Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism

 , ,
, ,
Abstract
1. Introduction
2. G Protein-Coupled Receptors (GPCRs)
3. Histone Deacetylases (HDACs)
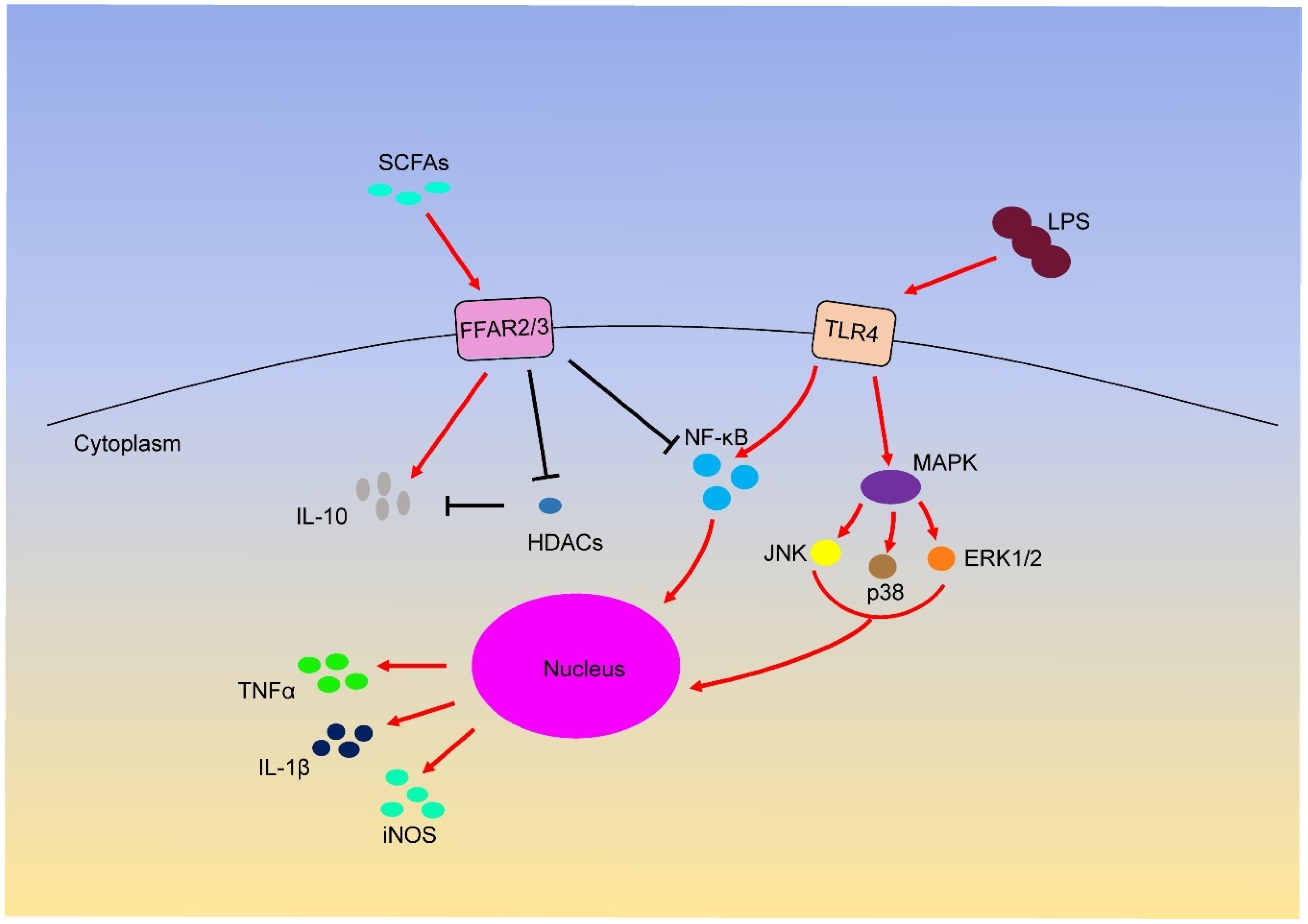
4. SCFAs and Inflammation
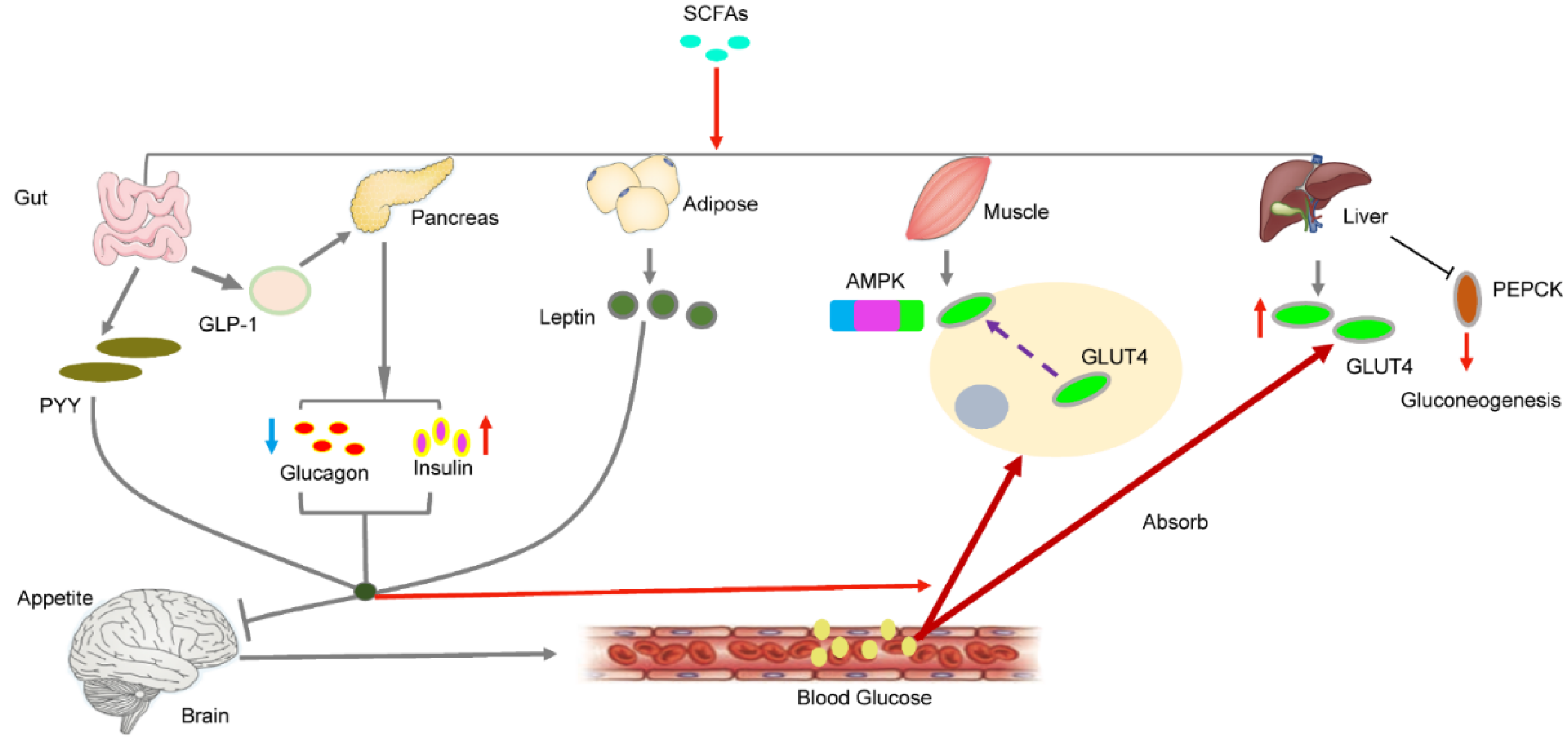
5. SCFAs and Glucose Metabolism
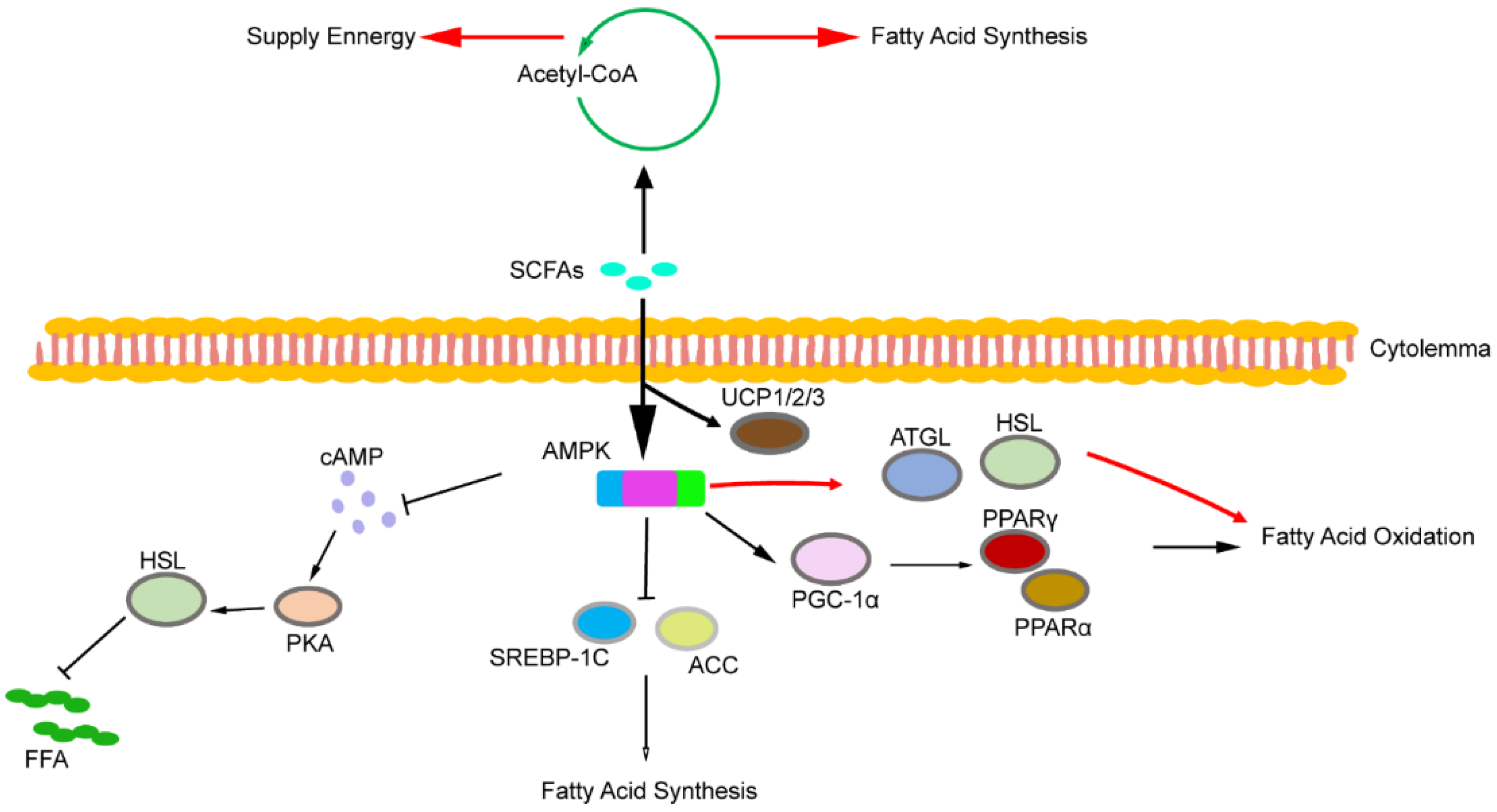
6. SCFAs and Lipid Metabolism
7. Summary
Funding
Conflicts of Interest
References
- Cook, S.I.; Sellin, J.H. Review Article: Short Chain Fatty Acids in Health and Disease. Aliment. Pharmacol Ther. 1998, 12, 499–507. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short Chain Fatty Acids in Human Large Intestine, Portal, Hepatic and Venous Blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Bergman, E.N. Energy Contributions of Volatile Fatty Acids from the Gastrointestinal Tract in Various Species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G.T. Regulation of Short-Chain Fatty Acid Production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Aceituno, L.; Esteban-Torres, M.; James, K.; Moreno, F.J.; Sinderen, D.V. Metabolism of Biosynthetic Oligosaccharides by Human-Derived Bifidobacterium Breve Ucc2003 and Bifidobacterium Longum Ncimb 8809. Int. J. Food Microbiol. 2020, 316, 108476. [Google Scholar] [CrossRef]
- Horiuchi, H.; Kamikado, K.; Aoki, R.; Suganuma, N.; Nishijima, T.; Nakatani, A.; Kimura, I. Bifidobacterium Animalis Subsp. Lactis Gcl2505 Modulates Host Energy Metabolism Via the Short-Chain Fatty Acid Receptor Gpr43. Sci. Rep. 2020, 10, 4158. [Google Scholar] [CrossRef]
- Smith, E.A.; Macfarlane, G.T. Enumeration of amino acid fermenting bacteria in the human large intestine: Effects of pH and starch on peptide metabolism and dissimilation of amino acids. FEMS Microbiol. Ecol. 1998, 25, 355–368. [Google Scholar] [CrossRef]
- Dai, Z.L.; Wu, G.; Zhu, W.Y. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar] [CrossRef]
- Duncan, S.H.; Louis, P.; Thomson, J.M.; Flint, H.J. The role of pH in determining the species composition of the human colonic microbiota. Environ. Microbiol. 2009, 11, 2112–2122. [Google Scholar] [CrossRef]
- Youngblut, N.D.; Reischer, G.H.; Walters, W.A.; Schuster, N.; Walzer, C.; Stalder, G.; Ley, R.E.; Farnleitner, A.H. Host diet and evolutionary history explain different aspects of gut microbiome diversity among vertebrate clades. Nat. Commun. 2019, 10, 2200. [Google Scholar] [CrossRef] [PubMed]
- Besten, G.D.; Eunen, K.V.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate Improves Insulin Sensitivity and Increases Energy Expenditure in Mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, T.; Suruga, K.; Oshima, Y.; Fukiharu, M.; Tsukamoto, Y.; Goda, T. Dietary Acetic Acid Reduces Serum Cholesterol and Triacylglycerols in Rats Fed a Cholesterol-Rich Diet. Br. J. Nutr. 2006, 95, 916–924. [Google Scholar] [CrossRef]
- Demigné, C.; Morand, C.; Levrat, M.A.; Besson, C.; Moundras, C.; Rémésy, C. Effect of Propionate on Fatty Acid and Cholesterol Synthesis and on Acetate Metabolism in Isolated Rat Hepatocytes. Br. J. Nutr. 1995, 74, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Todesco, T.; Rao, A.V.; Bosello, O.; Jenkins, D.J. Propionate Lowers Blood Glucose and Alters Lipid Metabolism in Healthy Subjects. Am. J. Clin. Nutr. 1991, 54, 860–865. [Google Scholar] [CrossRef]
- Brown, L.; Rosner, B.; Willett, W.W.; Sacks, F.M. Cholesterol-Lowering Effects of Dietary Fiber: A Meta-Analysis. Am. J. Clin. Nutr. 1999, 69, 30–42. [Google Scholar] [CrossRef]
- Causey, J.L.; Feirtag, J.M.; Gallaher, D.D.; Tungland, B.C.; Slavin, J.L. Effects of Dietary Inulin on Serum Lipids, Blood Glucose and the Gastrointestinal Environment in Hypercholesterolemic Men. Nutr. Res. 2000, 20, 191–201. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids in Control of Body Weight and Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Vadder, F.D.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-Produced Succinate Improves Glucose Homeostasis Via Intestinal Gluconeogenesis. Cell Metab. 2016, 24, 151–157. [Google Scholar] [CrossRef]
