CpG Site-Specific Regulation of Metallothionein-1 Gene Expression


Abstract
1. Introduction
2. Results and Discussion
2.1. Characterization of P1798 Cells Treated with 5-AzaC Plus Cd
2.2. Effect of CpG Methylation in the 5′-Flanking Region of the MT1 Gene on Lucia Luciferase Reporter Gene Expression
2.3. Involvement of the MTF1–MRE Pathway on the Inhibition of MT1 Expression by CpG Methylation
3. Materials and Methods
3.1. Cell Culture and Treatment
3.2. Quantitative Reverse Transcription PCR
3.3. PrimeFlow RNA Assay
3.4. Bisulfite Genomic Sequencing Assay
3.5. Luciferase Reporter Assay
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MT | Metallothionein |
| ROS | Reactive oxygen species |
| MRE | Metal response element |
| MTF1 | MRE-binding transcription factor-1 |
| HAT | histone acetyltransferase |
| 5-AzaC | 5-azacytidine |
| MEF | Mouse embryonic fibroblast |
| MTF1-KO | MTF1-knockout |
| qPCR | quantitative PCR |
References
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef]
- Michalska, A.E.; Choo, K.H. Targeting and germ-line transmission of a null mutation at the metallothionein I and II loci in mouse. Proc. Natl. Acad. Sci. USA 1993, 90, 8088–8092. [Google Scholar] [CrossRef] [PubMed]
- Masters, B.A.; Kelly, E.J.; Quaife, C.J.; Brinster, R.L.; Palmiter, R.D. Targeted disruption of metallothionein I and II genes increases sensitivity to cadmium. Proc. Natl. Acad. Sci. USA 1994, 91, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.D.; Liu, J.; Diwan, B.A. Metallothionein protection of cadmium toxicity. Toxicol. Appl. Pharmacol. 