Platelets Extracellular Vesicles as Regulators of Cancer Progression—An Updated Perspective


Abstract
1. Introduction
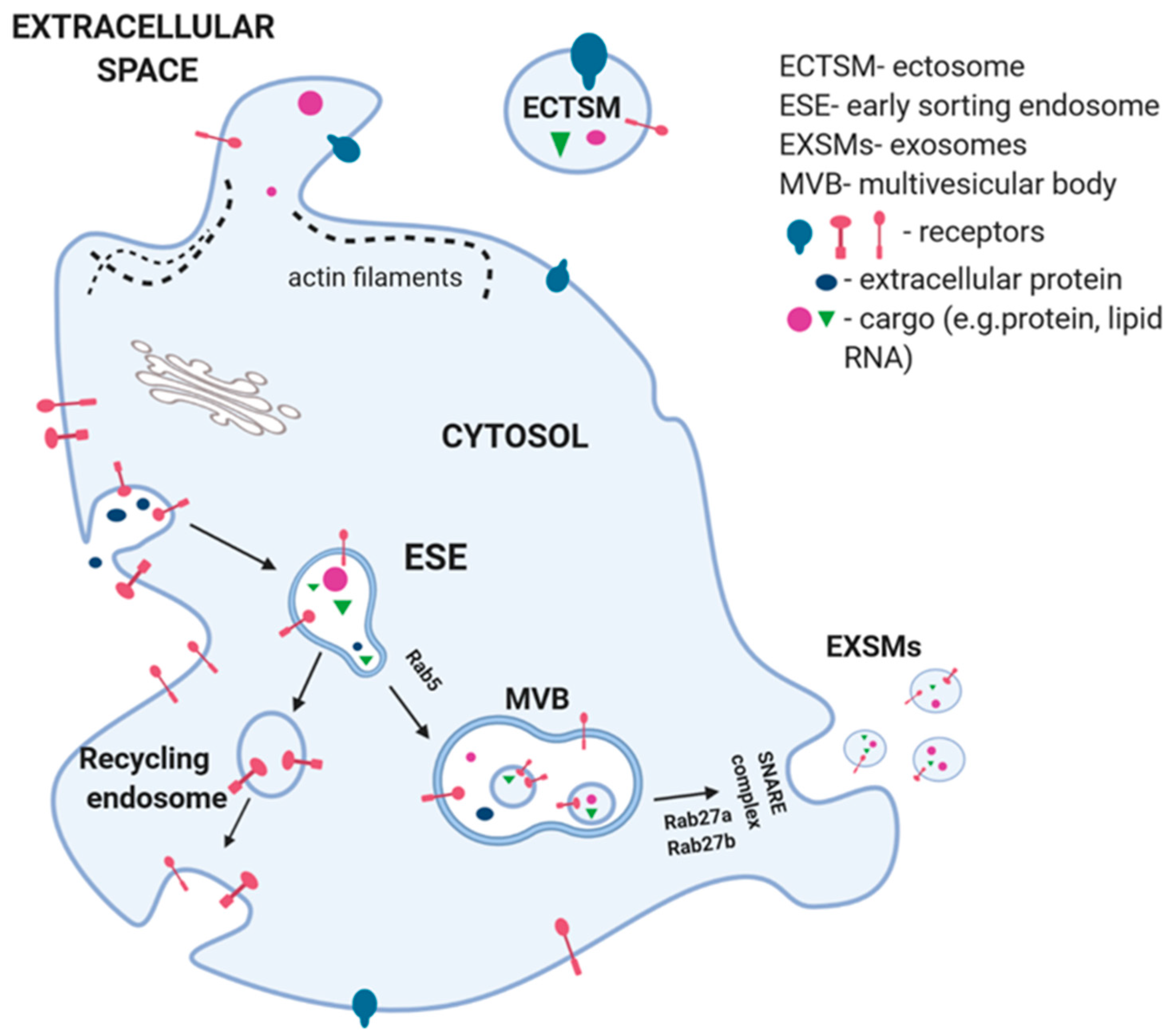
2. EVs Biogenesis and Elimination
2.1. ECTSMs Formation
2.2. EXSMs Formation
2.3. EVs Elimination and Impact of Storage Conditions on PEVs Number
3. Content of Platelet Extracellular Vesicles
4. PEVs in Cancer Progression
4.1. PEVs in Tumor Angiogenesis
4.2. PEVs in Migration, Invasion, and Metastasis
5. The Potential of PEVs as Diagnostics Cancer Biomarkers
6. The Potential of PEVs in Cancer Therapy
7. Summary
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABs | apoptotic bodies |
| ALL | acute lymphoblastic leukemia |
| CD | cluster of differentiation |
| CCL | cancer cell line |
| CLEC-2 | C-type lectin domain family 1-member B |
| ECTSMs | ectosomes |
| ESCRT | endosomal sorting complex required for transport |
| ESE | early sorting endosomes |
| EVs | extracellular vesicles |
| EXSMs | exosomes |
| HAECs | human aortic endothelial cells |
| HUVECs | human umbilical vein endothelial cells |
| ILVs | intraluminal vesicles |
| LA | lactadherin |
| LAMP-1 | lysosome-associated glycoprotein-1 |
| LPS | lipopolysaccharide |
| LXA4 | lipoxin A4 |
| MAPK | mitogen-activated protein kinase |
| Mk-EVs | EVs derived from megakaryocytes |
| MMP | metalloproteinase |
| MVBs | multivesicular bodies |
| MPs | microparticles |
| NSCLC | non-small cell lung cancer |
| OSCC | oral squamous cell carcinoma |
| PBMCs | peripheral blood mononuclear cells |
| PdEXSMs | platelet derived exosomes |
| PEVs | platelets extracellular vesicles |
| PLTs | platelets |
| PMPs | platelets microparticles |
| PUFAs | polyunsaturated fatty acids |
| RT | room temperature |
| TEVs | tumor derived extracellular vesicles |
| TF | tissue factor |
| TGF-β1 | transforming growth factor beta 1 |
| THBS-1 | thrombospondin-1 |
| TLR-4 | toll-like receptor 4 |
| TME | tumor microenvironment |
| TPM3 | tropomyosin 3 |
| VEGF | vascular endothelial growth factor |
| vWf | von Willebrand factor |
References
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 4, 213–228. [Google Scholar] [CrossRef]
- Dauros Singorenko, P.; Chang, V.; Whitcombe, A.; Simonov, D.; Hong, J.; Phillips, A.; Swift, S.; Blenkiron, C. Isolation of membrane vesicles from prokaryotes: A technical and biological comparison reveals heterogeneity. J. Extracell. Vesicles 2017, 1, 1324731. [Google Scholar] [CrossRef] [PubMed]
- Dovizio, M.; Bruno, A.; Contursi, A.; Grande, R.; Patrignani, P. Platelets and extracellular vesicles in cancer: Diagnostic and therapeutic implications. Cancer Metastasis Rev. 2018, 37, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Wojtukiewicz, M.Z.; Sierko, E.; Hempel, D.; Tucker, S.C.; Honn, K. Platelets and cancer angiogenesis nexus. Cancer Metastasis Rev. 2017, 2, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Tableros, V.; Gomez, Y.; Camussi, G.; Brizzi, M.F. Extracellular vesicles: New players in lymphomas. Int. J. Mol. Sci. 2018, 21, 41. [Google Scholar] [CrossRef] [PubMed]
- Lorenc, T.; Klimczyk, K.; Michalczewska, I.; Słomka, M.; Kubiak-Tomaszewska, G.; Olejarz, W. Exosomes in prostate cancer diagnosis, prognosis and therapy. Int. J. Mol. Sci. 2020, 21, 2118. [Google Scholar] [CrossRef]
- Meldolesi, J. Extracellular vesicles, news about their role in immune cells: Physiology, pathology and diseases. Clin. Exp. Immunol. 2019, 13, 318–327. [Google Scholar] [CrossRef]
- Stahl, P.D.; Raposo, G. Extracellular vesicles: Exosomes and microvesicles, integrators of homeostasis. Physiology 2019, 3, 169–177. [Google Scholar] [CrossRef]
- Frydrychowicz, M.; Kolecka-Bednarczyk, A.; Madejczyk, M.; Yasar, S.; Dworacki, G. Exosomes—Structure, biogenesis and biological role in non-small-cell lung cancer. Scand. J. Immunol. 2015, 81, 2–10. [Google Scholar] [CrossRef]
- Żmigrodzka, M.; Guzera, M.; Miśkiewicz, A.; Jagielski, D.; Winnicka, A. The biology of extracellular vesicles with focus on platelet microparticles and their role in cancer development and progression. Tumour. Biol. 2016, 11, 14391–14401. [Google Scholar] [CrossRef]
