Expression and Role of Biosynthetic, Transporter, Receptor, and Responsive Genes for Auxin Signaling during Clubroot Disease Development

 ,
, 

Abstract
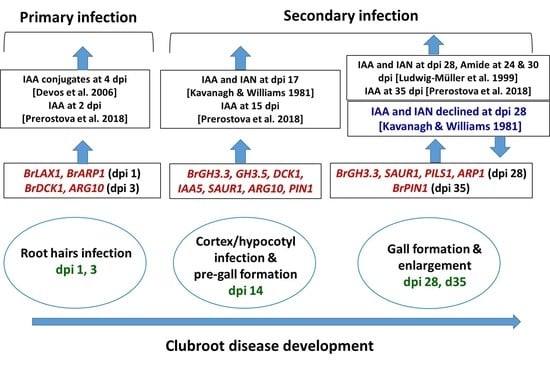
1. Introduction
2. Results
2.1. Properties of Auxin-Related Proteins
2.2. Variation in Expression between Leaves and Roots
2.3. Expression Profiles of Auxin-Responsive GH3 Family and BrDCK1 Genes
2.4. Expression Profiles of BrIAA and BrRACK Genes
2.5. Expression Profiles of BrSAUR and BrNIT Signaling Genes
2.6. Expression Profiles of Other Auxin-Related Genes
3. Discussion
3.1. Expression Level Difference in Leaf vs. Root Tissue
3.2. Expression Level Difference and Role of Auxin Signaling and Biosynthesis (BrGH3, BrIAA, BrNIT, and BrSAUR) Genes
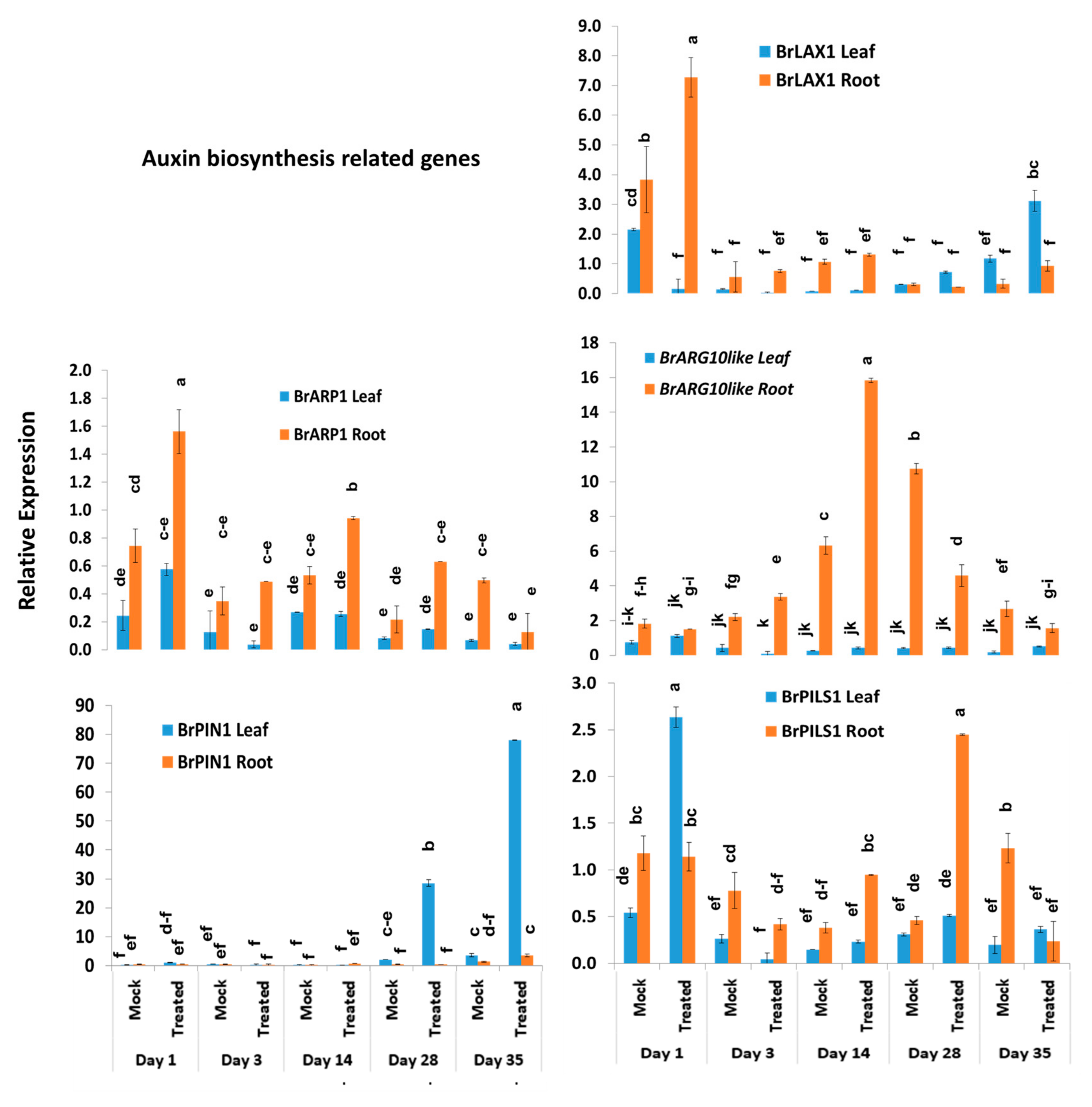
3.3. Expression Level Difference and Role of Auxin Transporter (BrPIN, BrPILS, and BrLAX) Genes
3.4. Expression Level Difference and Role of Auxin Receptor (BrDCK and BrRACK) Genes
3.5. Expression Level Difference and Role of BrARP1 Gene
4. Materials and Methods
4.1. Preparation of Plant Materials
4.2. Gall Collection, Spore Preparation, and Inoculation of Plant Materials
4.3. Sampling of Inoculated Roots and Leaves for Expression Analysis
4.4. Extraction of RNA
4.5. In Silico Analysis of Auxin-Related Genes
4.6. Quantitative RT-PCR for Expression Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ludwig-Müller, J. Auxin homeostasis, signaling, and interaction with other growth hormones during the clubroot disease of Brassicaceae. Plant Signal Behav. 2014, 9, e28593. [Google Scholar] [CrossRef] [PubMed]
- Wallenhammar, A.C. Prevalence of Plasmodiophora brassicae in a spring oilseed rape growing area in central Sweden and factors influencing soil infestation levels. Plant Pathol. 1996, 45, 710–719. [Google Scholar] [CrossRef]
- Devos, S.; Laukens, K.; Deckers, P.; Van Der Straeten, D.; Beeckman, T.; Inzé, D.; Van Onckelen, H.; Witters, E.; Prinsen, E. A hormone and proteome approach to picturing the initial metabolic events during Plasmodiophora brassicae infection on Arabidopsis. Mol. Plant Microbe Interact. 2006, 19, 1431–1443. [Google Scholar] [CrossRef]
- Kageyama, K.; Asano, T. Life cycle of Plasmodiophora brassicae. J. Plant Growth Regul. 2009, 28, 203–211. [Google Scholar] [CrossRef]
- Devos, S.; Vissenberg, K.; Verbelen, J.P.; Prinsen, E. Infection of Chinese cabbage by Plasmodiophora brassicae leads to a stimulation of plant growth: Impacts on cell wall metabolism and hormone balance. New Phytol. 2005, 166, 241–250. [Google Scholar] [CrossRef]
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Ludwig-Müller, J.; Pieper, K.; Ruppel, M.; Cohen, J.D.; Epstein, E.; Kiddle, G.; Bennett, R. Indole glucosinolate and auxin biosynthesis in Arabidopsis thaliana (L.) Heynh. glucosinolate mutants and the development of clubroot disease. Planta 1999, 208, 409–419. [Google Scholar]
- Ludwig-Müller, J.; Prinsen, E.; Rolfe, S.A.; Scholes, J.D. Metabolism and plant hormone action during clubroot disease. J. Plant Growth Regul. 2009, 28, 229–244. [Google Scholar] [CrossRef]
- Robin, A.H.K.; Hossain, M.R.; Kim, H.T.; Nou, I.S.; Park, J.I. Role of Cytokinins in Clubroot Disease Development. Plant Breed. Biotech. 2019, 7, 73–82. [Google Scholar] [CrossRef]
- Dekhuijzen, H.M. The occurrence of free and bound cytokinins in plasmodia of Plasmodiophora brassicae isolated from tissue cultures of clubroots. Plant Cell Rep. 1981, 1, 18–20. [Google Scholar] [CrossRef]