2009, 238, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J.; Vašák, M. Possible role for metallothionein in protection against radiation-induced oxidative stress. Kinetics and mechanism of its reaction with superoxide and hydroxyl radicals. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. 1985, 827, 36–44. [Google Scholar] [CrossRef]
- Chiaverini, N.; De Ley, M. Protective effect of metallothionein on oxidative stress-induced DNA damage. Free Radic. Res. 2010, 44, 605–613. [Google Scholar] [CrossRef]
- Ling, X.-B.; Wei, H.-W.; Wang, J.; Kong, Y.-Q.; Wu, Y.-Y.; Guo, J.-L.; Li, T.-F.; Li, J.-K. Mammalian Metallothionein-2A and Oxidative Stress. Int. J. Mol. Sci. 2016, 17, 1483. [Google Scholar] [CrossRef]
- Kimura, T.; Kambe, T. The Functions of Metallothionein and ZIP and ZnT Transporters: An Overview and Perspective. Int. J. Mol. Sci. 2016, 17, 336. [Google Scholar] [CrossRef]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef]
- Henikoff, S.; Greally, J.M. Epigenetics, cellular memory and gene regulation. Curr. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Luu, P.-L.; Stirzaker, C.; Clark, S.J. Methyl-CpG-binding domain proteins: Readers of the epigenome. Epigenomics 2015, 7, 1051–1073. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Itoh, N.; Andrews, G.K. Mechanisms of Heavy Metal Sensing by Metal Response Element-binding Transcription Factor-1. J. Health Sci. 2009, 55, 484–494. [Google Scholar] [CrossRef]
