Rho GTPase ROP1 Interactome Analysis Reveals Novel ROP1-Associated Pathways for Pollen Tube Polar Growth in Arabidopsis

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Isolation of ROP1-Associated Interacting Proteins from Arabidopsis Pollen Tubes
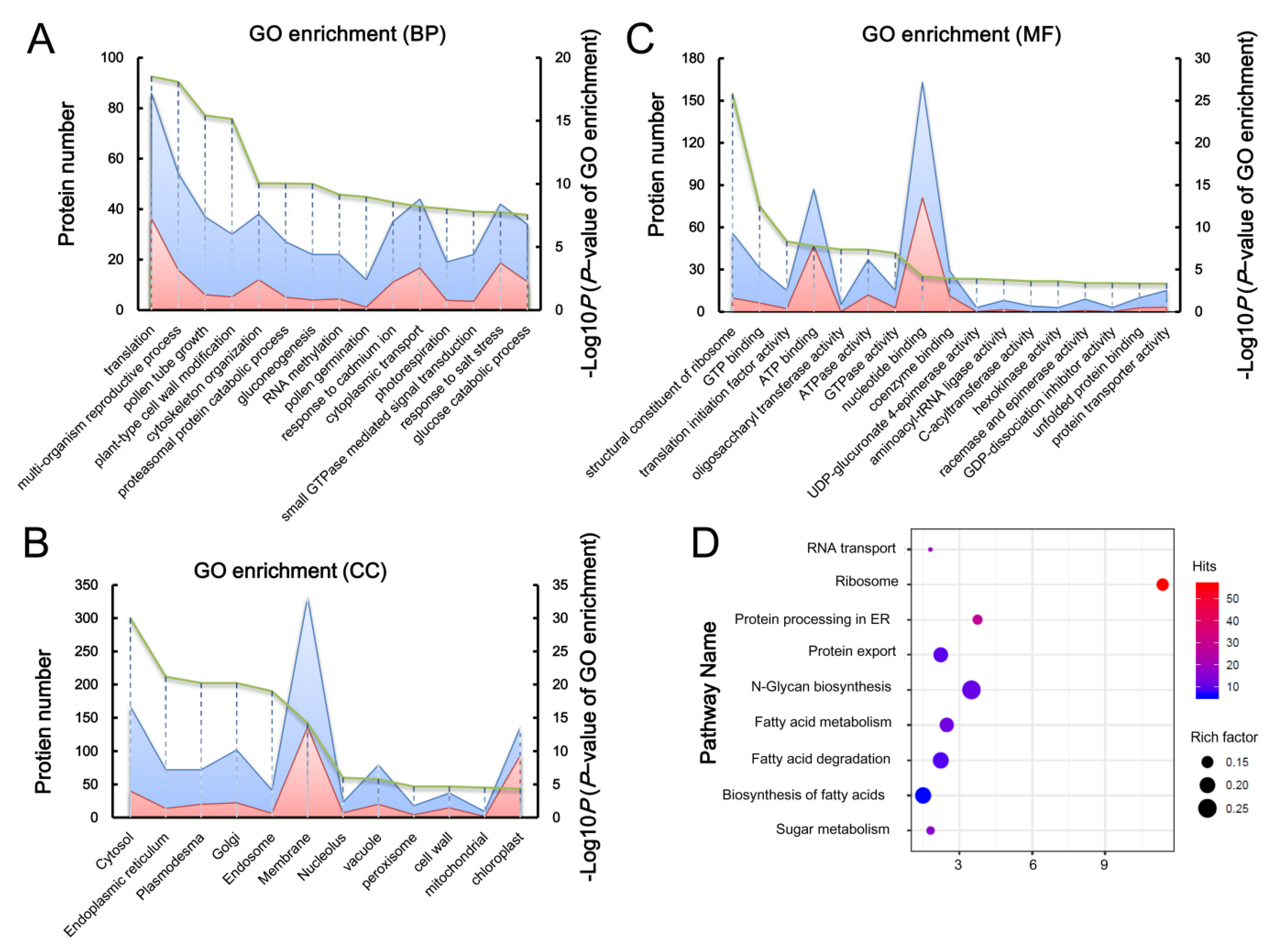
2.2. Bioinformatic Analysis of ROP1-Associated Proteins in Arabidopsis
2.3. Coexpression Assay of ROP1 and ROP1-Associated Proteins in Arabidopsis Pollen Tubes
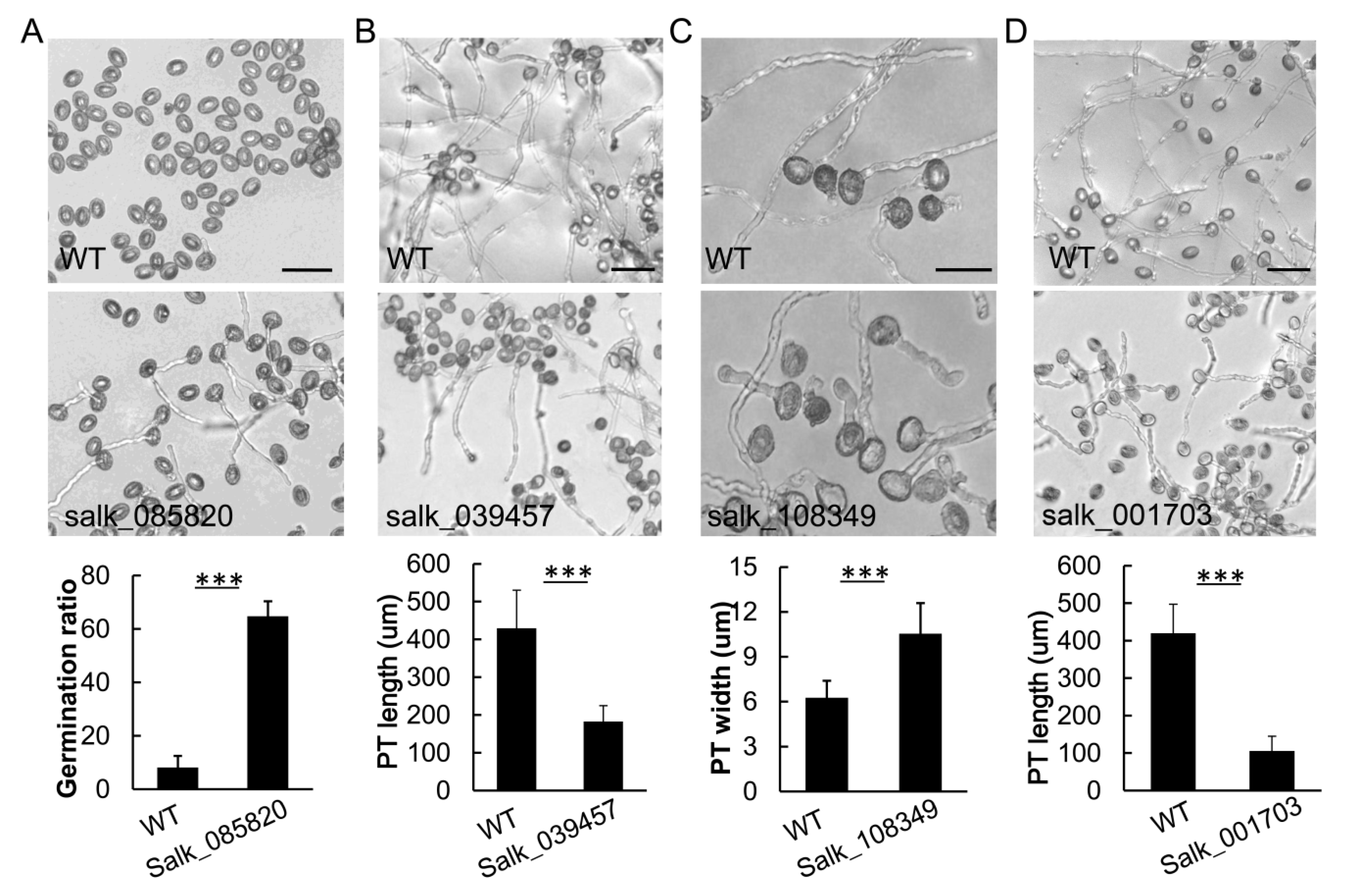
2.4. Functional Assay and Confirmation of ROP1 Interactors in Arabidopsis Pollen Tube Tip Growth
2.5. Protein Network Analysis of ROP1–Associated Proteins
3. Discussion
3.1. Cell Wall Modification and ROP1 Signaling
3.2. Post-Translational Modifications and ROP1 Signaling
3.3. Material Transport and ROP1 Signaling in Pollen Tube
3.4. ROP1 Signaling and Endocytosis
4. Material and Method
4.1. Plant Materials and Pollen Tube Cultivation
4.2. Protein Extraction and Affinity Purification of Target Proteins
4.3. Western Blots Assay
4.4. Identification of ROP1-Binding Protein by LC MS/MS
4.5. Data Processing and Analysis
4.6. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yang, Z. Cell polarity signaling in Arabidopsis. Annu. Rev. Cell Dev. Biol. 2008, 24, 551–575. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, M. Tip Growth in Filamentous Fungi: A Road Trip to the Apex. Annu. Rev. Cell Dev. Biol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Rounds, C.M.; Bezanilla, M. Growth Mechanisms in Tip-Growing Plant Cells. Annu. Rev. Cell Dev. Biol. 2013, 64, 243–265. [Google Scholar] [CrossRef] [PubMed]
- Arkowitz, R.A. Polarized growth and movement: How to generate new shapes and structures. Semin. Cell Dev. Biol. 2012, 22, 789. [Google Scholar] [CrossRef]
- Lee, Y.J.; Yang, Z. Tip growth: Signaling in the apical dome. Curr. Opin. Plant Biol. 2008, 11, 662–671. [Google Scholar] [CrossRef]
- Qin, Y.; Yang, Z. Rapid tip growth: Insights from pollen tubes. Semin. Cell Dev. Biol. 2011, 22, 816–824. [Google Scholar] [CrossRef]
- Guan, Y.; Guo, J.; Li, H.; Yang, Z. Signaling in pollen tube growth: Crosstalk, feedback, and missing links. Mol. Plant 2013, 6, 1053–1064. [Google Scholar] [CrossRef]
- Takeuchi, H.; Higashiyama, T. Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis. Nature 2016, 531, 245–248. [Google Scholar] [CrossRef]
- Kanaoka, M.M.; Higashiyama, T. Peptide signaling in pollen tube guidance. Curr. Opin. Plant Biol. 2015, 28, 127–136. [Google Scholar] [CrossRef]
