Total Recall: Lateral Habenula and Psychedelics in the Study of Depression and Comorbid Brain Disorders



{kind=link}
Abstract
1. Introduction
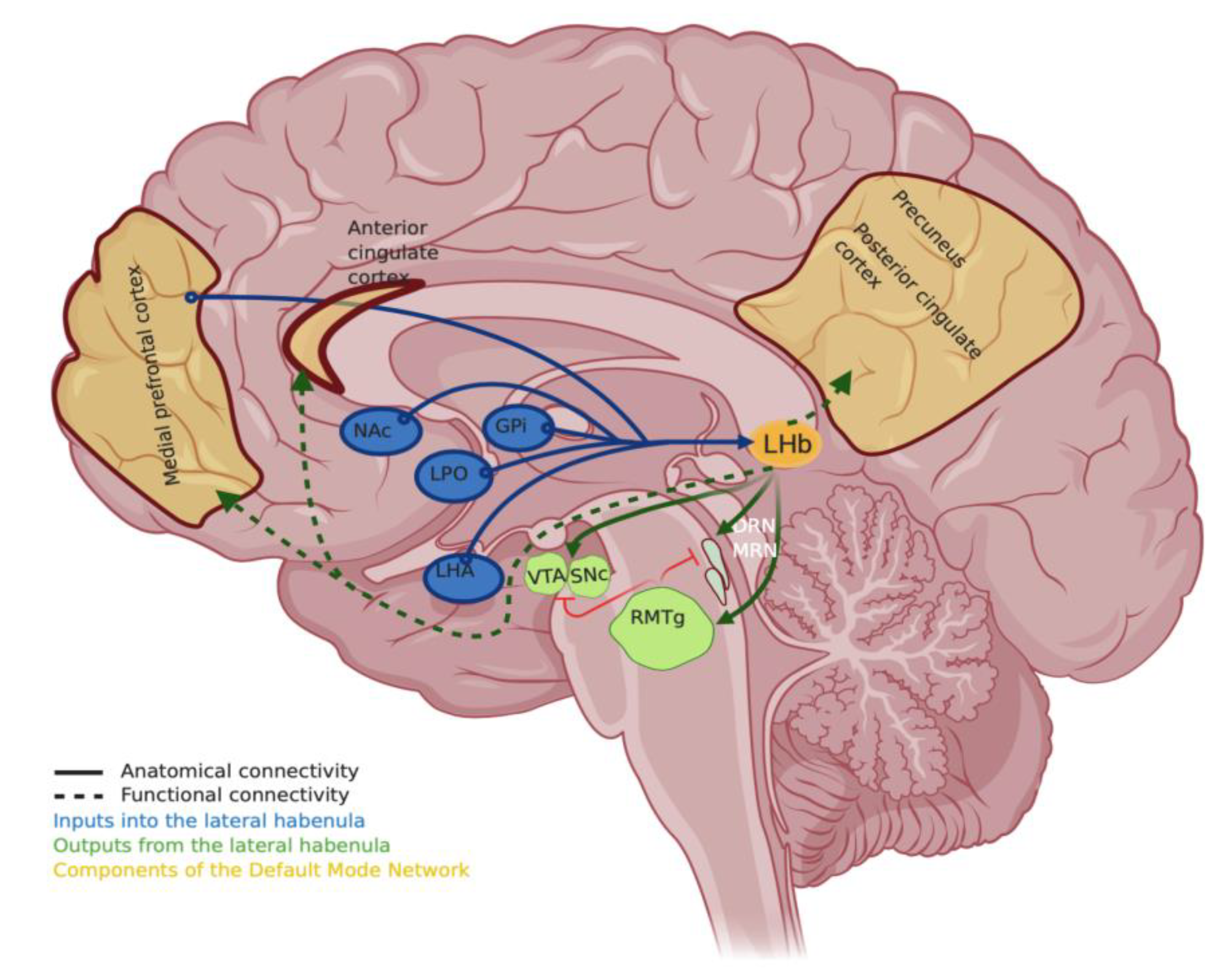
2. Lateral Habenula, “Negative-Reward”, and the Relation to Depression
3. Targeting the Hyperactivity of Lateral Habenula Provides Antidepressant Effects in Animals
4. Lateral Habenula Activity in Human Mdd Patients
5. Perils of Reliance on Single Models to Understand the Neurobiology of Habenula Function in Depression
6. Comorbidity of Depression with Neuropsychiatric Disorders: An Underexplored Avenue of Research
7. Psychedelics—A “New” Approach to Antidepressant Treatment
8. Molecular Mechanisms of Psychedelic Action and the Future of Antidepressant Discovery
9. Conclusions
Funding
Conflicts of Interest
References
- Liu, Q.; He, H.; Yang, J.; Feng, X.; Zhao, F.; Lyu, J. Changes in the global burden of depression from 1990 to 2017: Findings from the Global Burden of Disease study. J. Psychiatr. Res. 2020, 126, 134–140. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, M.; Olsson, C.A.; Jorm, A.F.; Romaniuk, H.; Patton, G.C. Association of adolescent symptoms of depression and anxiety with daily smoking and nicotine dependence in young adulthood: Findings from a 10-year longitudinal study. Addiction 2010, 105, 1652–1659. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A.R.; Hogarth, L.; Leventhal, A.M.; Cook, J.W.; Hitsman, B. Cigarette smoking and depression comorbidity: Systematic review and proposed theoretical model. Addiction 2017, 112, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Vanyukov, M.M.; Tarter, R.E.; Kirisci, L.; Kirillova, G.P.; Maher, B.S.; Clark, D.B. Liability to substance use disorders: 1. Common mechanisms and manifestations. Neurosci. Biobehav. Rev. 2003, 27, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Fracalanza, K.; McCabe, R.E.; Taylor, V.H.; Antony, M.M. The effect of comorbid major depressive disorder or bipolar disorder on cognitive behavioral therapy for social anxiety disorder. J. Affect. Disord. 2014, 162, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Finan, P.H.; Smith, M.T. The comorbidity of insomnia, chronic pain, and depression: Dopamine as a putative mechanism. Sleep Med. Rev. 2013, 17, 173–183. [Google Scholar] [CrossRef]
- Manber, R.; Edinger, J.D.; Gress, J.L.; San Pedro-Salcedo, M.G.; Kuo, T.F.; Kalista, T. Cognitive behavioral therapy for insomnia enhances depression outcome in patients with comorbid major depressive disorder and insomnia. Sleep 2008, 31, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Möller, H.-J. Occurrence and treatment of depressive comorbidity/cosyndromality in schizophrenic psychoses: Conceptual and treatment issues. World J. Biol. Psychiatry 2005, 6, 247–263. [Google Scholar] [CrossRef]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef]
- Lawson, R.P.; Nord, C.L.; Seymour, B.; Thomas, D.L.; Dayan, P.; Pilling, S.; Roiser, J.P. Disrupted habenula function in major depression. Mol. Psychiatry 2017, 22, 202–208. [Google Scholar] [CrossRef]