- Searle, P.F.; Stuart, G.W.; Palmiter, R.D. Building a metal-responsive promoter with synthetic regulatory elements. Mol. Cell. Biol. 1985, 5, 1480–1489. [Google Scholar] [CrossRef] [PubMed]
- Labbé, S.; Prévost, J.; Remondelli, P.; Leone, A.; Séguin, C. A nuclear factor binds to the metal regulatory elements of the mouse gene encoding metallothionein-I. Nucleic Acids Res. 1991, 19, 4225–4231. [Google Scholar] [CrossRef] [PubMed]
- Heuchel, R.; Radtke, F.; Georgiev, O.; Stark, G.; Aguet, M.; Schaffner, W. The transcription factor MTF-1 is essential for basal and heavy metal-induced metallothionein gene expression. EMBO J. 1994, 13, 2870–2875. [Google Scholar] [CrossRef]
- Li, Y.; Andrews, G.K.; Laity, J.H.; Kimura, T.; Huyck, R.W. Zinc-Induced Formation of a Coactivator Complex Containing the Zinc-Sensing Transcription Factor MTF-1, p300/CBP, and Sp1. Mol. Cell. Biol. 2008, 28, 4275–4284. [Google Scholar] [CrossRef]
- Kimura, T.; Li, Y.; Okumura, F.; Itoh, N.; Nakanishi, T.; Sone, T.; Isobe, M.; Andrews, G.K. Chromium (VI) inhibits mouse metallothionein-I gene transcription by preventing the zinc-dependent formation of an MTF-1-p300 complex. Biochem. J. 2008, 415, 477–482. [Google Scholar] [CrossRef]
- Kimura, T.; Okumura, F.; Onodera, A.; Nakanishi, T.; Itoh, N.; Isobe, M. Chromium (VI) inhibits mouse metallothionein-I gene transcription by modifying the transcription potential of the co-activator p300. J. Toxicol. Sci. 2011, 36, 173–180. [Google Scholar] [CrossRef]