- Müller, P.; Hilgenberg, W. Isomers of zeatin and zeatin riboside in clubroot tissue: Evidence for trans-zeatin biosynthesis by Plasmodiophora brassicae. Physiol. Plant. 1986, 66, 245–250. [Google Scholar] [CrossRef]
- Davies, P.J. The Plant Hormones: Their Nature, Occurrence, and Functions. In Plant Hormones; Springer: Dordrecht, The Netherlands, 2010; pp. 1–15. [Google Scholar]
- Ludwig-Müller, J. Bacteria and fungi controlling plant growth by manipulating auxin: Balance between development and defense. J. Plant Physiol. 2015, 172, 4–12. [Google Scholar] [CrossRef]
- Fu, J.; Wang, S. Insights into auxin signaling in plant–pathogen interactions. Front. Plant Sci. 2011, 2, 74. [Google Scholar] [CrossRef]
- Butcher, D.N.; El-Tigani, S.; Ingram, D.S. The role of indole glucosinolates in the clubroot disease of the Cruciferae. Physiol. Plant Pathol. 1974, 4, 127–140. [Google Scholar] [CrossRef]
- Mousdale, D.M.A. Endogenous indolyl-3-acetic acid and pathogen-induced plant growth disorders: Distinction between hyperplasia and neoplastic development. Experientia 1981, 37, 972–973. [Google Scholar] [CrossRef]
- Neuhaus, K.; Grsic-Rausch, S.; Sauerteig, S.; Ludwig-Müller, J. Arabidopsis plants transformed with nitrilase 1 or 2 in antisense direction are delayed in clubroot development. J. Plant Physiol. 2000, 156, 756–761. [Google Scholar]
- Raa, J. Indole-3-acetic acid levels and the role of indole-3-acetic acid oxidase in the normal root and club-root of cabbage. Physiol. Plant. 1971, 25, 130–134. [Google Scholar] [CrossRef]
- Jahn, L.; Mucha, S.; Bergmann, S.; Horn, C.; Siemens, J.; Staswick, P.; Steffens, B.; Ludwig-Müller, J. The clubroot pathogen (Plasmodiophora brassicae) influences auxin signaling to regulate auxin homeostasis. Plants 2013, 2, 726–749. [Google Scholar] [CrossRef] [PubMed]
- Siemens, J.; Keller, I.; Sarx, J.; Kunz, S.; Schuller, A.; Nagel, W.; Schmülling, T.; Parniske, M.; Ludwig-Müller, J. Transcriptome analysis of Arabidopsis clubroots indicate a key role for cytokinins in disease development. Mol. Plant Microbe Interact. 2006, 19, 480–494. [Google Scholar] [CrossRef]
- Schuller, A.; Kehr, J.; Ludwig-Müller, J. Laser microdissection coupled to transcriptional profiling of Arabidopsis roots inoculated by Plasmodiophora brassicae indicates a role for brassinosteroids in clubroot formation. Plant Cell Physiol. 2014, 55, 392–411. [Google Scholar] [CrossRef]
- Ludwig-Müller, J.; Auer, S.; Jülke, S.; Marschollek, S. Manipulation of auxin and cytokinin balance during the Plasmodiophora brassicae—Arabidopsis thaliana interaction. In Auxins and Cytokinins in Plant Biology; Humana Press: Totowa, NJ, USA, 2017; pp. 41–60. [Google Scholar]
- Grsic-Rausch, S.; Kobelt, P.; Siemens, J.; Bischoff, M.; Ludwig-Müller, J. Expression andlocalization of nitrilase during symptom development of the clubroot disease in Arabidopsis thaliana. Plant Physiol. 2000, 122, 369–378. [Google Scholar] [CrossRef]
- Päsold, S.; Siegel, I.; Seidel, C.; Ludwig-Müller, J. Flavonoid accumulation in Arabidopsis thaliana root galls caused by the obligate biotrophic pathogen Plasmodiophora brassicae. Mol. Plant Pathol. 2010, 11, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Grsic, S.; Kirchheim, B.; Pieper, K.; Fritsch, M.; Hilgenberg, W.; Ludwig-Müller, J. Induction of auxin biosynthetic enzymes by jasmonic acid and in clubroot diseased Chinese cabbage plants. Physiol. Plant. 1999, 105, 521–531. [Google Scholar] [CrossRef]
- Schuller, A.; Ludwig-Müller, J. A family of auxin conjugate hydrolases from Brassica rapa: Characterization and expression during clubroot disease. New Phytol. 2006, 171, 145–158. [Google Scholar] [CrossRef]