- Hu, H.; Cui, Y.; Yang, Y. Circuits and functions of the lateral habenula in health and in disease. Nat. Rev. Neurosci. 2020, 21, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Yang, Y.; Ni, Z.; Dong, Y.; Cai, G.; Foncelle, A.; Ma, S.; Sang, K.; Tang, S.; Li, Y.; et al. Astroglial Kir4.1 in the lateral habenula drives neuronal bursts in depression. Nature 2018, 554, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.E.; Chandra, J.; Sheriff, S.; Malinow, R. Opioid system is necessary but not sufficient for antidepressive actions of ketamine in rodents. Proc. Natl. Acad. Sci. USA 2020, 117, 2656–2662. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cui, Y.; Sang, K.; Dong, Y.; Ni, Z.; Ma, S.; Hu, H. Ketamine blocks bursting in the lateral habenula to rapidly relieve depression. Nature 2018, 554, 317–322. [Google Scholar] [CrossRef]
- Amat, J.; Sparks, P.D.; Matus-Amat, P.; Griggs, J.; Watkins, L.R.; Maier, S.F. The role of the habenular complex in the elevation of dorsal raphe nucleus serotonin and the changes in the behavioral responses produced by uncontrollable stress. Brain Res. 2001, 917, 118–126. [Google Scholar] [CrossRef]
- Lecourtier, L.; Kelly, P.H. A conductor hidden in the orchestra? Role of the habenular complex in monoamine transmission and cognition. Neurosci. Biobehav. Rev. 2007, 31, 658–672. [Google Scholar] [CrossRef]
- Hikosaka, O. The habenula: From stress evasion to value-based decision-making. Nat. Rev. Neurosci. 2010, 11, 503–513. [Google Scholar] [CrossRef]
- Stephenson-Jones, M.; Floros, O.; Robertson, B.; Grillner, S. Evolutionary conservation of the habenular nuclei and their circuitry controlling the dopamine and 5-hydroxytryptophan (5-HT) systems. Proc. Natl. Acad. Sci. USA 2012, 109, E164–E173. [Google Scholar] [CrossRef]
- Grillner, S.; von Twickel, A.; Robertson, B. The blueprint of the vertebrate forebrain—With special reference to the habenulae. Semin. Cell Dev. Biol. 2018, 78, 103–106. [Google Scholar] [CrossRef]
- Lee, H.W.; Yang, S.H.; Kim, J.Y.; Kim, H. The Role of the Medial Habenula Cholinergic System in Addiction and Emotion-Associated Behaviors. Front. Psychiatry 2019, 10, 100. [Google Scholar] [CrossRef]
- Mathuru, A.S. A little rein on addiction. Semin. Cell Dev. Biol. 2018, 78, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zhou, T.; Liao, L.; Yang, Z.; Wong, C.; Henn, F.; Malinow, R.; Yates, J.R., 3rd; Hu, H. βCaMKII in lateral habenula mediates core symptoms of depression. Science 2013, 341, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Shabel, S.J.; Wang, C.; Monk, B.; Aronson, S.; Malinow, R. Stress transforms lateral habenula reward responses into punishment signals. Proc. Natl. Acad. Sci. USA 2019, 116, 12488–12493. [Google Scholar] [CrossRef] [PubMed]
- Shabel, S.J.; Proulx, C.D.; Piriz, J.; Malinow, R. Mood regulation. GABA/glutamate co-release controls habenula output and is modified by antidepressant treatment. Science 2014, 345, 1494–1498. [Google Scholar] [CrossRef] [PubMed]
- Proulx, C.D.; Hikosaka, O.; Malinow, R. Reward processing by the lateral habenula in normal and depressive behaviors. Nat. Neurosci. 2014, 17, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Lipina, T.V.; Khrapova, M.V.; Serykh, A.; Dubrovina, N.I.; Petrova, E.S.; Mikhnevich, N.; Starostina, M.V.; Amstyslavskaja, T.G. The increased density of the habenular neurons, high impulsivity, aggression and resistant fear memory in Disc1-Q31 L genetic mouse model of depression. Behav. Brain Res. 2020, 392, 112693. [Google Scholar]
