Alternative Splicing Regulation of an Alzheimer’s Risk Variant in CLU



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Regulatory Function of rs7982 in Splicing
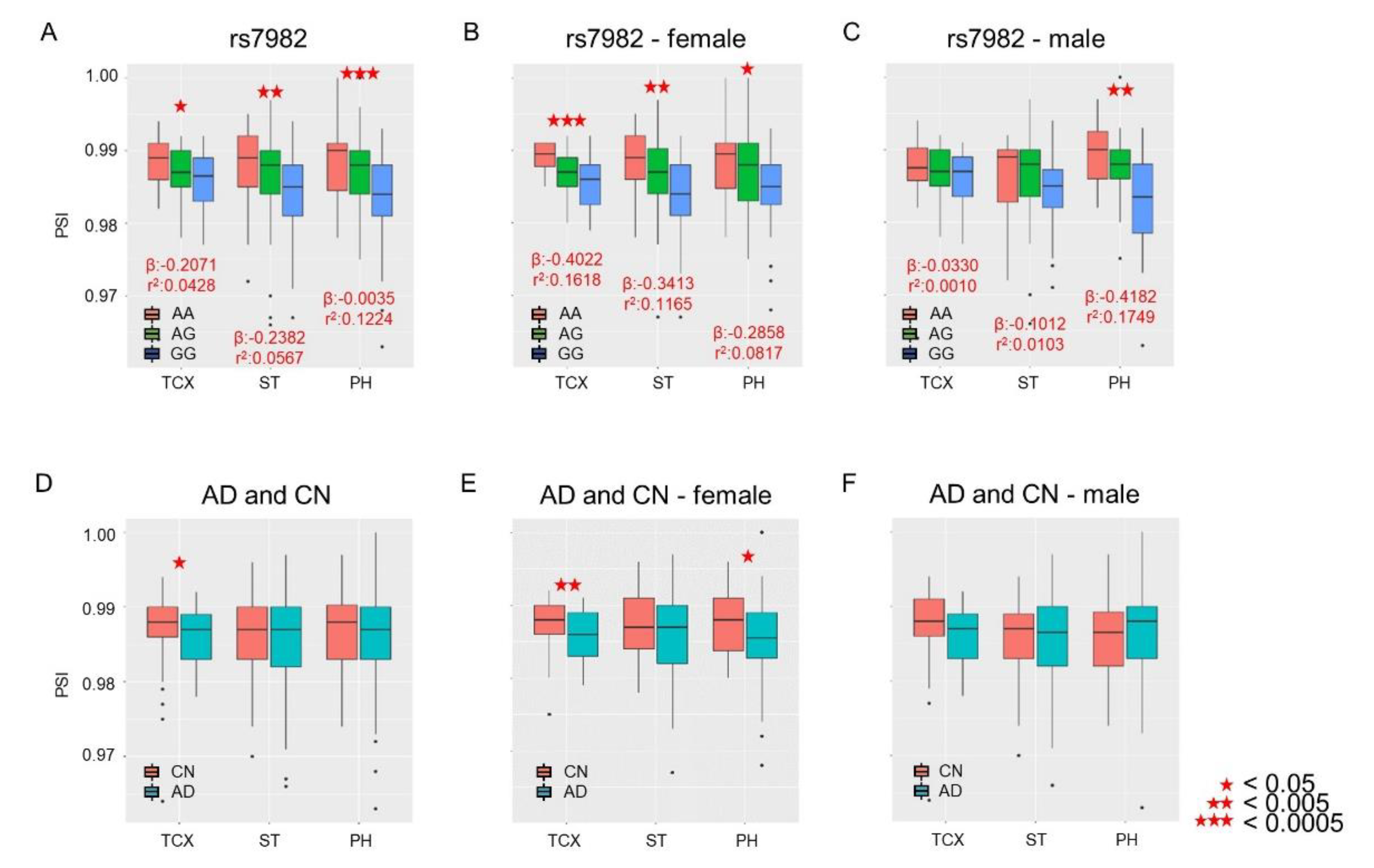
2.2. Association of Intron Retention (IR) with rs7982
2.3. Sex-Dependent Association of IR with rs7982
3. Discussion
4. Materials and Methods
4.1. RNA-Seq and WGS Analysis
4.2. Implication of rs7982 in Splicing Regulation
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TCX | Temporal cortex |
| ST | Superior temporal gyrus |
| PH | Parahippocampal gyrus |
| OR | Odds ratio |
| AD | Alzheimer’s disease |
| AMP-AD | Accelerating Medicines Partnership–Alzheimer’s Disease |
| ESE | Exon skipping enhancer |
| IR | Intron retention |
| AS | Alternative splicing |
| sQTLs | Splicing quantitative trait loci |
References
- Buee, L.; Bussiere, T.; Buee-Scherrer, V.; Delacourte, A.; Hof, P.R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res. Rev. 2000, 33, 95–130. [Google Scholar] [CrossRef]
- Rockenstein, E.M.; McConlogue, L.; Tan, H.; Power, M.; Masliah, E.; Mucke, L. Levels and alternative splicing of amyloid beta protein precursor (APP) transcripts in brains of APP transgenic mice and humans with Alzheimer’s disease. J. Biol. Chem. 1995, 270, 28257–28267. [Google Scholar] [PubMed]
- Koch, L. Altered splicing in Alzheimer transcriptomes. Nat. Rev. Genet. 2018, 19, 738–739. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.S.; Cooper, T.A. Splicing in disease: Disruption of the splicing code and the decoding machinery. Nat. Rev. Genet. 2007, 8, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Raj, T.; Li, Y.I.; Wong, G.; Humphrey, J.; Wang, M.; Ramdhani, S.; Wang, Y.C.; Ng, B.; Gupta, I.; Haroutunian, V.; et al. Integrative transcriptome analyses of the aging brain implicate altered splicing in Alzheimer’s disease susceptibility. Nat. Genet. 2018, 50, 1584–1592. [Google Scholar] [CrossRef]
- Harold, D.; Abraham, R.; Hollingworth, P.; Sims, R.; Gerrish, A.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; Dowzell, K.; Williams, A.; et al. Genome-Wide association study identifies variants at CLU and PICALM associated with Alzheimer’s disease. Nat. Genet. 2009, 41, 1088–1093. [Google Scholar] [CrossRef]
- Yu, J.T.; Tan, L. The role of clusterin in Alzheimer’s disease: Pathways, pathogenesis, and therapy. Mol. Neurobiol. 2012, 45, 314–326. [Google Scholar] [CrossRef]
- Szymanski, M.; Wang, R.; Bassett, S.S.; Avramopoulos, D. Alzheimer’s risk variants in the clusterin gene are associated with alternative splicing. Transl. Psychiatry 2011, 1, e18. [Google Scholar] [CrossRef]