- Majumder, S.; Ghoshal, K.; Li, Z.; Bo, Y.; Jacob, S.T. Silencing of metallothionein-I gene in mouse lymphosarcoma cells by methylation. Oncogene 1999, 18, 6287–6295. [Google Scholar] [CrossRef]
- Ghoshal, K.; Majumder, S.; Li, Z.; Dong, X.; Jacob, S.T. Suppression of Metallothionein Gene Expression in a Rat Hepatoma Because of Promoter-specific DNA Methylation. J. Biol. Chem. 2000, 275, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, K.; Datta, J.; Majumder, S.; Bai, S.; Dong, X.; Parthun, M.; Jacob, S.T. Inhibitors of Histone Deacetylase and DNA Methyltransferase Synergistically Activate the Methylated Metallothionein I Promoter by Activating the Transcription Factor MTF-1 and Forming an Open Chromatin Structure. Mol. Cell. Biol. 2002, 22, 8302–8319. [Google Scholar] [CrossRef] [PubMed]
- Majumder, S.; Kutay, H.; Datta, J.; Summers, D.; Jacob, S.T.; Ghoshal, K. Epigenetic regulation of metallothionein-I gene expression: Differential regulation of methylated and unmethylated promoters by DNA methyltransferases and methyl CpG binding proteins. J. Cell. Biochem. 2006, 97, 1300–1316. [Google Scholar] [CrossRef] [PubMed]
- Okumura, F.; Li, Y.; Itoh, N.; Nakanishi, T.; Isobe, M.; Andrews, G.K.; Kimura, T. The zinc-sensing transcription factor MTF-1 mediates zinc-induced epigenetic changes in chromatin of the mouse metallothionein-I promoter. Biochim. Biophys. Acta 2011, 1809, 56–62. [Google Scholar] [CrossRef]
- Kurita, H.; Ohsako, S.; Hashimoto, S.; Yoshinaga, J.; Tohyama, C. Prenatal zinc deficiency-dependent epigenetic alterations of mouse metallothionein-2 gene. J. Nutr. Biochem. 2013, 24, 256–266. [Google Scholar] [CrossRef]
