Comparative Proteomic Analysis Identifies EphA2 as a Specific Cell Surface Marker for Wharton’s Jelly-Derived Mesenchymal Stem Cells

 ,
,  , ,
, ,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
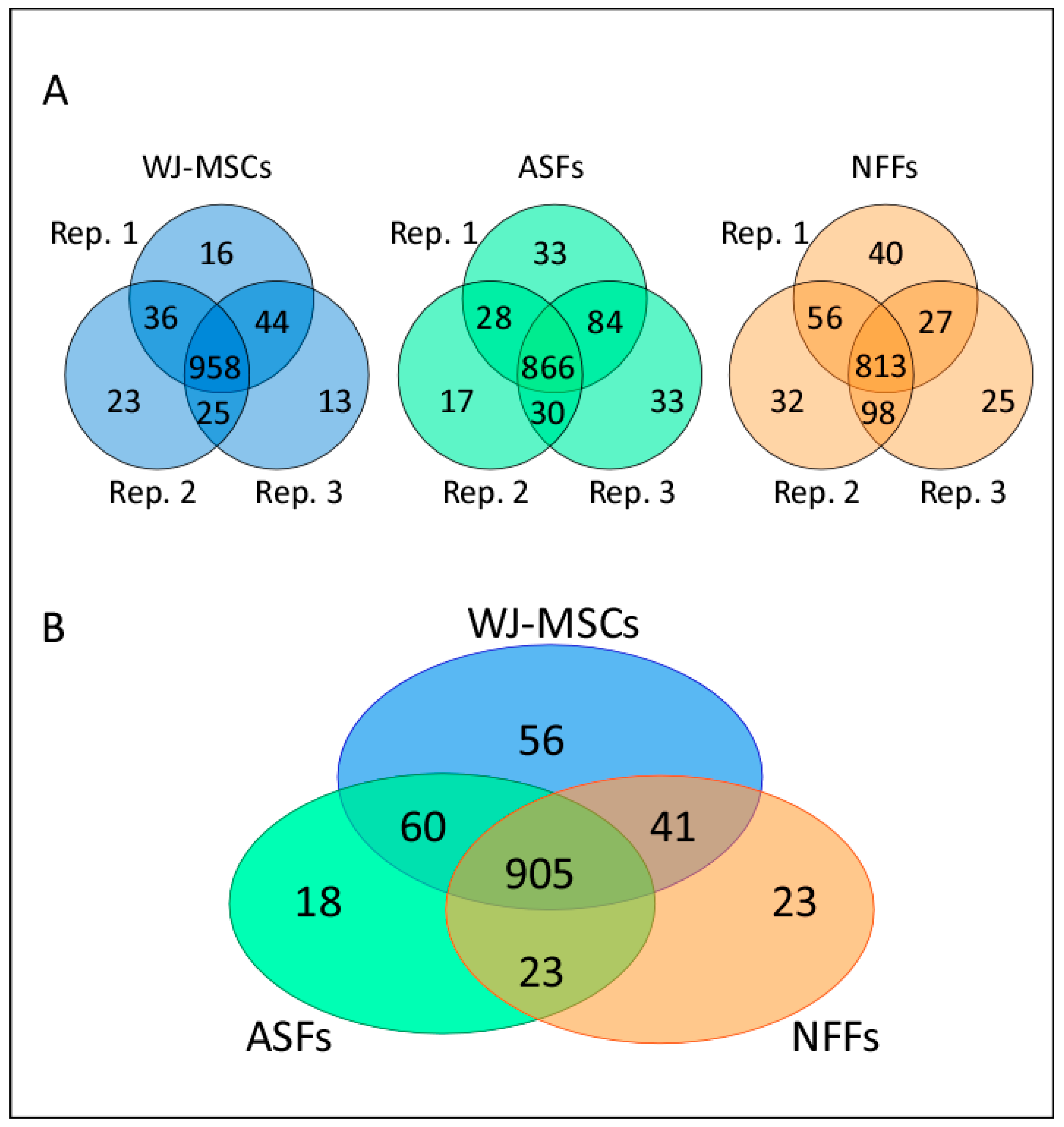
2.1. Analysis of Differentially Expressed Proteins Detected by Nano LC-MS/MS
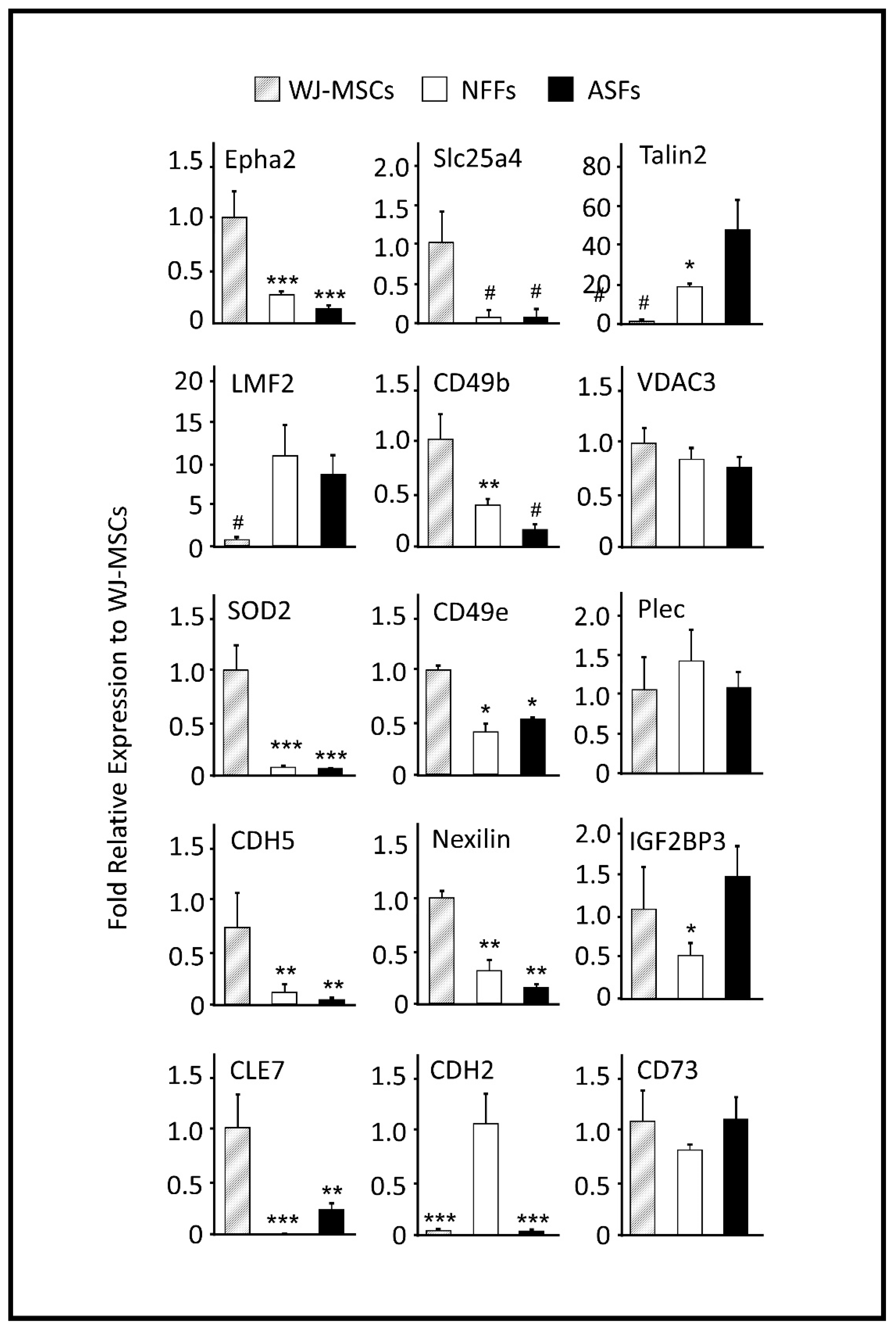
2.2. Quantitative RT-PCR Analysis of Gene Products (Proteins) Identified by Mass spectrometry (MS) Screening
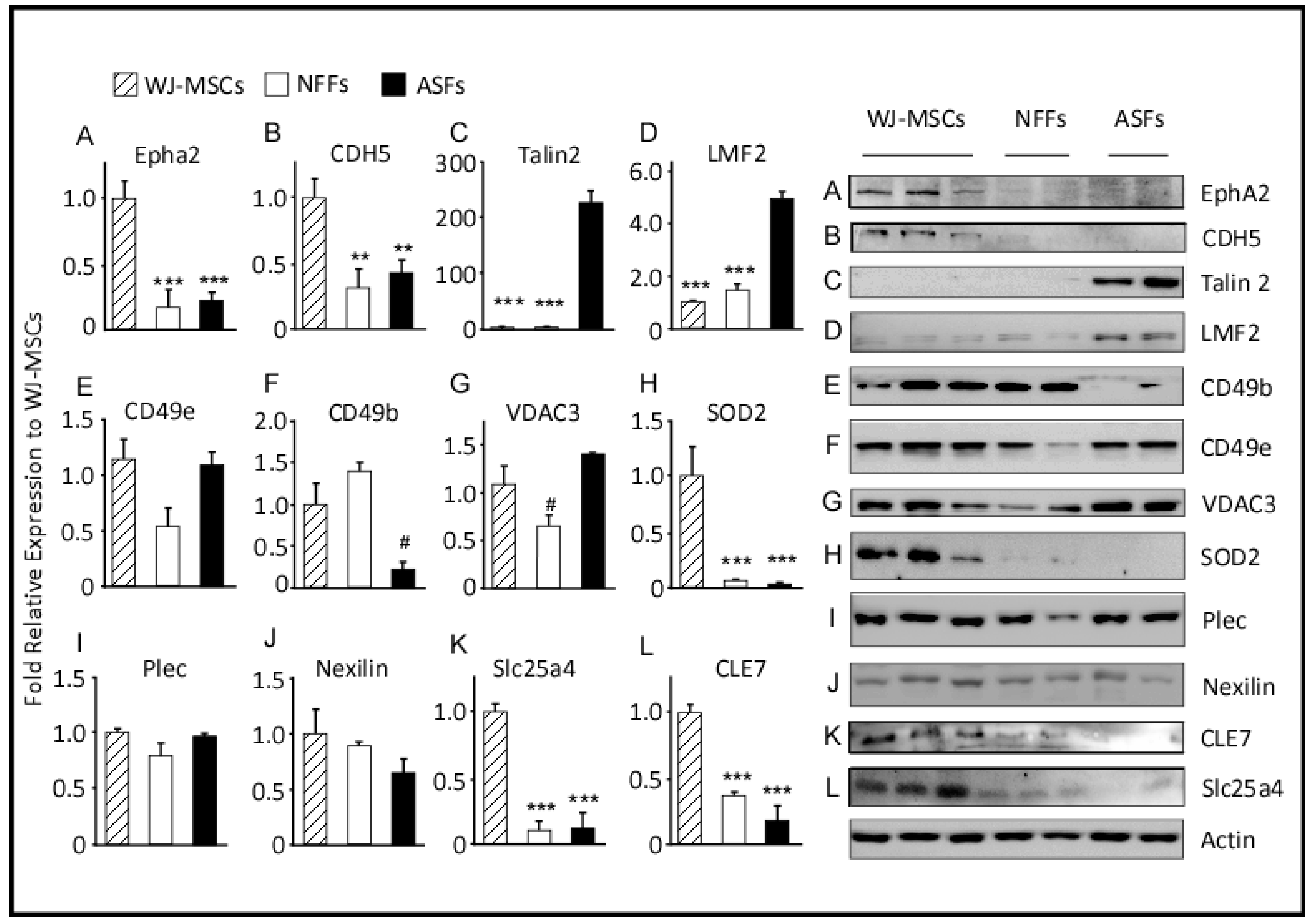
2.3. Western Blot Analysis
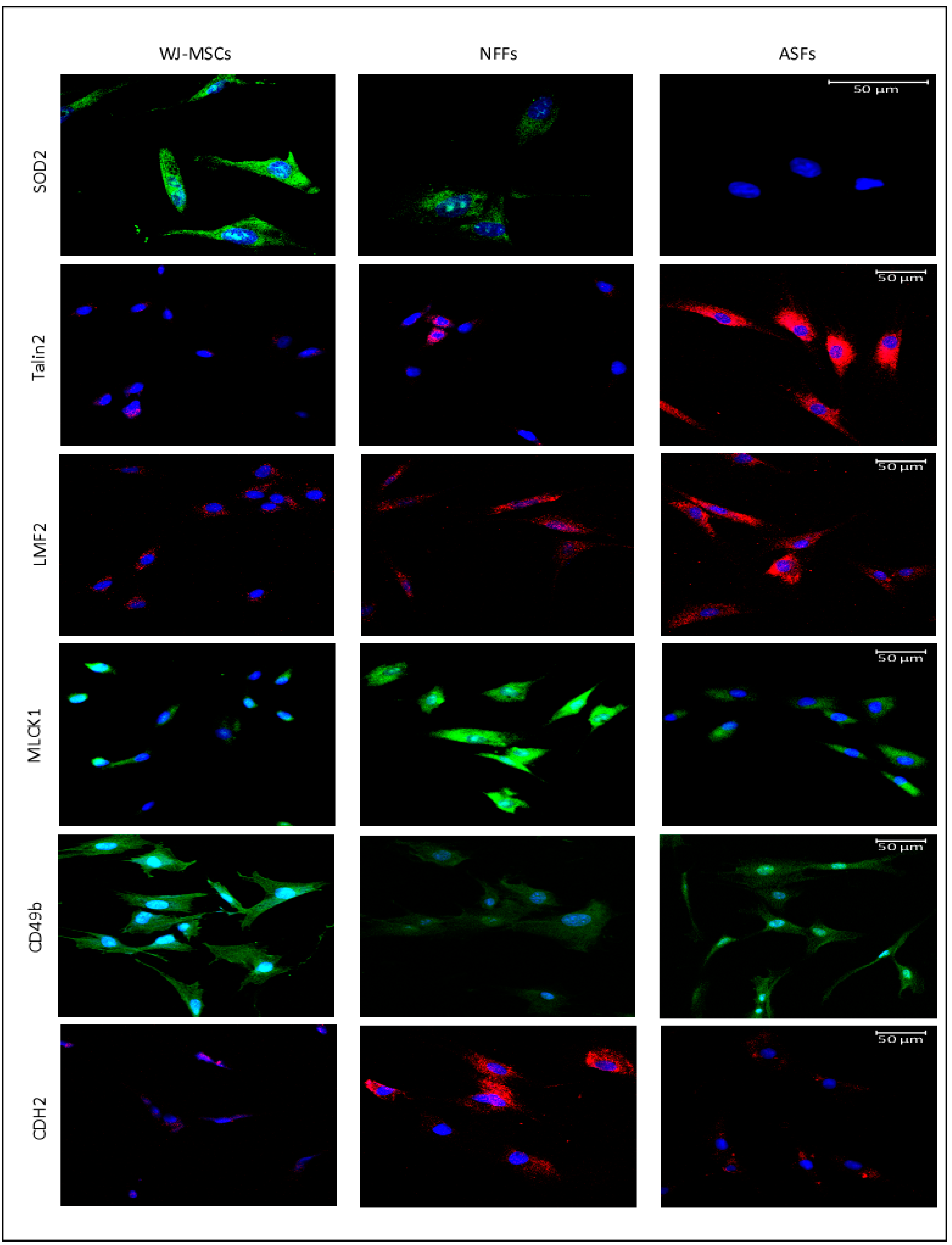
2.4. Fluorescence Microscopy
3. Discussion
4. Materials and Methods
4.1. Ethical Permission and Procurement of Human Samples
4.2. Culture and Maintenance of WJ-MSCs, NFFs, and ASFs
4.3. Preparation of Protein Extract for MS Analysis
4.4. Nano Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS) Analysis
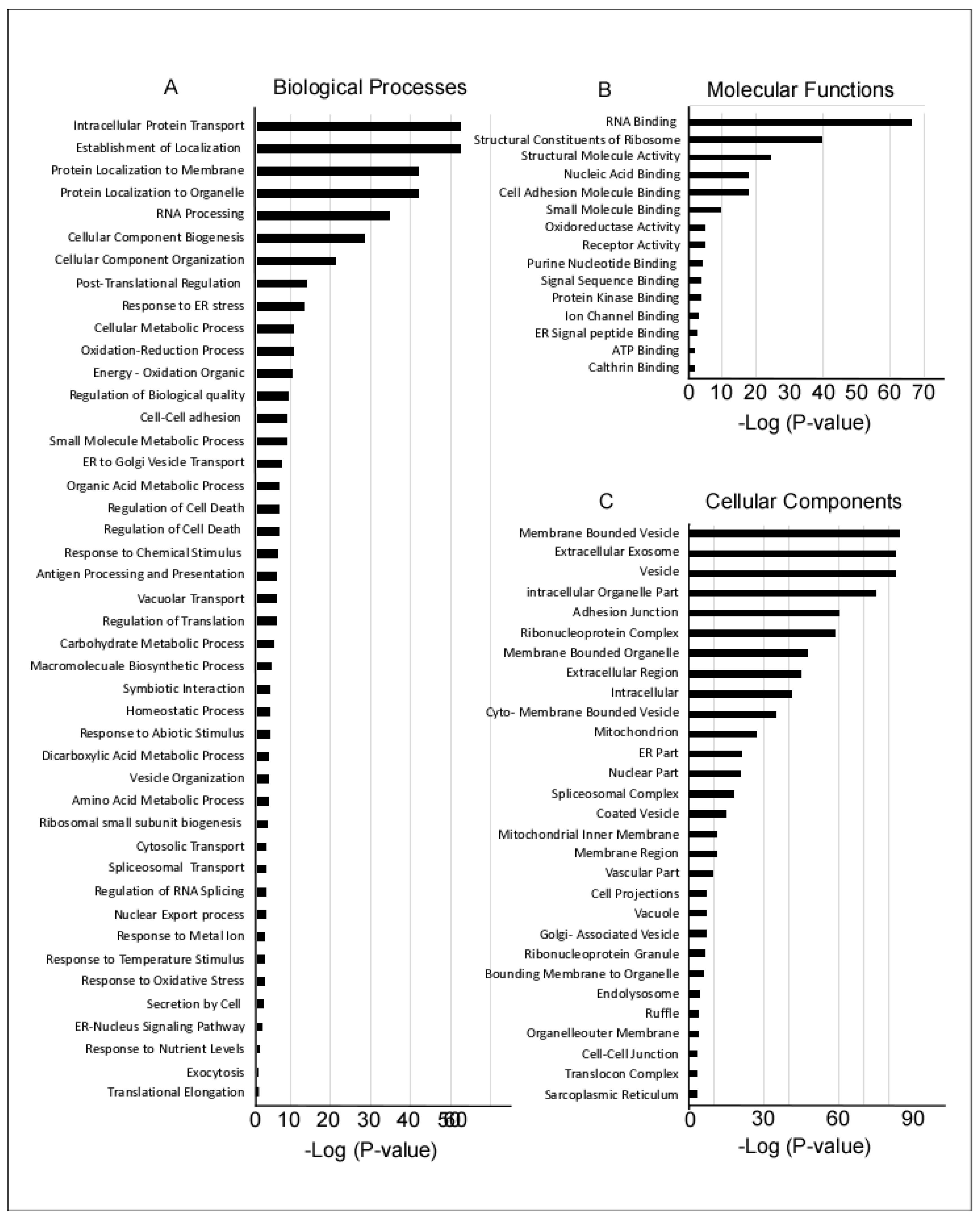
4.5. Functional Enrichment Analysis
4.6. Western Blot and Immunofluorescence Assays
4.7. RNA Extraction, cDNA Synthesis, and qRT-PCR Reactions
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASFs | Adult skin fibroblasts |
| MS | Mass spectrometry |
| MSCs | Mesenchymal stem cells |
| NFFs | Neonate foreskin fibroblasts |
| WJ-MSCs | Wharton’s jelly-derived mesenchymal stem cells |
| Nano LC-MS/MS | Nanoscale liquid chromatography coupled to tandem mass spectrometry |
| GO | Gene ontology |
| EphA2 | Ephrin type-A receptor 2 |
| TLN2 | Talin 2 |
| CDH2 | Cadherin-2 or N-cadherin |
| VDAC-3 | Voltage-dependent anion-selective channel protein-3 |
| SOD-2 | Superoxide dismutase-2 |
| ITGA-2 | Integrin alpha subunit-2 |
| LMF-2 | Lipase maturation factor-2 |
| SLC25A4 | Solute Carrier Family 25 Member 4 |
| PLEC1 | Plectin-1 |
| MLCK1 | Myosin light chain kinase-1 |
| ANTs | Adenine nucleotide translocases |
| HPA | Human protein atlas |
| qRT-PCR | Real-time quantitative reverse transcription polymerase chain reaction |
References
- Ali, H.; Al-Yatama, M.K.; Abu-Farha, M.; Behbehani, K.; Al Madhoun, A. Multi-lineage differentiation of human umbilical cord Wharton’s Jelly Mesenchymal Stromal Cells mediates changes in the expression profile of stemness markers. PLoS ONE 2015, 10, e0122465. [Google Scholar] [CrossRef] [PubMed]
- Alt, E.; Yan, Y.; Gehmert, S.; Song, Y.H.; Altman, A.; Gehmert, S.; Vykoukal, D.; Bai, X. Fibroblasts share mesenchymal phenotypes with stem cells, but lack their differentiation and colony-forming potential. Biol. Cell 2011, 103, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Gnecchi, M.; Zhang, Z.; Ni, A.; Dzau, V.J. Paracrine mechanisms in adult stem cell signaling and therapy. Circ. Res. 2008, 103, 1204–1219. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Piatetzky-Shapiro, I.I.; Petrakova, K.V. Osteogenesis in transplants of bone marrow cells. J. Embryol. Exp. Morphol. 1966, 16, 381–390. [Google Scholar] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Atari, M.; Barajas, M.; Hernandez-Alfaro, F.; Gil, C.; Fabregat, M.; Ferres Padro, E.; Giner, L.; Casals, N. Isolation of pluripotent stem cells from human third molar dental pulp. Histol. Histopathol. 2011, 26, 1057–1070. [Google Scholar] [PubMed]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, e264–e277. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Ghetu, N.; Huang, W.C.; Wang, Y.L.; Wallace, C.G.; Wen, C.J.; Chen, H.C.; Shih, L.Y.; Lin, C.F.; Hwang, S.M.; et al. Syngeneic adipose-derived stem cells with short-term immunosuppression induce vascularized composite allotransplantation tolerance in rats. Cytotherapy 2014, 16, 369–380. [Google Scholar] [CrossRef]