- Lewis, K.; Lutgendorff, F.; Phan, V.; Söderholm, J.D.; Sherman, P.M.; McKay, D.M. Enhanced Translocation of Bacteria across Metabolically Stressed Epithelia Is Reduced by Butyrate. Inflamm. Bowel Dis. 2010, 16, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Li, Z.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly Via Activation of Amp-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [PubMed]
- Gaudier, E.; Rival, M.; Buisine, M.P.; Robineau, I.; Hoebler, C. Butyrate Enemas Upregulate Muc Genes Expression but Decrease Adherent Mucus Thickness in Mice Colon. Physiol. Res. 2009, 58, 111–119. [Google Scholar] [PubMed]
- O’Keefe, S.J.D. Diet, Microorganisms and Their Metabolites, and Colon Cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef]
- Brown, E.M.; Sadarangani, M.; Finlay, B.B. The Role of the Immune System in Governing Host-Microbe Interactions in the Intestine. Nat. Immunol. 2013, 14, 660–667. [Google Scholar] [CrossRef]
- Li, Z.; Yi, C.; Katiraei, S.; Kooijman, S.; Zhou, E.; Chung, C.K.; Gao, Y.; Heuvel, J.V.D.; Meijer, O.C.; Berbée, J.F.P.; et al. Butyrate Reduces Appetite and Activates Brown Adipose Tissue Via the Gut-Brain Neural Circuit. Gut 2018, 67, 1269–1279. [Google Scholar] [CrossRef]
- Mollica, M.P.; Raso, G.M.; Cavaliere, G. Butyrate Regulates Liver Mitochondrial Function, Efficiency, and Dynamics in Insulin-Resistant Obese Mice. Diabetes 2017, 66, 1405–1418. [Google Scholar] [CrossRef]
- Vadder, F.D.; Kovatcheva-Datchary, P.; Goncalve, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Generated Metabolites Promote Metabolic Benefits Via Gut-Brain Neural Circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef]
- Szentirmai, É.; Millican, N.S.; Massie, A.R.; Kapás, L. Butyrate, a Metabolite of Intestinal Bacteria, Enhances Sleep. Sci. Rep. 2019, 9, 7035. [Google Scholar] [CrossRef]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G Protein-Coupled Receptors Gpr41 and Gpr43 Are Activated by Propionate and Other Short Chain Carboxylic Acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef]
- Poul, E.L.; Loison, C.; Struyf, S.; Springael, J.; Lannoy, V.; Decobecq, M.; Brezillon, S.; Dupriez, V.; Vassart, G.; Damme, J.V.; et al. Functional Characterization of Human Receptors for Short Chain Fatty Acids and Their Role in Polymorphonuclear Cell Activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G.; Stoddart, L.A.; Smith, N.J. Agonism and Allosterism: The Pharmacology of the Free Fatty Acid Receptors Ffa2 and Ffa3. Br. J. Pharmacol. 2009, 158, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Jupe, S.; Briscoe, C.P. A Family of Fatty Acid Binding Receptors. DNA Cell Biol. 2005, 24, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Kebede, M.A.; Alquier, T.; Latour, M.G.; Poitout, V. Lipid Receptors and Islet Function: Therapeutic Implications? Diabetes Obes. Metab. 2009, 4, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Halpern, K.B.; Veprik, A.; Rubins, N.; Naaman, O.; Walker, M.D. Gpr41 Gene Expression Is Mediated by Internal Ribosome Entry Site (Ires)-Dependent Translation of Bicistronic Mrna Encoding Gpr40 and Gpr41 Proteins. J. Biol. Chem. 2012, 287, 20154–20163. [Google Scholar] [CrossRef]
- Tazoe, H.; Otomo, Y.; Karaki, S.; Kato, I.; Fukami, Y.; Terasaki, M.; Kuwahara, A. Expression of Short-Chain Fatty Acid Receptor Gpr41 in the Human Colon. Biomed. Res. 2009, 30, 149–156. [Google Scholar] [CrossRef]
- Sykaras, A.G.; Demenis, C.; Case, R.M.; McLaughlin, J.T.; Smith, C.P. Duodenal Enteroendocrine I-Cells Contain Mrna Transcripts Encoding Key Endocannabinoid and Fatty Acid Receptors. PLoS ONE 2012, 7, e42373. [Google Scholar] [CrossRef]
- Karaki, S.; Mitsui, R.; Hayashi, H.; Kato, I.; Sugiya, H.; Iwanaga, T.; Furness, J.B.; Kuwahara, A. Short-Chain Fatty Acid Receptor, Gpr43, Is Expressed by Enteroendocrine Cells and Mucosal Mast Cells in Rat Intestine. Cell Tissue Res. 2006, 324, 353–360. [Google Scholar] [CrossRef]
- Hong, Y.; Nishimura, Y.; Hishikawa, D.; Tsuzuki, H.; Miyahara, H.; Gotoh, C.; Choi, K.; Feng, D.D.; Chen, C.; Lee, H.; et al. Acetate and Propionate Short Chain Fatty Acids Stimulate Adipogenesis Via Gpcr43. Endocrinology 2005, 146, 5092–5099. [Google Scholar] [CrossRef]