- Carlson, P.J.; Diazgranados, N.; Nugent, A.C.; Ibrahim, L.; Luckenbaugh, D.A.; Brutsche, N.; Herscovitch, P.; Manji, H.K.; Zarate, C.A., Jr.; Drevets, W.C. Neural correlates of rapid antidepressant response to ketamine in treatment-resistant unipolar depression: A preliminary positron emission tomography study. Biol. Psychiatry 2013, 73, 1213–1221. [Google Scholar] [CrossRef]
- Dandekar, M.P.; Fenoy, A.J.; Carvalho, A.F.; Soares, J.C.; Quevedo, J. Deep brain stimulation for treatment-resistant depression: An integrative review of preclinical and clinical findings and translational implications. Mol. Psychiatry 2018, 23, 1094–1112. [Google Scholar] [CrossRef]
- Sartorius, A.; Henn, F.A. Deep brain stimulation of the lateral habenula in treatment resistant major depression. Med. Hypotheses 2007, 69, 1305–1308. [Google Scholar] [CrossRef]
- Drobisz, D.; Damborská, A. Deep brain stimulation targets for treating depression. Behav. Brain Res. 2019, 359, 266–273. [Google Scholar] [CrossRef]
- Torres-Sanchez, S.; Perez-Caballero, L.; Berrocoso, E. Cellular and molecular mechanisms triggered by Deep Brain Stimulation in depression: A preclinical and clinical approach. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kiening, K.; Sartorius, A. A new translational target for deep brain stimulation to treat depression. EMBO Mol. Med. 2013, 5, 1151–1153. [Google Scholar] [CrossRef] [PubMed]
- Sartorius, A.; Kiening, K.L.; Kirsch, P.; von Gall, C.C.; Haberkorn, U.; Unterberg, A.W.; Henn, F.A.; Meyer-Lindenberg, A. Remission of major depression under deep brain stimulation of the lateral habenula in a therapy-refractory patient. Biol. Psychiatry 2010, 67, e9–e11. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.; Wang, Y.; Huang, M.; Lin, W.; Wang, S.; Zhang, B. Chronic deep brain stimulation of the lateral habenula nucleus in a rat model of depression. Brain Res. 2011, 1422, 32–38. [Google Scholar] [CrossRef]
- Jakobs, M.; Pitzer, C.; Sartorius, A.; Unterberg, A.; Kiening, K. Acute 5 Hz deep brain stimulation of the lateral habenula is associated with depressive-like behavior in male wild-type Wistar rats. Brain Res. 2019, 1721, 146283. [Google Scholar] [CrossRef]
- Nuno-Perez, A.; Tchenio, A.; Mameli, M.; Lecca, S. Lateral Habenula Gone Awry in Depression: Bridging Cellular Adaptations with Therapeutics. Front. Neurosci. 2018, 12, 485. [Google Scholar] [CrossRef]
- Belouin, S.J.; Henningfield, J.E. Psychedelics: Where we are now, why we got here, what we must do. Neuropharmacology 2018, 142, 7–19. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L.; Roseman, L.; Bolstridge, M.; Demetriou, L.; Pannekoek, J.N.; Wall, M.B.; Tanner, M.; Kaelen, M.; McGonigle, J.; Murphy, K.; et al. Psilocybin for treatment-resistant depression: fMRI-measured brain mechanisms. Sci. Rep. 2017, 7, 13187. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L.; Bolstridge, M.; Rucker, J.; Day, C.M.J.; Erritzoe, D.; Kaelen, M.; Bloomfield, M.; Rickard, J.A.; Forbes, B.; Feilding, A.; et al. Psilocybin with psychological support for treatment-resistant depression: An open-label feasibility study. Lancet Psychiatry 2016, 3, 619–627. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L.; Bolstridge, M.; Day, C.M.J.; Rucker, J.; Watts, R.; Erritzoe, D.E.; Kaelen, M.; Giribaldi, B.; Bloomfield, M.; Pilling, S.; et al. Psilocybin with psychological support for treatment-resistant depression: Six-month follow-up. Psychophamacology 2018, 235, 399–408. [Google Scholar] [CrossRef]