- Mueller, S.M.; Glowacki, J. Age-related decline in the osteogenic potential of human bone marrow cells cultured in three-dimensional collagen sponges. J. Cell Biochem. 2001, 82, 583–590. [Google Scholar] [CrossRef]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor age negatively impacts adipose tissue-derived mesenchymal stem cell expansion and differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Al-Mulla, F. Defining umbilical cord blood stem cells. Stem Cell Discov. 2012, 2, 15–23. [Google Scholar] [CrossRef]
- Joerger-Messerli, M.S.; Marx, C.; Oppliger, B.; Mueller, M.; Surbek, D.V.; Schoeberlein, A. Mesenchymal Stem Cells from Wharton’s Jelly and Amniotic Fluid. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 31, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.S.; Hung, S.C.; Peng, S.T.; Huang, C.C.; Wei, H.M.; Guo, Y.J.; Fu, Y.S.; Lai, M.C.; Chen, C.C. Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem Cells 2004, 22, 1307–1330. [Google Scholar] [CrossRef] [PubMed]
- Bagher, Z.; Azami, M.; Ebrahimi-Barough, S.; Mirzadeh, H.; Solouk, A.; Soleimani, M.; Ai, J.; Nourani, M.R.; Joghataei, M.T. Differentiation of Wharton’s Jelly-Derived Mesenchymal Stem Cells into Motor Neuron-Like Cells on Three-Dimensional Collagen-Grafted Nanofibers. Mol. Neurobiol. 2016, 53, 2397–2408. [Google Scholar] [CrossRef]
- Weiss, M.L.; Anderson, C.; Medicetty, S.; Seshareddy, K.B.; Weiss, R.J.; Vanderwerff, I.; Troyer, D.; Mcintosh, K.R. Immune Properties of Human Umbilical Cord Wharton’s Jelly-Derived Cells. Stem Cells 2008, 26, 2865–2874. [Google Scholar] [CrossRef]
- Al Madhoun, A.; Alkandari, S.; Ali, H.; Carrio, N.; Atari, M.; Bitar, M.S.; Al-Mulla, F. Chemically Defined Conditions Mediate an Efficient Induction of Mesodermal Lineage from Human Umbilical Cord- and Bone Marrow- Mesenchymal Stem Cells and Dental Pulp Pluripotent-Like Stem Cells. Cell Reprogram. 2018, 20, 9–16. [Google Scholar] [CrossRef]
- Al Madhoun, A.; Ali, H.; AlKandari, S.; Atizado, V.L.; Akhter, N.; Al-Mulla, F.; Atari, M. Defined three-dimensional culture conditions mediate efficient induction of definitive endoderm lineage from human umbilical cord Wharton’s jelly mesenchymal stem cells. Stem Cell Res. Ther. 2016, 7, 165. [Google Scholar] [CrossRef]
- Liang, J.; Wu, S.; Zhao, H.; Li, S.L.; Liu, Z.X.; Wu, J.; Zhou, L. Human umbilical cord mesenchymal stem cells derived from Wharton’s jelly differentiate into cholinergic-like neurons in vitro. Neurosci. Lett. 2013, 532, 59–63. [Google Scholar] [CrossRef]
- Bhandari, D.R.; Seo, K.W.; Sun, B.; Seo, M.S.; Kim, H.S.; Seo, Y.J.; Marcin, J.; Forraz, N.; Roy, H.L.; Larry, D.; et al. The simplest method for in vitro beta-cell production from human adult stem cells. Differentiation 2011, 82, 144–152. [Google Scholar] [CrossRef]