- Palanivelu, R.; Brass, L.; Edlund, A.F.; Preuss, D. Pollen tube growth and guidance is regulated by POP2, an Arabidopsis gene that controls GABA levels. Cell 2003, 114, 47–59. [Google Scholar] [CrossRef]
- Cole, R.A.; Fowler, J.E. Polarized growth: Maintaining focus on the tip. Curr. Opin. Plant Biol. 2006, 9, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Scheible, N.; McCubbin, A. Signaling in Pollen Tube Growth: Beyond the Tip of the Polarity Iceberg. Plants 2019, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Yang, Z. Exocytosis and endocytosis: Coordinating and fine-tuning the polar tip growth domain in pollen tubes. J. Exp. Bot. 2020, 71, 2428–2438. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. Small GTPases: Versatile signaling switches in plants. Plant Cell 2002, 14, S375–S388. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Song, J.; Tian, X.; Zhang, H.; Li, L.; Zhu, H. Arabidopsis PRK6 interacts specifically with AtRopGEF8/12 and induces depolarized growth of pollen tubes when overexpressed. Sci. China Life Sci. 2018, 61, 100–112. [Google Scholar] [CrossRef]
- Chang, F.; Gu, Y.; Ma, H.; Yang, Z. AtPRK2 Promotes ROP1 Activation via RopGEFs in the Control of Polarized Pollen Tube Growth. Mol. Plant 2012. [Google Scholar] [CrossRef]
- Li, E.; Cui, Y.; Ge, F.R.; Chai, S.; Zhang, W.T.; Feng, Q.N.; Jiang, L.; Li, S.; Zhang, Y. AGC1.5 Kinase Phosphorylates RopGEFs to Control Pollen Tube Growth. Mol. Plant 2018, 11, 1198–1209. [Google Scholar] [CrossRef]
- Hwang, J.U.; Vernoud, V.; Szumlanski, A.; Nielsen, E.; Yang, Z. A tip-localized RhoGAP controls cell polarity by globally inhibiting Rho GTPase at the cell apex. Curr. Biol. 2008, 18, 1907–1916. [Google Scholar] [CrossRef]
- Feng, Q.N.; Kang, H.; Song, S.J.; Ge, F.R.; Zhang, Y.L.; Li, E.; Li, S.; Zhang, Y. Arabidopsis RhoGDIs Are Critical for Cellular Homeostasis of Pollen Tubes. Plant Physiol. 2016, 170, 841–856. [Google Scholar] [CrossRef]
- Hwang, J.U.; Wu, G.; Yan, A.; Lee, Y.J.; Grierson, C.S.; Yang, Z. Pollen-tube tip growth requires a balance of lateral propagation and global inhibition of Rho-family GTPase activity. J. Cell Sci. 2010, 123 Pt 3, 340–350. [Google Scholar] [CrossRef]
