Characterization of Integrin αIIbβ3-Mediated Outside-in Signaling by Protein Kinase Cδ in Platelets



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. The Role of Integrin αIIbβ3-Mediated Signaling in PKCδ Phosphorylation
2.2. αIIbβ3-Mediated PKCδ Phosphorylation is Regulated by Src and Pyk2
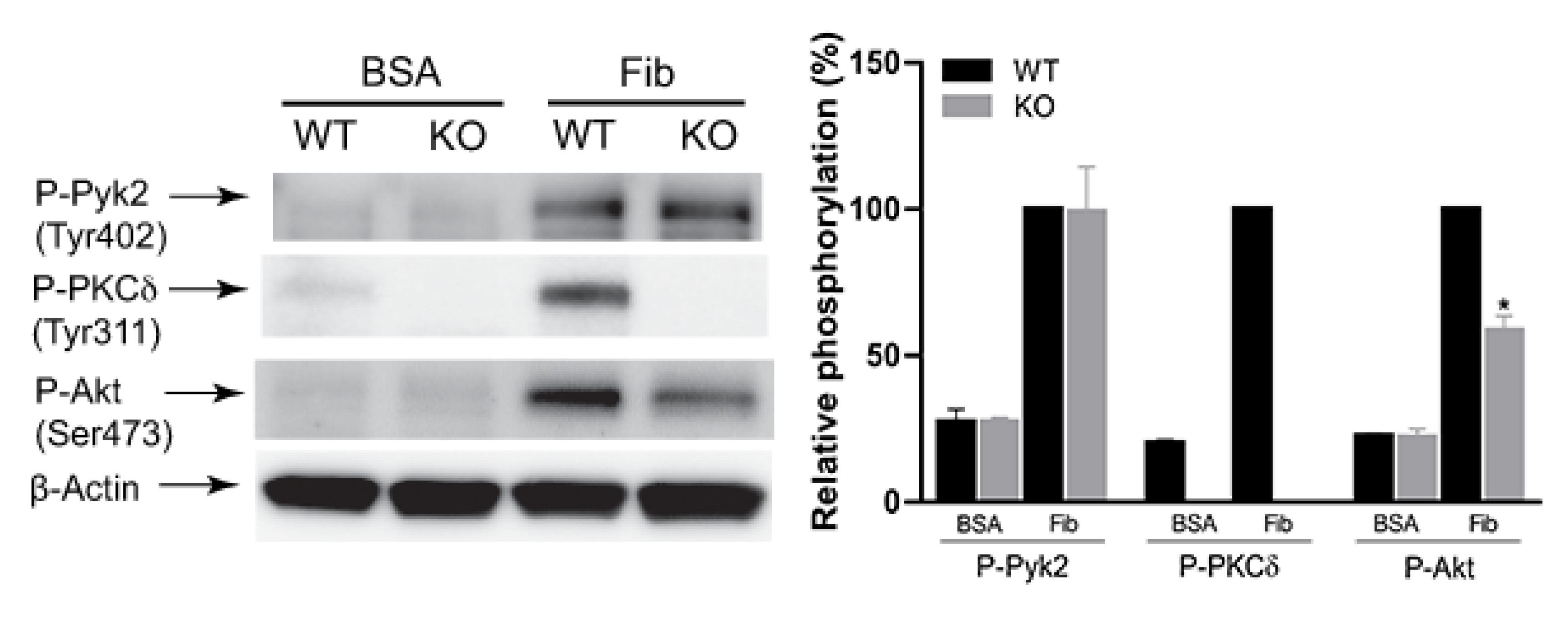
2.3. The Effect of PKCδ on αIIbβ3-Mediated Pyk2 and Akt Phosphorylation
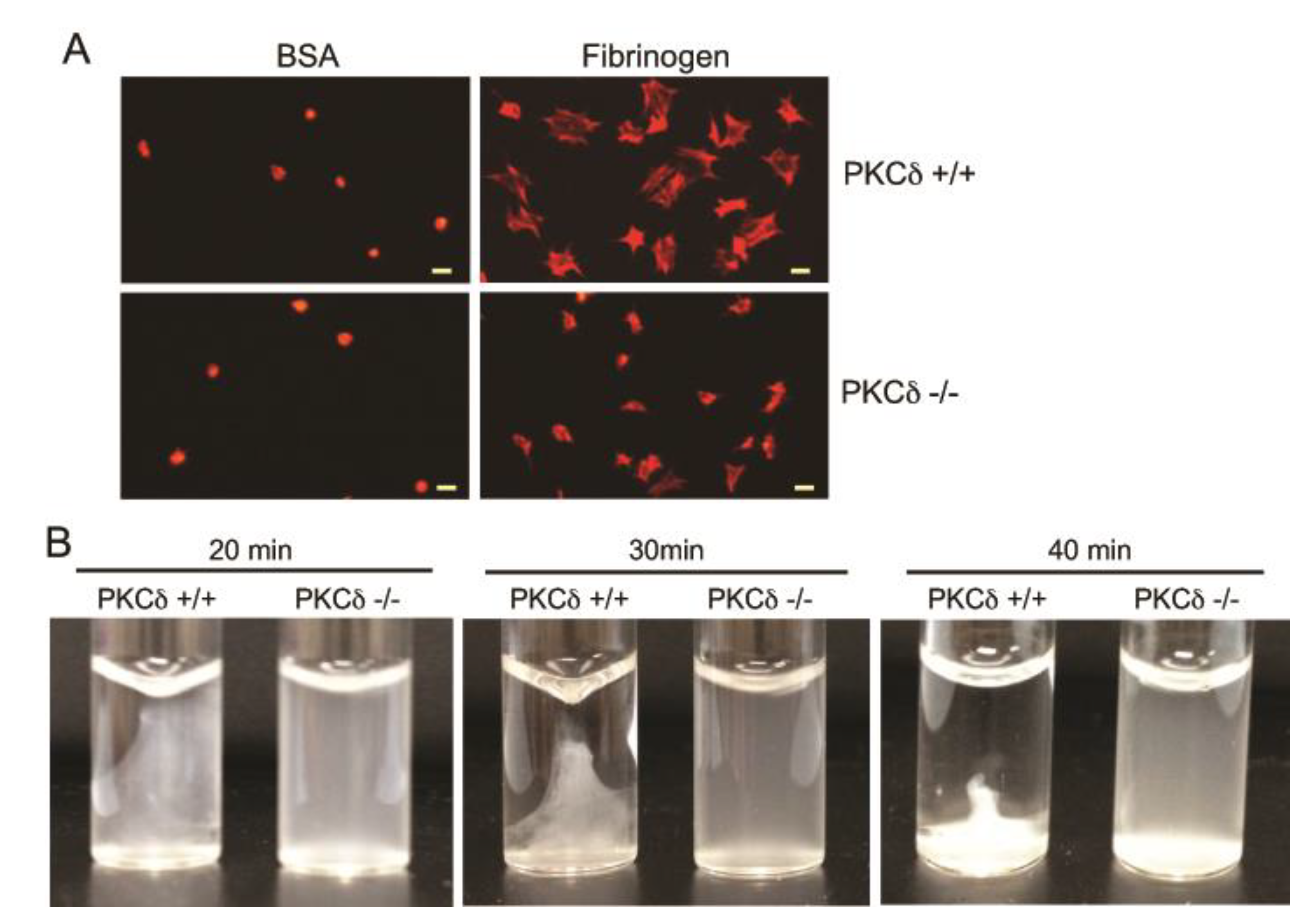
2.4. The Effect of PKCδ on Platelet Spreading and Clot Retraction
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Preparation of Murine Platelets
4.4. Western Blotting
4.5. Platelet Spreading and Clot Retraction
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PKCδ | Protein kinase C |
| Pyk2 | Proline-rich tyrosine kinase 2 |
| PLCγ2 | Phospholipase Cγ2 |
| TxA2 | Thromboxane A2 |
| PI3K | Phosphoinositide 3-kinase |
| FAK | Focal adhesion kinase |
| DAG | Diacylglycerol |
| GPVI | Glycoprotein VI |
| PAR | Protease-activated receptor |
| 2-MeSADP | 2-Methylthio-adenosine diphosphate |
| vWF | von Willebrand factor |
| SFKs | Src family kinases |
| PRP | Platelet-rich plasma |
References
- Nesbitt, W.S.; Kulkarni, S.; Giuliano, S.; Goncalves, I.; Dopheide, S.M.; Yap, C.L.; Harper, I.S.; Salem, H.H.; Jackson, S.P. Distinct glycoprotein Ib/V/IX and integrin αIIbβ3-dependent calcium signals cooperatively regulate platelet adhesion under flow. J. Biol. Chem. 2002, 277, 2965–2972. [Google Scholar] [CrossRef]
- Leisner, T.M.; Wencel-Drake, J.D.; Wang, W.; Lam, S.C.-T. Bidirectional transmembrane modulation of integrin αIIbβ3 conformations. J. Biol. Chem. 1999, 274, 12945–12949. [Google Scholar] [CrossRef] [PubMed]
- Shattil, S.J.; Kashiwagi, H.; Pampori, N. Integrin signaling: The platelet paradigm. Blood 1998, 91, 2645–2657. [Google Scholar] [CrossRef] [PubMed]
- Arias-Salgado, E.G.; Lizano, S.; Sarkar, S.; Brugge, J.S.; Ginsberg, M.H.; Shattil, S.J. Src kinase activation by direct interaction with the integrin β cytoplasmic domain. Proc. Natl. Acad. Sci. USA 2003, 100, 13298–13302. [Google Scholar] [CrossRef]
- Obergfell, A.; Eto, K.; Mocsai, A.; Buensuceso, C.; Moores, S.L.; Brugge, J.S.; Lowell, C.A.; Shattil, S.J. Coordinate interactions of Csk, Src, and Syk kinases with αIIbβ3 initiate integrin signaling to the cytoskeleton. J. Cell Biol. 2002, 157, 265–275. [Google Scholar] [CrossRef]