- Stern-Straeter, J.; Bonaterra, G.A.; Juritz, S.; Birk, R.; Goessler, U.R.; Bieback, K.; Bugert, P.; Schultz, J.; Hormann, K.; Kinscherf, R.; et al. Evaluation of the effects of different culture media on the myogenic differentiation potential of adipose tissue- or bone marrow-derived human mesenchymal stem cells. Int. J. Mol. Med. 2014, 33, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Thej, C.; Venugopal, P.; Priya, N.; Zakaria, Z.; Sundarraj, S.; Majumdar, A.S. Higher propensity of Wharton’s jelly derived mesenchymal stromal cells towards neuronal lineage in comparison to those derived from adipose and bone marrow. Cell Biol. Int. 2013, 37, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Song, T.; Wu, P.; Chen, Y.; Fan, X.; Chen, H.; Zhang, J.; Huang, C. Differentiation potential of human mesenchymal stem cells derived from adipose tissue and bone marrow to sinus node-like cells. Mol. Med. Rep. 2012, 5, 108–113. [Google Scholar]
- Meligy, F.Y.; Shigemura, K.; Behnsawy, H.M.; Fujisawa, M.; Kawabata, M.; Shirakawa, T. The efficiency of in vitro isolation and myogenic differentiation of MSCs derived from adipose connective tissue, bone marrow, and skeletal muscle tissue. Vitro Cell Dev. Biol. Anim. 2012, 48, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Gauthaman, K.; Fong, C.Y.; Suganya, C.A.; Subramanian, A.; Biswas, A.; Choolani, M.; Bongso, A. Extra-embryonic human Wharton’s jelly stem cells do not induce tumorigenesis, unlike human embryonic stem cells. Reprod. Biomed. Online 2012, 24, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Zeddou, M.; Briquet, A.; Relic, B.; Josse, C.; Malaise, M.G.; Gothot, A.; Lechanteur, C.; Beguin, Y. The umbilical cord matrix is a better source of mesenchymal stem cells (MSC) than the umbilical cord blood. Cell Biol. Int. 2010, 34, 693–701. [Google Scholar] [CrossRef]
- Fong, C.Y.; Chak, L.L.; Biswas, A.; Tan, J.H.; Gauthaman, K.; Chan, W.K.; Bongso, A. Human Wharton’s jelly stem cells have unique transcriptome profiles compared to human embryonic stem cells and other mesenchymal stem cells. Stem Cell Rev. 2011, 7, 1–16. [Google Scholar] [CrossRef]
- Hsieh, J.Y.; Fu, Y.S.; Chang, S.J.; Tsuang, Y.H.; Wang, H.W. Functional module analysis reveals differential osteogenic and stemness potentials in human mesenchymal stem cells from bone marrow and Wharton’s jelly of umbilical cord. Stem. Cells Dev. 2010, 19, 1895–1910. [Google Scholar] [CrossRef]
- Jiang, W.; Xu, J. Immune modulation by mesenchymal stem cells. Cell Prolif. 2020, 53, e12712. [Google Scholar] [CrossRef]
- Abumaree, M.; Al Jumah, M.; Pace, R.A.; Kalionis, B. Immunosuppressive properties of mesenchymal stem cells. Stem Cell Rev. Rep. 2012, 8, 375–392. [Google Scholar] [CrossRef]
- Melief, S.M.; Geutskens, S.B.; Fibbe, W.E.; Roelofs, H. Multipotent stromal cells skew monocytes towards an anti-inflammatory function: The link with key immunoregulatory molecules. Haematologica 2013, 98, e121–e122. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.M.; Capobianco, A.; Abdelrazik, H.; Becchetti, F.; Mingari, M.C.; Moretta, L. Mesenchymal stem cells inhibit natural killer-cell proliferation, cytotoxicity, and cytokine production: Role of indoleamine 2,3-dioxygenase and prostaglandin E2. Blood 2008, 111, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Obermajer, N.; Popp, F.C.; Soeder, Y.; Haarer, J.; Geissler, E.K.; Schlitt, H.J.; Dahlke, M. H Conversion of Th17 into IL-17A(neg) regulatory T cells: A novel mechanism in prolonged allograft survival promoted by mesenchymal stem cell-supported minimized immunosuppressive therapy. J. Immunol. 2014, 193, 4988–4999. [Google Scholar] [CrossRef] [PubMed]
- Yoo, K.H.; Jang, I.K.; Lee, M.W.; Kim, H.E.; Yang, M.S.; Eom, Y.; Lee, J.E.; Kim, Y.J.; Yang, S.K.; Jung, H.L.; et al. Comparison of immunomodulatory properties of mesenchymal stem cells derived from adult human tissues. Cell Immunol. 2009, 259, 150–156. [Google Scholar] [CrossRef]