- Klahre, U.; Becker, C.; Schmitt, A.C.; Kost, B. Nt-RhoGDI2 regulates Rac/Rop signaling and polar cell growth in tobacco pollen tubes. Plant J. 2006, 46, 1018–1031. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, N.; Wang, W.; Liu, Z.; Chen, J.; Zhao, L.; Tan, L.; Wang, C.; Qin, Y.; Li, C.; et al. The REN4 rheostat dynamically coordinates the apical and lateral domains of Arabidopsis pollen tubes. Nat. Commun. 2018, 9, 2573. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Gu, Y.; Yan, A.; Lord, E.; Yang, Z.B. RIP1 (ROP Interactive Partner 1)/ICR1 marks pollen germination sites and may act in the ROP1 pathway in the control of polarized pollen growth. Mol. Plant 2008, 1, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Fu, Y.; Dowd, P.; Li, S.; Vernoud, V.; Gilroy, S.; Yang, Z. A Rho family GTPase controls actin dynamics and tip growth via two counteracting downstream pathways in pollen tubes. J. Cell Biol. 2005, 169, 127–138. [Google Scholar] [CrossRef]
- Lee, Y.J.; Szumlanski, A.; Nielsen, E.; Yang, Z. Rho-GTPase-dependent filamentous actin dynamics coordinate vesicle targeting and exocytosis during tip growth. J. Cell Biol. 2008, 181, 1155–1168. [Google Scholar] [CrossRef]
- Li, H.; Lin, Y.; Heath, R.M.; Zhu, M.X.; Yang, Z. Control of pollen tube tip growth by a Rop GTPase-dependent pathway that leads to tip-localized calcium influx. Plant Cell 1999, 11, 1731–1742. [Google Scholar]
- Lin, Y.; Yang, Z. Inhibition of Pollen Tube Elongation by Microinjected Anti-Rop1Ps Antibodies Suggests a Crucial Role for Rho-Type GTPases in the Control of Tip Growth. Plant Cell 1997, 9, 1647–1659. [Google Scholar] [CrossRef]
- Gu, Y.; Vernoud, V.; Fu, Y.; Yang, Z. ROP GTPase regulation of pollen tube growth through the dynamics of tip-localized F-actin. J. Exp. Bot. 2003, 54, 93–101. [Google Scholar] [CrossRef]
- Hwang, J.U.; Gu, Y.; Lee, Y.J.; Yang, Z. Oscillatory ROP GTPase activation leads the oscillatory polarized growth of pollen tubes. Mol. Biol. Cell 2005, 16, 5385–5399. [Google Scholar] [CrossRef]
- Klahre, U.; Kost, B. Tobacco RhoGTPase ACTIVATING PROTEIN1 spatially restricts signaling of RAC/Rop to the apex of pollen tubes. Plant Cell 2006, 18, 3033–3046. [Google Scholar] [CrossRef]
- Dai, S.; Chen, T.; Chong, K.; Xue, Y.; Liu, S.; Wang, T. Proteomics identification of differentially expressed proteins associated with pollen germination and tube growth reveals characteristics of germinated Oryza sativa pollen. Mol. Cell. Proteom. MCP 2007, 6, 207–230. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Gu, Y.; Li, S.; Yang, Z. A genome-wide analysis of Arabidopsis Rop-interactive CRIB motif-containing proteins that act as Rop GTPase targets. Plant Cell 2001, 13, 2841–2856. [Google Scholar] [PubMed]