- Kimura, T.; Hosaka, T.; Nakanishi, T.; Aozasa, O. Long-term cadmium exposure enhances metallothionein-1 induction after subsequent exposure to high concentrations of cadmium in P1798 mouse lymphosarcoma cells. J. Toxicol. Sci. 2019, 44, 309–316. [Google Scholar] [CrossRef]
- de Andres, M.C.; Imagawa, K.; Hashimoto, K.; Gonzalez, A.; Roach, H.I.; Goldring, M.B.; Oreffo, R.O. Loss of methylation in CpG sites in the NF-kappaB enhancer elements of inducible nitric oxide synthase is responsible for gene induction in human articular chondrocytes. Arthritis Rheum. 2013, 65, 732–742. [Google Scholar] [CrossRef]
- Ling, L.; Ren, M.; Yang, C.; Lao, G.; Chen, L.; Luo, H.; Feng, Z.; Yan, L. Role of site-specific DNA demethylation in TNFalpha-induced MMP9 expression in keratinocytes. J. Mol. Endocrinol. 2013, 50, 279–290. [Google Scholar] [CrossRef]
- Han, W.; Shi, M.; Spivack, S.D. Site-specific methylated reporter constructs for functional analysis of DNA methylation. Epigenetics 2013, 8, 1176–1187. [Google Scholar] [CrossRef]
- Urbano, A.; Smith, J.; Weeks, R.J.; Chatterjee, A. Gene-Specific Targeting of DNA Methylation in the Mammalian Genome. Cancers 2019, 11, 1515. [Google Scholar] [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef] [PubMed]
- Henning, A.L.; Sampson, J.N.B.; McFarlin, B.K. Measurement of Low-Abundance Intracellular mRNA Using Amplified FISH Staining and Image-Based Flow Cytometry. Curr. Protoc. Cytom. 2016, 76. [Google Scholar] [CrossRef] [PubMed]