- Maslowski, K.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of Inflammatory Responses by Gut Microbiota and Chemoattractant Receptor Gpr43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Nilsson, C.; Swolin-Eide, D.; Ohlsso, C.; Eriksson, E.; Ho, H.; Björntorp, P.; Holmäng, A. Reductions in Adipose Tissue and Skeletal Growth in Rat Adult Offspring after Prenatal Leptin Exposure. J. Endocrinol. 2003, 176, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Pingitore, A.; Gonzalez-Abuin, N.; Ruz-Maldonado, I.; Huang, G.C.; Frost, G.; Persaud, S.J. Short Chain Fatty Acids Stimulate Insulin Secretion and Reduce Apoptosis in Mouse and Human Islets in Vitro: Role of Free Fatty Acid Receptor 2. Diabetes Obes. Metab. 2019, 21, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Tsoulou, C.; Ward, E.; Gower, E.; Bhudia, N.; Chowdhury, F.; Dean, T.W.; Faucher, N.; Gangar, A.; Dowell, S.J. Pharmacological Properties of Acid N-Thiazolylamide Ffa2 Agonists. Pharmacol. Res. Perspect. 2015, 3, e00141. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.; Stafford, S.; Coope, G.; Heffron, H.; Real, K.; Newman, R.; Davenport, R.; Barnes, M.; Grosse, J.; Cox, H. Selective Ffa2 Agonism Appears to Act Via Intestinal Pyy to Reduce Transit and Food Intake but Does Not Improve Glucose Tolerance in Mouse Models. Diabetes 2015, 64, 3763–3771. [Google Scholar] [CrossRef] [PubMed]
- Houslay, M.D.; Milligan, G. Tailoring Camp-Signalling Responses through Isoform Multiplicity. Trends Biochem. Sci. 1997, 22, 217–224. [Google Scholar] [CrossRef]
- Kimura, I.; Inoue, D.; Hirano, K.; Tsujimoto, G. The SCFA Receptor Gpr43 and Energy Metabolism. Front. Endocrinol. 2014, 5, 85. [Google Scholar] [CrossRef] [PubMed]
- Pirozzi, C.; Francisco, V.; Guida, F.D.; Gómez, R.; Lago, F.; Pino, J.; Meli, R.; Gualillo, O. Butyrate Modulates Inflammation in Chondrocytes Via Gpr43 Receptor. Cell Physiol. Biochem. 2018, 51, 228–243. [Google Scholar] [CrossRef]
- Nakajima, A.; Nakatani, A.; Hasegawa, S.; Irie, J.; Ozawa, K.; Tsujimoto, G.; Suganami, T.; Itoh, H.; Kimura, I. The Short Chain Fatty Acid Receptor Gpr43 Regulates Inflammatory Signals in Adipose Tissue M2-Type Macrophages. PLoS ONE 2017, 12, e0179696. [Google Scholar] [CrossRef]
- Mizuta, K.; Matoba, A.; Shibata, S.; Masaki, E.; Sr, C.W.E. Obesity-Induced Asthma: Role of Free Fatty Acid Receptors. Jpn. Dent. Sci. Rev. 2019, 55, 103–107. [Google Scholar] [CrossRef]
- Samuel, B.S.; Shaito, A.; Motoike, T.; Rey, F.E.; Backhed, F.; Manchester, J.K.; Hammer, R.E.; Williams, S.C.; Crowley, J.; Yanagisawa, M.; et al. Effects of the Gut Microbiota on Host Adiposity Are Modulated by the Short-Chain Fatty-Acid Binding G Protein-Coupled Receptor, Gpr41. Proc. Natl. Acad. Sci. USA 2008, 105, 16767–16772. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-Chain Fatty Acids and Ketones Directly Regulate Sympathetic Nervous System Via G Protein-Coupled Receptor 41 (Gpr41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef] [PubMed]
- Ohira, H.; Fujioka, Y.; Katagiri, C.; Mamoto, R.; Aoyama-Ishikawa, M.; Amako, K.; Izumi, Y.; Nishiumi, S.; Yoshida, M.; Usami, M.; et al. Butyrate Attenuates Inflammation and Lipolysis Generated by the Interaction of Adipocytes and Macrophages. J. Atheroscler. Thromb. 2013, 20, 425–442. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Esch, B.C.A.M.; Wagenaar, G.T.M.; Garssen, J.; Folkerts, G.; Henricks, P.A.J. Pro- and Anti-Inflammatory Effects of Short Chain Fatty Acids on Immune and Endothelial Cells. Eur. J. Pharmacol. 2018, 831, 52–59. [Google Scholar] [CrossRef]
- Miyamoto, J.; Kasubuchi, M.; Nakajima, A.; Kimura, I. Anti-Inflammatory and Insulin-Sensitizing Effects of Free Fatty Acid Receptors. Handb. Exp. Pharmacol. 2016, 236, 221–231. [Google Scholar]
- Nilsson, N.E.; Kotarsky, K.; Owman, C.; Olde, B. Identification of a Free Fatty Acid Receptor, Ffa2r, Expressed on Leukocytes and Activated by Short-Chain Fatty Acids. Biochem. Biophys. Res. Commun. 2003, 303, 1047–1052. [Google Scholar] [CrossRef]
- Stoddart, L.A.; Smith, N.J.; Jenkins, L.; Brown, A.J.; Milligan, G. Conserved Polar Residues in Transmembrane Domains V, Vi, and Vii of Free Fatty Acid Receptor 2 and Free Fatty Acid Receptor 3 Are Required for the Binding and Function of Short Chain Fatty Acids. J. Biol. Chem. 2008, 283, 32913–32924. [Google Scholar] [CrossRef]