- Procissi, A.; Guyon, A.; Pierson, E.S.; Giritch, A.; Knuiman, B.; Grandjean, O.; Tonelli, C.; Derksen, J.; Pelletier, G.; Bonhomme, S. KINKY POLLEN encodes a SABRE-like protein required for tip growth in Arabidopsis and conserved among eukaryotes. Plant J. 2003, 36, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Steinebrunner, I.; Wu, J.; Sun, Y.; Corbett, A.; Roux, S.J. Disruption of Apyrases Inhibits Pollen Germination in Arabidopsis. Plant Physiol. 2003, 131, 1638. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.-G.; Liang, L.; Jia, P.-F.; Wang, Y.-C.; Li, H.-J.; Yang, W.-C. Integration of ovular signals and exocytosis of a Ca2+ channel by MLOs in pollen tube guidance. Nat. Plants 2020, 6, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Han, S.; Siao, W.; Song, C.; Xiang, Y.; Wu, X.; Cheng, P.; Li, H.; Jásik, J.; Mičieta, K.; et al. Arabidopsis Synaptotagmin 2 Participates in Pollen Germination and Tube Growth and Is Delivered to Plasma Membrane via Conventional Secretion. Mol. Plant 2015, 8, 1737–1750. [Google Scholar] [CrossRef]
- Zhang, Y.; McCormick, S. The regulation of vesicle trafficking by small GTPases and phospholipids during pollen tube growth. Sex Plant Reprod. 2010, 23, 87–93. [Google Scholar] [CrossRef]
- Vogler, H.; Santos-Fernandez, G.; Mecchia, M.A.; Grossniklaus, U. To preserve or to destroy, that is the question: The role of the cell wall integrity pathway in pollen tube growth. Curr. Opin. Plant. Biol. 2019, 52, 131–139. [Google Scholar] [CrossRef]
- Chebli, Y.; Kaneda, M.; Zerzour, R.; Geitmann, A. The cell wall of the Arabidopsis pollen tube--spatial distribution, recycling, and network formation of polysaccharides. Plant Physiol. 2012, 160, 1940–1955. [Google Scholar] [CrossRef]
- Rockel, N.; Wolf, S.; Kost, B.; Rausch, T.; Greiner, S. Elaborate spatial patterning of cell-wall PME and PMEI at the pollen tube tip involves PMEI endocytosis, and reflects the distribution of esterified and de-esterified pectins. Plant J. 2008, 53, 133–143. [Google Scholar] [CrossRef]
- Bosch, M.; Cheung, A.Y.; Hepler, P.K. Pectin methylesterase, a regulator of pollen tube growth. Plant Physiol. 2005, 138, 1334–1346. [Google Scholar] [CrossRef]
- Luo, N.; Yan, A.; Liu, G.; Guo, J.; Rong, D.; Kanaoka, M.M.; Xiao, Z.; Xu, G.; Higashiyama, T.; Cui, X.; et al. Exocytosis-coordinated mechanisms for tip growth underlie pollen tube growth guidance. Nat. Commun. 2017, 8, 1687. [Google Scholar] [CrossRef] [PubMed]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Tkachenko, E.; Sabouri-Ghomi, M.; Pertz, O.; Kim, C.; Gutierrez, E.; Machacek, M.; Groisman, A.; Danuser, G.; Ginsberg, M.H. Protein kinase A governs a RhoA-RhoGDI protrusion-retraction pacemaker in migrating cells. Nat. Cell Biol. 2011, 13, 660–667. [Google Scholar] [CrossRef]
