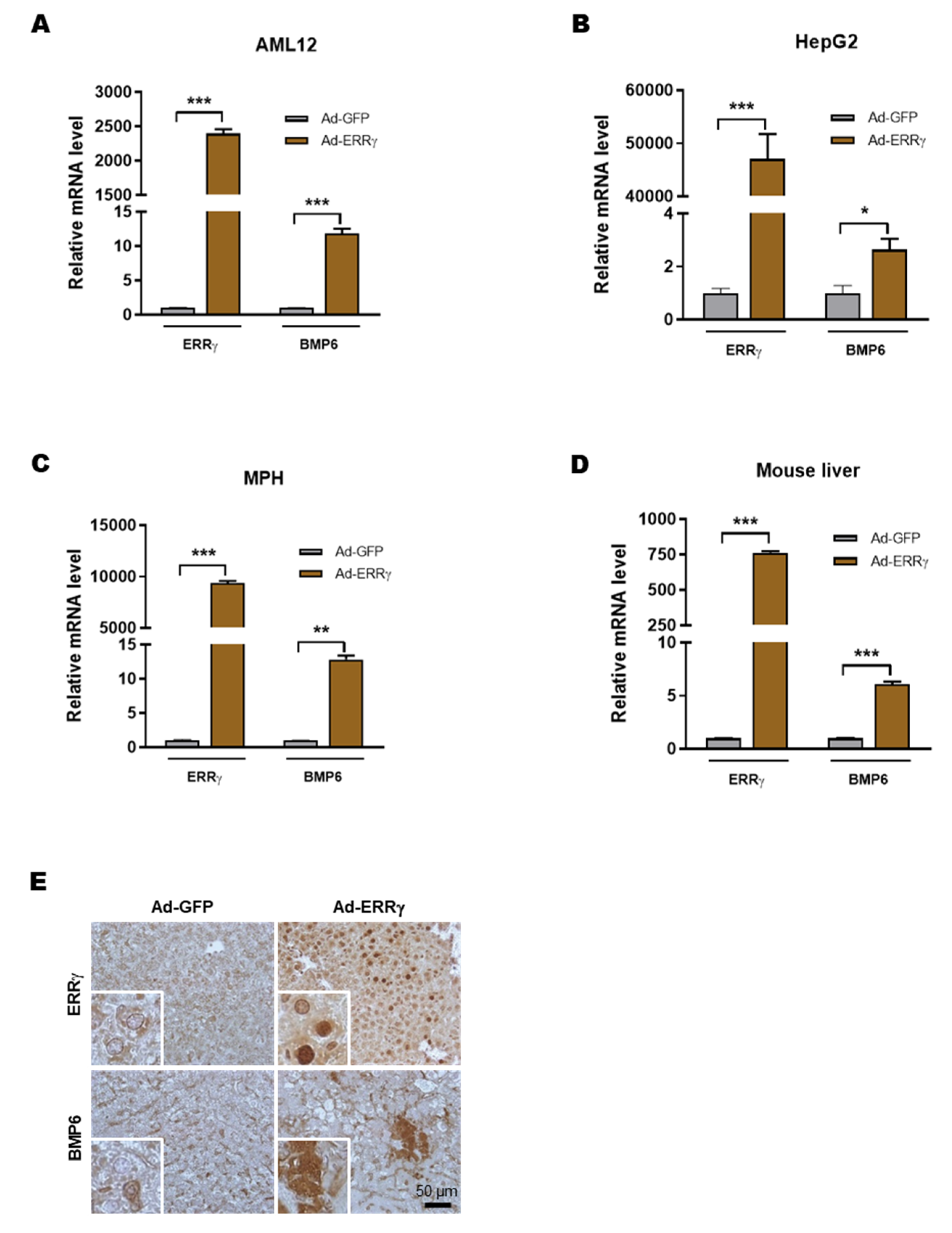
2.2. Overexpression of ERRγ Increases BMP6 Gene Expression in Liver
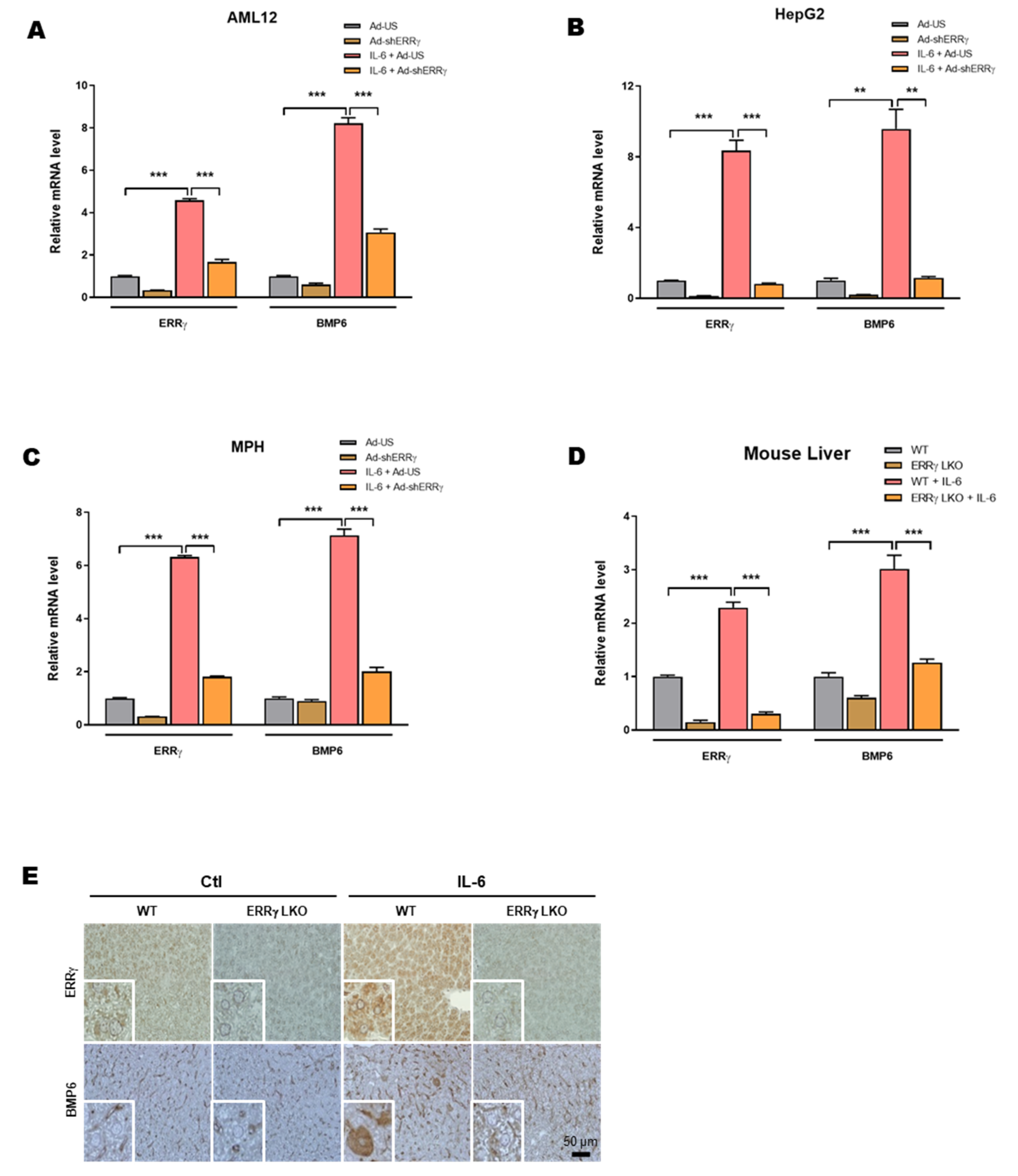
2.3. Liver Specific ERRγ Knock-Out Mice Fail to Increase Hepatic BMP6 Gene Expression in Response to IL-6 Treatment
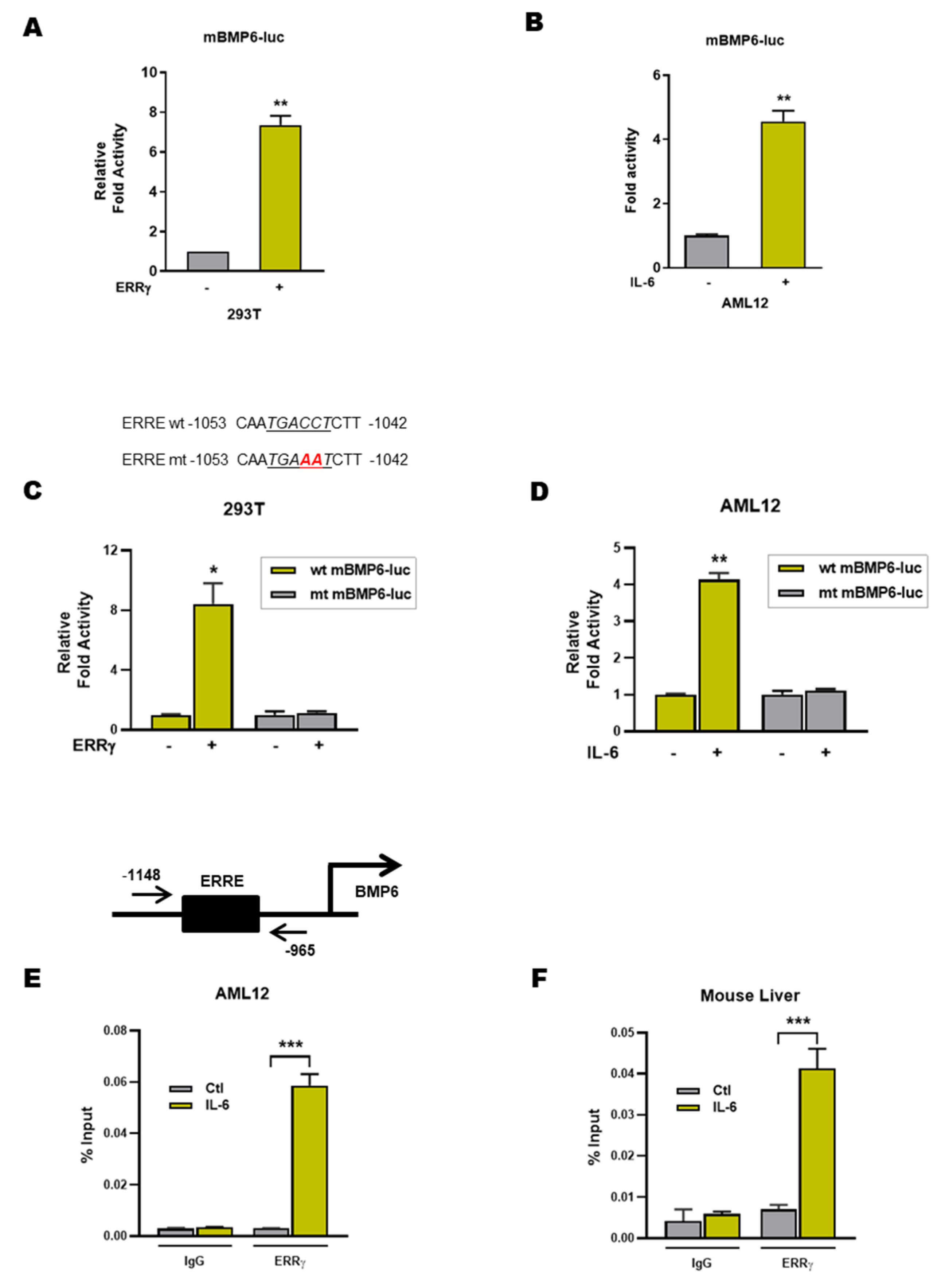
2.4. ERRγ Activates the BMP6 Gene Promoter
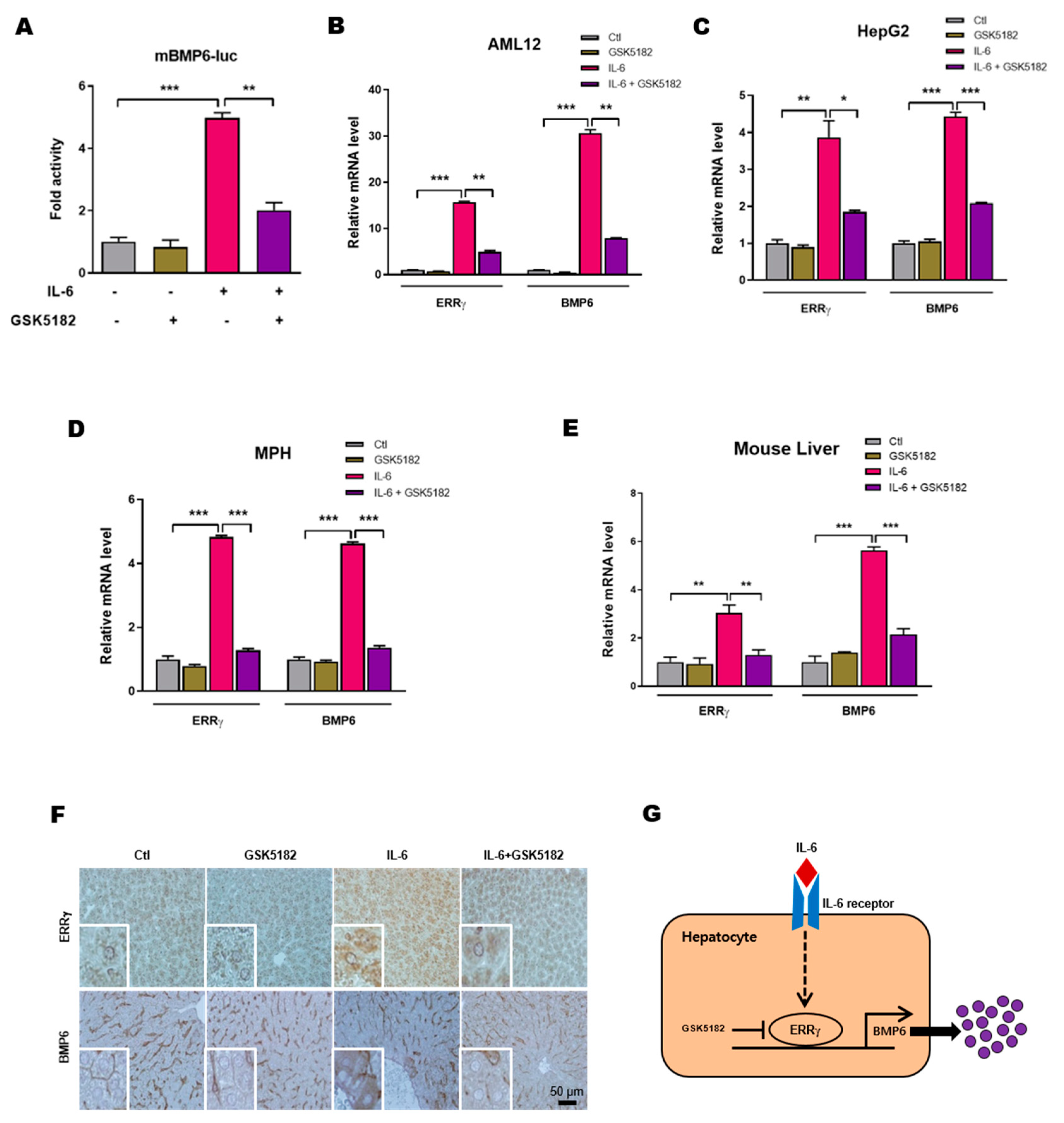
2.5. An Inverse Agonist of ERRγ Inhibits IL-6 Induced Hepatic BMP6 Gene Expression
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. Cell Culture, Transient Transfection and Luciferase Assay
4.3. Isolation of Primary Hepatocytes
4.4. Chemicals and Antibodies
4.5. DNA Cloning
4.6. Quantitative PCR
4.7. Chromatin Immunoprecipitation (ChIP) Assay
4.8. Immunohistochemistry
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMP6 | Bone morphogenetic protein 6 |
| ChIP | Chromatin immunoprecipitation |
| ERRγ | Estrogen-related receptor γ |
| ERRE | ERR responsive element |
| IL-6 | Interleukin 6 |
| NPCs | Non-parenchymal cells |
| 4-OHT | 4-hydroxytamoxifen |
References
- Bragdon, B.; Moseychuk, O.; Saldanha, S.; King, D.; Julian, J.; Nohe, A. Bone morphogenetic proteins: A critical review. Cell. Signal. 2011, 23, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Attisano, L. The TGFbeta superfamily signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, T.; Watabe, T. Bone morphogenetic proteins. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.Q. Consequences of knocking out BMP signaling in the mouse. Genesis 2003, 35, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Sykaras, N.; Opperman, L.A. Bone morphogenetic proteins (BMPs): How do they function and what can they offer the clinician? J. Oral Sci. 2003, 45, 57–73. [Google Scholar] [CrossRef]