- Van der Pol, E.; Böing, A.N.; Gool, E.L.; Nieuwland, R. Recent developments in the nomenclature, presence, isolation, detection and clinical impact of extracellular vesicles. J. Thromb. Haemost. 2016, 14, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Menck, K.; Bleckmann, A.; Wachter, A.; Hennies, B.; Ries, L.; Schulz, M.; Balkenhol, M.; Pukrop, T.; Schatlo, B.; Rost, U.; et al. Characterisation of tumour-derived microvesicles in cancer patients’ blood and correlation with clinical outcome. J. Extracell. Vesicles 2017, 1, 1340745. [Google Scholar] [CrossRef] [PubMed]
- Laroche, M.; Dunois, C.; Vissac, A.M.; Amiral, J. Update on functional and genetic laboratory assays for the detection of platelet microvesicles. Platelets 2017, 3, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Italiano, J.E., Jr.; Mairuhu, A.T.; Flaumenhaft, R. Clinical relevance of microparticles from platelets and megakaryocytes. Curr. Opin. Hematol. 2010, 6, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Berckmans, R.J.; Nieuwland, R.; Böing, A.N.; Romijn, F.P.; Hack, C.E.; Sturk, A. Cell-derived microparticles circulate in healthy humans and support low grade thrombin generation. Thromb. Haemost. 2001, 4, 639–646. [Google Scholar]
- Wolf, P. The nature and significance of platelet products in human plasma. Br. J. Haematol. 1967, 3, 269–288. [Google Scholar] [CrossRef]
- Melki, I.; Tessandier, N.; Zufferey, A.; Boilard, E. Platelet microvesicles in health and disease. Platelets 2017, 3, 214–221. [Google Scholar] [CrossRef]
- Van der Pol, E.; Harrison, P. From platelet dust to gold dust: Physiological importance and detection of platelet microvesicles. Platelets 2017, 3, 211–213. [Google Scholar] [CrossRef]
- Sinauridze, E.I.; Kireev, D.A.; Popenko, N.Y.; Pichugin, A.V.; Panteleev, M.A.; Krymskaya, O.V.; Ataullakhanov, F.I. Platelet microparticle membranes have 50- to 100-fold higher specific procoagulant activity than activated platelets. Thromb. Haemost. 2007, 97, 425–434. [Google Scholar]
- Aatonen, M.; Grönholm, M.; Siljander, P.R. Platelet-derived microvesicles: Multitalented participants in intercellular communication. Semin. Thromb. Hemost. 2012, 1, 102–113. [Google Scholar] [CrossRef]
- Zubairova, L.D.; Nabiullina, R.M.; Nagaswami, C.; Zuev, Y.F.; Mustafin, I.G.; Litvinov, R.I.; Weisel, J.W. Circulating microparticles alter formation, structure, and properties of fibrin clots. Sci. Rep. 2015, 5, 17611. [Google Scholar] [CrossRef] [PubMed]
- Castaman, G.; Li, Y.-F.; Battistin, E.; Rodeghiero, F. Characterization of a novel bleeding disorder with isolated prolonged bleeding time and deficiency of platelet microvesicle generation. Br. J. Haematol. 1997, 96, 458–463. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, Y.; Lin, Z.; Xiao, X.; He, C.; Bihl, J.C.; Zhao, B.; Ma, X.; Chen, Y. The role of circulating platelets microparticles and platelet parameters in acute ischemic stroke patients. J. Stroke Cereb. Dis. 2015, 10, 2313–2320. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Román, M.T.; Fernández-Bello, I.; Jiménez-Yuste, V.; Martín-Salces, M.; Arias-Salgado, E.G.; Rivas Pollmar, M.I.; Justo Sanz, R.; Butta, N.V. Procoagulant profile in patients with immune thrombocytopenia. Br. J. Haematol. 2016, 5, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Song, K.S.; Park, Y.S.; Kang, Y.H.; Lee, Y.J.; Lee, K.R.; Kim, H.K.; Ryu, K.W.; Bae, J.M.; Kim, S. Elevated levels of circulating platelet microparticles, VEGF, IL-6 and RANTES in patients with gastric cancer: Possible role of a metastasis predictor. Eur. J. Cancer 2003, 2, 184–191. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 19, 9412–9420. [Google Scholar]
- Edelstein, L.C. The role of platelet microvesicles in intercellular communication. Platelets 2017, 3, 222–227. [Google Scholar] [CrossRef]
- Schoenwaelder, S.M.; Yuan, Y.; Josefsson, E.C.; White, M.J.; Yao, Y.; Mason, K.D.; O’Reilly, L.A.; Henley, K.J.; Ono, A.; Hsiao, S.; et al. Two distinct pathways regulate platelet phosphatidylserine exposure and procoagulant function. Blood 2009, 3, 663–666. [Google Scholar] [CrossRef]