- Colina, C.; Puhl, H.L.; Ikeda, S.R. Selective Tracking of Ffar3-Expressing Neurons Supports Receptor Coupling to N-Type Calcium Channels in Mouse Sympathetic Neurons. Sci. Rep. 2018, 8, 17379. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut Microbiota Metabolism of Dietary Fiber Influences Allergic Airway Disease and Hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.; Rey, F.; Wang, T.; et al. Olfactory Receptor Responding to Gut Microbiota-Derived Signals Plays a Role in Renin Secretion and Blood Pressure Regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef]
- Pluznick, J. A Novel SCFA Receptor, the Microbiota, and Blood Pressure Regulation. Gut Microb. 2014, 5, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Mizuta, K.; Sasaki, H.; Zhang, Y.; Matoba, A.; Sr, C.W.E. The Short-Chain Free Fatty Acid Receptor Ffar3 Is Expressed and Potentiates Contraction in Human Airway Smooth Muscle. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 318, L1248–L1260. [Google Scholar] [CrossRef] [PubMed]
- Ørgaard, A.; Jepsen, S.L.; Holst, J.J. Short-Chain Fatty Acids and Regulation of Pancreatic Endocrine Secretion in Mice. Islets 2019, 11, 103–111. [Google Scholar] [CrossRef]
- Priyadarshini, M.; Layden, B.T. Ffar3 Modulates Insulin Secretion and Global Gene Expression in Mouse Islets. Islets 2015, 7, e1045182. [Google Scholar] [CrossRef]
- Shimizu, H.; Masujima, Y.; Ushiroda, C.; Mizushima, R.; Taira, S.; Ohue-Kitano, R.; Kimura, I. Dietary Short-Chain Fatty Acid Intake Improves the Hepatic Metabolic Condition Via Ffar3. Sci. Rep. 2019, 9, 16574. [Google Scholar] [CrossRef] [PubMed]
- Wanders, D.; Graff, E.C.; Judd, R.L. Effects of High Fat Diet on Gpr109A and Gpr81 Gene Expression. Biochem. Biophys. Res. Commun. 2012, 425, 278–283. [Google Scholar] [CrossRef]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. Gpr109A Is a G-Protein-Coupled Receptor for the Bacterial Fermentation Product Butyrate and Functions as a Tumor Suppressor in Colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef]
- Benyó, Z.; Gille, A.; Kero, J.; Csiky, M.; Suchánková, M.C.; Nüsing, R.M.; Moers, A.; Pfeffer, K.; Offermanns, S. Gpr109A (Puma-G/Hm74a) Mediates Nicotinic Acid-Induced Flushing. J. Clin. Investig. 2005, 115, 3634–3640. [Google Scholar] [CrossRef]
- Zellner, C.; Pullinger, C.R.; Aouizerat, B.E.; Frost, P.H.; Kwok, P.; Malloy, M.J.; Kane, J.P. Variations in Human Hm74 (Gpr109b) and Hm74a (Gpr109A) Niacin Receptors. Human Mutat. 2005, 25, 18–21. [Google Scholar] [CrossRef]
- Offermanns, S. Hydroxy-Carboxylic Acid Receptor Actions in Metabolism. Trends Endocrinol. Metab. 2017, 28, 227–236. [Google Scholar] [CrossRef]
- Rezq, S.; Abdel-Rahman, A.A. Central Gpr109A Activation Mediates Glutamate-Dependent Pressor Response in Conscious Rats. J. Pharmacol. Exp. Ther. 2016, 356, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, X.; Lin, S.; Chen, Y.; Ma, S.; Fu, Y.; Wei, C.; Xu, W. Nicotinic Acid Receptor Gpr109A Exerts Anti-Inflammatory Effects through Inhibiting the Akt/mTOR Signaling Pathway in Min6 Pancreatic Β Cells. Ann. Clin. Lab. Sci. 2017, 47, 729–737. [Google Scholar] [PubMed]
- Elangovan, S.; Pathania, R.; Ramachandran, S.; Ananth, S.; Padia, R.N.; Lan, L.; Singh, N.; Martin, P.M.; Hawthorn, L.; Prasad, P.D.; et al. The Niacin/Butyrate Receptor Gpr109A Suppresses Mammary Tumorigenesis by Inhibiting Cell Survival. Cancer Res. 2014, 74, 1166–1178. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, M.; Kotlo, K.U.; Dudeja, P.K.; Layden, B.T. Role of Short Chain Fatty Acid Receptors in Intestinal Physiology and Pathophysiology. Compr. Physiol. 2018, 8, 1091–1115. [Google Scholar] [PubMed]
- Kaye, D.M.; Shihata, W.A.; Jama, H.A.; Tsyganov, K.; Ziemann, M.; Kiriazis, H.; Horlock, D.; Vijay, A.; Giam, B.; Vinh, A.; et al. Deficiency of Prebiotic Fiber and Insufficient Signaling through Gut Metabolite-Sensing Receptors Leads to Cardiovascular Disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef]