- Woodside, D.G.; Obergfell, A.; Leng, L.; Wilsbacher, J.L.; Miranti, C.K.; Brugge, J.S.; Shattil, S.J.; Ginsberg, M.H. Activation of Syk protein tyrosine kinase through interaction with integrin β cytoplasmic domains. Curr. Biol. 2001, 11, 1799–1804. [Google Scholar] [CrossRef]
- Judd, B.A.; Myung, P.S.; Leng, L.; Obergfell, A.; Pear, W.S.; Shattil, S.J.; Koretzky, G.A. Hematopoietic reconstitution of SLP-76 corrects hemostasis and platelet signaling through αIIbβ3 and collagen receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 12056–12061. [Google Scholar] [CrossRef] [PubMed]
- Miranti, C.K.; Leng, L.; Maschberger, P.; Brugge, J.S.; Shattil, S.J. Identification of a novel integrin signaling pathway involving the kinase Syk and the guanine nucleotide exchange factor Vav1. Curr. Biol. 1998, 8, 1289–1299. [Google Scholar] [CrossRef]
- Goncalves, I.; Hughan, S.C.; Schoenwaelder, S.M.; Yap, C.L.; Yuan, Y.; Jackson, S.P. Integrin αIIbβ3-dependent calcium signals regulate platelet-fibrinogen interactions under flow involvement of phospholipase Cγ2. J. Biol. Chem. 2003, 278, 34812–34822. [Google Scholar] [CrossRef] [PubMed]
- Wonerow, P.; Pearce, A.C.; Vaux, D.J.; Watson, S.P. A critical role for phospholipase Cγ2 in αIIbβ3-mediated platelet spreading. J. Biol. Chem. 2003, 278, 37520–37529. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.P.; Yap, C.L.; Anderson, K.E. Phosphoinositide 3-kinases and the regulation of platelet function. Biochem. Soc. Trans. 2004, 32, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y. Protein kinase C and lipid signaling for sustained cellular responses. FASEB J. 1995, 9, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Murugappan, S.; Tuluc, F.; Dorsam, R.T.; Shankar, H.; Kunapuli, S.P. Differential role of protein kinase Cδ isoform in agonist-induced dense granule secretion in human platelets. J. Biol. Chem. 2004, 279, 2360–2367. [Google Scholar] [CrossRef] [PubMed]