- Dale, G.L.; Remenyi, G.; Friese, P. Tetraspanin CD9 is required for microparticle release from coated-platelets. Platelets 2009, 20, 361–366. [Google Scholar] [CrossRef]
- Brisson, A.R.; Tan, S.; Linares, R.; Gounou, C.; Arraud, N. Extracellular vesicles from activated platelets: A semiquantitative cryo-electron microscopy and immuno-gold labeling study. Platelets 2017, 3, 263–271. [Google Scholar] [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 75. [Google Scholar] [CrossRef] [PubMed]
- Meldolesi, J. Exosomes and ectosomes in intercellular communication. Curr. Biol. 2018, 8, R435–R444. [Google Scholar] [CrossRef] [PubMed]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell Mol. Life Sci. 2018, 2, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Catalano, M.; O’Driscoll, L. Inhibiting extracellular vesicles formation and release: A review of EV inhibitors. J. Extracell. Vesicles 2019, 9, 1703244. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, S.; Luo, L.; Norstrom, E.; Braun, O.O.; Morgelin, M.; Thorlacius, H. Platelet-derived microparticles regulates thrombin generation via phophatidylserine in abdominal sepsis. J. Cell Physiol. 2018, 2, 1051–1060. [Google Scholar] [CrossRef]
- Aatonen, M.T.; Ohman, T.; Nyman, T.A.; Laitinen, S.; Grönholm, M.; Siljander, P.R. Isolation and characterization of platelet-derived extracellular vesicles. J. Extracell. Vesicles 2014. [Google Scholar] [CrossRef]
- Lea, J.; Sharma, R.; Yang, F.; Zhu, H.; Ward, E.S.; Schroit, A.J. Detection of phosphatidylserine-positive exosomes as a diagnostic marker for ovarian malignancies: A proof of concept study. Oncotarget 2017, 9, 14395–14407. [Google Scholar] [CrossRef]
- Rand, M.L.; Wang, H.; Bang, K.W.; Packham, M.A.; Freedman, J. Rapid clearance of procoagulant platelet-derived microparticles from the circulation of rabbits. J. Thromb. Haemost. 2006, 7, 1621–1623. [Google Scholar] [CrossRef]
- Rank, A.; Nieuwland, R.; Crispin, A.; Grützner, S.; Iberer, M.; Toth, B.; Pihusch, R. Clearance of platelet microparticles in vivo. Platelets 2011, 2, 111–116. [Google Scholar] [CrossRef]
- Flaumenhaft, R. Formation and fate of platelet microparticles. Blood Cells Mol. Dis. 2006, 2, 182–187. [Google Scholar] [CrossRef]
- Abdel-Monem, H.; Dasgupta, S.K.; Le, A.; Prakasam, A.; Thiagarajan, P. Phagocytosis of platelet microvesicles and beta2- glycoprotein I. Thromb. Haemost. 2010, 2, 335–341. [Google Scholar]
- Dasgupta, S.K.; Le, A.; Chavakis, T.; Rumbaut, R.E.; Thiagarajan, P. Developmental endothelial locus-1 (Del-1) mediates clearance of platelet microparticles by the endothelium. Circulation 2012, 13, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Record, M.; Silvente-Poirot, S.; Poirot, M.; Wakelam, M.J.O. Extracellular vesicles: Lipids as key components of their biogenesis and functions. J. Lipid. Res. 2018, 8, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- Charoenviriyakul, C.; Takahashi, Y.; Morishita, M.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Cell type-specific and common characteristics of exosomes derived from mouse cell lines: Yield, physicochemical properties, and pharmacokinetics. Eur. J. Pharm. Sci. 2017, 1, 316–322. [Google Scholar] [CrossRef]
- Wisgrill, L.; Lamm, C.; Hartmann, J.; Preiβing, F.; Dragostis, K.; Bee, A.; Hell, L.; Thaler, J.; Ay, C.; Pabinger, I.; et al. Peripheral blood microvesicles secretion is influenced by storage time, temperature, and anticoagulants. Cytometry A 2016, 7, 663–672. [Google Scholar] [CrossRef]
- Flaumenhaft, R.; Dilks, J.R.; Richardson, J.; Alden, E.; Patel-Hett, S.R.; Battinelli, E.; Klement, G.L.; Sola-Visner, M.; Italiano, J.E., Jr. Megakaryocyte-derived microparticles: Direct visualization and distinction from platelet-derived microparticles. Blood 2009, 5, 1112–1121. [Google Scholar] [CrossRef]
- Vajen, T.; Mause, S.F.; Koenen, R.R. Microvesicles from platelets: Novel drivers of vascular inflammation. Thromb. Haemost. 2015, 2, 228–236. [Google Scholar] [CrossRef]