- Agarwal, A.; Kaul, V.; Faggian, R.; Rookes, J.E.; Ludwig-Müller, J.; Cahill, D.M. Analysis of global host gene expression during the primary phase of the Arabidopsis thaliana–Plasmodiophora brassicae interaction. Funct. Plant Biol. 2011, 38, 462–478. [Google Scholar] [CrossRef]
- Jia, H.; Wei, X.; Yang, Y.; Yuan, Y.; Wei, F.; Zhao, Y.; Yang, S.; Yao, Q.; Wang, Z.; Tian, B.; et al. Root RNA-seq analysis reveals a distinct transcriptome landscape between clubroot-susceptible and clubroot-resistant Chinese cabbage lines after Plasmodiophora brassicae infection. Plant Soil. 2017, 421, 93–105. [Google Scholar] [CrossRef]
- Laila, R.; Robin, A.H.K.; Park, J.-I.; Saha, G.; Kim, H.-T.; Kayum, M.A.; Nou, I.-S. Expression and Role of Response Regulating, Biosynthetic and Degrading Genes for Cytokinin Signaling during Clubroot Disease Development. Int. J. Mol. Sci. 2020, 21, 3896. [Google Scholar] [CrossRef]
- Park, B.S.; Sang, W.G.; Song, J.T.; Lee, B.H.; Kim, J.H.; Seo, H.S. Auxin is involved in the regulation of leaf and root development by LAF1 under short day conditions. Biol. Plant. 2011, 55, 647. [Google Scholar] [CrossRef]
- Ruzza, V.; Sessa, G.; Sassi, M.; Morelli, G.; Ruberti, I. Auxin coordinates shoot and root development during shade avoidance response. In Auxin and its role in plant development; Zažímalová, E., Petrášek, J., Benková, E., Eds.; Springer: Vienna, Austria, 2014; pp. 389–412. [Google Scholar]
- Aloni, R.; Aloni, E.; Langhans, M.; Ullrich, C.I. Role of cytokinin and auxin in shaping root architecture: Regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann. Bot. 2006, 97, 883–893. [Google Scholar] [CrossRef]
- Overvoorde, P.; Fukaki, H.; Beeckman, T. Auxin control of root development. Cold Spring Harb. Perspect. Biol. 2010, 2, a001537. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J. Auxin and plant-microbe interactions Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar]
- Hagen, G.; Guilfoyle, T. Auxin-responsive gene expression: Genes, promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Berleth, T.; Krogan, N.T.; Scarpella, E. Auxin signals—turning genes on and turning cells around. Curr. Opin. Plant Biol. 2004, 7, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Ludwig-Müller, J. Auxin conjugates: Their role for plant development and in the evolution of land plants. J. Exp. Bot. 2011, 62, 1757–1773. [Google Scholar] [CrossRef]
- De Rybel, B.; Vassileva, V.; Parizot, B.; Demeulenaere, M.; Grunewald, W.; Audenaert, D.; van Camenhout, J.; Overvoorde, P.; Jansen, L.; Vanneste, S.; et al. A novel Aux/IAA28 signaling cascade activates GATA23-dependent specification of lateral root founder cell identity. Curr. Biol. 2010, 20, 1697–1706. [Google Scholar] [CrossRef]
- Bartling, D.; Seedorf, M.; Schmidt, R.C.; Weiler, E.W. Molecular characterization of two cloned nitrilases from Arabidopsis thaliana: Key enzymes in biosynthesis of the plant hormone indole-3-acetic acid. Proc. Natl. Acad. Sci. USA 1994, 91, 6021–6025. [Google Scholar] [CrossRef]
- Bartel, B.; Fink, G.R. Differential regulation of an auxin-producing nitrilase gene family in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1994, 91, 6649–6653. [Google Scholar] [CrossRef]
- Müller, A.; Hillebrand, H.; Weiler, E.W. Indole-3-acetic acid is synthesized from L-tryptophan in roots of Arabidopsis thaliana. Planta 1998, 206, 362–369. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Konradyova, V.; Knirsch, V.; Gaudinova, A.; Kramna, B.; Kazda, J.; Ludwig-Müller, J.; Vankova, R. Hormonal responses to Plasmodiophora brassicae infection in Brassica napus cultivars differing in their pathogen resistance. Int. J. Mol. Sci. 2018, 19, 4024. [Google Scholar] [CrossRef]