- Zhao, L.; Mao, Z.; Woody, S.K.; Brinton, R.D. Sex differences in metabolic aging of the brain: Insights into female susceptibility to Alzheimer’s disease. Neurobiol. Aging 2016, 42, 69–79. [Google Scholar] [CrossRef]
- Rohne, P.; Prochnow, H.; Wolf, S.; Renner, B.; Koch-Brandt, C. The chaperone activity of clusterin is dependent on glycosylation and redox environment. Cell Physiol. Biochem. 2014, 34, 1626–1639. [Google Scholar] [CrossRef]
- Bettens, K.; Vermeulen, S.; Van Cauwenberghe, C.; Heeman, B.; Asselbergh, B.; Robberecht, C.; Engelborghs, S.; Vandenbulcke, M.; Vandenberghe, R.; De Deyn, P.P.; et al. Reduced secreted clusterin as a mechanism for Alzheimer-associated CLU mutations. Mol. Neurodegener. 2015, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Yamamoto, M. Modification of the alternative splicing process of testosterone-repressed prostate message-2 (TRPM-2) gene by protein synthesis inhibitors and heat shock treatment. Biochim. Biophys. Acta 1996, 1307, 83–88. [Google Scholar] [CrossRef]
- Zhou, Y.; Hayashi, I.; Wong, J.; Tugusheva, K.; Renger, J.J.; Zerbinatti, C. Intracellular clusterin interacts with brain isoforms of the bridging integrator 1 and with the microtubule-associated protein Tau in Alzheimer’s disease. PLoS ONE 2014, 9, e103187. [Google Scholar] [CrossRef] [PubMed]
- Weyn-Vanhentenryck, S.M.; Feng, H.; Ustianenko, D.; Duffie, R.; Yan, Q.; Jacko, M.; Martinez, J.C.; Goodwin, M.; Zhang, X.; Hengst, U.; et al. Precise temporal regulation of alternative splicing during neural development. Nat. Commun. 2018, 9, 2189. [Google Scholar] [CrossRef] [PubMed]
- Su, C.H.; Tarn, W.Y. Alternative Splicing in Neurogenesis and Brain Development. Front. Mol. Biosci. 2018, 5, 12. [Google Scholar] [CrossRef]
- Cederholm, J.M.E.; Kim, Y.; von Jonquieres, G.; Housley, G.D. Human Brain Region-Specific Alternative Splicing of TRPC3, the Type 3 Canonical Transient Receptor Potential Non-Selective Cation Channel. Cerebellum 2019, 18, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Raj, B.; Blencowe, B.J. Alternative Splicing in the Mammalian Nervous System: Recent Insights into Mechanisms and Functional Roles. Neuron 2015, 87, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Jansen, I.; Savage, J.; Watanabe, K.; Bryois, J.; Williams, D.; Steinberg, S.; Sealock, J.; Karlsson, I.; Hagg, S.; Athanasiu, L.; et al. Genetic meta-analysis identifies 10 novel loci and functional pathways for Alzheimer’s disease risk. bioRxiv 2018, 258533. [Google Scholar] [CrossRef]