- Simic, P.; Vukicevic, S. Bone morphogenetic proteins in development and homeostasis of kidney. Cytokine Growth Factor Rev. 2005, 16, 299–308. [Google Scholar] [CrossRef]
- Oumi, N.; Taniguchi, K.A.; Kanai, A.M.; Yasunaga, M.; Nakanishi, T.; Sato, K. A crucial role of bone morphogenetic protein signaling in the wound healing response in acute liver injury induced by carbon tetrachloride. Int. J. Hepatol. 2012, 2012, 476820. [Google Scholar] [CrossRef]
- Wu, M.; Chen, G.; Li, Y.P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Umulis, D.; O’Connor, M.B.; Blair, S.S. The extracellular regulation of bone morphogenetic protein signaling. Development 2009, 136, 3715–3728. [Google Scholar] [CrossRef]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-β/BMP signaling and other molecular events: Regulation of osteoblastogenesis and bone formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef]
- Celeste, A.J.; Iannazzi, J.A.; Taylor, R.C.; Hewick, R.M.; Rosen, V.; Wang, E.A.; Wozney, J.M. Identification of transforming growth factor beta family members present in bone-inductive protein purified from bovine bone. Proc. Natl. Acad. Sci. USA 1990, 87, 9843–9847. [Google Scholar] [CrossRef] [PubMed]
- Gitelman, S.E.; Kobrin, M.S.; Ye, J.Q.; Lopez, A.R.; Lee, A.; Derynck, R. Recombinant Vgr-1/BMP-6-expressing tumors induce fibrosis and endochondral bone formation in vivo. J. Cell Biol. 1994, 126, 1595–1609. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Luo, Z.; Zhang, Z.; Yang, H.; Lai, B.; Yao, Q.; Xiao, L.; Wang, N. Homocysteine upregulates hepcidin expression through BMP6/SMAD signaling pathway in hepatocytes. Biochem. Biophys. Res. Commun. 