- Valencia, J.; Blanco, B.; Yanez, R.; Vazquez, M.; Herrero Sanchez, C.; Fernandez-Garcia, M.; Rodriguez Serrano, C.; Pescador, D.; Blanco, J.F.; Hernando-Rodriguez, M.; et al. Comparative analysis of the immunomodulatory capacities of human bone marrow- and adipose tissue-derived mesenchymal stromal cells from the same donor. Cytotherapy 2016, 18, 1297–1311. [Google Scholar] [CrossRef]
- Melief, S.M.; Zwaginga, J.J.; Fibbe, W.E.; Roelofs, H. Adipose tissue-derived multipotent stromal cells have a higher immunomodulatory capacity than their bone marrow-derived counterparts. Stem Cells Transl. Med. 2013, 2, 455–463. [Google Scholar] [CrossRef]
- Petrenko, Y.; Vackova, I.; Kekulova, K.; Chudickova, M.; Koci, Z.; Turnovcova, K.; Kupcova Skalnikova, H.; Vodicka, P.; Kubinova, S. A Comparative Analysis of Multipotent Mesenchymal Stromal Cells derived from Different Sources, with a Focus on Neuroregenerative Potential. Sci. Rep. 2020, 10, 4290. [Google Scholar] [CrossRef]
- Ribeiro, A.; Laranjeira, P.; Mendes, S.; Velada, I.; Leite, C.; Andrade, P.; Santos, F.; Henriques, A.; Graos, M.; Cardoso, C.M.; et al. Mesenchymal stem cells from umbilical cord matrix, adipose tissue and bone marrow exhibit different capability to suppress peripheral blood B, natural killer and T cells. Stem Cell Res. 2013, 4, 125. [Google Scholar] [CrossRef]
- Kalaszczynska, I.; Ferdyn, K. Wharton’s jelly derived mesenchymal stem cells: Future of regenerative medicine? Recent findings and clinical significance. Biomed Res. Int. 2015, 2015, 430847. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Q.; Wang, Z.; Tong, H.; Ma, L.; Zhang, Y.; Shan, F.; Meng, Y.; Yuan, Z. Comparative analysis of human mesenchymal stem cells from fetal-bone marrow, adipose tissue, and Warton’s jelly as sources of cell immunomodulatory therapy. Hum. Vaccin. Immunother. 2016, 12, 85–96. [Google Scholar] [CrossRef]
- Kendall, R.T.; Feghali-Bostwick, C. A Fibroblasts in fibrosis: Novel roles and mediators. Front. Pharmacol. 2014, 5, 123. [Google Scholar] [CrossRef] [PubMed]
- Halfon, S.; Abramov, N.; Grinblat, B.; Ginis, I. Markers distinguishing mesenchymal stem cells from fibroblasts are downregulated with passaging. Stem Cells Dev. 2011, 20, 53–66. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Abreu, R.; Penalva, L.O.; Marcotte, E.M.; Vogel, C. Global signatures of protein and mRNA expression levels. Mol. Biosyst. 2009, 5, 1512–1526. [Google Scholar] [CrossRef]
- Brohem, C.A.; de Carvalho, C.M.; Radoski, C.L.; Santi, F.C.; Baptista, M.C.; Swinka, B.B.; de A. Urban, C.; de Araujo, L.R.; Graf, R.M.; Feferman, I.H.; et al. Comparison between fibroblasts and mesenchymal stem cells derived from dermal and adipose tissue. Int. J. Cosmet. Sci. 2013, 35, 448–457. [Google Scholar] [CrossRef]
- Denu, R.A.; Nemcek, S.; Bloom, D.D.; Goodrich, A.D.; Kim, J.; Mosher, D.F.; Hematti, P. Fibroblasts and Mesenchymal Stromal/Stem Cells Are Phenotypically Indistinguishable. Acta Haematol. 2016, 136, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, K.; Sicker, M.; Schmelzer, E.; Rupf, T.; Salvetter, J.; Schulz-Siegmund, M.; Bader, A. Multilineage differentiation potential of human dermal skin-derived fibroblasts. Exp. Dermatol. 2008, 17, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Sudo, K.; Kanno, M.; Miharada, K.; Ogawa, S.; Hiroyama, T.; Saijo, K.; Nakamura, Y. Mesenchymal progenitors able to differentiate into osteogenic, chondrogenic, and/or adipogenic cells in vitro are present in most primary fibroblast-like cell populations. Stem Cells 2007, 25, 1610–1617. [Google Scholar] [CrossRef]