- Park, J.E.; Kim, Y.S.; Yoon, H.K.; Park, C.M. Functional characterization of a small auxin-up RNA gene in apical hook development in Arabidopsis. Plant Sci. 2007, 172, 150–157. [Google Scholar] [CrossRef]
- Markakis, M.N.; Boron, A.K.; Van Loock, B.; Saini, K.; Cirera, S.; Verbelen, J.P.; Vissenberg, K. Characterization of a small auxin-up RNA (SAUR)-like gene involved in Arabidopsis thaliana development. PLoS ONE 2013, 8, e82596. [Google Scholar] [CrossRef]
- Hou, K.; Wu, W.; Gan, S.S. SAUR36, a small auxin up RNA gene, is involved in the promotion of leaf senescence in Arabidopsis. Plant Physiol. 2013, 161, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Irani, S.; Trost, B.; Waldner, M.; Nayidu, N.; Tu, J.; Kusalik, A.J.; Todd, C.D.; Wei, Y.; Bonham-Smith, P.C. Transcriptome analysis of response to Plasmodiophora brassicae infection in the Arabidopsis shoot and root. BMC Genom. 2018, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Ciaghi, S.; Schwelm, A.; Neuhauser, S. Transcriptomic response in symptomless roots of clubroot infected kohlrabi (Brassica oleracea var. gongylodes) mirrors resistant plants. BMC Plant Biol. 2019, 19, 288. [Google Scholar] [CrossRef] [PubMed]
- Friml, J. Subcellular trafficking of PIN auxin efflux carriers in auxin transport. Eur. J. Cell Biol. 2010, 89, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: Action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Swarup, R.; Péret, B. AUX/LAX family of auxin influx carriers—an overview. Front. Plant Sci. 2012, 3, 225. [Google Scholar] [CrossRef]
- Li, D.H.; Shen, F.J.; Li, H.Y.; Li, W. Kale BoRACK1 is involved in the plant response to salt stress and Peronospora brassicae Gaumann. J. Plant Physiol. 2017, 213, 188–198. [Google Scholar] [CrossRef]
- Chen, J.G.; Ullah, H.; Temple, B.; Liang, J.; Guo, J.; Alonso, J.M.; Ecker, J.R.; Jones, A.M. RACK1 mediates multiple hormone responsiveness and developmental processes in Arabidopsis. J. Exp. Bot. 2006, 57, 2697–2708. [Google Scholar] [CrossRef]
- Cai, Z.; Zeng, D.E.; Liao, J.; Cheng, C.; Sahito, Z.A.; Xiang, M.; Fu, M.; Chen, Y.; Wang, D. Genome-wide analysis of auxin receptor family genes in Brassica juncea var. tumida. Genes 2019, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Tatematsu, K.; Ward, S.; Leyser, O.; Kamiya, Y.; Nambara, E. Identification of cis-Elements That Regulate Gene Expression during Initiation of Axillary Bud Outgrowth in Arabidopsis. Plant Physiol. 2005, 138, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Mantri, N.L.; Ford, R.; Coram, T.E.; Pang, E.C.K. Transcriptional Profiling of Chickpea Genes Differentially Regulated in Response to High-Salinity, Cold and Drought. BMC Genom. 2007, 8, 303. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Han, C.-T.; Hur, Y. Molecular Characterization of the Brassica rapa Auxin-Repressed, Superfamily Genes, BrARP1 and BrDRM1. Mol. Biol. Rep. 2013, 40, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.W.; Kim, K.A.; Park, S.C.; Jeong, M.J.; Byun, M.O.; Kwon, H.B. Expression Profiles of Hot Pepper (Capsicum annum) Genes under Cold Stress Condition. J. Biosci. 2005, 30, 657–667. [Google Scholar] [CrossRef]
- Wu, L.; Yu, M.; Holowachuk, J.; Sharpe, A.; Lydiate, D.; Hegedus, D.; Gruber, M. Evaluation of a Brassica napus auxin-repressed gene induced by flea beetle damage and Sclerotinia sclerotiorum infection. Am. J. Plant Sci. 2017, 8, 1921–1952. [Google Scholar] [CrossRef][Green Version]