- Mithoefer, M.C.; Grob, C.S.; Brewerton, T.D. Novel psychopharmacological therapies for psychiatric disorders: Psilocybin and MDMA. Lancet Psychiatry 2016, 3, 481–488. [Google Scholar] [CrossRef]
- Martin, D.A.; Nichols, C.D. The Effects of Hallucinogens on Gene Expression. Curr. Top. Behav. Neurosci. 2018, 36, 137–158. [Google Scholar] [PubMed]
- Bogenschutz, M.P.; Forcehimes, A.A.; Pommy, J.A.; Wilcox, C.E.; Barbosa, P.C.R.; Strassman, R.J. Psilocybin-assisted treatment for alcohol dependence: A proof-of-concept study. J. Psychopharmacol. 2015, 29, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.W.; Garcia-Romeu, A.; Cosimano, M.P.; Griffiths, R.R. Pilot study of the 5-HT2AR agonist psilocybin in the treatment of tobacco addiction. J. Psychopharmacol. 2014, 28, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Hikosaka, O. Lateral habenula as a source of negative reward signals in dopamine neurons. Nature 2007, 447, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Dopamine in motivational control: Rewarding, aversive, and alerting. Neuron 2010, 68, 815–834. [Google Scholar] [CrossRef] [PubMed]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Distinct tonic and phasic anticipatory activity in lateral habenula and dopamine neurons. Neuron 2010, 67, 144–155. [Google Scholar] [CrossRef]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hong, S.; Hikosaka, O. A pallidus-habenula-dopamine pathway signals inferred stimulus values. J. Neurophysiol. 2010, 104, 1068–1076. [Google Scholar] [CrossRef]
- Matsumoto, M.; Hikosaka, O. Two types of dopamine neuron distinctly convey positive and negative motivational signals. Nature 2009, 459, 837–841. [Google Scholar] [CrossRef]
- Matsumoto, M.; Hikosaka, O. Representation of negative motivational value in the primate lateral habenula. Nat. Neurosci. 2009, 12, 77–84. [Google Scholar] [CrossRef]
- Herkenham, M.; Nauta, W.J. Efferent connections of the habenular nuclei in the rat. J. Comp. Neurol. 1979, 187, 19–47. [Google Scholar] [CrossRef] [PubMed]
- Christoph, G.R.; Leonzio, R.J.; Wilcox, K.S. Stimulation of the lateral habenula inhibits dopamine-containing neurons in the substantia nigra and ventral tegmental area of the rat. J. Neurosci. 1986, 6, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Agetsuma, M.; Aizawa, H.; Aoki, T.; Nakayama, R.; Takahoko, M.; Goto, M.; Sassa, T.; Amo, R.; Shiraki, T.; Kawakami, K.; et al. The habenula is crucial for experience-dependent modification of fear responses in zebrafish. Nat. Neurosci. 2010, 13, 1354–1356. [Google Scholar] [CrossRef] [PubMed]
- Mathuru, A.S.; Jesuthasan, S. The medial habenula as a regulator of anxiety in adult zebrafish. Front. Neural Circuits 2013, 7, 99. [Google Scholar] [CrossRef]
- Lee, A.; Mathuru, A.S.; Teh, C.; Kibat, C.; Korzh, V.; Penney, T.B.; Jesuthasan, S. The habenula prevents helpless behavior in larval zebrafish. Curr. Biol. 2010, 20, 2211–2216. [Google Scholar] [CrossRef] [PubMed]