- Kumaki, Y.; Oda, M.; Okano, M. QUMA: Quantification tool for methylation analysis. Nucleic Acids Res. 2008, 36, W170–W175. [Google Scholar] [CrossRef] [PubMed]






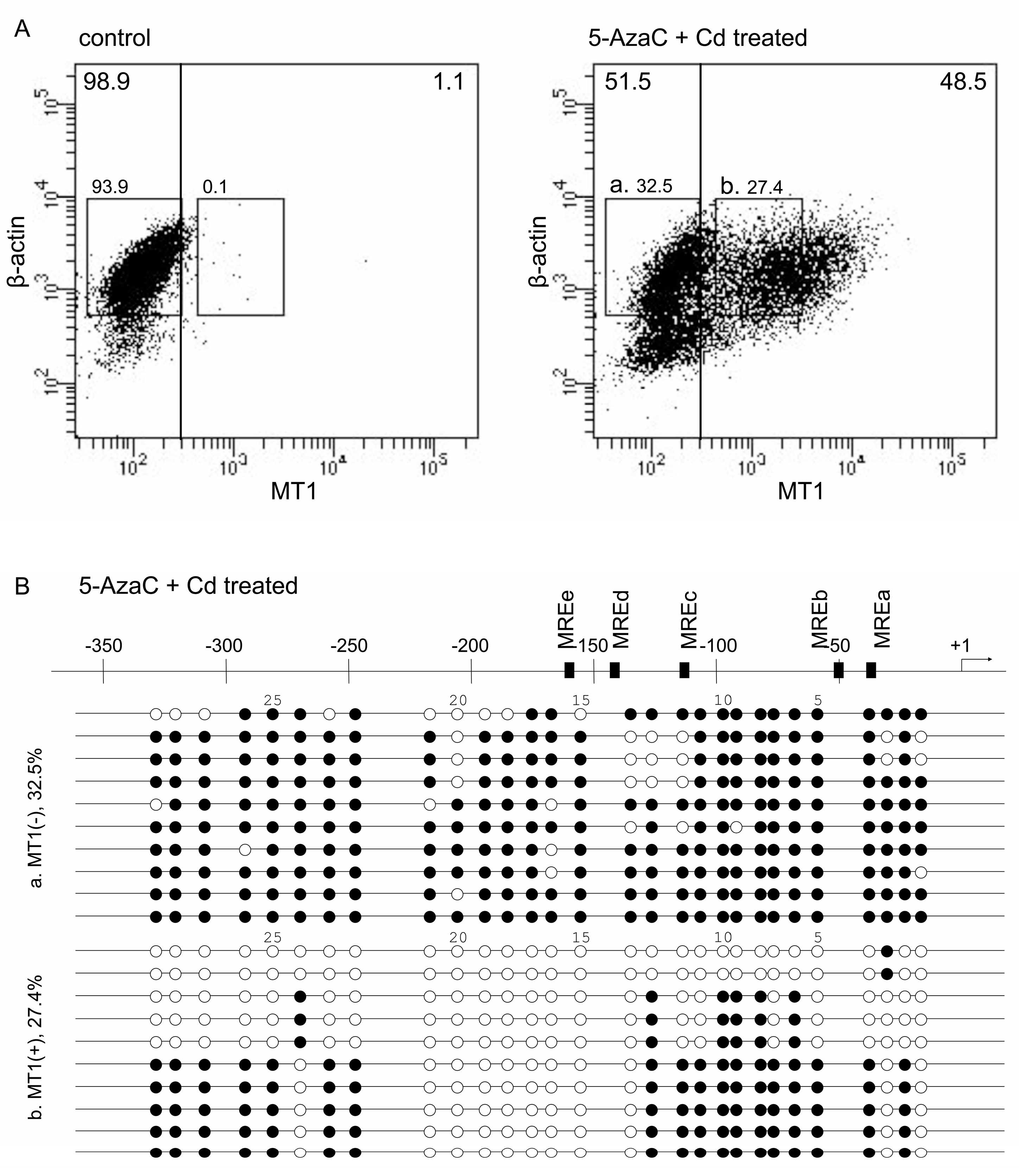{kind=link}
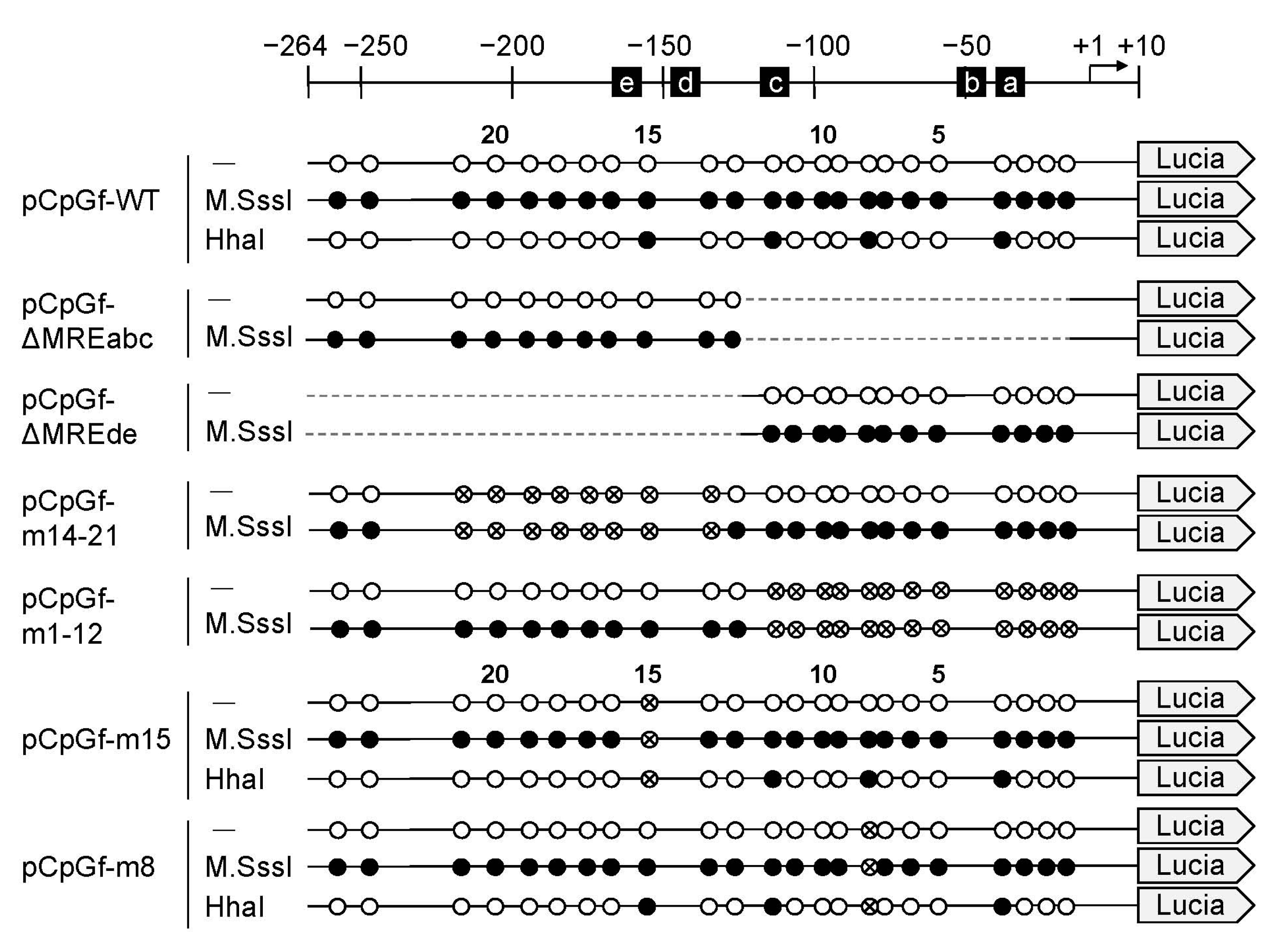{kind=link}
{kind=link}
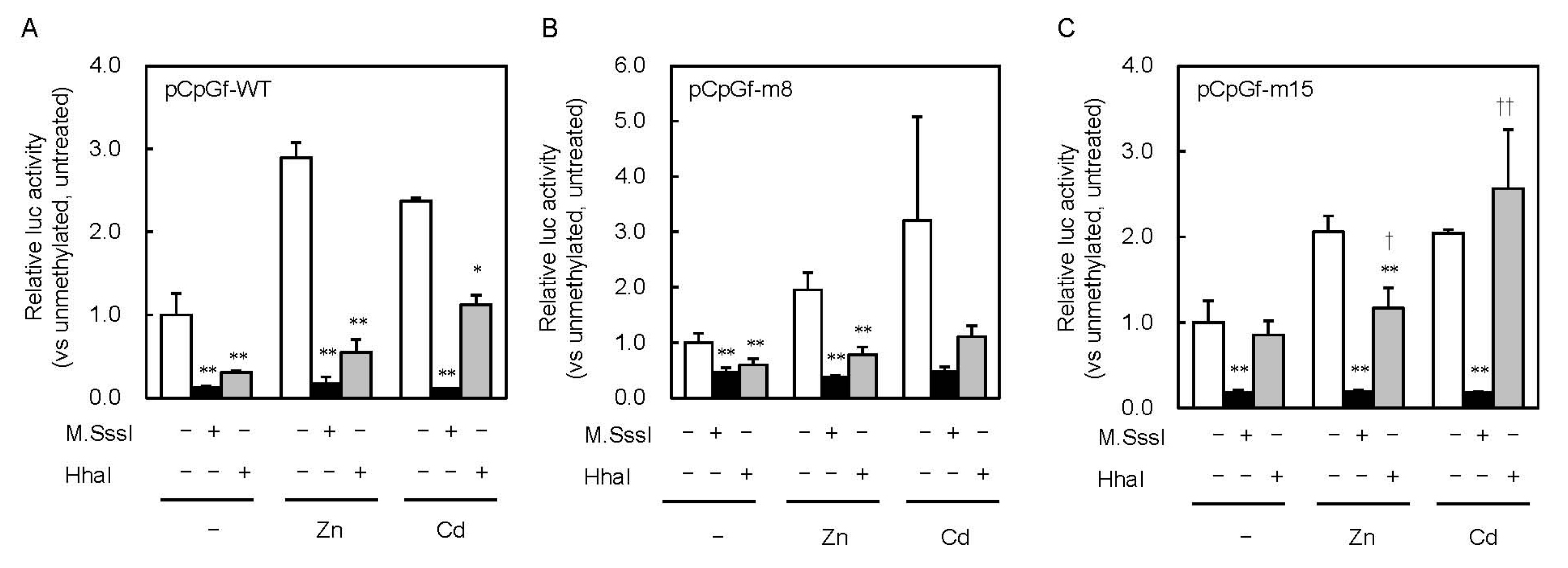{kind=link}
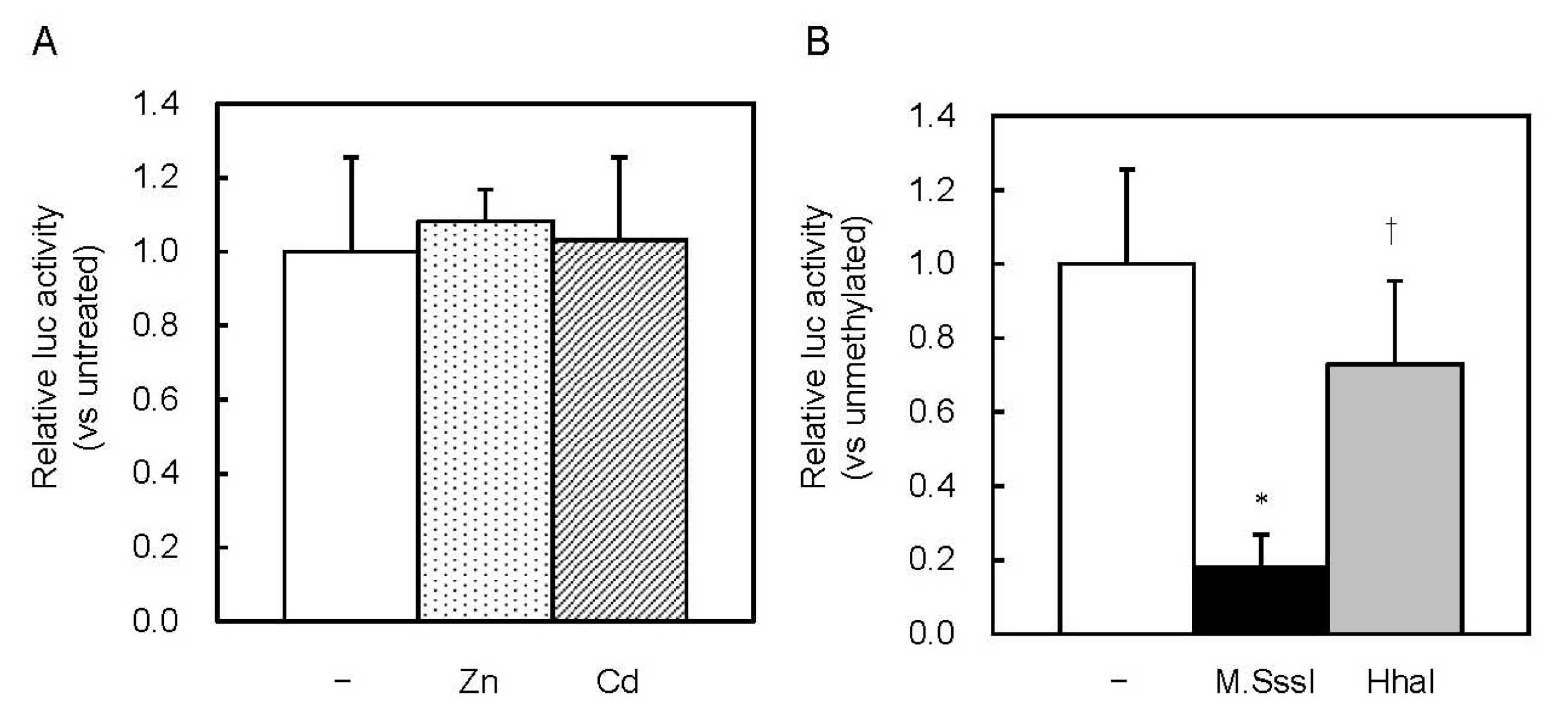{kind=link}
| Gene | Control | Cd | 5-AzaC + Cd |
|---|---|---|---|
| MT1 | 1.0 ± 1.9 | 0.9 ± 0.7 | 31,214.4 ± 16,310.7 * |
| β-actin | 1.0 ± 0.6 | 0.9 ± 0.3 | 0.9 ± 0.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogushi, S.; Yoshida, Y.; Nakanishi, T.; Kimura, T. CpG Site-Specific Regulation of Metallothionein-1 Gene Expression. Int. J. Mol. Sci. 2020, 21, 5946. https://doi.org/10.3390/ijms21175946
Ogushi S, Yoshida Y, Nakanishi T, Kimura T. CpG Site-Specific Regulation of Metallothionein-1 Gene Expression. International Journal of Molecular Sciences. 2020; 21(17):5946. https://doi.org/10.3390/ijms21175946
Chicago/Turabian StyleOgushi, Shoko, Yuya Yoshida, Tsuyoshi Nakanishi, and Tomoki Kimura. 2020. "CpG Site-Specific Regulation of Metallothionein-1 Gene Expression" International Journal of Molecular Sciences 21, no. 17: 5946. https://doi.org/10.3390/ijms21175946
APA StyleOgushi, S., Yoshida, Y., Nakanishi, T., & Kimura, T. (2020). CpG Site-Specific Regulation of Metallothionein-1 Gene Expression. International Journal of Molecular Sciences, 21(17), 5946. https://doi.org/10.3390/ijms21175946