- Kavanagh, J.A.; Williams, P.H. Indole auxins in Plasmodiophora infected cabbage roots and hypocotyls. Trans. Br. Mycol. Soc. 1981, 77, 125–129. [Google Scholar] [CrossRef]
- Feng, J.; Hwang, S.F.; Strelkov, S.E. Studies into primary and secondary infection processes by Plasmodiophora brassicae on canola. Plant Path. 2013, 62, 177–183. [Google Scholar] [CrossRef]
- Cheng, F.; Liu, S.; Wu, J.; Fang, L.; Sun, S.; Liu, B.; Li, P.; Hua, W.; Wang, X. BRAD, the genetics and genomics database for Brassica plants. BMC Plant Biol. 2011, 11, 136. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy—the proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Robin, A.H.K.; Yi, G.E.; Laila, R.; Yang, K.; Park, J.I.; Kim, H.R.; Nou, I.S. Expression profiling of glucosinolate biosynthetic genes in Brassica oleracea L. var. capitata inbred lines reveals their association with glucosinolate content. Molecules 2016, 21, 787. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | BRAD ID | Chromosomal Location | Start Codon | Stop Codon | St. | Gene Identity | (aa) | Pi | MW (kDa) | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| BrDCK1 | Bra008038 | A02 | 11763540 | 11765539 | − | Receptor kinase | 447 | 9.06 | 50.44 | Cell membrane |
| BrDCK1.1 | Bra016050 | A07 | 22960115 | 22961810 | − | Protein kinase | 451 | 9.24 | 50.77 | Cell membrane |
| BrDCK1.2 | Bra003875 | A07 | 18779483 | 18780917 | + | 396 | 8.74 | 44.61 | Cell membrane | |
| BrLAX1 | Bra008325 | A02 | 13732347 | 13734803 | − | Auxin efflux carrier/ Transporter | 472 | 8.95 | 53.53 | Cell membrane |
| BrLAX1.1 | Bra003674 | A07 | 17668825 | 17671143 | + | 471 | 8.86 | 53.45 | Cell membrane | |
| BrPIN1 | Bra015983 | A07 | 23234264 | 23237329 | + | 618 | 9.14 | 66.82 | Cell membrane | |
| BrPIN1.1 | Bra008105 | A02 | 12154076 | 12156932 | + | 616 | 9.10 | 66.61 | Cell membrane | |
| BrPILS1 | Bra014265 | A08 | 2101045 | 2103020 | − | 431 | 9.26 | 46.66 | Lysosome | |
| BrPILS1.1 | Bra030441 | A05 | 11329601 | 11331470 | + | 423 | 8.86 | 46.08 | Cell membrane | |
| BrRACK1 | Bra032261 | A05 | 12304734 | 12306049 | + | Receptor C kinase | 327 | 6.31 | 35.70 | Nucleus |
| BrRACK1.1 | Bra018745 | A06 | 2234607 | 2235718 | + | 298 | 6.06 | 32.47 | Nucleus | |
| BrRACK1.2 | Bra022282 | A05 | 19371512 | 19372720 | + | 325 | 7.59 | 35.46 | Nucleus | |
| BrRACK1.3 | Bra021294 | A01 | 21899789 | 21900926 | + | 326 | 8.33 | 35.58 | Nucleus | |
| BrGH3.1 | Bra039186 | A09 | 33153634 | 33155891 | + | Auxin-responsive GH3 (auxin signaling) | 596 | 6.28 | 67.77 | Cytoplasm |
| BrGH3.2 | Bra008833 | A10 | 12836737 | 12839205 | − | 594 | 7.81 | 67.08 | Cytoplasm | |
| BrGH3.3 | Bra041046 | Scaffold000414 | 2738 | 4832 | − | 587 | 5.98 | 66.03 | Cytoplasm | |
| BrGH3.31 | Bra023405 | A02 | 2218215 | 2221058 | + | 595 | 5.80 | 67.02 | Cytoplasm | |
| BrGH3.4 | Bra034205 | A09 | 1271960 | 1274617 | − | 610 | 6.28 | 68.54 | Cytoplasm | |
| BrGH3.41 | Bra013086 | A03 | 20567114 | 20569305 | − | 557 | 5.43 | 62.48 | Cytoplasm | |
| BrGH3.5 | Bra019060 | A03 | 26541024 | 26543324 | + | 612 | 5.98 | 69.39 | Cytoplasm | |
| BrGH3.51 | Bra026365 | A01 | 9560154 | 9562651 | + | 612 | 6.05 | 69.36 | Cytoplasm | |
| BrGH3.52 | Bra022725 | A02 | 6752640 | 6754845 | + | 612 | 5.87 | 69.09 | Cytoplasm | |
| BrGH3.53 | Bra029023 | A03 | 5931112 | 5933213 | + | 608 | 5.74 | 68.63 | Cytoplasm | |
| BrIAA1 | Bra039855 | A08 | 6303677 | 6305559 | − | Auxin signaling | 233 | 8.68 | 25.85 | Nucleus |