- Tye, K.M.; Mirzabekov, J.J.; Warden, M.R.; Ferenczi, E.A.; Tsai, H.-C.; Finkelstein, J.; Kim, S.-Y.; Adhikari, A.; Thompson, K.R.; Andalman, A.S.; et al. Dopamine neurons modulate neural encoding and expression of depression-related behaviour. Nature 2013, 493, 537–541. [Google Scholar] [CrossRef]
- Hu, H. Reward and Aversion. Annu. Rev. Neurosci. 2016, 39, 297–324. [Google Scholar] [CrossRef]
- Cerniauskas, I.; Winterer, J.; de Jong, J.W.; Lukacsovich, D.; Yang, H.; Khan, F.; Peck, J.R.; Obayashi, S.K.; Lilascharoen, V.; Lim, B.K.; et al. Chronic Stress Induces Activity, Synaptic, and Transcriptional Remodeling of the Lateral Habenula Associated with Deficits in Motivated Behaviors. Neuron 2019, 104, 899–915.e8. [Google Scholar] [CrossRef]
- Andalman, A.S.; Burns, V.M.; Lovett-Barron, M.; Broxton, M.; Poole, B.; Yang, S.J.; Grosenick, L.; Lerner, T.N.; Chen, R.; Benster, T.; et al. Neuronal Dynamics Regulating Brain and Behavioral State Transitions. Cell 2019, 177, 970–985.e20. [Google Scholar] [CrossRef]
- Tchenio, A.; Lecca, S.; Valentinova, K.; Mameli, M. Limiting habenular hyperactivity ameliorates maternal separation-driven depressive-like symptoms. Nat. Commun. 2017, 8, 1135. [Google Scholar] [CrossRef]
- Lecca, S.; Pelosi, A.; Tchenio, A.; Moutkine, I.; Lujan, R.; Hervé, D.; Mameli, M. Rescue of GABAB and GIRK function in the lateral habenula by protein phosphatase 2A inhibition ameliorates depression-like phenotypes in mice. Nat. Med. 2016, 22, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.-S.; Zhong, P.; Liu, A.; Yan, Z.; Greengard, P. Elevation of p11 in lateral habenula mediates depression-like behavior. Mol. Psychiatry 2018, 23, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Molendijk, M.L.; de Kloet, E.R. Coping with the forced swim stressor: Current state-of-the-art. Behav. Brain Res. 2019, 364, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Hu, S.; Hu, H. Lateral Habenular Burst Firing as a Target of the Rapid Antidepressant Effects of Ketamine. Trends Neurosci. 2019, 42, 179–191. [Google Scholar] [CrossRef]
- Skandalakis, G.P.; Koutsarnakis, C.; Kalyvas, A.V.; Skandalakis, P.; Johnson, E.O.; Stranjalis, G. The habenula in neurosurgery for depression: A convergence of functional neuroanatomy, psychiatry and imaging. Brain Res. 2018, 1694, 13–18. [Google Scholar] [CrossRef]
- Huang, L.; Xi, Y.; Peng, Y.; Yang, Y.; Huang, X.; Fu, Y.; Tao, Q.; Xiao, J.; Yuan, T.; An, K.; et al. A Visual Circuit Related to Habenula Underlies the Antidepressive Effects of Light Therapy. Neuron 2019, 102, 128–142.e8. [Google Scholar] [CrossRef]
- Hennigan, K.; D’Ardenne, K.; McClure, S.M. Distinct midbrain and habenula pathways are involved in processing aversive events in humans. J. Neurosci. 2015, 35, 198–208. [Google Scholar] [CrossRef]
- Lawson, R.P.; Seymour, B.; Loh, E.; Lutti, A.; Dolan, R.J.; Dayan, P.; Weiskopf, N.; Roiser, J.P. The habenula encodes negative motivational value associated with primary punishment in humans. Proc. Natl. Acad. Sci. USA 2014, 111, 11858–11863. [Google Scholar] [CrossRef]
- Salas; Salas BOLD responses to negative reward prediction errors in human habenula. Front. Hum. Neurosci. 2010, 4, 36.