- Alfaro, D.; Zapata, A.G. Eph/Ephrin-mediated stimulation of human bone marrow mesenchymal stromal cells correlates with changes in cell adherence and increased cell death. Stem Cell Res. 2018, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.; Campbell, J.; Nobes, C.D. Ephs and ephrins. Curr. Biol. 2017, 27, R90–R95. [Google Scholar] [CrossRef]
- Goldshmit, Y.; McLenachan, S.; Turnley, A. Roles of Eph receptors and ephrins in the normal and damaged adult CNS. Brain Res. Rev. 2006, 52, 327–345. [Google Scholar] [CrossRef]
- Binda, E.; Visioli, A.; Giani, F.; Lamorte, G.; Copetti, M.; Pitter, K.L.; Huse, J.T.; Cajola, L.; Zanetti, N.; DiMeco, F.; et al. The EphA2 receptor drives self-renewal and tumorigenicity in stem-like tumor-propagating cells from human glioblastomas. Cancer Cell 2012, 22, 765–780. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.H.; Lee, S.J.; Oh, S.Y.; Lee, H.J.; Ryu, J.M.; Han, H.J. Oleic acid enhances the motility of umbilical cord blood derived mesenchymal stem cells through EphB2-dependent F-actin formation. Biochim. Biophys. Acta 2015, 1853, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.P.; Liu, W.T.; Lin, Y.; Li, Y.T.; Chang, C.H.; Chang, F.W.; Wang, L.M.; Teng, S.W.; Hsuan, Y. EphA2 is a biomarker of hMSCs derived from human placenta and umbilical cord. Taiwan J. Obs. Gynecol. 2015, 54, 749–756. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wen, Y.C.; Du, M.K.; Li, M.W.; Hsuan, Y.C.; Su, Y.C.; Lin, W. EphA2-positive human umbilical cord-derived mesenchymal stem cells exert anti-fibrosis and immunomodulatory activities via secretion of prostaglandin E2. Taiwan J. Obs. Gynecol. 2018, 57, 722–725. [Google Scholar] [CrossRef]
- Toran, J.L.; Lopez, J.A.; Gomes-Alves, P.; Aguilar, S.; Torroja, C.; Trevisan-Herraz, M.; Moscoso, I.; Sebastiao, M.J.; Serra, M.; Brito, C.; et al. Definition of a cell surface signature for human cardiac progenitor cells after comprehensive comparative transcriptomic and proteomic characterization. Sci. Rep. 2019, 9, 4647. [Google Scholar] [CrossRef]
- Choi, M.; Lee, H.S.; Naidansaren, P.; Kim, H.K.; O, E.; Cha, J.H.; Ahn, H.Y.; Yang, P.I.; Shin, J.C.; Joe, Y.A. Proangiogenic features of Wharton’s jelly-derived mesenchymal stromal/stem cells and their ability to form functional vessels. Int. J. Biochem. Cell Biol. 2013, 45, 560–570. [Google Scholar] [CrossRef]
- Dejana, E.; Orsenigo, F.; Molendini, C.; Baluk, P.; McDonald, D.M. Organization and signaling of endothelial cell-to-cell junctions in various regions of the blood and lymphatic vascular trees. Cell Tissue. Res. 2009, 335, 17–25. [Google Scholar] [CrossRef]
- Yoo, J.K.; Kim, J.; Choi, S.J.; Noh, H.M.; Kwon, Y.D.; Yoo, H.; Yi, H.S.; Chung, H.M.; Kim, J.K. Discovery and characterization of novel microRNAs during endothelial differentiation of human embryonic stem cells. Stem Cells Dev. 2012, 21, 2049–2057. [Google Scholar] [CrossRef]
- Adorno-Cruz, V.; Liu, H. Regulation and functions of integrin alpha2 in cell adhesion and disease. Genes Dis. 2019, 6, 16–24. [Google Scholar] [CrossRef]
- Ginsberg, M.H. Integrin activation. BMB Rep. 2014, 47, 655–659. [Google Scholar] [CrossRef]