- Rank, A.; Nieuwland, R.; Delker, R.; Kohler, A.; Toth, B.; Pihusch, V. Cellular origin of platelet-derived microparticles In Vivo. Thromb. Res. 2010, 126, e255–e259. [Google Scholar] [CrossRef]
- Heijnen, H.F.; Schiel, A.E.; Fijnheer, R.; Geuze, H.J.; Sixma, J.J. Activated platelets release two types of membrane vesicles: Microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood 1999, 11, 3791–3799. [Google Scholar] [CrossRef]
- Dean, W.L.; Lee, M.J.; Cummins, T.D.; Schultz, D.J.; Powell, D.W. Proteomic and functional characterisation of platelet microparticle size classes. Thromb. Haemost. 2009, 102, 711–718. [Google Scholar] [CrossRef]
- Menter, D.G.; Kanikarla-Marie, P.; Lam, M.; Davis, J.S.; Kopetz, S. Platelet microparticles: Small payloads with profound effects on tumor growth. Noncoding RNA Investig. 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Sadallah, S.; Eken, C.; Martin, P.J.; Schifferli, J.A. Microparticles (ectosomes) shed by stored human platelets downregulate macrophages and modify the development of dendritic cells. J. Immunol. 2011, 11, 6543–6552. [Google Scholar] [CrossRef] [PubMed]
- Falati, S.; Liu, Q.; Gross, P.; Merrill-Skoloff, G.; Chou, J.; Vandendries, E.; Celi, A.; Croce, K.; Furie, B.C.; Furie, B. Accumulation of tissue factor into developing thrombi in vivo is dependent upon microparticle Pselectin glycoprotein ligand 1 and platelet P-selectin. J. Exp. Med. 2003, 11, 1585–1598. [Google Scholar] [CrossRef] [PubMed]
- Diamant, M.; Nieuwland, R.; Pablo, R.F.; Sturk, A.; Smit, J.W.; Radder, J.K. Elevated numbers of tissue-factor exposing microparticles correlate with components of the metabolic syndrome in uncomplicated type 2 diabetes mellitus. Circulation 2002, 19, 2442–2447. [Google Scholar] [CrossRef]
- Tang, K.; Liu, J.; Yang, Z.; Zhang, B.; Zhang, H.; Huang, C.; Ma, J.; Shen, G.X.; Ye, D.; Huang, B. Microparticles mediate enzyme transfer from platelets to mast cells: A new pathway for lipoxin a4 biosynthesis. Biochem. Biophys. Res. Commun. 2010, 3, 432–436. [Google Scholar] [CrossRef]
- Kailashiya, J.; Gupta, V.; Dash, D. Engineered human platelet-derived microparticles as natural vectors for targeted drug delivery. Oncotarget 2019, 56, 5835–5846. [Google Scholar] [CrossRef]
- Baj-Krzyworzeka, M.; Majka, M.; Pratico, D.; Ratajczak, J.; Vilaire, G.; Kijowski, J.; Reca, R.; Janowska-Wieczorek, A.; Ratajczak, M.Z. Platelet-derived microparticles stimulate proliferation, survival, adhesion, and chemotaxis of hematopoietic cells. Exp. Hematol. 2002, 5, 450–459. [Google Scholar] [CrossRef]
- Gelderman, M.P.; Simak, J. Flow cytometric analysis of cell membrane microparticles. Methods Mol. Biol. 2008, 484, 79–93. [Google Scholar]
- Abid, H.M.N.; Meesters, E.W.; Osmanovic, N.; Romijn, F.P.; Nieuwland, R.; Sturk, A. Antigenic characterization of endothelial cellderived microparticles and their detection ex vivo. J. Thromb. Haemost. 2003, 11, 2434–2443. [Google Scholar] [CrossRef]
- Thiagarajan, P.; Tait, J.F. Collagen-induced exposure of anionic phospholipid in platelets and platelet-derived microparticles. J. Biol. Chem. 1991, 36, 24302–24307. [Google Scholar]
- Rozmyslowicz, T.; Majka, M.; Kijowski, J.; Murphy, S.L.; Conover, D.O.; Poncz, M.; Ratajczak, J.; Gaulton, G.N.; Ratajczak, M.Z. Platelet- and megakaryocyte-derived microparticles transfer CXCR4 receptor to CXCR4-null cells and make them susceptible to infection by X4-HIV. AIDS 2003, 1, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gasperi, W.; Vangapandu, C.; Savini, I.; Ventimiglia, G.; Adoro, G.; Catani, M.V. Polyunsaturated fatty acids modulate the delivery of platelet microvesicle-derived microRNAs into human breast cancer cell lines. J. Nutr. Biochem. 2019, 74, 108242. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, G.E.; Sims, P.J.; Wiedmer, T.; Furie, B.; Furie, B.C.; Shattil, S.J. Platelet-derivedmicroparticles express high affinity receptors for factor VIII. J. Biol. Chem. 1991, 26, 17261–17268. [Google Scholar]