- Wang, N.; Guo, D.; Tian, X.; Lin, H.; Li, Y.; Chen, S.; Fu, Y.; Xu, W.; Wei, C. Niacin Receptor Gpr109A Inhibits Insulin Secretion and Is Down-Regulated in Type 2 Diabetic Islet Beta-Cells. Gen. Comp. Endocrinol. 2016, 237, 98–108. [Google Scholar] [CrossRef]
- Shi, Y.; Lai, X.; Ye, L.; Chen, K.; Cao, Z.; Gong, W.; Jin, L.; Wang, C.; Liu, M.; Liao, Y.; et al. Activated Niacin Receptor Hca2 Inhibits Chemoattractant-Mediated Macrophage Migration Via Gβγ/PKC/ERK1/2 Pathway and Heterologous Receptor Desensitization. Sci. Rep. 2017, 7, 42279. [Google Scholar] [CrossRef]
- Borthakur, A.; Priyamvada, S.; Kumar, A.; Natarajan, A.A.; Gill, R.K.; Alrefai, W.A.; Dudeja, P.K. A Novel Nutrient Sensing Mechanism Underlies Substrate-Induced Regulation of Monocarboxylate Transporter-1. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1126–G1133. [Google Scholar] [CrossRef]
- Linda Sealy, R.C. The Effect of Sodium Butyrate on Histone Modification. Cell 1978, 14, 115–121. [Google Scholar] [CrossRef]
- Boffa, L.C.; Vidali, G.; Mann, R.S.; Allfrey, V.G. Suppression of Histone Deacetylation in Vivo and in Vitro by Sodium Butyrate. J. Biol. Chem. 1978, 253, 3364–3366. [Google Scholar]
- Waldecker, M.; Kautenburger, T.; Daumann, H.; Busch, C.; Schrenk, D. Inhibition of Histone-Deacetylase Activity by Short-Chain Fatty Acids and Some Polyphenol Metabolites Formed in the Colon. J. Nutr. Biochem. 2008, 19, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.L.; Rosenberger, T.A. Acetate Supplementation Increases Brain Histone Acetylation and Inhibits Histone Deacetylase Activity and Expression. Mol. Cell Biochem. 2011, 352, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Bulusu, V.; Tumanov, S.; Michalopoulou, E.; Broek, N.J.; MacKay, G.; Nixon, C.; Dhayade, S.; Schug, Z.T.; Voorde, J.V.; Blyth, K.; et al. Acetate Recapturing by Nuclear Acetyl-CoA Synthetase 2 Prevents Loss of Histone Acetylation during Oxygen and Serum Limitation. Cell Rep. 2017, 18, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The Microbial Metabolite Butyrate Regulates Intestinal Macrophage Function Via Histone Deacetylase Inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota Metabolite Short Chain Fatty Acids, Gpcr, and Inflammatory Bowel Diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, Z.; Hu, Y.; Dong, S. Butyrate-Induced Gpr41 Activation Inhibits Histone Acetylation and Cell Growth. J. Genet. Genom. 2012, 39, 375–384. [Google Scholar] [CrossRef]
- Czepiel, J.; Biesiada, G.; Brzozowski, T.; Ptak-Belowska, A.; Perucki, W.; Birczynska, M.; Jurczyszyn, A.; Strzalka, M.; Targosz, A.; Garlicki, A. The Role of Local and Systemic Cytokines in Patients Infected with Clostridium Difficile. J. Physiol. Pharmacol. 2014, 65, 695–703. [Google Scholar]
- Park, B.S.; Lee, J. Recognition of Lipopolysaccharide Pattern by TLR4 Complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef]
- Hagar, J.A.; Powell, D.A.; Aachoui, Y.; Ernst, R.K.; Miao, E.A. Cytoplasmic LPS Activates Caspase-11: Implications in TLR4-Independent Endotoxic shock. Science 2013, 341, 1250–1253. [Google Scholar] [CrossRef]
- Adrie, C.; Pinsky, M.R. The Inflammatory Balance in Human Sepsis. Intensive Care Med. 2000, 26, 364–375. [Google Scholar] [CrossRef]
- Baeuerle, P.A.; Baichwal, V.R. NF-Kappa B as a Frequent Target for Immunosuppressive and Anti-Inflammatory Molecules. Adv. Immunol. 1997, 65, 111–137. [Google Scholar]
- Pahl, H.L. Activators and Target Genes of Rel/NF-Kappab Transcription Factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.F.; Malik, A.B. NF-Kappa B Activation as a Pathological Mechanism of Septic Shock and Inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L622–L645. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Feng, G.; Wang, G.; Liu, G. p38MAPK Inhibition Attenuates LPS-induced Acute Lung Injury Involvement of NF-kappaB Pathway. Eur. J. Pharmacol. 2008, 584, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Du, R.; Zhu, X.; Yin, S.; Wang, J.; Cui, H.; Cao, W.; Lowenstein, C.J. Histone Deacetylase Isoforms Regulate Innate Immune Responses by Deacetylating Mitogen-Activated Protein Kinase Phosphatase-1. J. Leukoc. Biol. 2014, 95, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Subedi, L.; Venkatesan, R.; Kim, S.Y. Neuroprotective and Anti-Inflammatory Activities of Allyl Isothiocyanate Through Attenuation of JNK/NF-κB/TNF-α Signaling. Int. J. Mol. Sci. 2017, 18, 1423. [Google Scholar] [CrossRef]