- Lawson, R.P.; Drevets, W.C.; Roiser, J.P. Defining the habenula in human neuroimaging studies. Neuroimage 2013, 64, 722–727. [Google Scholar] [CrossRef]
- Kumar, P.; Goer, F.; Murray, L.; Dillon, D.G.; Beltzer, M.L.; Cohen, A.L.; Brooks, N.H.; Pizzagalli, D.A. Impaired reward prediction error encoding and striatal-midbrain connectivity in depression. Neuropsychopharmacology 2018, 43, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Stamps, M.T.; Go, S.; Mathuru, A.S. Computational geometric tools for quantitative comparison of locomotory behavior. Sci. Rep. 2019, 9, 16585. [Google Scholar] [CrossRef] [PubMed]
- Botvinik-Nezer, R.; Holzmeister, F.; Camerer, C.F.; Dreber, A.; Huber, J.; Johannesson, M.; Kirchler, M.; Iwanir, R.; Mumford, J.A.; Adcock, R.A.; et al. Variability in the analysis of a single neuroimaging dataset by many teams. Nature 2020. [Google Scholar] [CrossRef]
- Ekstrom, A. How and when the fMRI BOLD signal relates to underlying neural activity: The danger in dissociation. Brain Res. Rev. 2010, 62, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Shekhar, K.; Regev, A.; Schier, A.F. Comprehensive Identification and Spatial Mapping of Habenular Neuronal Types Using Single-Cell RNA-Seq. Curr. Biol. 2018, 28, 1052–1065.e7. [Google Scholar] [CrossRef]
- Lim, C.H.; Kaur, P.; Teo, E.; Lam, V.Y.M.; Zhu, F.; Kibat, C.; Gruber, J.; Mathuru, A.S.; Tolwinski, N.S. Application of optogenetic Amyloid-β distinguishes between metabolic and physical damages in neurodegeneration. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Mathuru, A.S.; Libersat, F.; Vyas, A.; Teseo, S. Why behavioral neuroscience still needs diversity? A curious case of a persistent need. Neurosci. Biobehav. Rev. 2020, 116, 130–141. [Google Scholar] [CrossRef]
- Blanchard, R.J.; Hebert, M.A.; Ferrari, P.F.; Palanza, P.; Figueira, R.; Blanchard, D.C.; Parmigiani, S. Defensive behaviors in wild and laboratory (Swiss) mice: The mouse defense test battery. Physiol. Behav. 1998, 65, 201–209. [Google Scholar] [CrossRef]
- Sun, P.; Smith, A.S.; Lei, K.; Liu, Y.; Wang, Z. Breaking bonds in male prairie vole: Long-term effects on emotional and social behavior, physiology, and neurochemistry. Behav. Brain Res. 2014, 265, 22–31. [Google Scholar] [CrossRef]
- Dubnau, J.; Grady, L.; Kitamoto, T.; Tully, T. Disruption of neurotransmission in Drosophila mushroom body blocks retrieval but not acquisition of memory. Nature 2001, 411, 476–480. [Google Scholar] [CrossRef]
- Yoshihara, M.; Yoshihara, M. “Necessary and sufficient” in biology is not necessarily necessary—Confusions and erroneous conclusions resulting from misapplied logic in the field of biology, especially neuroscience. J. Neurogenet. 2018, 32, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, M.; Cheok, C.; Lee, I.-M.; Pek, E.; Verma, S.; Wong, J.; Chong, S.A. Nicotine dependence and psychiatric disorders among young males in Singapore. Nicotine Tob. Res. 2009, 11, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Cohn, A.M. Never, non-daily, and daily smoking status and progression to daily cigarette smoking as correlates of major depressive episode in a national sample of youth: Results from the National Survey of Drug Use and Health 2013 to 2015. Addict. Behav. 2018, 84, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Zbozinek, T.D.; Rose, R.D.; Wolitzky-Taylor, K.B.; Sherbourne, C.; Sullivan, G.; Stein, M.B.; Roy-Byrne, P.P.; Craske, M.G. Diagnostic overlap of generalized anxiety disorder and major depressive disorder in a primary care sample. Depress. Anxiety 2012, 29, 1065–1071. [Google Scholar] [CrossRef]
- Cryan, J.F.; Kaupmann, K. Don’t worry “B”happy! A role for GABAB receptors in anxiety and depression. Trends Pharmacol. Sci. 2005, 26, 36–43. [Google Scholar] [CrossRef]
- Jacobson, L.H.; Vlachou, S.; Slattery, D.A.; Li, X.; Cryan, J.F. The Gamma-Aminobutyric Acid B Receptor in Depression and Reward. Biol. Psychiatry 2018, 83, 963–976. [Google Scholar] [CrossRef]
- Varani, A.P.; Pedrón, V.T.; Aon, A.J.; Höcht, C.; Acosta, G.B.; Bettler, B.; Balerio, G.N. Nicotine-induced molecular alterations are modulated by GABAB receptor activity. Addict. Biol. 2018, 23, 230–246. [Google Scholar] [CrossRef]