- Tan, L.; Wang, H.F.; Tan, M.S.; Tan, C.C.; Zhu, X.C.; Miao, D.; Yu, W.J.; Jiang, T.; Tan, L.; Yu, J.T.; et al. Effect of CLU genetic variants on cerebrospinal fluid and neuroimaging markers in healthy, mild cognitive impairment and Alzheimer’s disease cohorts. Sci. Rep. 2016, 6, 26027. [Google Scholar] [CrossRef] [PubMed]
- Kralovicova, J.; Houngninou-Molango, S.; Kramer, A.; Vorechovsky, I. Branch site haplotypes that control alternative splicing. Hum. Mol. Genet. 2004, 13, 3189–3202. [Google Scholar] [CrossRef] [PubMed]
- Katz, Y.; Wang, E.T.; Airoldi, E.M.; Burge, C.B. Analysis and design of RNA sequencing experiments for identifying isoform regulation. Nat. Methods 2010, 7, 1009–1015. [Google Scholar] [CrossRef]
- Allen, M.; Carrasquillo, M.M.; Funk, C.; Heavner, B.D.; Zou, F.; Younkin, C.S.; Burgess, J.D.; Chai, H.S.; Crook, J.; Eddy, J.A.; et al. Human whole genome genotype and transcriptome data for Alzheimer’s and other neurodegenerative diseases. Sci. Data 2016, 3, 160089. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Beckmann, N.D.; Roussos, P.; Wang, E.; Zhou, X.; Wang, Q.; Ming, C.; Neff, R.; Ma, W.; Fullard, J.F.; et al. The Mount Sinai cohort of large-scale genomic, transcriptomic and proteomic data in Alzheimer’s disease. Sci. Data 2018, 5, 180185. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Park, J.W.; Lu, Z.X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef]
- Lee, Y.; Gamazon, E.R.; Rebman, E.; Lee, Y.; Lee, S.; Dolan, M.E.; Cox, N.J.; Lussier, Y.A. Variants affecting exon skipping contribute to complex traits. PLoS Genet. 2012, 8, e1002998. [Google Scholar] [CrossRef]
- Smith, P.J.; Zhang, C.; Wang, J.; Chew, S.L.; Zhang, M.Q.; Krainer, A.R. An increased specificity score matrix for the prediction of SF2/ASF-specific exonic splicing enhancers. Hum. Mol Genet. 2006, 15, 2490–2508. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.; Nho, K.; Lee, Y. Alternative Splicing Regulation of an Alzheimer’s Risk Variant in CLU. Int. J. Mol. Sci. 2020, 21, 7079. https://doi.org/10.3390/ijms21197079
Han S, Nho K, Lee Y. Alternative Splicing Regulation of an Alzheimer’s Risk Variant in CLU. International Journal of Molecular Sciences. 2020; 21(19):7079. https://doi.org/10.3390/ijms21197079
Chicago/Turabian StyleHan, Seonggyun, Kwangsik Nho, and Younghee Lee. 2020. "Alternative Splicing Regulation of an Alzheimer’s Risk Variant in CLU" International Journal of Molecular Sciences 21, no. 19: 7079. https://doi.org/10.3390/ijms21197079
APA StyleHan, S., Nho, K., & Lee, Y. (2020). Alternative Splicing Regulation of an Alzheimer’s Risk Variant in CLU. International Journal of Molecular Sciences, 21(19), 7079. https://doi.org/10.3390/ijms21197079