- Vinolo, M.A.R.; Rodrigues, H.G.; Hatanaka, E.; Sato, F.T.; Sampaio, S.C.; Curi, R. Suppressive Effect of Short-Chain Fatty Acids on Production of Proinflammatory Mediators by Neutrophils. J. Nutr. Biochem. 2011, 22, 849–855. [Google Scholar] [CrossRef]
- Xiao, T.; Wu, S.; Yan, C.; Zhao, C.; Jin, H.; Yan, N.; Xu, J.; Wu, Y.; Li, C.; Shao, Q.; et al. Butyrate Upregulates the TLR4 Expression and the Phosphorylation of MAPKs and NK-κB in Colon Cancer Cell in vitro. Oncol. Lett. 2018, 16, 4439–4447. [Google Scholar] [CrossRef]
- Masui, R.; Sasaki, M.; Funaki, Y.; Ogasawara, N.; Mizuno, M.; Iida, A.; Izawa, S.; Kondo, Y.; Ito, Y.; Tamura, Y.; et al. G Protein-Coupled Receptor 43 Moderates Gut Inflammation through Cytokine Regulation from Mononuclear Cells. Inflamm. Bowel Dis. 2013, 19, 2848–2856. [Google Scholar] [CrossRef]
- Aoyama, M.; Kotani, J.; Usami, M. Butyrate and Propionate Induced Activated or Non-Activated Neutrophil Apoptosis via HDAC Inhibitor Activity but Without Activating GPR-41/GPR-43 Pathways. Nutrition 2010, 26, 653–661. [Google Scholar] [CrossRef]
- Vinolo, M.A.R.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of Inflammation by Short Chain Fatty Acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Halnes, I.; Baines, K.J.; Berthon, B.S.; MacDonald-Wicks, L.K.; Gibson, P.G.; Wood, L.G. Soluble Fibre Meal Challenge Reduces Airway Inflammation and Expression of GPR43 and GPR41 in Asthma. Nutrients 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Schaub, A.; Fütterer, A.; Pfeffer, K. PUMA-G, an IFN-gamma-inducible Gene in Macrophages Is a Novel Member of the Seven Transmembrane Spanning Receptor Superfamily. Eur. J. Immunol. 2001, 31, 3714–3739. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-chain Fatty Acids Activate GPR41 and GPR43 on Intestinal Epithelial Cells to Promote Inflammatory Responses in Mice. Gastroenterology 2013, 145, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Ribola, F.A.; Cançado, F.B.; Schoueri, J.H.M.; Toni, V.F.; Medeiros, V.H.; Feder, D. Effects of SGLT2 Inhibitors on Weight Loss in Patients With Type 2 Diabetes Mellitus. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 199–211. [Google Scholar]
- Batterham, R.L.; Cowley, M.A.; Small, C.J.; Herzog, H.; Cohen, M.A.; Dakin, C.L.; Wren, A.M.; Brynes, A.E.; Low, M.J.; Ghatei, M.A.; et al. Gut Hormone PYY(3–36) Physiologically Inhibits Food Intake. Nature 2002, 418, 650–654. [Google Scholar] [CrossRef]
- Hoek, A.M.; Heijboer, A.C.; Corssmit, E.P.M.; Voshol, P.J.; Romijn, J.A.; Havekes, L.M.; Pijl, H. PYY3–36 Reinforces Insulin Action on Glucose Disposal in Mice Fed a High-Fat Diet. Diabetes 2004, 53, 1949–1952. [Google Scholar] [CrossRef]
- Mazibuko, S.E.; Muller, C.J.F.; Joubert, E.; Beer, D.; Johnson, R.; Opoku, A.R.; Louw, J. Amelioration of Palmitate-Induced Insulin Resistance in C₂C₁₂ Muscle Cells by Rooibos (Aspalathus Linearis). Phytomedicine 2013, 20, 813–822. [Google Scholar] [CrossRef]
- Barrera, J.G.; Sandoval, D.A.; D’Alessio, D.A.; Seeley, R.J. GLP-1 and Energy Balance: An Integrated Model of Short-Term and Long-Term Control. Nat. Rev. Endocrinol. 2011, 7, 507–516. [Google Scholar] [CrossRef]
- Sakakibara, S.; Yamauchi, T.; Oshima, Y.; Tsukamoto, Y.; Kadowaki, T. Acetic acid activates hepatic AMPK and reduces hyperglycemia in diabetic KK-A(y) mice. Biochem. Biophys. Res. Commun. 2006, 344, 597–604. [Google Scholar] [CrossRef]
- Fujikawa, T.; Berglund, E.D.; Patel, V.R.; Ramadori, G.; Vianna, C.R.; Vong, L.; Thorel, F.; Chera, S.; Herrera, P.L.; Lowell, B.B.; et al. Leptin Engages a Hypothalamic Neurocircuitry to Permit Survival in the Absence of Insulin. Cell Metab. 2013, 18, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, T.; Chuang, J.; Sakata, I.; Ramadori, G.; Coppari, R. Leptin Therapy Improves Insulin-Deficient Type 1 Diabetes by Cns-Dependent Mechanisms in Mice. Proc. Natl. Acad. Sci. USA 2010, 107, 17391–17396. [Google Scholar] [CrossRef] [PubMed]