- Murugappan, S.; Shankar, H.; Bhamidipati, S.; Dorsam, R.T.; Jin, J.; Kunapuli, S.P. Molecular mechanism and functional implications of thrombin-mediated tyrosine phosphorylation of PKCδ in platelets. Blood 2005, 106, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Buensuceso, C.S.; Obergfell, A.; Soriani, A.; Eto, K.; Kiosses, W.B.; Arias-Salgado, E.G.; Kawakami, T.; Shattil, S.J. Regulation of outside-in signaling in platelets by integrin-associated protein kinase C beta. J. Biol. Chem. 2005, 280, 644–653. [Google Scholar] [CrossRef]
- Chari, R.; Getz, T.; Nagy Jr, B.; Bhavaraju, K.; Mao, Y.; Bynagari, Y.S.; Murugappan, S.; Nakayama, K.; Kunapuli, S.P. Protein kinase Cδ differentially regulates platelet functional responses. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 699–705. [Google Scholar] [CrossRef]
- Pula, G.; Schuh, K.; Nakayama, K.; Nakayama, K.I.; Walter, U.; Poole, A.W. PKCδ regulates collagen-induced platelet aggregation through inhibition of VASP-mediated filopodia formation. Blood 2006, 108, 4035–4044. [Google Scholar] [CrossRef]
- Crosby, D.; Poole, A.W. Physical and functional interaction between protein kinase C δ and Fyn tyrosine kinase in human platelets. J. Biol. Chem. 2003, 278, 24533–24541. [Google Scholar] [CrossRef]
- Hall, K.J.; Jones, M.L.; Poole, A.W. Coincident regulation of PKCδ in human platelets by phosphorylation of Tyr311 and Tyr565 and phospholipase C signalling. Biochem. J. 2007, 406, 501–509. [Google Scholar] [CrossRef]
- Kostyak, J.C.; Bhavanasi, D.; Liverani, E.; McKenzie, S.E.; Kunapuli, S.P. Protein kinase C δ deficiency enhances megakaryopoiesis and recovery from thrombocytopenia. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2579–2585. [Google Scholar] [CrossRef]
- Liverani, E.; Mondrinos, M.J.; Sun, S.; Kunapuli, S.P.; Kilpatrick, L.E. Role of Protein Kinase C-delta in regulating platelet activation and platelet-leukocyte interaction during sepsis. PLoS ONE 2018, 13, e0195379. [Google Scholar] [CrossRef]
- Ivaska, J.; Kermorgant, S.; Whelan, R.; Parsons, M.; Ng, T.; Parker, P. Integrin–protein kinase C relationships. Biochem. Soc. Trans. 2003, 31, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Soriani, A.; Moran, B.; de Virgilio, M.; Kawakami, T.; Altman, A.; Lowell, C.; Eto, K.; Shattil, S.J. A role for PKCtheta in outside-in alpha(IIb)beta3 signaling. J. Thromb. Haemost. 2006, 4, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Moussazadeh, M.; Haimovich, B. Protein kinase C–delta activation and tyrosine phosphorylation in platelets. FEBS Lett. 1998, 438, 225–230. [Google Scholar] [CrossRef]