- Brill, A.; Dashevsky, O.; Rivo, J.; Gozal, Y.; Varon, D. Platelet-derived microparticles induce angiogenesis and stimulate post-ischemic revascularization. Cardiovasc. Res. 2005, 1, 30–38. [Google Scholar] [CrossRef]
- Fox, J.E.; Austin, C.D.; Boyles, J.K.; Steffen, P.K. Role of the membrane skeleton in preventing the shedding of procoagulant-rich microvesicles from the platelet plasma membrane. J. Cell Biol. 1990, 2, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Podor, T.J.; Singh, D.; Chindemi, P.; Foulon, D.M.; McKelvie, R.; Weitz, J.I.; Austin, R.; Boudreau, G.; Davies, R. Vimentin exposed on activated platelets and platelet microparticles localizes vitronectin and plasminogen activator inhibitor complexes on their surface. J. Biol. Chem. 2002, 9, 7529–7539. [Google Scholar] [CrossRef]
- Barry, O.P.; Praticò, D.; Savani, R.C.; FitzGerald, G.A. Modulation of monocyte-endothelial cell interactions by platelet microparticles. J. Clin. Investig. 1998, 1, 136–144. [Google Scholar] [CrossRef]
- Barry, O.P.; Kazanietz, M.G.; Praticò, D.; FitzGerald, G.A. Arachidonic acid in platelet microparticles up-regulates cyclooxygenase-2-dependent prostaglandin formation via a protein kinase C/mitogen-activated protein kinase-dependent pathway. J. Biol. Chem. 1999, 11, 7545–7556. [Google Scholar] [CrossRef]
- Boilard, E.; Nigrovic, P.A.; Larabee, K.; Watts, G.F.M.; Coblyn, J.S.; Weinblatt, M.E.; Massarotti, E.M.; Remold-O’Donnell, E.; Farndale, R.W.; Ware, J.; et al. Platelets amplify inflammation in arthritis via collagen-dependent microparticle production. Science 2010, 5965, 580–583. [Google Scholar] [CrossRef]
- Sims, P.J.; Faioni, E.M.; Wiedmer, T.; Shattil, S.J. Complement proteins C5b-9 cause release of membrane vesicles from the platelet surface that are enriched in the membrane receptor for coagulation factor Va and express prothrombinase activity. J. Biol. Chem. 1988, 34, 18205–18212. [Google Scholar]
- Raturi, A.; Miersch, S.; Hudson, J.W.; Mutus, B. Platelet microparticleassociated protein disulfide isomerase promotes platelet aggregation and inactivates insulin. Biochim. Biophys. Acta 2008, 12, 2790–2796. [Google Scholar] [CrossRef] [PubMed]
- Böing, A.N.; Hau, C.M.; Sturk, A.; Nieuwland, R. Platelet microparticles contain active caspase 3. Platelets 2008, 19, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Mause, S.F.; von Hundelshausen, P.; Zernecke, A.; Koenen, R.R.; Weber, C. Platelet microparticles: A transcellular delivery system for RANTES promoting monocyte recruitment on endothelium. Arter. Thromb. Vasc. Biol. 2005, 7, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Maroney, S.A.; Haberichter, S.L.; Friese, P.; Collins, M.L.; Ferrel, J.P.; Dale, G.L.; Mast, A.E. Active tissue factor pathway inhibitor is expressed on the surface of coated platelets. Blood 2007, 5, 1931–1937. [Google Scholar] [CrossRef] [PubMed]
- Gambim, M.H.; do Carmo, A.; Marti, L.; Veríssimo-Filho, S.; Lopes, L.R.; Janiszewski, M. Platelet-derived exosomes induce endothelial cell apoptosis through peroxynitrite generation: Experimental evidence for a novel mechanism of septic vascular dysfunction. Crit. Care 2007, 5, R107. [Google Scholar] [CrossRef] [PubMed]
- Pfister, S.L. Role of platelet microparticles in the production of thromboxane by rabbit pulmonary artery. Hypertension 2004, 2, 428–433. [Google Scholar] [CrossRef]
- Salanova, B.; Choi, M.; Rolle, S.; Wellner, M.; Luft, F.C.; Kettritz, R. Beta2-integrins and acquired glycoprotein IIb/IIIa (GPIIb/IIIa) receptors cooperate in NF-kappaB activation of human neutrophils. J. Biol. Chem. 2007, 38, 27960–27969. [Google Scholar] [CrossRef]
- Kim, H.K.; Song, K.S.; Chung, J.H.; Lee, K.R.; Lee, S.N. Platelet microparticles induce angiogenesis in vitro. Br. J. Haematol. 2004, 3, 376–384. [Google Scholar] [CrossRef]
- Saber, S.H.; Ali, H.E.A.; Gaballa, R.; Gaballah, M.; Ali, H.I.; Zerfaoui, M.; Abd Elmageed, Z.Y. Exosomes are the driving force in preparing the soil for the metastatic seeds: Lessons from the prostate cancer. Cells 2020, 3, 564. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 7, 727. [Google Scholar] [CrossRef]