2016, 471, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Benn, A.; Hiepen, C.; Osterland, M.; Schutte, C.; Zwijsen, A.; Knaus, P. Role of bone morphogenetic proteins in sprouting angiogenesis: Differential BMP receptor-dependent signaling pathways balance stalk vs. tip cell competence. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 4720–4733. [Google Scholar] [CrossRef]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) signaling in development and human diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef]
- Wang, R.H.; Li, C.; Xu, X.; Zheng, Y.; Xiao, C.; Zerfas, P.; Cooperman, S.; Eckhaus, M.; Rouault, T.; Mishra, L.; et al. A role of SMAD4 in iron metabolism through the positive regulation of hepcidin expression. Cell Metab. 2005, 2, 399–409. [Google Scholar] [CrossRef]
- Babitt, J.L.; Huang, F.W.; Wrighting, D.M.; Xia, Y.; Sidis, Y.; Samad, T.A.; Campagna, J.A.; Chung, R.T.; Schneyer, A.L.; Woolf, C.J.; et al. Bone morphogenetic protein signaling by hemojuvelin regulates hepcidin expression. Nat. Genet. 2006, 38, 531–539. [Google Scholar] [CrossRef]
- Andriopoulos, B., Jr.; Corradini, E.; Xia, Y.; Faasse, S.A.; Chen, S.; Grgurevic, L.; Knutson, M.D.; Pietrangelo, A.; Vukicevic, S.; Lin, H.Y.; et al. BMP6 is a key endogenous regulator of hepcidin expression and iron metabolism. Nat. Genet. 2009, 41, 482–487. [Google Scholar] [CrossRef]
- Tremblay, A.M.; Giguere, V. The NR3B subgroup: An ovERRview. Nucl. Recept. Signal. 2007, 5, e009. [Google Scholar] [CrossRef]
- Deblois, G.; Giguere, V. Nuclear receptor location analyses in mammalian genomes: From gene regulation to regulatory networks. Mol. Endocrinol. 2008, 22, 1999–2011. [Google Scholar] [CrossRef] [PubMed]