- Minokoshi, Y.; Kim, Y.; Peroni, O.D.; Fryer, L.G.; Müller, C.; Carling, D.; Kahn, B.B. Leptin Stimulates Fatty-Acid Oxidation by Activating Amp-Activated Protein Kinase. Nature 2002, 415, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Deng, B. Study the Mechanism of Calcium Signal- and Short Chain Fatty Acids-Regulated Glucose Metabolism. Ph.D. Thesis, Tianjin Medical University, Tianjin, China, 2018. [Google Scholar]
- Hong, J.; Yimin Jia, Y.; Pan, S.; Jia, L.; Li, H.; Han, Z.; Cai, D.; Zhao, R. Butyrate Alleviates High Fat Diet-Induced Obesity through Activation of Adiponectin-Mediated Pathway and Stimulation of Mitochondrial Function in the Skeletal Muscle of Mice. Oncotarget 2016, 7, 56071–56082. [Google Scholar] [CrossRef]
- Besten, G.; Gerding, A.; Dijk, T.H.; Ciapaite, J.; Bleeker, A.; Eunen, K.; Havinga, R.; Groen, A.K.; Reijngoud, D.J.; Bakker, B.M. Protection against the Metabolic Syndrome by Guar Gum-Derived Short-Chain Fatty Acids Depends on Peroxisome Proliferator-Activated Receptor Γ and Glucagon-Like Peptide-1. PLoS ONE 2015, 10, e0136364. [Google Scholar] [CrossRef]
- Merrill, G.F.; Kurth, E.J.; Hardie, D.G.; Winder, W.W. Aica Riboside Increases Amp Activated Protein Kinase, Fatty Acid Oxidati on, and Glucose Uptake in Rat Muscle. Am. J. Physiol. 1997, 273, E1107–E1119. [Google Scholar]
- Hardie, D.G.; Hawley, S.A.; Scott, J.W. AMP-activated Protein Kinase—Development of the Energy Sensor Concept. J. Physiol. 2006, 574, 7–15. [Google Scholar] [CrossRef]
- Kondo, T.; Kishi, M.; Fushimi, T.; Kaga, T. Acetic Acid Upregulates the Expression of Genes for Fatty Acid Oxidation Enzymes in Liver to Suppress Body Fat Accumulation. J. Agric. Food Chem. 2009, 57, 5982–5986. [Google Scholar] [CrossRef]
- Kim, G.T.; Lee, S.H.; Kim, Y.M. Quercetin Regulates Sestrin 2-Ampk-Mtor Signaling Pathway and Induces Apoptosis Via Increased Intracellular Ros in Hct116 Colon Cancer Cells. J. Cancer Prev. 2013, 18, 264–270. [Google Scholar] [CrossRef]
- Liu, H.; Peng, H.; Xiang, H.; Guo, L.; Chen, R.; Zhao, S.; Chen, W.; Chen, P.; Lu, H.; Chen, S. TWEAK/Fn14 Promotes Oxidative Stress Through AMPK/PGC-1α/MnSOD Signaling Pathway in Endothelial Cells. Mol. Med. Rep. 2017, 17, 1998–2004. [Google Scholar] [CrossRef]
- Ma, W.; Wang, J.; Bu, X.; Zhang, H.; Zhang, J.; Zhang, X.; He, Y.; Wang, D.; Zhang, Z.; Meng, F. Effects of Polygonum Cuspidatum on AMPK-FOXO3α Signaling Pathway in Rat Model of Uric Acid-Induced Renal Damage. Chin. J. Integr. Med. 2019, 25, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Zambell, K.L.; Fitch, M.D.; Fleming, S.E. Acetate and Butyrate Are the Major Substrates for De Novo Lipogenesis in Rat Colonic Epithelial Cells. J. Nutr. 2003, 133, 3509–3515. [Google Scholar] [CrossRef] [PubMed]
- Kindt, A.; Liebisch, G.; Clacel, T.; Haller, D.; Hörmannsperger, G.; Yoon, H.; Kolmeder, D.; Sigruener, A.; Krautbauer, S.; Seeliger, C.; et al. The gut microbiota promotes hepatic fatty acid desaturation and elongation in mice. Nat. Commun. 2018, 9, 3760. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.B.; Lamb, J.D.; Hurst, R.W.; Chesser, D.G.; Ellingson, W.J.; Greenwood, L.J.; Porter, B.B.; Herway, S.T.; Winder, W.W. Endurance Training Increases Skeletal Muscle LKB1 and PGC-1alpha Protein Abundance: Effects of Time and Intensity. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E960–E968. [Google Scholar] [CrossRef]
- Terada, S.; Goto, M.; Kato, M.; Kawanaka, K.; Shimokawa, T.; Tabata, I. Effects of Low-Intensity Prolonged Exercise on PGC-1 mRNA Expression in Rat Epitrochlearis Muscle. Biochem. Biophys. Res. Commun. 2002, 296, 350–354. [Google Scholar] [CrossRef]
- Wan, Z.; Root-McCaig, J.; Castellani, L.; Kemp, B.E.; Steinberg, G.R.; Wright, D.C. Evidence for the Role of AMPK in Regulating PGC-1 Alpha Expression and Mitochondrial Proteins in Mouse Epididymal Adipose Tissue. Obesity 2014, 22, 730–738. [Google Scholar] [CrossRef]
- Yan, M.; Audet-Walsh, É.; Manteghi, S.; Dufour, C.R.; Walker, B.; Baba, M.; St-Pierre, J.; Giguère, V.; Pause, A. Chronic AMPK Activation via Loss of FLCN Induces Functional Beige Adipose Tissue Through PGC-1α/ERRα. Genes Dev. 2016, 30, 1034–1046. [Google Scholar] [CrossRef]