- Nishida, N.; Yano, H.; Nishida, T.; Kamura, T.; Kojiro, M. Angiogenesis in cancer. Vasc. Health Risk Manag. 2006, 3, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Happonen, K.E.; Tran, S.; Mörgelin, M.; Prince, R.; Calzavarini, S.; Angelillo-Scherrer, A.; Dählback, B. The Gas6-Axl protein interaction mediates endothelial uptake of platelet microparticles. J. Biol. Chem. 2016, 20, 10586–10601. [Google Scholar] [CrossRef] [PubMed]
- Janowska-Wieczorek, A.; Wysoczynski, M.; Kijowski, J.; Marquez-Curtis, L.; Machalinski, B.; Ratajczak, J.; Ratajczak, M.Z. Microvesicles derived from activated platelets induce metastasis and angiogenesis in lung cancer. Int. J. Cancer 2005, 5, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Michael, J.V.; Wurtzel, J.G.T.; Mao, G.F.; Rao, A.K.; Kolpakov, M.A.; Sabri, A.; Hoffman, N.E.; Rajan, S.; Tomar, D.; Madesh, M.; et al. Platelet microparticles infiltrating solid tumors transfer miRNAs that suppress tumor growth. Blood 2017, 5, 567–580. [Google Scholar] [CrossRef]
- Anene, C.; Graham, A.M.; Boyne, J.; Roberts, W. Platelet microparticle delivered microRNA-Let-7a promotes the angiogenic switch. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 8, 2633–2643. [Google Scholar] [CrossRef]
- Lazar, S.; Goldfinger, L.E. Platelet microparticles and miRNA transfer in cancer progression: Many targets, modes of action, and effects across cancer stages. Front. Cardiovasc. Med. 2018, 5, 13. [Google Scholar] [CrossRef]
- Pan, Y.; Liang, H.; Liu, H.; Li, D.; Chen, X.; Li, L.; Zhang, C.Y.; Zen, K. Platelet-secreted microRNA-223 promotes endothelial cell apoptosis induced by advanced glycation end products via targeting the insulin-like growth factor 1 receptor. J. Immunol. 2014, 1, 437–446. [Google Scholar] [CrossRef]
- Laffont, B.; Corduan, A.; Rousseau, M.; Duchez, A.C.; Lee, C.H.C.; Boilard, E.; Provost, P. Platelet microparticles reprogram macrophage gene expression and function. Thromb. Haemost. 2016, 2, 311–323. [Google Scholar]
- Bakewell, S.J.; Nestor, P.; Prasad, S.; Tomasson, M.H.; Dowland, N.; Mehrorta, M.; Scarborough, R.; Kanter, J.; Abe, K.; Phillips, D.; et al. Platelet and osteoclast beta3 integrins are critical for bone metastasis. Proc. Natl. Acad. Sci. USA 2003, 24, 14205–14210. [Google Scholar] [CrossRef]
- Liu, S.C.; Bassi, D.E.; Zhang, S.Y.; Holoran, D.; Conti, C.J.; Klein-Szanto, A.J. Overexpression of cyclin D2 is associated with increased in vivo invasiveness of human squamous carcinoma cells. Mol. Carcinog. 2002, 3, 131–139. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Omega-3 polyunsaturated fatty acids and their health benefits. Ann. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Fisslthaler, B.; Zippel, N.; Frömel, T.; Hu, J.; Elgheznawy, A.; Heide, H.; Popp, R.; Fleming, I. MicroRNA-223 antagonizes angiogenesis by targeting β1 integrin and preventing growth factor signaling in endothelial cells. Circ. Res. 2013, 113, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.L.; Jiang, L.; Lin, Y.Y.; Wu, X.L.; Wang, K.; He, Q.Z.; Wang, X.P.; Li, W.P. Platelet microparticle mediated transfer of miR-939 to epithelial ovarian cancer cells promotes epithelial to mesenchymal transition. Oncotarget 2017, 8, 97464–97475. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Qu, S.; Hu, R.; Gao, W.; Jin, S.; Ju, J.; Zhao, Q. Delivery of platelet TPM3 mRNA into breast cancer cells via microvesicles enhances metastasis. FEBS Open Biol. 2019, 12, 2159–2169. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.B.; Gao, Q.; Lin, W.W.; Kang, M.Q. Proteomic analysis indicates the importance of TPM3 in esophageal squamous cell carcinoma invasion and metastasis. Mol. Med. Rep. 2017, 15, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Zarà, M.; Guidetti, G.F.; Boselli, D.; Villa, C.; Canobbio, I.; Seppi, C.; Visconte, C.; Canio, J.; Torti, M. Release of prometastatic platelet-derived microparticles induced by breast cancer cells: A novel positive feedback mechanism for metastasis. TH Open 2017, 2, e155–e163. [Google Scholar] [CrossRef]