- Eichner, L.J.; Giguere, V. Estrogen related receptors (ERRs): A new dawn in transcriptional control of mitochondrial gene networks. Mitochondrion 2011, 11, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Giguere, V.; Yang, N.; Segui, P.; Evans, R.M. Identification of a new class of steroid hormone receptors. Nature 1988, 331, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Eudy, J.D.; Yao, S.; Weston, M.D.; Ma-Edmonds, M.; Talmadge, C.B.; Cheng, J.J.; Kimberling, W.J.; Sumegi, J. Isolation of a gene encoding a novel member of the nuclear receptor superfamily from the critical region of Usher syndrome type IIa at 1q41. Genomics 1998, 50, 382–384. [Google Scholar] [CrossRef] [PubMed]
- Lui, K.; Huang, Y.; Choi, H.L.; Yu, S.; Wong, K.B.; Chen, S.; Chan, F.L. Molecular cloning and functional study of rat estrogen receptor-related receptor γ in rat prostatic cells. Prostate 2006, 66, 1600–1619. [Google Scholar] [CrossRef]
- Xie, Y.B.; Park, J.H.; Kim, D.K.; Hwang, J.H.; Oh, S.; Park, S.B.; Shong, M.; Lee, I.K.; Choi, H.S. Transcriptional corepressor SMILE recruits SIRT1 to inhibit nuclear receptor estrogen receptor-related receptor γ transactivation. J. Biol. Chem. 2009, 284, 28762–28774. [Google Scholar] [CrossRef]
- Misra, J.; Kim, D.K.; Choi, H.S. ERRγ: A junior orphan with a senior role in metabolism. Trends Endocrinol. Metab. TEM 2017, 28, 261–272. [Google Scholar] [CrossRef]
- Kim, D.K.; Kim, J.R.; Koh, M.; Kim, Y.D.; Lee, J.M.; Chanda, D.; Park, S.B.; Min, J.J.; Lee, C.H.; Park, T.S.; et al. Estrogen-related receptor γ (ERRγ) is a novel transcriptional regulator of phosphatidic acid phosphatase, LIPIN1, and inhibits hepatic insulin signaling. J. Biol. Chem. 2011, 286, 38035–38042. [Google Scholar] [CrossRef]
