A Mechanosensitive Channel, Mouse Transmembrane Channel-Like Protein 1 (mTMC1) Is Translated from a Splice Variant mTmc1ex1 but Not from the Other Variant mTmc1ex2


Abstract
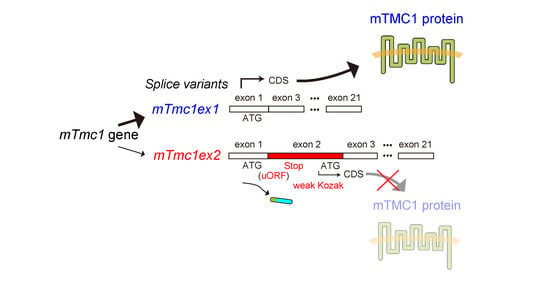
1. Introduction
2. Results
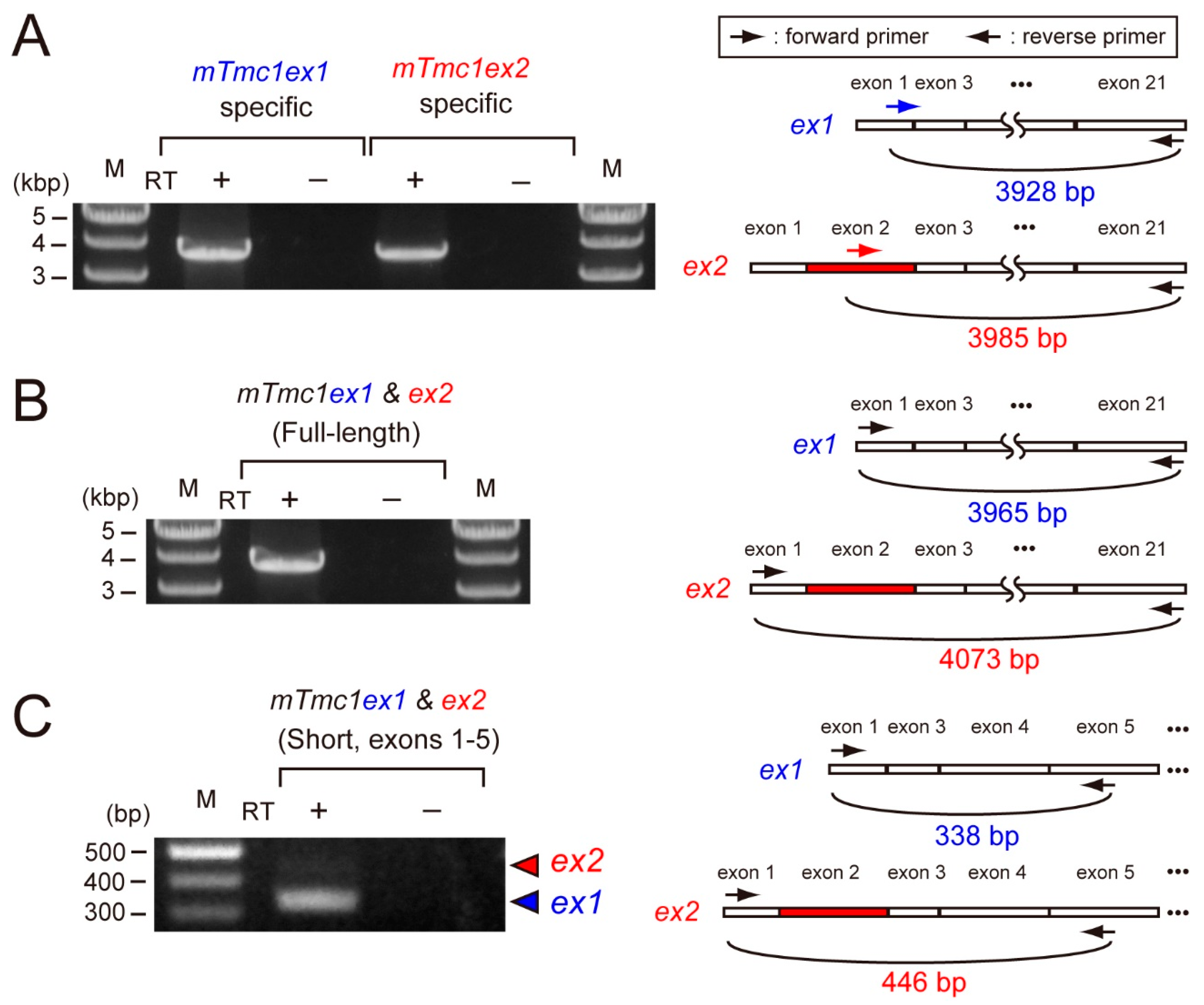
2.1. Expression of mTmc1ex1 and mTmc1ex2 in the Cochlea of 5-Week-Old Mice
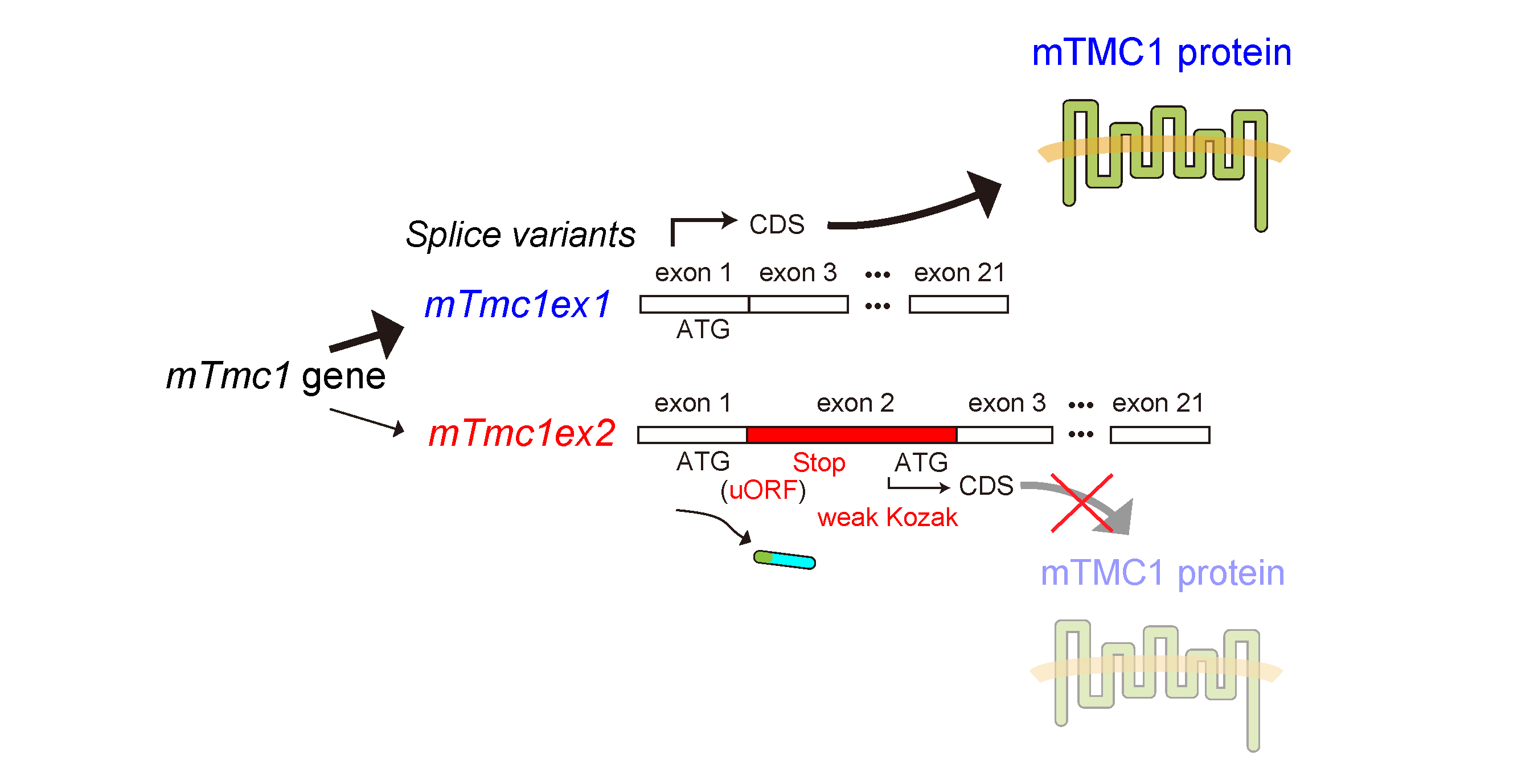
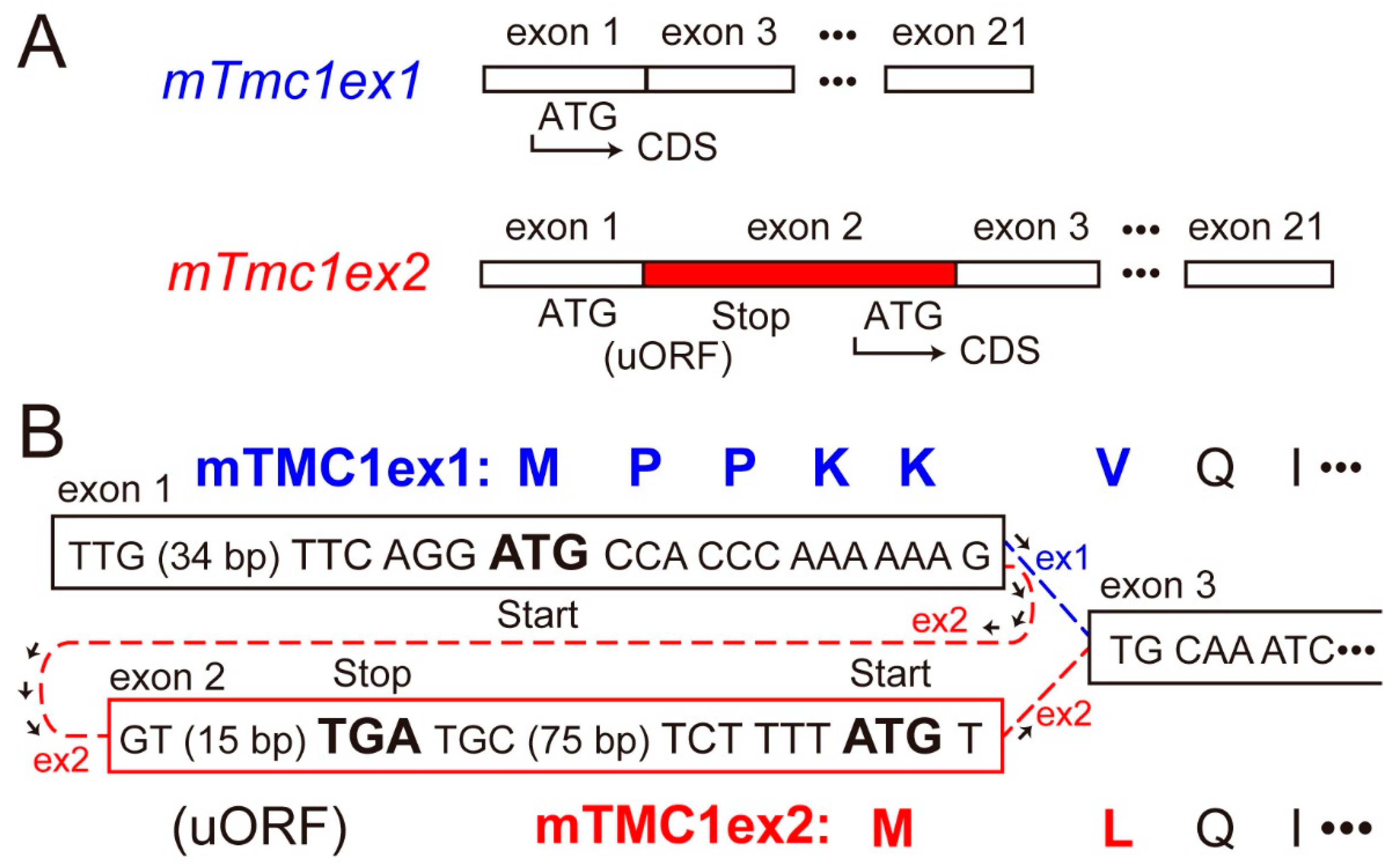
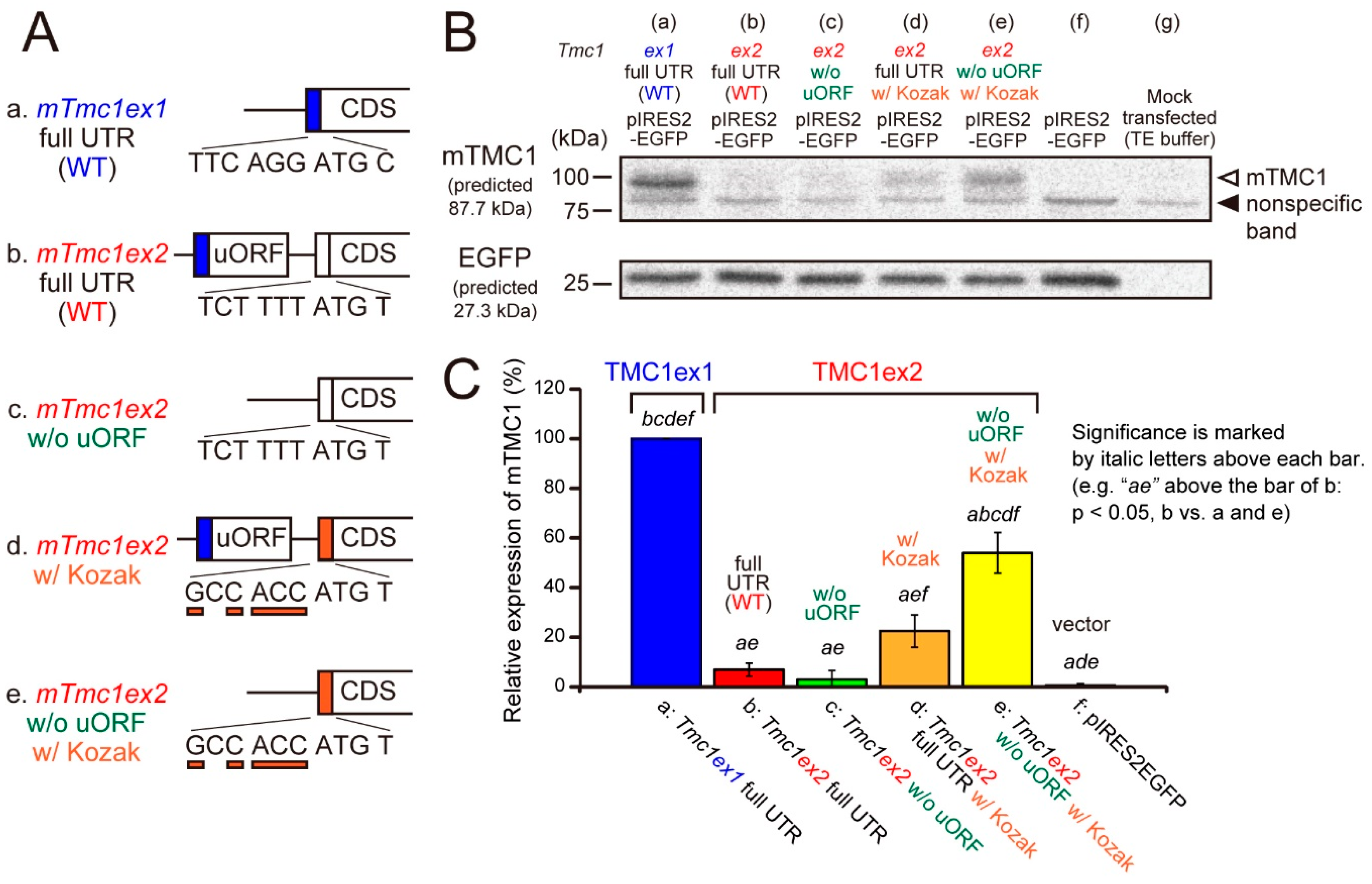
2.2. mTMC1 Is Translated from mTmc1ex1 but Not from mTmc1ex2 due to the Presence of uORF and the Lack of Kozak Sequence
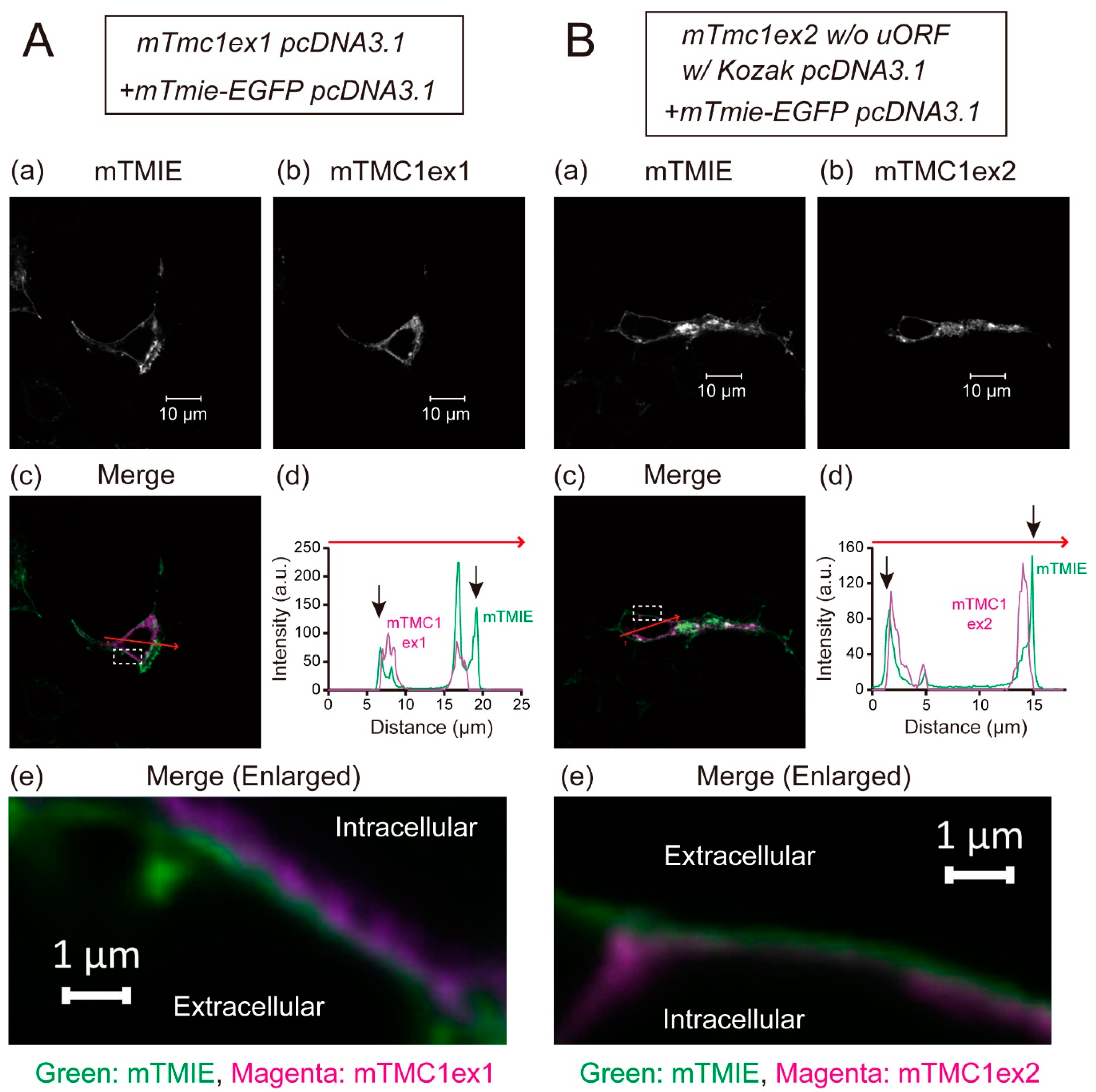
2.3. Both the Heterologously Expressed mTMC1ex1 and mTMC1ex2 Were Retained in Cytoplasm
3. Discussion
4. Materials and Methods
4.1. Animal Ethics Approval
4.2. RT-PCR
4.3. Molecular Cloning and Site-Directed Mutagenesis
4.4. Cell Culture and Transfection