- Burnouf, T.; Goubran, H.A.; Chou, M.L.; Devos, D.; Radosevic, M. Platelet microparticles: Detection and assessment of their paradoxical functional roles in disease and regenerative medicine. Blood Rev. 2014, 4, 155–166. [Google Scholar] [CrossRef]
- Vasina, E.M.; Cauwenberghs, S.; Feijge, M.A.; Heemskerk, J.W.; Weber, C.; Koenen, R.R. Microparticles from apoptotic platelets promote resident macrophage differentiation. Cell Death Dis. 2011, 9, e211. [Google Scholar] [CrossRef]
- Li, J.; Guo, Y.; Liang, X.; Sun, M.; Wang, G.; De, W.; Wu, W. MicroRNA-223 functions as an oncogene in human gastric cancer by targeting FBXW7/hCdc4. J. Cancer Res. Clin. Oncol. 2012, 5, 763–774. [Google Scholar] [CrossRef]
- Yaftian, M.; Yari, F.; Ghasemzadeh, M.; Fallah, A.V.; Haghighi, M. Induction of apoptosis in cancer cells of pre-B ALL patients after exposure to platelets, platelet-derived microparticles and soluble CD40 ligand. Cell J. 2018, 1, 120–126. [Google Scholar]
- Dashevsky, O.; Varon, D.; Brill, A. Platelet-derived microparticles promote invasiveness of prostate cancer cells via upregulation of MMP-2 production. Int. J. Cancer 2009, 8, 1773–1777. [Google Scholar] [CrossRef] [PubMed]
- Sadallah, S.; Schmied, L.; Eken, C.; Charoudeh, H.N.; Amicarella, F.; Schifferli, J.A. Platelet-derived ectosomes reduce NK cell function. J. Immunol. 2016, 5, 1663–1671. [Google Scholar] [CrossRef] [PubMed]
- Laffont, B.; Corduan, A.; Plé, H.; Duchez, A.C.; Cloutier, N.; Boilard, E.; Provost, P. Activated platelets can deliver mRNA regulatory Ago2 microRNA complexes to endothelial cells via microparticles. Blood 2013, 2, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Kang, K.W. Phosphatidylserine receptor-targeting therapies for the treatment of cancer. Arch. Pharm. Res. 2019, 7, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Antonyak, M.A.; Zhang, J.; Cerione, R.A. RhoA triggers a specific signaling pathway that generates transforming microvesicles in cancer cells. Oncogene 2012, 45, 4740–4749. [Google Scholar] [CrossRef]
- Zhang, W.; Qi, J.; Zhao, S.; Shen, W.; Dai, L.; Han, W.; Huang, M.; Wang, Z.; Ruan, C.; Wu, D.; et al. Clinical significance of circulating microparticles in Ph- myeloproliferative neoplasms. Oncol. Lett. 2017, 2, 2531–2536. [Google Scholar] [CrossRef]
- Ren, J.G.; Man, Q.W.; Zhang, W.; Li, C.; Xiong, X.P.; Zhu, J.Y.; Wang, W.M.; Sun, Z.J.; Jia, J.; Zhang, W.F.; et al. Elevated level of circulating platelet-derived microparticles in oral cancer. J. Dent. Res. 2016, 1, 87–93. [Google Scholar] [CrossRef]
- Dymicka-Piekarska, V.; Gryko, M.; Lipska, A.; Korniluk, A.; Siergiejko, E.; Kemona, H. Platelet-derived microparticles in patients with colorectal cancer. J. Cancer Ther. 2012, 6, 898–901. [Google Scholar] [CrossRef]
- Wang, C.C.; Tseng, C.C.; Chang, H.C.; Huang, K.T.; Fang, W.F.; Chen, Y.M.; Yang, C.T.; Hsiao, C.C.; Lin, M.C.; Ho, C.K.; et al. Circulating microparticles are prognostic biomarkers in advanced non-small cell lung cancer patients. Oncotarget 2017, 44, 75952–75967. [Google Scholar] [CrossRef]
- Mege, D.; Panicot-Dubois, L.; Ouaissi, M.; Robert, S.; Sielezneff, I.; Sastre, B.; Digant-George, F.; Dubois, C. The origin and concentration of circulating microparticles differ according to cancer type and evolution: A prospective single-center study. Int. J. Cancer 2016, 4, 939–948. [Google Scholar] [CrossRef]
- Yenigürbüz, F.D.; Kızmazoğlu, D.; Ateş, H.; Erdem, M.; Tufekci, O.; Yilmaz, S.; Oren, H. Analysis of apoptotic, platelet-derived, endothelial-derived, and tissue factor-positive microparticles of children with acute lymphoblastic leukemia during induction therapy. Blood Coagul. Fibrinolysis 2019, 4, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.; Nugent, K. Microparticles in Hematological Malignancies: Role in Coagulopathy and Tumor Pathogenesis. Am. J. Med. Sci. 2018, 3, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Tjon-Kon-Fat, L.A.; Lundholm, M.; Schroder, M.; Wurdinger, T.; Thellenberg-Karlsson, C.; Widmark, A.; Wikstrom, P.; Nilsson, R.J.A. Platelets harbor prostate cancer biomarkers and the ability to predict therapeutic response to abiraterone in castration resistant patients. Prostate 2018, 1, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Boerrigter, E.; Groen, L.N.; Van Erp, N.P.; Verhaegh, G.W.; Schalken, J.A. Clinical utility of emerging biomarkers in prostate cancer liquid biopsies. Expert Rev. Mol. Diagn. 2020, 2, 219–230. [Google Scholar] [CrossRef]