- Ye, F.; Snider, A.K.; Ginsberg, M.H.; Ye, F.; Snider, A.K.; Ginsberg, M.H. Talin and kindlin: The one-two punch in integrin activation. Front Med. 2014, 8, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Cappellesso-Fleury, S.; Puissant-Lubrano, B.; Apoil, P.A.; Titeux, M.; Winterton, P.; Casteilla, L.; Bourin, P.; Blancher, A. Human fibroblasts share immunosuppressive properties with bone marrow mesenchymal stem cells. J. Clin. Immunol. 2010, 30, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Thul, P.J.; Akesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Bjork, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356. [Google Scholar] [CrossRef] [PubMed]
- Mafi, P.; Hindocha, S.; Mafi, R.; Griffin, M.; Khan, W.S. Adult mesenchymal stem cells and cell surface characterization—A systematic review of the literature. Open Orthop. J. 2011, 5, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wang, F.; Wang, Y.; Ning, Z.; Hou, W.; Wright, T.G.; Sundaram, M.; Zhong, S.; Yao, Z.; Figeys, D. Improved recovery and identification of membrane proteins from rat hepatic cells using a centrifugal proteomic reactor. Mol. Cell Proteomics 2011, 10, O111 008425. [Google Scholar] [CrossRef]
- Abu-Farha, M.; Tiss, A.; Abubaker, J.; Khadir, A.; Al-Ghimlas, F.; Al-Khairi, I.; Baturcam, E.; Cherian, P.; Elkum, N.; Hammad, M.; et al. Proteomics analysis of human obesity reveals the epigenetic factor HDAC4 as a potential target for obesity. PLoS ONE 2013, 8, e75342. [Google Scholar] [CrossRef] [PubMed]
- Al Madhoun, A.S.; Mehta, V.; Li, G.; Figeys, D.; Wiper-Bergeron, N.; Skerjanc, I. Skeletal myosin light chain kinase regulates skeletal myogenesis by phosphorylation of MEF2C. EMBO J. 2011, 30, 2477–2489. [Google Scholar] [CrossRef]
- Voronova, A.; Al Madhoun, A.; Fischer, A.; Shelton, M.; Karamboulas, C.; Skerjanc, I.S. Gli2 and MEF2C activate each other’s expression and function synergistically during cardiomyogenesis in vitro. Nucleic Acids Res. 2012, 40, 3329–3347. [Google Scholar] [CrossRef]
- Khadir, A.; Tiss, A.; Abubaker, J.; Abu-Farha, M.; Al-Khairi, I.; Cherian, P.; John, J.; Kavalakatt, S.; Warsame, S.; Al-Madhoun, A.; et al. MAP kinase phosphatase DUSP1 is overexpressed in obese humans and modulated by physical exercise. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E71–E83. [Google Scholar] [CrossRef]
- Al-Madhoun, A.S.; van der Wilt, C.L.; Loves, W.J.; Padron, J.M.; Eriksson, S.; Talianidis, I.; Peters, G.J. Detection of an alternatively spliced form of deoxycytidine kinase mRNA in the 2′-2′-difluorodeoxycytidine (gemcitabine)-resistant human ovarian cancer cell line AG6000. Biochem. Pharmacol. 2004, 68, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Seed, B. A PCR primer bank for quantitative gene expression analysis. Nucleic Acids Res. 2003, 31, e154. [Google Scholar] [CrossRef] [PubMed]
- Al Madhoun, A.S.; Voronova, A.; Ryan, T.; Zakariyah, A.; McIntire, C.; Gibson, L.; Shelton, M.; Ruel, M.; Skerjanc, I.S. Testosterone enhances cardiomyogenesis in stem cells and recruits the androgen receptor to the MEF2C and HCN4 genes. J. Mol. Cell Cardiol. 2013, 60, 164–171. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular Localization | Unique to WJ-MSCs | Unique to NFFs | Unique to ASFs | WJ-MSCs and NFFs | WJ-MSCs and ASFs | ASFs and NFFs |
|---|---|---|---|---|---|---|