- Muoio, D.M.; Way, J.M.; Tanner, C.J.; Winegar, D.A.; Kliewer, S.A.; Houmard, J.A.; Kraus, W.E.; Dohm, G.L. Peroxisome Proliferator-Activated Receptor-Alpha Regulates Fatty Acid Utilization in Primary Human Skeletal Muscle Cells. Diabetes 2002, 51, 901–909. [Google Scholar] [CrossRef]
- Lin, J.; Handschin, C.; Spiegelman, B.M. Metabolic Control through the PGC-1 Family of Transcription Coactivators. Cell Metab. 2005, 1, 361–370. [Google Scholar] [CrossRef]
- Jäger, S.; Handschin, C.; St-Pierre, J.; Spiegelman, B.M. AMP-activated Protein Kinase (AMPK) Action in Skeletal Muscle via Direct Phosphorylation of PGC-1alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar] [CrossRef]
- Huang, M.; Zhou, C.; Zhang, Y.; Zhang, X.; Xu, W.; Lin, J.; Wang, P. Salvianolic Acid B Ameliorates Hyperglycemia and Dyslipidemia in db/db Mice Through the AMPK Pathway. Cell Physiol. Biochem. 2016, 40, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Auwerx, J. AMP-activated Protein Kinase and Its Downstream Transcriptional Pathways. Cell Mol. Life Sci. 2010, 67, 3407–3423. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Song, J.; Li, J.; Wang, H.; Zhang, Y.; Suo, H. Synbiotic Consisting of Lactobacillus Plantarum S58 and Hull-Less Barley β-glucan Ameliorates Lipid Accumulation in Mice Fed With a High-Fat Diet by Activating AMPK Signaling and Modulating the Gut Microbiota. Carbohydr. Polym. 2020, 243, 116398. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Kang, J.S.; Park, Y.H.; Je, B.; Lee, Y.; Kang, N.; Park, S.; Hwang, D.Y.; Choi, Y.W. S-petasin Inhibits Lipid Accumulation in Oleic Acid-Induced HepG2 Cells Through Activation of the AMPK Signaling Pathway. Food Funct. 2020, 11, 5664–5673. [Google Scholar] [CrossRef] [PubMed]
- Carmen, G.; Víctor, S. Signalling Mechanisms Regulating Lipolysis. Cell Signal. 2006, 18, 401–408. [Google Scholar] [CrossRef]
- Jocken, J.W.E.; Hernández, M.A.G.; Hoebers, N.T.H.; Beek, C.M.; Essers, Y.P.G.; Blaak, E.E.; Canfora, E.E. Short-Chain Fatty Acids Differentially Affect Intracellular Lipolysis in a Human White Adipocyte Model. Front. Endocrinol. 2018, 8, 372. [Google Scholar] [CrossRef]
- Yap, F.; Craddock, L.; Yang, J. Mechanism of AMPK Suppression of LXR-dependent Srebp-1c Transcription. Int. J. Biol. Sci. 2011, 7, 645–650. [Google Scholar] [CrossRef]
- Besten, G.; Bleeker, A.; Gerding, A.; Eunenet, K.; Havinga, R.; Dijk, T.; Oosterveer, M.; Jonker, J.; Groen, A.; Reijngoud, D.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARγ-Dependent Switch From Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| SCFAs Receptors | Alternative Names | G-Protein Coupling | Affinity | Expression | References |
|---|---|---|---|---|---|
| FFAR2 | FFA2; GPR43 | Gq/G11; Gi | C2=C3>C4>C5=C1 | white adipocytes, immune cells, islet α and β cells, intestinal enteroendocrine cells | [34,35,36,37,38,39,40,41,42] |
| FFAR3 | FFA3; GPR41 | Gi | C3=C4=C5>C2>C1 | immune cells, enteric neurons and sympathetic ganglia, intestinal enteroendocrine cells, islet α and β cells, | [34,35,36,37,51,52] |
| GPR109A | HCA2 | Gi | C4, niacin | Macrophages, immune cells, Adipocytes, β cells microvascular endothelial cells, microglial cells | [66,67,68,69,70,71,72,73] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. https://doi.org/10.3390/ijms21176356
He J, Zhang P, Shen L, Niu L, Tan Y, Chen L, Zhao Y, Bai L, Hao X, Li X, et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. International Journal of Molecular Sciences. 2020; 21(17):6356. https://doi.org/10.3390/ijms21176356
Chicago/Turabian StyleHe, Jin, Peiwen Zhang, Linyuan Shen, Lili Niu, Ya Tan, Lei Chen, Ye Zhao, Lin Bai, Xiaoxia Hao, Xuewei Li, and et al. 2020. "Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism" International Journal of Molecular Sciences 21, no. 17: 6356. https://doi.org/10.3390/ijms21176356
APA StyleHe, J., Zhang, P., Shen, L., Niu, L., Tan, Y., Chen, L., Zhao, Y., Bai, L., Hao, X., Li, X., Zhang, S., & Zhu, L. (2020). Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. International Journal of Molecular Sciences, 21(17), 6356. https://doi.org/10.3390/ijms21176356