- Kang, S.; Li, J.; Bekker, A.; Ye, J.-H. Rescue of glutamate transport in the lateral habenula alleviates depression- and anxiety-like behaviors in ethanol-withdrawn rats. Neuropharmacology 2018, 129, 47–56. [Google Scholar] [CrossRef]
- Centanni, S.W.; Bedse, G.; Patel, S.; Winder, D.G. Driving the Downward Spiral: Alcohol-Induced Dysregulation of Extended Amygdala Circuits and Negative Affect. Alcohol. Clin. Exp. Res. 2019, 43, 2000–2013. [Google Scholar] [CrossRef]
- Heal, D.J.; Henningfield, J.; Frenguelli, B.G.; Nutt, D.J.; Smith, S.L. Psychedelics—Re-opening the doors of perception. Neuropharmacology 2018, 142, 1–6. [Google Scholar] [CrossRef]
- Byock, I. Taking Psychedelics Seriously. J. Palliat. Med. 2018, 21, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Carhart-Harris, R.L.; Muthukumaraswamy, S.; Roseman, L.; Kaelen, M.; Droog, W.; Murphy, K.; Tagliazucchi, E.; Schenberg, E.E.; Nest, T.; Orban, C.; et al. Neural correlates of the LSD experience revealed by multimodal neuroimaging. Proc. Natl. Acad. Sci. USA 2016, 113, 4853–4858. [Google Scholar] [CrossRef] [PubMed]
- Palhano-Fontes, F.; Andrade, K.C.; Tofoli, L.F.; Santos, A.C.; Crippa, J.A.S.; Hallak, J.E.C.; Ribeiro, S.; de Araujo, D.B. The psychedelic state induced by ayahuasca modulates the activity and connectivity of the default mode network. PLoS ONE 2015, 10, e0118143. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L. How do psychedelics work? Curr. Opin. Psychiatry 2019, 32, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, E.; Roseman, L.; Kaelen, M.; Orban, C.; Muthukumaraswamy, S.D.; Murphy, K.; Laufs, H.; Leech, R.; McGonigle, J.; Crossley, N.; et al. Increased Global Functional Connectivity Correlates with LSD-Induced Ego Dissolution. Curr. Biol. 2016, 26, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Petri, G.; Expert, P.; Turkheimer, F.; Carhart-Harris, R.; Nutt, D.; Hellyer, P.J.; Vaccarino, F. Homological scaffolds of brain functional networks. J. R. Soc. Interface 2014, 11, 20140873. [Google Scholar] [CrossRef] [PubMed]
- von Hohenberg, C.C.; von Hohenberg, C.C.; Weber-Fahr, W.; Lebhardt, P.; Ravi, N.; Braun, U.; Gass, N.; Becker, R.; Sack, M.; Linan, A.C.; et al. Lateral habenula perturbation reduces default-mode network connectivity in a rat model of depression. Transl. Psychiatry 2018, 8, 1–9. [Google Scholar]
- Torrisi, S.; Nord, C.L.; Balderston, N.L.; Roiser, J.P.; Grillon, C.; Ernst, M. Resting state connectivity of the human habenula at ultra-high field. Neuroimage 2017, 147, 872–879. [Google Scholar] [CrossRef]
- Ely, B.A.; Xu, J.; Goodman, W.K.; Lapidus, K.A.; Gabbay, V.; Stern, E.R. Resting-state functional connectivity of the human habenula in healthy individuals: Associations with subclinical depression. Hum. Brain Mapp. 2016, 37, 2369–2384. [Google Scholar] [CrossRef]
- van der Meer, J.; van der Meer, J.; Breakspear, M. Neuroscience: Modeling the Brain on Acid. Curr. Biol. 2018, 28, R1157–R1160. [Google Scholar] [CrossRef]
- Vollenweider, F.X.; Vollenweider-Scherpenhuyzen, M.F.I.; Bäbler, A.; Vogel, H.; Hell, D. Psilocybin induces schizophrenia-like psychosis in humans via a serotonin-2 agonist action. NeuroReport 1998, 9, 3897–3902. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Cruzat, J.; Cabral, J.; Knudsen, G.M.; Carhart-Harris, R.L.; Whybrow, P.C.; Logothetis, N.K.; Kringelbach, M.L. Whole-Brain Multimodal Neuroimaging Model Using Serotonin Receptor Maps Explains Non-linear Functional Effects of LSD. Curr. Biol. 2018, 28, 3065–3074.e6. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, C.A.; Gao, R.; Negraes, P.D.; Gu, J.; Buchanan, J.; Preissl, S.; Wang, A.; Wu, W.; Haddad, G.G.; Chaim, I.A.; et al. Complex Oscillatory Waves Emerging from Cortical Organoids Model Early Human Brain Network Development. Cell Stem Cell 2019, 25, 558–569.e7. [Google Scholar] [CrossRef] [PubMed]
- Knoblich, J.A. Lab-Built Brains. Sci. Am. 2016, 316, 26–31. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef]
- Dang, J.; Tiwari, S.K.; Agrawal, K.; Hui, H.; Qin, Y.; Rana, T.M. Glial cell diversity and methamphetamine-induced neuroinflammation in human cerebral organoids. Mol. Psychiatry 2020. [Google Scholar] [CrossRef]