- Hotton, S.K.; Callis, J. Regulation of cullin RING ligases. Annu. Rev. Plant Biol. 2008, 59, 467–489. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Shen, Y.; Sullivan, J.A.; Rubio, V.; Xiong, Y.; Sun, T.P.; Deng, X.W. Arabidopsis CAND1, an unmodified CUL1-interacting protein, is involved in multiple developmental pathways controlled by ubiquitin/proteasome-mediated protein Degradation. Plant Cell 2004, 16, 1870–1882. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Park, J.; Choi, H.; Burla, B.; Kretzschmar, T.; Lee, Y.; Martinoia, E. Plant ABC Transporters. Arab. Book 2011, 9, e0153. [Google Scholar] [CrossRef]
- Do, T.H.T.; Choi, H.; Palmgren, M.; Martinoia, E.; Hwang, J.-U.; Lee, Y. Arabidopsis ABCG28 is required for the apical accumulation of reactive oxygen species in growing pollen tubes. Proc. Natl. Acad. Sci. USA 2019, 116, 12540. [Google Scholar] [CrossRef]
- Zhou, L.; Lan, W.; Jiang, Y.; Fang, W.; Luan, S. A Calcium-Dependent Protein Kinase Interacts with and Activates A Calcium Channel to Regulate Pollen Tube Growth. Mol. Plant 2013. [Google Scholar] [CrossRef]
- Hoffmann, R.D.; Portes, M.T.; Olsen, L.I.; Damineli, D.S.C.; Hayashi, M.; Nunes, C.O.; Pedersen, J.T.; Lima, P.T.; Campos, C.; Feijó, J.A.; et al. Plasma membrane H+-ATPases sustain pollen tube growth and fertilization. Nat. Commun. 2020, 11, 2395. [Google Scholar] [CrossRef]
- Lu, Y.; Chanroj, S.; Zulkifli, L.; Johnson, M.A.; Uozumi, N.; Cheung, A.; Sze, H. Pollen Tubes Lacking a Pair of K+ Transporters Fail to Target Ovules in Arabidopsis. Plant Cell 2011, 23, 81. [Google Scholar] [PubMed]
- Luo, N.; Yan, A.; Yang, Z. Measuring Exocytosis Rate Using Corrected Fluorescence Recovery After Photoconversion. Traffic 2016, 17, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Muro, K.; Matsuura-Tokita, K.; Tsukamoto, R.; Kanaoka, M.M.; Ebine, K.; Higashiyama, T.; Nakano, A.; Ueda, T. ANTH domain-containing proteins are required for the pollen tube plasma membrane integrity via recycling ANXUR kinases. Commun. Biol. 2018, 1, 152. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_ID | Protein Name | Function | Reference |
|---|---|---|---|
| AT3G07880 | SCN1 | Remove of membrane-distriubtion ROP1 | Feng et al., 2016 |