| BrIAA2 | Bra033886 | A05 | 15764615 | 15766543 | − | 242 | 8.78 | 26.43 | Nucleus | |
| BrIAA2.1 | Bra001900 | A03 | 19263301 | 19265148 | + | 237 | 8.54 | 26.16 | Nucleus | |
| BrIAA3 | Bra001634 | A03 | 17624668 | 17625966 | − | Phytochrome -associated | 272 | 7.68 | 30.43 | Nucleus |
| BrIAA3.1 | Bra021184 | A01 | 22676134 | 22677514 | + | 269 | 9.20 | 30.28 | Nucleus | |
| BrIAA4 | Bra009867 | A06 | 18399781 | 18400911 | + | Auxin signaling | 176 | 8.80 | 20.33 | Nucleus |
| BrIAA4.1 | Bra036557 | A09 | 2815416 | 2816543 | − | 175 | 9.24 | 20.00 | Cytoplasm | |
| BrIAA5 | Bra032521 | A09 | 38191234 | 38191967 | + | Transcriptional regulator | 180 | 5.11 | 20.36 | Cytoplasm |
| BrIAA5.1 | Bra015297 | A10 | 2137104 | 2137826 | + | 186 | 5.31 | 21.10 | Cytoplasm | |
| BrSAUR1 | Bra011560 | A01 | 1680948 | 1681271 | + | Auxin-responsive protein (auxin signaling) | 107 | 6.82 | 12.11 | Mitochondrion |
| BrSAUR1.1 | Bra034651 | A08 | 11488514 | 11488828 | + | 104 | 8.60 | 12.03 | Mitochondrion | |
| BrSAUR1.2 | Bra017676 | A03 | 29740606 | 29740929 | − | 107 | 6.82 | 12.14 | Mitochondrion | |
| BrSAUR1.3 | Bra013061 | A03 | 20715121 | 20715447 | − | 108 | 7.73 | 12.36 | Mitochondrion | |
| BrSAUR2 | Bra039345 | Scaffold000164 | 28744 | 29145 | − | 133 | 9.03 | 15.34 | Mitochondrion | |
| BrSAUR3 | Bra039264 | A04 | 18962112 | 18962471 | + | 119 | 7.81 | 13.67 | Cell membrane | |
| BrSAUR3.1 | Bra004515 | A05 | 560573 | 560941 | − | 122 | 8.79 | 14.06 | Cell membrane | |
| BrSAUR4 | Bra014476 | A04 | 882244 | 882756 | − | 170 | 10.96 | 19.75 | Mitochondrion | |
| BrNIT1 | Bra028932 | A03 | 5467727 | 5469454 | − | Auxin biosynthesis | 343 | 5.54 | 37.73 | Cytoplasm |
| BrNIT1.1 | Bra021681 | A04 | 13934901 | 13936735 | + | 350 | 5.48 | 38.48 | Cytoplasm | |
| BrNIT2 | Bra020184 | A02 | 4417472 | 4419242 | + | 357 | 5.30 | 38.94 | Cytoplasm | |
| BrARG10-like | Bra026348 | A01 | 9659581 | 9660796 | − | Auxin responsive | 250 | 5.52 | 27.55 | Cytoplasm |
| BrARG10-like1 | Bra010404 | A08 | 13579603 | 13580693 | + | 250 | 6.07 | 27.49 | Cytoplasm | |
| BrARP1 | Bra013191 | A03 | 19982046 | 19983742 | + | Auxin repressed protein | 403 | 9.70 | 43.86 | Cell membrane |
| Sources of Variation | df | Genes | MS | F Statistic | p Value | Genes | MS | F Statistic | p Value | Genes | MS | F Statistic | p Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time-point | 4 | BrGH3.1 | 2.68 | 83.2 | < 0.01 | BrIAA4 | 1.01 | 63.4 | < 0.01 | BrDCK1 | 132.7 | 76.3 | < 0.01 |
| Treatment | 1 | 0.94 | 29.2 | < 0.01 | 1.28 | 80.2 | < 0.01 | 22.1 | 12.7 | < 0.01 | |||
| Tm × Tr | 4 | 1.34 | 41.8 | < 0.01 | 1.35 | 84.5 | < 0.01 | 28.5 | 16.4 | < 0.01 | |||
| LvR (Tm Tr) | 10 | 1.21 | 37.7 | < 0.01 | 2.14 | 134.3 | < 0.01 | 265 | 152.9 | < 0.01 | |||
| Error | 40 | 0.032 | 0.016 | 1.74 | |||||||||
| Time-point | 4 | BrGH3.2 | 1519 | 19.8 | < 0.01 | BrIAA5 | 15.5 | 71.7 | < 0.01 | BrRACK1 | 11.67 | 69.7 | < 0.01 |
| Treatment | 1 | 2169 | 28.2 | < 0.01 | 1.14 | 5.28 | 0.027 | 0.116 | 0.70 | 0.409 | |||
| Tm × Tr | 4 | 1184 | 15.4 | < 0.01 | 16.7 | 77.3 | < 0.01 | 1.11 | 6.66 | < 0.01 | |||
| LvR (Tm Tr) | 10 | 1408 | 18.3 | < 0.01 | 14.2 | 65.9 | < 0.01 | 8.06 | 48.2 | < 0.01 | |||
| Error | 40 | 76.8 | 0.22 | 0.167 | |||||||||
| Time-point | 4 | BrGH3.3 | 4,699,114 | 2767 | < 0.01 | BrSAUR1 | 0.268 | 406.6 | < 0.01 | BrLAX1 | 14.2 | 128.2 | < 0.01 |
| Treatment | 1 | 1,822,177 | 1073 | < 0.01 | 0.345 | 522.1 | < 0.01 | 3.52 | 31.7 | < 0.01 | |||
| Tm × Tr | 4 | 4,623,051 | 2723 | < 0.01 | 0.533 | 805.8 | < 0.01 | 0.80 | 7.24 | < 0.01 | |||