- Kim, D.K.; Ryu, D.; Koh, M.; Lee, M.W.; Lim, D.; Kim, M.J.; Kim, Y.H.; Cho, W.J.; Lee, C.H.; Park, S.B.; et al. Orphan nuclear receptor estrogen-related receptor γ (ERRγ) is key regulator of hepatic gluconeogenesis. J. Biol. Chem. 2012, 287, 21628–21639. [Google Scholar] [CrossRef]
- Kim, D.K.; Gang, G.T.; Ryu, D.; Koh, M.; Kim, Y.N.; Kim, S.S.; Park, J.; Kim, Y.H.; Sim, T.; Lee, I.K.; et al. Inverse agonist of nuclear receptor ERRγ mediates antidiabetic effect through inhibition of hepatic gluconeogenesis. Diabetes 2013, 62, 3093–3102. [Google Scholar] [CrossRef][Green Version]
- Kim, D.K.; Kim, Y.H.; Jang, H.H.; Park, J.; Kim, J.R.; Koh, M.; Jeong, W.I.; Koo, S.H.; Park, T.S.; Yun, C.H.; et al. Estrogen-related receptor γ controls hepatic CB1 receptor-mediated CYP2E1 expression and oxidative liver injury by alcohol. Gut 2013, 62, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Jeong, J.H.; Lee, J.M.; Kim, K.S.; Park, S.H.; Kim, Y.D.; Koh, M.; Shin, M.; Jung, Y.S.; Kim, H.S.; et al. Inverse agonist of estrogen-related receptor γ controls Salmonella typhimurium infection by modulating host iron homeostasis. Nat. Med. 2014, 20, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Chao, E.Y.; Collins, J.L.; Gaillard, S.; Miller, A.B.; Wang, L.; Orband-Miller, L.A.; Nolte, R.T.; McDonnell, D.P.; Willson, T.M.; Zuercher, W.J. Structure-guided synthesis of tamoxifen analogs with improved selectivity for the orphan ERRγ. Bioorg. Med. Chem. Lett. 2006, 16, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Woo, S.Y.; Im, C.Y.; Yoo, E.K.; Lee, S.; Kim, H.J.; Hwang, H.J.; Cho, J.H.; Lee, W.S.; Yoon, H.; et al. Insights of a lead optimization study and biological evaluation of novel 4-hydroxytamoxifen analogs as estrogen-related receptor γ (ERRγ) inverse agonists. J. Med. Chem. 2016, 59, 10209–10227. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Song, J.; Ji, H.D.; Yoo, E.K.; Lee, J.E.; Lee, S.B.; Oh, J.M.; Lee, S.; Hwang, J.S.; Yoon, H.; et al. Discovery of potent, selective, and orally bioavailable estrogen-related receptor-γ inverse agonists to restore the sodium iodide symporter function in anaplastic thyroid cancer. J. Med. Chem. 2019, 62, 1837–1858. [Google Scholar] [CrossRef] [PubMed]
- Lim, P.J.; Duarte, T.L.; Arezes, J.; Garcia-Santos, D.; Hamdi, A.; Pasricha, S.R.; Armitage, A.E.; Mehta, H.; Wideman, S.; Santos, A.G.; et al. Nrf2 controls iron homeostasis in haemochromatosis and thalassaemia via Bmp6 and hepcidin. Nat. Metab. 2019, 1, 519–531. [Google Scholar] [CrossRef]
- Steinbicker, A.U.; Sachidanandan, C.; Vonner, A.J.; Yusuf, R.Z.; Deng, D.Y.; Lai, C.S.; Rauwerdink, K.M.; Winn, J.C.; Saez, B.; Cook, C.M.; et al. Inhibition of bone morphogenetic protein signaling attenuates anemia associated with inflammation. Blood 2011, 117, 4915–4923. [Google Scholar] [CrossRef]
- Kautz, L.; Besson-Fournier, C.; Meynard, D.; Latour, C.; Roth, M.P.; Coppin, H. Iron overload induces BMP6 expression in the liver but not in the duodenum. Haematologica 2011, 96, 199–203. [Google Scholar] [CrossRef]
- Enns, C.A.; Ahmed, R.; Wang, J.; Ueno, A.; Worthen, C.; Tsukamoto, H.; Zhang, A.S. Increased iron loading induces Bmp6 expression in the non-parenchymal cells of the liver independent of the BMP-signaling pathway. PLoS ONE 2013, 8, e60534. [Google Scholar] [CrossRef][Green Version]
- Knittel, T.; Fellmer, P.; Müller, L.; Ramadori, G. Bone morphogenetic protein-6 is expressed in nonparenchymal liver cells and upregulated by transforming growth factor-beta 1. Exp. Cell Res. 1997, 232, 263–269. [Google Scholar] [CrossRef]