- Tarasov, V.V.; Svistunov, A.A.; Chubarev, V.N. Extracellular vesicles in cancer nanomedicine. Semin. Cancer Biol. 2019. [Google Scholar] [CrossRef]
- Wiklander, O.P. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef]
- Dilsiz, N. Role of exosomes and exosomal microRNAs in cancer. Future Sci. OA 2020, 4, FSO465. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, D.; Ma, X.; Wang, J.; Hou, W.; Zhang, W. Exosomes as drug carriers for cancer therapy and challenges regarding exosome uptake. Biomed. Pharmacother. 2020, 128, 110237. [Google Scholar] [CrossRef]


{kind=link}
| Function or Category Name | ||||||||
|---|---|---|---|---|---|---|---|---|
| Clotting | Enzymes | Adhesion Molecules | Bioactive Lipids | Programmed Cell Death | Growth Factors | Chemokines /Cytokines | Immune Response | |
| PEVs Cargo (Ref) | TF [52,53,54] | 12-LO [55] | CD41/61 [56,57,58,59] CD31 [49,59] | PS [60] | caspase-3 [58] | TGF β1 [50] | CXCR4-(PF-4) [57,61] | CD 154 [32,62] |
| FVa, FVIII [60,63] | heparynase [64] | CD62P [57,59,65,66] | AA [67,68] | CD95 [57] | PDGF bFGF [64] | IL-1β [69] | C5b-9 [70] | |
| PAR-1 [57] | PDI [71] | fibrinogen, vWF, vitronectin [65] | LPA [70] | caspase-9 [72] | VEGF [64] | CCL5, CCL23 [50,73] | CD55, CD59 [52] | |
| TFPI [74] | NADPH oxidase [75] | CD42a, CD42b [49,59] | TXA2 [76] | CX3CR1 [73] | Factor H [52] | |||
| Target Cell | PEVs Derived Factors/Molecules | Functional Changes (References) |
|---|---|---|
| A549, CRL 2066, CRL 2062, HTB 183, HTB 177 lung CCL; LCC * CCL | CD41, CD61 CD184 | (+) adhesion to fibrinogen and HUVECs [83] (+) metastatic potential [83] (+) mRNA expression of angiogenic factors (MMP-9, VEGF, IL-8) [83] (+) proliferation and chemoinvasion [83] |
| HUVECs | miRNA Let-7a miRNA-223 | (−) synthesis THBS-1 anti-angiogenic molecule [85] (+) apoptosis by IGF-1 [4,87] |
| MC-38 colon CCL, LCC * CCL | miRNA-24 | (+) apoptosis [84] |
| BT549 breast CCL | miRNA-123 miRNA-233 | (−) migration [62] (−) cell cycle [62] |
| SKOV3 ovarian CCL | miRNA-939 | (+) invasion via TPM3 [94] (+) progression [94] |
| MDM-MB-231 breast CCL | (+) invasion [96] (+) migration [94] | |
| SGC7901 gastric CCL | miRNA-223 | (+) proliferation and invasion [4,105] (−) apoptosis [4,105] |
| PBMCs from patients with ALL | CD95 Caspase-3 | (+) apoptosis [100] |
| Cl-1 prostate CCL | MMP-2 miRNA? | (+) migration [101] |
| macrophages | PS, gpIIb/IIIa miR-126-3p | polarization into macrophage M2 [97] (−)CCL4, CSF1, TNF [88] |
| NK cells | miR-183 | (−) cytolysis [102] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żmigrodzka, M.; Witkowska-Piłaszewicz, O.; Winnicka, A. Platelets Extracellular Vesicles as Regulators of Cancer Progression—An Updated Perspective. Int. J. Mol. Sci. 2020, 21, 5195. https://doi.org/10.3390/ijms21155195
Żmigrodzka M, Witkowska-Piłaszewicz O, Winnicka A. Platelets Extracellular Vesicles as Regulators of Cancer Progression—An Updated Perspective. International Journal of Molecular Sciences. 2020; 21(15):5195. https://doi.org/10.3390/ijms21155195
Chicago/Turabian StyleŻmigrodzka, Magdalena, Olga Witkowska-Piłaszewicz, and Anna Winnicka. 2020. "Platelets Extracellular Vesicles as Regulators of Cancer Progression—An Updated Perspective" International Journal of Molecular Sciences 21, no. 15: 5195. https://doi.org/10.3390/ijms21155195
APA StyleŻmigrodzka, M., Witkowska-Piłaszewicz, O., & Winnicka, A. (2020). Platelets Extracellular Vesicles as Regulators of Cancer Progression—An Updated Perspective. International Journal of Molecular Sciences, 21(15), 5195. https://doi.org/10.3390/ijms21155195