| LvR (Tm Tr) | 10 | 6,320,827 | 3723 | < 0.01 | 0.505 | 765.3 | < 0.01 | 9.12 | 82.2 | < 0.01 | |||
| Error | 40 | 1698 | 0.0007 | 0.11 | |||||||||
| Time-point | 4 | BrGH3.4 | 1978.9 | 582.8 | < 0.01 | BrSAUR2 | 3.56 | 46.24 | < 0.01 | BrARP1 | 0.726 | 158 | < 0.01 |
| Treatment | 1 | 54.83 | 17.0 | < 0.01 | 3.63 | 47.22 | < 0.01 | 0.422 | 91.8 | < 0.01 | |||
| Tm × Tr | 4 | 161.8 | 50.2 | < 0.01 | 2.93 | 38.03 | < 0.01 | 0.243 | 52.9 | < 0.01 | |||
| LvR (Tm Tr) | 10 | 1369.7 | 424.8 | < 0.01 | 5.61 | 72.92 | < 0.01 | 0.368 | 80.1 | < 0.01 | |||
| Error | 40 | 3.22 | 0.077 | 0.004 | |||||||||
| Time-point | 4 | BrGH3.5 | 396.0 | 72.82 | < 0.01 | BrSAUR3 | 4.84 | 355.6 | < 0.01 | BrARG | 52.95 | 741.5 | < 0.01 |
| Treatment | 1 | 71.8 | 13.21 | < 0.01 | 3.37 | 247.5 | < 0.01 | 2.62 | 36.6 | < 0.01 | |||
| Tm × Tr | 4 | 215.8 | 39.69 | < 0.01 | 2.06 | 151.0 | < 0.01 | 26.39 | 369.6 | < 0.01 | |||
| LvR (Tm Tr) | 10 | 357.9 | 65.82 | < 0.01 | 2.96 | 218.0 | < 0.01 | 65.54 | 917.8 | < 0.01 | |||
| Error | 40 | 5.44 | 0.013 | 0.071 | |||||||||
| Time-point | 4 | BrIAA1 | 200.9 | 192.5 | < 0.01 | BrSAUR4 | 0.518 | 121.1 | < 0.01 | BrPIN1 | 260.1 | 13.2 | < 0.01 |
| Treatment | 1 | 76.18 | 73.0 | < 0.01 | 0.057 | 13.44 | < 0.01 | 298.8 | 15.1 | < 0.01 | |||
| Tm × Tr | 4 | 213.8 | 204.8 | < 0.01 | 0.121 | 28.35 | < 0.01 | 168.5 | 8.54 | < 0.01 | |||
| LvR (Tm Tr) | 10 | 266.2 | 255.1 | < 0.01 | 0.857 | 200.5 | < 0.01 | 206.0 | 10.4 | < 0.01 | |||
| Error | 40 | 1.04 | 0.004 | 19.7 | |||||||||
| Time-point | 4 | BrIAA2 | 0.017 | 12.15 | < 0.01 | BrNIT1 | 14844 | 165.8 | < 0.01 | BrPILS1 | 0.969 | 233.9 | < 0.01 |
| Treatment | 1 | 0.013 | 9.57 | < 0.01 | 9378 | 104.8 | < 0.01 | 0.789 | 190.5 | < 0.01 | |||
| Tm × Tr | 4 | 0.173 | 120.9 | < 0.01 | 6064 | 67.7 | < 0.01 | 0.695 | 167.6 | < 0.01 | |||
| LvR (Tm Tr) | 10 | 0.129 | 90.6 | < 0.01 | 15511 | 173.3 | < 0.01 | 0.584 | 140.9 | < 0.01 | |||
| Error | 40 | 0.0014 | 89.5 | 0.004 | |||||||||
| Time-point | 4 | BrIAA3 | 47.3 | 632.4 | < 0.01 | BrNIT2 | 1.55 | 22.95 | < 0.01 | ||||
| Treatment | 1 | 8.28 | 110.7 | < 0.01 | 0.85 | 12.67 | < 0.01 | ||||||
| Tm × Tr | 4 | 8.16 | 109.2 | < 0.01 | 1.05 | 15.52 | < 0.01 | ||||||
| LvR (Tm Tr) | 10 | 40.1 | 535.5 | < 0.01 | 2.78 | 41.08 | < 0.01 | ||||||
| Error | 40 | 0.075 | 0.067 |
| Sample ID | Time Point | Treatment | Sample Type |
|---|---|---|---|
| 1 | Day 1 | Mock | Leaf |
| 2 | Day 1 | Mock | Root |
| 3 | Day 1 | Seosan-inoculated | Leaf |
| 4 | Day 1 | Seosan-inoculated | Root |
| 5 | Day 3 | Mock | Leaf |
| 6 | Day 3 | Mock | Root |
| 7 | Day 3 | Seosan-inoculated | Leaf |
| 8 | Day 3 | Seosan-inoculated | Root |
| 9 | Day 14 | Mock | Leaf |
| 10 | Day 14 | Mock | Root |
| 11 | Day 14 | Seosan-inoculated | Leaf |
| 12 | Day 14 | Seosan-inoculated | Root |
| 13 | Day 28 | Mock | Leaf |
| 14 | Day 28 | Mock | Root |
| 15 | Day 28 | Seosan-inoculated | Leaf |
| 16 | Day 28 | Seosan-inoculated | Root |
| 17 | Day 35 | Mock | Leaf |
| 18 | Day 35 | Mock | Root |
| 19 | Day 35 | Seosan-inoculated | Leaf |
| 20 | Day 35 | Seosan-inoculated | Root |
| Arabidopsis Homolog (Accession Number) | Gene Name | BRAD ID | cDNA Size (bp) | Primer Forward (F) and Reverse(R) | Product Size (bp) |
|---|---|---|---|---|---|
| At2g23170 | BrGH3.1 | Bra039186 | 1791 | AAGGACTTGAAAGCTCTCAG | 156 |
| CTTAAATGTTTTCCGGTCGG | |||||
| At1g72540 | BrDCK1 | Bra008038 | 1344 | ATCCAGACAAACCTCTGTTC | 177 |
| AATCCTCCTTCACCGAGATA | |||||
| At5g13360 | BrGH3.2 | Bra008833 | 1758 | AGTGAGACAGCCTCTAGAAT | 177 |
| TCGTTGTATAAGCCCACAAA | |||||