- Arndt, S.; Wacker, E.; Dorn, C.; Koch, A.; Saugspier, M.; Thasler, W.E.; Hartmann, A.; Bosserhoff, A.K.; Hellerbrand, C. Enhanced expression of BMP6 inhibits hepatic fibrosis in non-alcoholic fatty liver disease. Gut 2015, 64, 973–981. [Google Scholar] [CrossRef]
- Rausa, M.; Pagani, A.; Nai, A.; Campanella, A.; Gilberti, M.E.; Apostoli, P.; Camaschella, C.; Silvestri, L. Bmp6 expression in murine liver non parenchymal cells: A mechanism to control their high iron exporter activity and protect hepatocytes from iron overload? PLoS ONE 2015, 10, e0122696. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.D.; Park, K.G.; Lee, Y.S.; Park, Y.Y.; Kim, D.K.; Nedumaran, B.; Jang, W.G.; Cho, W.J.; Ha, J.; Lee, I.K.; et al. Metformin inhibits hepatic gluconeogenesis through AMP-activated protein kinase-dependent regulation of the orphan nuclear receptor SHP. Diabetes 2008, 57, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.H.; Satoh, H.; Herzig, S.; Lee, C.H.; Hedrick, S.; Kulkarni, R.; Evans, R.M.; Olefsky, J.; Montminy, M. PGC-1 promotes insulin resistance in liver through PPAR-alpha-dependent induction of TRB-3. Nat. Med. 2004, 10, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.D.; Song, J.; Kim, J.; Chin, J.; Ji, H.D.; Lee, J.-E.; Lee, S.B.; Yoon, H.; Yu, J.H.; Kim, S.K.; et al. A novel orally active inverse agonist of estrogen-related receptor γ (ERRγ), DN200434, A booster of NIS in anaplastic thyroid cancer. Clin. Cancer Res. 2019, 25, 5069–5081. [Google Scholar] [CrossRef] [PubMed]