- Ormel, P.R.; Vieira de Sá, R.; van Bodegraven, E.J.; Karst, H.; Harschnitz, O.; Sneeboer, M.A.M.; Johansen, L.E.; van Dijk, R.E.; Scheefhals, N.; Berdenis van Berlekom, A.; et al. Microglia innately develop within cerebral organoids. Nat. Commun. 2018, 9, 4167. [Google Scholar] [CrossRef]
- Quadrato, G.; Nguyen, T.; Macosko, E.Z.; Sherwood, J.L.; Min Yang, S.; Berger, D.R.; Maria, N.; Scholvin, J.; Goldman, M.; Kinney, J.P.; et al. Cell diversity and network dynamics in photosensitive human brain organoids. Nature 2017, 545, 48–53. [Google Scholar] [CrossRef]
- Nutt, D.J.; King, L.A.; Nichols, D.E. Effects of Schedule I drug laws on neuroscience research and treatment innovation. Nat. Rev. Neurosci. 2013, 14, 577–585. [Google Scholar] [CrossRef]
- Hanks, J.B.; González-Maeso, J. Animal Models of Serotonergic Psychedelics. ACS Chem. Neurosci. 2013, 4, 33–42. [Google Scholar] [CrossRef]
- Dakic, V.; Minardi Nascimento, J.; Costa Sartore, R.; Maciel, R.d.M.; de Araujo, D.B.; Ribeiro, S.; Martins-de-Souza, D.; Rehen, S.K. Short term changes in the proteome of human cerebral organoids induced by 5-MeO-DMT. Sci. Rep. 2017, 7, 12863. [Google Scholar] [CrossRef] [PubMed]
- Osório, F.d.L.; Sanches, R.F.; Macedo, L.R.; dos Santos, R.G.; Maia-de-Oliveira, J.P.; Wichert-Ana, L.; de Araujo, D.B.; Riba, J.; Crippa, J.A.; Hallak, J.E. Antidepressant effects of a single dose of ayahuasca in patients with recurrent depression: A preliminary report. Braz. J. Psychiatry 2015, 37, 13–20. [Google Scholar]
- Quadrato, G.; Brown, J.; Arlotta, P. The promises and challenges of human brain organoids as models of neuropsychiatric disease. Nat. Med. 2016, 22, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Leuchter, A.F.; Cook, I.A.; Marangell, L.B.; Gilmer, W.S.; Burgoyne, K.S.; Howland, R.H.; Trivedi, M.H.; Zisook, S.; Jain, R.; McCracken, J.T.; et al. Comparative effectiveness of biomarkers and clinical indicators for predicting outcomes of SSRI treatment in Major Depressive Disorder: Results of the BRITE-MD study. Psychiatry Res. 2009, 169, 124–131. [Google Scholar] [CrossRef]
- Kessler, R.C.; Akiskal, H.S.; Ames, M.; Birnbaum, H.; Greenberg, P.; Robert, R.M.; Jin, R.; Merikangas, K.R.; Simon, G.E.; Wang, P.S. Prevalence and effects of mood disorders on work performance in a nationally representative sample of US workers. Am. J. Psychiatry 2006, 163, 1561–1568. [Google Scholar] [CrossRef]
- McGrath, C.L.; Kelley, M.E.; Holtzheimer, P.E.; Dunlop, B.W.; Craighead, W.E.; Franco, A.R.; Craddock, R.C.; Mayberg, H.S. Toward a neuroimaging treatment selection biomarker for major depressive disorder. JAMA Psychiatry 2013, 70, 821–829. [Google Scholar] [CrossRef]
- Leuchter, A.F.; Cook, I.A.; Hamilton, S.P.; Narr, K.L.; Toga, A.; Hunter, A.M.; Faull, K.; Whitelegge, J.; Andrews, A.M.; Loo, J.; et al. Biomarkers to predict antidepressant response. Curr. Psychiatry Rep. 2010, 12, 553–562. [Google Scholar] [CrossRef]
- Fatehullah, A.; Tan, S.H.; Barker, N. Organoids as an in vitro model of human development and disease. Nat. Cell Biol. 2016, 18, 246–254. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitkauskas, M.; Mathuru, A.S. Total Recall: Lateral Habenula and Psychedelics in the Study of Depression and Comorbid Brain Disorders. Int. J. Mol. Sci. 2020, 21, 6525. https://doi.org/10.3390/ijms21186525
Vitkauskas M, Mathuru AS. Total Recall: Lateral Habenula and Psychedelics in the Study of Depression and Comorbid Brain Disorders. International Journal of Molecular Sciences. 2020; 21(18):6525. https://doi.org/10.3390/ijms21186525
Chicago/Turabian StyleVitkauskas, Matas, and Ajay S. Mathuru. 2020. "Total Recall: Lateral Habenula and Psychedelics in the Study of Depression and Comorbid Brain Disorders" International Journal of Molecular Sciences 21, no. 18: 6525. https://doi.org/10.3390/ijms21186525
APA StyleVitkauskas, M., & Mathuru, A. S. (2020). Total Recall: Lateral Habenula and Psychedelics in the Study of Depression and Comorbid Brain Disorders. International Journal of Molecular Sciences, 21(18), 6525. https://doi.org/10.3390/ijms21186525