| At4g34760 | BrSAUR1 | Bra011560 | 324 | TTCCACTCGATGTACCAAAG | 197 |
| TGAGGTTAGGGTCTGAAAGA | |||||
| At1g77690 | BrLAX1 | Bra008325 | 1419 | CTGTGGAGATAATGCATGCCA | 161 |
| GAGTAGAGAGAGTGCATTGG | |||||
| At1g73590 | BrPIN1 | Bra015983 | 1857 | GGAAACAGAAAAGCAGCATT | 187 |
| GAGTAGAGAGAGTGCATTGG | |||||
| At1g48630 | BrRACK1 | Bra032261 | 984 | ACATGATCGTCACTTCTTCC | 243 |
| AGAGAACGTCTTTCGTGTGT | |||||
| At3g12830 | BrSAUR2 | Bra039345 | 402 | GAATCTTTCCTCCGGTCTTC | 187 |
| TTTGCTCGTAACCATACTCC | |||||
| At3g44300 | BrNIT1 | Bra028932 | 1032 | GAATCAGAGGGTCTCATCAC | 151 |
| TCGAAACGAATGTAACTGGT | |||||
| At5g22300 | BrNIT2 | Bra020184 | 1074 | GCAAGTATCTTGCTTCTGCT | 157 |
| TGACCATGTGAGTCAAAGAA | |||||
| At5g01990 | BrPILS1 | Bra014265 | 1296 | GAAGCAAATCATCCAACCTG | 167 |
| GTGCCAGTAATATACAGGGG | |||||
| At1g59500 | BrGH3.3 | Bra041046 | 1764 | TAAGCTTCCTCCAGAACAAC | 157 |
| GAGCAAACCTCTACCAAGAA | |||||
| At4g14550 | BrIAA1 | Bra039855 | 702 | CTCTACCAAAGAAGGATCCG | 163 |
| TTCGCTACACTTCTGATGAG | |||||
| At4g03400 | BrGH3.4 | Bra034205 | 1833 | ATGTTGAAGAGATGGACGAC | 208 |
| CTATGGCAAGTAAACGGAAC | |||||
| At3g23050 | BrIAA2 | Bra033886 | 729 | AGCTGCATTGGTTAAGGTAT | 184 |
| CAGATCCATTAGCTTGCTCT | |||||
| At4g27260 | BrGH3.5 | Bra019060 | 1839 | AAGAACGTGATGCCTGTTAT | 183 |
| TGATCTCCGGTCTAGTTCTT | |||||
| At4g27450 | BrARG10-like | Bra026348 | 753 | CTCTGACAACACTTTCTCCA | 175 |
| GGTCAGACCATATTGCTTG | |||||
| At3g16500 | BrIAA3 | Bra001634 | 819 | GAGGTCAAGTCAACAAGAGT | 171 |
| CTCTGAAGAGTTGGTCTACG | |||||
| At2g04850 | BrARP1 | Bra013191 | 1212 | GAATGTGTTCCAAGGGTTTG | 189 |
| CTGCTGCATCTATCATCAGT | |||||
| At2g46690 | BrSAUR3 | Bra039264 | 360 | GCTCTGAAAGGCTTCCATTTAC | 222 |
| AACGTCGCATGGAATTGTGATG | |||||
| At5g25890 | BrIAA4 | Bra009867 | 531 | TTCGTCGGATCTAAACAGAC | 167 |
| CCTTGCAGTTCTTTCTAGGT | |||||
| At1g04240 | BrIAA5 | Bra032521 | 543 | ATCCCCTCCTAGAAAGACTC | 152 |
| TTTTCCTCAAGTATGGTGCA | |||||
| At3g60690 | BrSAUR4 | Bra014476 | 513 | ATACCAATTGGGAAGGAACC | 172 |
| CAAACTCTTCCTCTGCTTCT | |||||
| XM_009147610.2 | ACTIN1 | 1371 | F: CAACCAATCGTCTGTGACAA | 105 | |
| R: ATGTCTTGGCCTACCAACAA | |||||
| FJ969844.1 | ACTIN2 | 491 | F: AATGGTACCGGAATGGTCAA | 119 | |
| R: TCCTTCTGGTTCATCCCAAC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robin, A.H.K.; Saha, G.; Laila, R.; Park, J.-I.; Kim, H.-T.; Nou, I.-S. Expression and Role of Biosynthetic, Transporter, Receptor, and Responsive Genes for Auxin Signaling during Clubroot Disease Development. Int. J. Mol. Sci. 2020, 21, 5554. https://doi.org/10.3390/ijms21155554
Robin AHK, Saha G, Laila R, Park J-I, Kim H-T, Nou I-S. Expression and Role of Biosynthetic, Transporter, Receptor, and Responsive Genes for Auxin Signaling during Clubroot Disease Development. International Journal of Molecular Sciences. 2020; 21(15):5554. https://doi.org/10.3390/ijms21155554
Chicago/Turabian StyleRobin, Arif Hasan Khan, Gopal Saha, Rawnak Laila, Jong-In Park, Hoy-Taek Kim, and Ill-Sup Nou. 2020. "Expression and Role of Biosynthetic, Transporter, Receptor, and Responsive Genes for Auxin Signaling during Clubroot Disease Development" International Journal of Molecular Sciences 21, no. 15: 5554. https://doi.org/10.3390/ijms21155554
APA StyleRobin, A. H. K., Saha, G., Laila, R., Park, J.-I., Kim, H.-T., & Nou, I.-S. (2020). Expression and Role of Biosynthetic, Transporter, Receptor, and Responsive Genes for Auxin Signaling during Clubroot Disease Development. International Journal of Molecular Sciences, 21(15), 5554. https://doi.org/10.3390/ijms21155554