4.5. Western Blotting
4.6. Immunostaining
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TMC1 | Transmembrane channel-like protein 1 |
| Tmc1 | Gene, mRNA, or cDNA for TMC1 |
| mTMC1 | Mouse TMC1 |
| MET | Mechanoelectrical transduction |
| uORF | Upstream open reading frame |
| cDNA | Complementary DNA |
| CDS | Coding sequence |
| EGFP | Enhanced green fluorescent protein |
| RT | Reverse transcription |
| M | Marker |
| UTR | Untranslated region |
| WT | Wild type |
| hTMC1 | Human TMC1 |
References
- Corey, D.P.; Akyuz, N.; Holt, J.R. Function and Dysfunction of TMC Channels in Inner Ear Hair Cells. Cold Spring Harb. Perspect. Med. 2019, 9, a033506. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, Y.; Géléoc, G.S.G.; Kurima, K.; Labay, V.; Lelli, A.; Asai, Y.; Makishima, T.; Wu, D.K.; Santina, C.C.D.; Holt, J.R.; et al. Mechanotransduction in mouse inner ear hair cells requires transmembrane channel—Like genes. J. Clin. Investig. 2011, 121, 4796–4809. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. An analysis of 5′-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acids Res. 1987, 15, 8125–8148. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.J.; Rothnagel, J.A. Emerging evidence for functional peptides encoded by short open reading frames. Nat. Rev. Genet. 2014, 15, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Labay, V.; Weichert, R.M.; Makishima, T.; Griffith, A.J. Topology of transmembrane channel-like gene 1 protein. Biochemistry 2010, 49, 8592–8598. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wu, Z.; Grillet, N.; Yan, L.; Xiong, W.; Harkins-Perry, S.; Müller, U. TMIE is an essential component of the mechanotransduction machinery of cochlear hair cells. Neuron 2014, 84, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Géléoc, G.S.; Asai, Y.; Horwitz, G.C.; Kurima, K.; Ishikawa, K.; Kawashima, Y.; Griffith, A.J.; Holt, J.R. TMC1 and TMC2 are components of the mechanotransduction channel in hair cells of the mammalian inner ear. Neuron 2013, 79, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Keresztes, G.; Mutai, H.; Heller, S. TMC and EVER genes belong to a larger novel family, the TMC gene family encoding transmembrane proteins. BMC Genomics 2003, 4, 24. [Google Scholar] [CrossRef] [PubMed]
- Harkcom, W.T.; Papanikolaou, M.; Kanda, V.; Crump, S.M.; Abbott, G.W. KCNQ1 rescues TMC1 plasma membrane expression but not mechanosensitive channel activity. J. Cell. Physiol. 2019, 234, 13361–13369. [Google Scholar] [CrossRef] [PubMed]
- Kurima, K.; Ebrahim, S.; Pan, B.; Sedlacek, M.; Sengupta, P.; Millis, B.A.; Cui, R.; Nakanishi, H.; Fujikawa, T.; Kawashima, Y.; et al. TMC1 and TMC2 Localize at the Site of Mechanotransduction in Mammalian Inner Ear Hair Cell Stereocilia. Cell Rep. 2015, 12, 1606–1617. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Tanimoto, A.; Iwasa, S.; Otsuguro, K.I. TRPM4 and TRPM5 channels share crucial amino acid residues for Ca2+ sensitivity but not significance of Pi(4,5)p2. Int. J. Mol. Sci. 2019, 20, 2012. [Google Scholar] [CrossRef] [PubMed]
- Fotiou, E.; Martin-Almedina, S.; Simpson, M.A.; Lin, S.; Gordon, K.; Brice, G.; Atton, G.; Jeffery, I.; Rees, D.C.; Mignot, C.; et al. Novel mutations in PIEZO1 cause an autosomal recessive generalized lymphatic dysplasia with non-immune hydrops fetalis. Nat. Commun. 2015, 6, 8805. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figures | Amplicon | Direction | Name | Nucleotide Sequence (5′ to 3′) |
|---|---|---|---|---|
| Figure 2A | mTmc1ex1 | Forward | mTmc1ex1_F | TTCAGGATGCCACCCAAAAAAGTGC |
| Reverse | mTmc1 3′UTR R | GAAATCAACACGATCTTTATTTGCTGC | ||
| mTmc1ex2 | Forward | mTmc1ex2_F | GCCTGTCTTCCTCTTAGCTCCTGTC | |
| Reverse | mTmc1 3′UTR R | GAAATCAACACGATCTTTATTTGCTGC | ||
| Figure 2B | mTmc1ex1 & mTmc1ex2 | Forward | mTmc1 5′UTR F | TTGCAATTCCTGATTAGAGACATTCTG |
| Reverse | mTmc1 3′UTR R | GAAATCAACACGATCTTTATTTGCTGC | ||
| Figure 2C | mTmc1ex1 & mTmc1ex2 | Forward | mTmc1 5′UTR F | TTGCAATTCCTGATTAGAGACATTCTG |
| Reverse | mTmc1 R | GTCTATTCTCATCGAGCAGTGCTTTTAAC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaguchi, S.; Hamamura, M.; Otsuguro, K.-i. A Mechanosensitive Channel, Mouse Transmembrane Channel-Like Protein 1 (mTMC1) Is Translated from a Splice Variant mTmc1ex1 but Not from the Other Variant mTmc1ex2. Int. J. Mol. Sci. 2020, 21, 6465. https://doi.org/10.3390/ijms21186465
Yamaguchi S, Hamamura M, Otsuguro K-i. A Mechanosensitive Channel, Mouse Transmembrane Channel-Like Protein 1 (mTMC1) Is Translated from a Splice Variant mTmc1ex1 but Not from the Other Variant mTmc1ex2. International Journal of Molecular Sciences. 2020; 21(18):6465. https://doi.org/10.3390/ijms21186465
Chicago/Turabian StyleYamaguchi, Soichiro, Maho Hamamura, and Ken-ichi Otsuguro. 2020. "A Mechanosensitive Channel, Mouse Transmembrane Channel-Like Protein 1 (mTMC1) Is Translated from a Splice Variant mTmc1ex1 but Not from the Other Variant mTmc1ex2" International Journal of Molecular Sciences 21, no. 18: 6465. https://doi.org/10.3390/ijms21186465
APA StyleYamaguchi, S., Hamamura, M., & Otsuguro, K.-i. (2020). A Mechanosensitive Channel, Mouse Transmembrane Channel-Like Protein 1 (mTMC1) Is Translated from a Splice Variant mTmc1ex1 but Not from the Other Variant mTmc1ex2. International Journal of Molecular Sciences, 21(18), 6465. https://doi.org/10.3390/ijms21186465