| Extracellular Exosome | ANP32B, ALDH1A3, ATP1B3, ATP2B1, DDX19B, DYNC2H1, EIF3E, GRHPR, HSPA2, ICAM1, NUCB2, PFKL, PFDN2, PDCD5, PCMT1, RAB5A, TMEM106B, TUBB3, VDAC3 | CDH2, EFHD1, RAB3B, RAB3B, SNRPE, TPP1 | AK1, ATP6V1A, GNAL, NDRG1, QDPR, SIRPA, TUBB4A, UBE2V2 | COLEC12, DSTN, DLST, ECHS1, IGF2R, IGFBP7, IDH1, IDH2, NDUFB4, PRDX3, PLD3, REEP5, SCAMP3, STX12, TMED9, TPM3, USP14 | ADH5, CAPN1, CAPN2, DNAJB4, EMILIN1, GLIPR2, GFPT1, ITSN2, LTA4H, HLA-B, HLA-C, MARS, PDLIM2, PEPD, PSMC6, PSMB6, RHOA, RHOC, RAC1, RAC2, RAC3, RNH1, RPS17, STK10, SOD2, TXNL1, TIMP3, FLJ44635, TCEB2, TPT1, UBE2L3 | ANXA4, CYB5A, GNB2, GNB4, LAMTOR1, PCK2, PTGS1, PRKCA, RPLP1, SNX18 |
| Cell–Cell Adhesion | DDX6, EPHA2, GCN1, LIMS1, BZW2, PCMT1 | CDH2, GOLGA3, STAT1, TMOD3 | TLN2 | IDH1, KTN1, MACF1 | EFHD2, SWAP70, MYH9, PSMB6, RHOA, TJP2, TWF1 | OXTR, SNX1 |
| Cellular Adherence | DDX6, ICAM1, LIMS1, TGFB1I1 | STAT1 | TNS3 | DPP4, IGF2R), ITGA2, IDH1 | CAPN2, RAC1, TLN2 | CAPN1 |
| Mitochondria | C1QBP, LETM1, PYCR1, PYCR2, SLC25A4, SQRDL, VDAC3 | EFHD1 | – | BRI3BP, NDUFB4, DLST, ECHS1, IDH2, PRDX3, PITRM1 | ALDH18A1, DLD, GLUD1, OGDH, SOD2 | CYB5A, PCK2, PRKCA |
| Mitochondrial Envelope | LETM1, SLC25A4, SQRDL, VDAC3 | EFHD1, SLC25A1 | – | NDUFB4 | ALDH18A1, OGDH, SOD2 | CYB5A, COX1, PRKCA |
| Endoplasmic Reticulum (ER) | APOL2 | MLEC, PML | – | PLD3, REEP5, TMED9 | CAPN2, HLA-B, HLA-C, RHOA, RAC1, SEC24D | CYB5A, LMF2, PTGS1 |
| ER Membrane | APOL2 | RAB2B | – | KTN1 | HLA-B, HLA-C, RAC1, SEC24D | CYB5A, PTGS1 |
| Nuclear Parts | C1QBP, DDX6, LSM2, NUP93, TGFB1I1, WDR36 | STAT1, SNRPE, SART3, TPP1 | – | ADAR, DLST, ECHS1, IGF2R, IDH1, NDUFB4, PITRM1, SPARC, TP53BP1, U2AF1 | GLUD1, OGDH, PPP3CA, SRRT, SF1, SOD2, TCEB2 | PRKCA |
| Plasma Membrane Raft | EPHA2, RAB5A | CDH2 | TLN2 | ATP1B3, MACF1, TPM1 | PRKAR1A, RAC1, TWF1 | – |
| Cytoplasm Membrane | RAB3B, TPP1 | PACS1 | ATP1B3, DPP4, IGF2R, SPARC, STX12, USP14 | HLA-B, HLA-C, RAC1, SEC24D | AP1B1 | |
| Intracellular Membranes | APOL2, C1QBP, DDX19A, DDX19B, DDX6, LETM1, LIMS1, LSM2, NUCB2, NUP93, PFKL, SLC25A4, SQRDL, TGFB1I1, VDAC3 | EFHD1, RAB2B, RAB3B, STAT1, SNRPE, SLC25A1, SART3, TPP1 | PACS1, RANBP2, RGPD3, RGPD4, RGPD5, RGPD6, RGPD8, TNS3 | ADAR, ATP1B3, DLST, DPP4, ECHS1, IGF2R, ITGA2, IDH1, KTN1, NDUFB4, PITRM1, SCRN1, SPARC, STX12, TPM2, TP53BP1, U2AF1, USP14 | ALDH18A1, CAPN1, CAPN2, GLUD1, GLUD2, HLA-B, HLA-C, OGDH, PRKAR1A, PPP3CA, RAC1, SEC24D, STRAP, SRRT, SF1, SOD2, TLN2, TCEB2, TPM3 | AP1B1, COX1, PGP, PTGS1, PRKCA, RPLP1, ADH5 |
| Spliceosome | LSM2 | SNRPE | – | ADAR | SNRPA, SF1 | – |
| Signaling Pathways | Unique to WJ-MSCs | Unique to NFFs | Unique to ASFs | WJ-MSCs and NFFs | WJ-MSCs and ASFs | ASFs and NFFs |
|---|---|---|---|---|---|---|
| Signal Transduction Pathway | ||||||
| Wnt | CTHRC1 | CDH2 | – | – | RHOA, UBA52 | – |
| NF-κB | – | – | – | – | PSMB6 | – |
| Notch | – | – | – | – | – | ANXA4 |
| Interferon | – | – | – | ADAR, STAT1 | HLA-B, HLA-C | – |
| Insulin/Glucose | NUP93, PFKL | – | ATP6V1A, RANBP2 | – | – | – |
| Amino Acid Biosynthesis | PYCR1, PYCR2 | – | – | – | ALDH18, GLUD1 | – |
| Tricarboxylic Acid Cycle | – | – | – | DLST, IDH1, IDH2, | DLD, OGDH | – |
| Intracellular Transport | DDX19A, DDX19B, NUP93, RPL9 | SNRPE | RANBP2, RGPD3, RGPD4, RGPD5, RGPD6, RGPD8 | ADAR, STX12, U2AF1 | PPP3CA, RPS17, SEC24D | AP1B1, RPLP1 |
| RNA Processing | C1QBP, LSM2, RPL9, WDR36 | SNRPE, SART3 | – | ADAR, RPS17, U2AF1 | STRAP, SRRT, SNRPA, SF1 | RPLP1 |