| AT4G24580 | REN1 | Deactivation of ROP1 | Hwang et al., 2008 |
| AT1G04450 | RIC3 | mediated ROP1-calcium signaling | Gu et al., 2005 |
| AT2G26490 | REN4 | Endocytosis of PM-localized active ROP1 | Li et al., 2018 |
| AT4G13240 | ROPGEF9 | Activation of ROP1 | Gu et al., 2006 |
| AT3G23380 | RIC5 | ROP1-depended actin dynamic | Wu et al., 2001 |
| AT1G12070 | GDI2 | Remove of membrane-distriubtion ROP1 | Hwang et al., 2010 |
| AT2G33460 | RIC1 | ROP1-mediated actin dynamic | Zhou et al., 2016 |
| AT1G62450 | GDI3 | Remove of membrane-distriubtion ROP1 | Feng et al., 2016 |
| AT3G42880 | PRK3 | activate GEF1&12 for ROP1 singaling | Takeuchi et al., 2016 |
| AT3G12690 | AGC1.5 | activate GEF1&12 for ROP1 singaling | Li et al., 2018 |
| AT1G79250 | AGC1.7 | Reduance with AGC1.5 for ROP1 singaling | Zhang et al., 2009 |
| AT5G49680 | KINKY POLLEN | Short PT | Procissi et al., 2003 |
| AT3G04080 | AtAPY1 | Inhibition of PG | Iris, et al., 2003 |
| AT1G20080 | SYT2 | Inhibition of PG and short PT | Wang, et al., 2015 |
| AT2G33670 | MLO5 | PT twist | Guo, et al., 2020 |
| AT2G01540 | Ca2+-dependent lipid-binding Protein | PT rupture | Wang, et al., 2018 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Hu, J.; Pang, J.; Zhao, L.; Yang, B.; Kang, X.; Wang, A.; Xu, T.; Yang, Z. Rho GTPase ROP1 Interactome Analysis Reveals Novel ROP1-Associated Pathways for Pollen Tube Polar Growth in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 7033. https://doi.org/10.3390/ijms21197033
Li H, Hu J, Pang J, Zhao L, Yang B, Kang X, Wang A, Xu T, Yang Z. Rho GTPase ROP1 Interactome Analysis Reveals Novel ROP1-Associated Pathways for Pollen Tube Polar Growth in Arabidopsis. International Journal of Molecular Sciences. 2020; 21(19):7033. https://doi.org/10.3390/ijms21197033
Chicago/Turabian StyleLi, Hui, Jinbo Hu, Jing Pang, Liangtao Zhao, Bing Yang, Xinlei Kang, Aimin Wang, Tongda Xu, and Zhenbiao Yang. 2020. "Rho GTPase ROP1 Interactome Analysis Reveals Novel ROP1-Associated Pathways for Pollen Tube Polar Growth in Arabidopsis" International Journal of Molecular Sciences 21, no. 19: 7033. https://doi.org/10.3390/ijms21197033
APA StyleLi, H., Hu, J., Pang, J., Zhao, L., Yang, B., Kang, X., Wang, A., Xu, T., & Yang, Z. (2020). Rho GTPase ROP1 Interactome Analysis Reveals Novel ROP1-Associated Pathways for Pollen Tube Polar Growth in Arabidopsis. International Journal of Molecular Sciences, 21(19), 7033. https://doi.org/10.3390/ijms21197033