- Alaynick, W.A.; Way, J.M.; Wilson, S.A.; Benson, W.G.; Pei, L.; Downes, M.; Yu, R.; Jonker, J.W.; Holt, J.A.; Rajpal, D.K.; et al. ERRγ regulates cardiac, gastric, and renal potassium homeostasis. Mol. Endocrinol. 2010, 24, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Kim, Y.H.; Hynx, D.; Wang, Y.; Yang, K.J.; Ryu, D.; Kim, K.S.; Yoo, E.K.; Kim, J.S.; Koo, S.H.; et al. PKB/Akt phosphorylation of ERRγ contributes to insulin-mediated inhibition of hepatic gluconeogenesis. Diabetologia 2014, 57, 2576–2585. [Google Scholar] [CrossRef][Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radhakrishnan, K.; Kim, Y.-H.; Jung, Y.S.; Kim, J.; Kim, D.-K.; Cho, S.J.; Lee, I.-K.; Dooley, S.; Lee, C.-H.; Choi, H.-S. Orphan Nuclear Receptor ERRγ Is a Novel Transcriptional Regulator of IL-6 Mediated Hepatic BMP6 Gene Expression in Mice. Int. J. Mol. Sci. 2020, 21, 7148. https://doi.org/10.3390/ijms21197148
Radhakrishnan K, Kim Y-H, Jung YS, Kim J, Kim D-K, Cho SJ, Lee I-K, Dooley S, Lee C-H, Choi H-S. Orphan Nuclear Receptor ERRγ Is a Novel Transcriptional Regulator of IL-6 Mediated Hepatic BMP6 Gene Expression in Mice. International Journal of Molecular Sciences. 2020; 21(19):7148. https://doi.org/10.3390/ijms21197148
Chicago/Turabian StyleRadhakrishnan, Kamalakannan, Yong-Hoon Kim, Yoon Seok Jung, Jina Kim, Don-Kyu Kim, Sung Jin Cho, In-Kyu Lee, Steven Dooley, Chul-Ho Lee, and Hueng-Sik Choi. 2020. "Orphan Nuclear Receptor ERRγ Is a Novel Transcriptional Regulator of IL-6 Mediated Hepatic BMP6 Gene Expression in Mice" International Journal of Molecular Sciences 21, no. 19: 7148. https://doi.org/10.3390/ijms21197148
APA StyleRadhakrishnan, K., Kim, Y.-H., Jung, Y. S., Kim, J., Kim, D.-K., Cho, S. J., Lee, I.-K., Dooley, S., Lee, C.-H., & Choi, H.-S. (2020). Orphan Nuclear Receptor ERRγ Is a Novel Transcriptional Regulator of IL-6 Mediated Hepatic BMP6 Gene Expression in Mice. International Journal of Molecular Sciences, 21(19), 7148. https://doi.org/10.3390/ijms21197148