- Yacoub, D.; Theoret, J.-F.; Villeneuve, L.; Abou-Saleh, H.; Mourad, W.; Allen, B.G.; Merhi, Y. Essential role of protein kinase Cδ in platelet signaling, αIIbβ3 activation, and thromboxane A2 release. J. Biol. Chem. 2006, 281, 30024–30035. [Google Scholar] [CrossRef]
- Kim, S.; Cipolla, L.; Guidetti, G.; Okigaki, M.; Jin, J.; Torti, M.; Kunapuli, S.P. Distinct role of Pyk2 in mediating thromboxane generation downstream of both G12/13 and integrin αIIbβ3 in platelets. J. Biol. Chem. 2013, 288, 18194–18203. [Google Scholar] [CrossRef]
- Steinberg, S.F. Distinctive activation mechanisms and functions for protein kinase Cδ. Biochem. J. 2004, 384, 449–459. [Google Scholar] [CrossRef]
- Wentworth, J.K.; Pula, G.; Poole, A.W. Vasodilator-stimulated phosphoprotein (VASP) is phosphorylated on Ser157 by protein kinase C-dependent and-independent mechanisms in thrombin-stimulated human platelets. Biochem. J. 2006, 393, 555–564. [Google Scholar] [CrossRef]
- Bhavanasi, D.; Badolia, R.; Manne, B.K.; Janapati, S.; Dangelmaier, C.T.; Mazharian, A.; Jin, J.; Kim, S.; Zhang, X.; Chen, X.; et al. Cross talk between serine/threonine and tyrosine kinases regulates ADP-induced thromboxane generation in platelets. Thromb. Haemost. 2015, 114, 558–568. [Google Scholar] [CrossRef]
- O’Brien, K.A.; Gartner, T.K.; Hay, N.; Du, X. ADP-stimulated activation of Akt during integrin outside-in signaling promotes platelet spreading by inhibiting glycogen synthase kinase-3β. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2232–2240. [Google Scholar] [CrossRef]
- Newman, D. The Y’s that bind: Negative regulators of Src family kinase activity in platelets. J. Thromb. Haemost. 2009, 7, 195–199. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Senis, Y.A.; Mazharian, A.; Mori, J. Src family kinases: At the forefront of platelet activation. Blood 2014, 124, 2013–2024. [Google Scholar] [CrossRef] [PubMed]
- Consonni, A.; Cipolla, L.; Guidetti, G.; Canobbio, I.; Ciraolo, E.; Hirsch, E.; Falasca, M.; Okigaki, M.; Balduini, C.; Torti, M.; et al. Role and regulation of phosphatidylinositol 3-kinase β in platelet integrin α2β1 signaling. Blood 2012, 119, 847–856. [Google Scholar] [CrossRef]