| Cell Component Biogenesis | DDX6, LIMS1, WDR36, C1QBP, ICAM1, NUP93, PFKL, VDAC3 | SNRPE, SART3 | – | NDUFB4, ADAR, ITGA2, STX12 | SEC24D, PRKAR1A, RAC1, STRAP, SF1, SOD2, TLN2, TCEB2 | PRKCA, RPLP1 |
| Oxidation-reduction Process | NUCB2, PFKL, SQRDL | – | – | NDUFB4, DLST, ECHS1, IDH | CYB5A, COX1, PTGS1ADH5, ALDH18A1, GLUD1, GLUD2, OGDH, SOD2 | – |
| Cell Adhesion | C1QBP, DDX6, ICAM1, TGFB1I1 | STAT1 | – | DPP4, ITGA2, IDH1, KTN1 | PRKAR1A, PPP3CA, RAC1 | PRKCA |
| Metabolic Processing | APOL2, NUCB2, PFKL | SLC25A1 | RANBP2 | DLST, ECHS1, IDH1, NDUFB4 | ADH5, ALDH18A1, GLUD1, GLUD2, OGDH | CYB5A, COX1, PGP, PTGS1, PRKCA |
| Post-Translational Modifications | C1QBP, DDX6 | – | – | ADAR | SRRT | PRKCA |
| Gene Symbol | Protein Name | Protein Expression Levels Determined by MS | mRNA Levels Determined by qRT-PCR | Protein Levels Determined by Western Blot | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| WJ-MSCs | NFFs | ASFs | WJ-MSCs | NFFs | ASFs | WJ-MSCs | NFFs | ASFs | ||
| EPHA2 | EPH receptor A2 | + | − | − | +++ | + | − | +++ | + | + |
| SLC25A4 | ADP/ATP translocase 1 | + | − | − | ++ | − | − | +++ | − | − |
| TLN2 | Talin2 | + | − | + | − | + | +++ | − | − | +++ |
| LMF2 | Lipase maturation factor 2 | − | + | + | − | ++ | ++ | + | + | ++ |
| ITGA2 | CD49b/Integrin subunit alpha 2 | + | + | − | ++ | + | − | +++ | ++ | − |
| VDAC3 | Voltage-dependent anion channel 3 | + | − | − | + | + | + | +++ | + | +++ |
| SOD2 | Superoxide desmutase | + | − | + | + | − | − | +++ | − | − |
| CDH2 | CD325/N-Cadherin | − | + | − | − | +++ | − | |||
| ITGA5 | CD49e/Integrin subunit alpha 5 | ++ | + | + | ++ | + | + | ++ | + | + |
| IGF2BP3 | Insulin-like growth factor 2 mRNA binding protein 3 | − | − | + | ++ | + | ++ | |||
| PLEC | Plectin | − | + | + | + | + | + | ++ | ++ | ++ |
| CLE7 | RNA transcription, translation and transport factor | + | − | − | + | − | − | +++ | + | + |
| CDH5 | CD144/VE-cadherin | ++ | + | − | ++ | + | + | |||
| NEXN | Nexilin | + | − | − | ++ | + | − | + | + | + |
| MLCK1 | Myosin light chain kinase | + | − | − | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Madhoun, A.; Marafie, S.K.; Haddad, D.; Melhem, M.; Abu-Farha, M.; Ali, H.; Sindhu, S.; Atari, M.; Al-Mulla, F. Comparative Proteomic Analysis Identifies EphA2 as a Specific Cell Surface Marker for Wharton’s Jelly-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2020, 21, 6437. https://doi.org/10.3390/ijms21176437
Al Madhoun A, Marafie SK, Haddad D, Melhem M, Abu-Farha M, Ali H, Sindhu S, Atari M, Al-Mulla F. Comparative Proteomic Analysis Identifies EphA2 as a Specific Cell Surface Marker for Wharton’s Jelly-Derived Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2020; 21(17):6437. https://doi.org/10.3390/ijms21176437
Chicago/Turabian StyleAl Madhoun, Ashraf, Sulaiman K. Marafie, Dania Haddad, Motasem Melhem, Mohamed Abu-Farha, Hamad Ali, Sardar Sindhu, Maher Atari, and Fahd Al-Mulla. 2020. "Comparative Proteomic Analysis Identifies EphA2 as a Specific Cell Surface Marker for Wharton’s Jelly-Derived Mesenchymal Stem Cells" International Journal of Molecular Sciences 21, no. 17: 6437. https://doi.org/10.3390/ijms21176437
APA StyleAl Madhoun, A., Marafie, S. K., Haddad, D., Melhem, M., Abu-Farha, M., Ali, H., Sindhu, S., Atari, M., & Al-Mulla, F. (2020). Comparative Proteomic Analysis Identifies EphA2 as a Specific Cell Surface Marker for Wharton’s Jelly-Derived Mesenchymal Stem Cells. International Journal of Molecular Sciences, 21(17), 6437. https://doi.org/10.3390/ijms21176437