- Cipolla, L.; Consonni, A.; Guidetti, G.; Canobbio, I.; Okigaki, M.; Falasca, M.; Ciraolo, E.; Hirsch, E.; Balduini, C.; Torti, M. The proline-rich tyrosine kinase P yk2 regulates platelet integrin α II bβ3 outside-in signaling. J Thromb. Haemost. 2013, 11, 345–356. [Google Scholar] [CrossRef]
- Canobbio, I.; Cipolla, L.; Consonni, A.; Momi, S.; Guidetti, G.; Oliviero, B.; Falasca, M.; Okigaki, M.; Balduini, C.; Gresele, P. Impaired thrombin-induced platelet activation and thrombus formation in mice lacking the Ca2+-dependent tyrosine kinase Pyk2. Blood 2013, 121, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Shattil, S.J.; Newman, P.J. Integrins: Dynamic scaffolds for adhesion and signaling in platelets. Blood 2004, 104, 1606–1615. [Google Scholar] [CrossRef]
- Kim, S.; Jin, J.; Kunapuli, S.P. Relative contribution of G-protein-coupled pathways to protease-activated receptor-mediated Akt phosphorylation in platelets. Blood 2006, 107, 947–954. [Google Scholar] [CrossRef]
- Kim, S.; Dangelmaier, C.; Bhavanasi, D.; Meng, S.; Wang, H.; Goldfinger, L.E.; Kunapuli, S.P. RhoG protein regulates glycoprotein VI-Fc receptor γ-chain complex-mediated platelet activation and thrombus formation. J. Biol. Chem. 2013, 288, 34230–34238. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhary, P.K.; Kim, S.; Jee, Y.; Lee, S.-H.; Kim, S. Characterization of Integrin αIIbβ3-Mediated Outside-in Signaling by Protein Kinase Cδ in Platelets. Int. J. Mol. Sci. 2020, 21, 6563. https://doi.org/10.3390/ijms21186563
Chaudhary PK, Kim S, Jee Y, Lee S-H, Kim S. Characterization of Integrin αIIbβ3-Mediated Outside-in Signaling by Protein Kinase Cδ in Platelets. International Journal of Molecular Sciences. 2020; 21(18):6563. https://doi.org/10.3390/ijms21186563
Chicago/Turabian StyleChaudhary, Preeti Kumari, Sanggu Kim, Youngheun Jee, Seung-Hun Lee, and Soochong Kim. 2020. "Characterization of Integrin αIIbβ3-Mediated Outside-in Signaling by Protein Kinase Cδ in Platelets" International Journal of Molecular Sciences 21, no. 18: 6563. https://doi.org/10.3390/ijms21186563
APA StyleChaudhary, P. K., Kim, S., Jee, Y., Lee, S.-H., & Kim, S. (2020). Characterization of Integrin αIIbβ3-Mediated Outside-in Signaling by Protein Kinase Cδ in Platelets. International Journal of Molecular Sciences, 21(18), 6563. https://doi.org/10.3390/ijms21186563