Abstract
Shoot apical meristems (SAM) are tissues that function as a site of continuous organogenesis, which indicates that a small pool of pluripotent stem cells replenishes into lateral organs. The coordination of intercellular and intracellular networks is essential for maintaining SAM structure and size and also leads to patterning and formation of lateral organs. Leaves initiate from the flanks of SAM and then develop into a flattened structure with variable sizes and forms. This process is mainly regulated by the transcriptional regulators and mechanical properties that modulate leaf development. Leaf initiation along with proper orientation is necessary for photosynthesis and thus vital for plant survival. Leaf development is controlled by different components such as hormones, transcription factors, miRNAs, small peptides, and epigenetic marks. Moreover, the adaxial/abaxial cell fate, lamina growth, and shape of margins are determined by certain regulatory mechanisms. The over-expression and repression of various factors responsible for leaf initiation, development, and shape have been previously studied in several mutants. However, in this review, we collectively discuss how these factors modulate leaf development in the context of leaf initiation, polarity establishment, leaf flattening and shape.
1. Introduction
Leaves are the primary organs responsible for photosynthesis and photoperception, and play a key role in plant growth. Their development starts from the shoot apical meristem (SAM), which have a central zone (CZ) that houses pluripotent cells, and a peripheral zone (PZ), responsible for the leaf initiation and their development into a flattened structure [1]. In most plants, the leaf functions as a solar panel, where photosynthesis converts carbon dioxide and water into carbohydrates and oxygen [2]. Leaves are an excellent example of learning how complex organs arise from a simple structure. All leaves are initiated by the recruitment of cells flanking the SAM as simple rod-like primordia, later on, to get their final shape. There are three principal axes in a leaf, along which intrinsic genetic programs control leaf cell division, differentiation, and expansion. However, leaf morphogenesis is strictly controlled not only by intrinsic genetic factors but also by hormonal factors. Numerous series of events demonstrate that plant hormones, mostly small and simple molecules, play crucial roles in plant growth and development [3]. It was hypothesized how hormonal and genetic networks regulate leaf morphogenesis to enable the transformation of simple primordium into a complex organ with consistent shape and size, and to elaborate how these genetic networks generate plasticity in response to both endogenous and environmental signals. Since a deeper understanding of leaf development contributes to our overall comprehension of plant biology, this understanding can also be used to improve crop production. Therefore, it is important to unveil the molecular and hormonal regulation of leaf morphogenesis, including the initiation of leaf primordia, the determination of leaf axes, and the regulation of cell division in model plant Arabidopsis thaliana. The main focus of this review is to summarize the current knowledge and discuss the several fundamental aspects of leaf development, including a genetic regulatory framework that contributes to leaf initiation, leaf polarity determination, and leaf outgrowth and flattening. Furthermore, we also emphasize recent results that have strengthened our understanding of leaf development. The information obtained from the studies of Arabidopsis thaliana simple leaves will continue to provide basic knowledge about the formation and genetic mechanisms involved in compound leaves [4,5].
1.1. Maintenance of Shoot Apical Meristems (SAM) and the Leaf Initiation Gene Network
The stem cells of plant meristems generate new organs and tissues throughout the life of the plant. The above-ground organs in plants are formed through SAM, whereas below-ground organs are created by root apical meristems (RAM). Beside these two apical meristems, various other types of meristems exist in plants, such as lateral meristems (e.g., axillary in the node of the leaves and flowers), intercalary meristems (at the base of the monocots leaf blade), and transient stem-like meristemoids (the precursor for guard cells) [6,7]. The SAM is a dome-shaped structure that comprises a reservoir of stem cells, provides cells that form the branches, leaves, and flowers of the plant, and also retains its own identity. The SAM is organized into several distinct cell layers and various zones [8] (Figure 1). The development and maintenance of SAM are crucial for determining the spatiotemporal arrangement (e.g., clockwise, anticlockwise, spiral and whorled) of aerial organs around the stem. The process of arranging different organs spatially is known as phyllotaxis, which is species and stage-dependent [9,10,11].
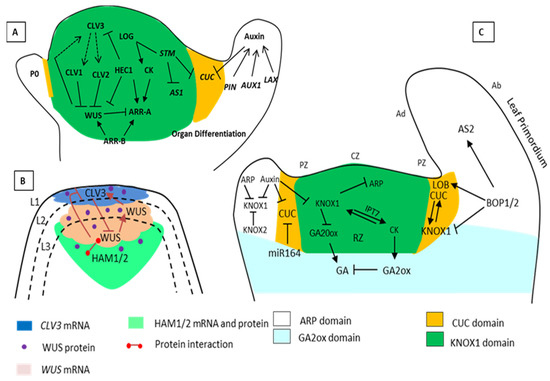
Figure 1.
Schematic representation of Shoot Apical Meristems (SAM) maintenance by the various interacting genes. (A) The pluripotent stage and a specific number of cells in the SAM are controlled by the (WUS/CLV3) negative-feedback loop. CLAVATA3 (CLV3) is a ligand attached to CLV1 and CLV2 for the restriction of WUS. However, WUS activates CLV3 and works as a stem cell-promoting protein. STM activated the biosynthesis of CK through LONELY GUY (LOG). The STM and KNOX related genes also keep the stem cells undifferentiated by suppressing the expression of AS1 and gibberellic acid (GA) biosynthesis. STM also restricts the expression of the CUC gene due to negative regulators in a specific area. (B) The HAM and WUS-CLV3 loop. The regulatory loop requires CLV3, WUS, and HAM; the CLV3 negatively regulates the WUS expression, and the WUS protein moves from the organizing center to the active central zone (stem cells) to activate CLV3 expression. HAM1/2 is an interacting partner of WUS, and together with WUS protein suppresses the expression of CLV3 in the rib meristems. The expression zone of the CLV3 gene (blue), WUS gene (pink), HAM proteins (green) and the dot marks the WUS protein. (C) Regulatory networks that control leaf initiation. The cells in the SAM are arranged into layers L1, L2, and L3 and further into a distinct group of either tunica or corpus. According to the expression of genes, the SAM architecture is organized with the central zone (CZ), peripheral zone (PZ), organizing zone (OZ), and rib zone (RZ). During leaf initiation, auxin maxima repress the expression of the KNOX1 domains (gene) indeterminate meristem domains. KNOX1 maintains a high level of CK and low levels of GA in the meristem. In the ARP domain that has leaf identity, the leaf primordium separates from SAM by expression of boundary specific genes CUC and BOP regulates the petiole specification and polarity (positive and negative regulations are indicated by pointed and T-shaped arrows).
In Arabidopsis thaliana, the SAM possesses three cell layers (L1–L3). An external two-cell layer forms tunica, where the L1 overlies L2. This layer divides by anticlinal cell division and grows in a two-dimensional fashion [12]. An inner layer (L3) divides both periclinal and anticlinal cell divisions in a mostly random fashion, which is commonly called corpus. These three histogenic layers are responsible for producing different parts; the L1 layer divides and forms the entire shoot epidermis while the L2 layer produces the photosynthesizing cells of the sub-epidermis. The L3 layer forms the internal tissue, pericycle, and other corpus cells [13].
The SAM layers are further subdivided into three functional domains or zones according to the function and division rate (Figure 1). The three zones include the central zone (CZ), peripheral zone (PZ), and rib zone (RZ), which is present below the CZ. The CZ is mainly responsible for the maintenance of SAM. The CZ contains both tunica and corpus cells in which the stem cells are present, and below the CZ is the organizing center (OC) [14]. The tunica and corpus cells of the CZ are symplasmically interconnected through the plasmodesmata [15,16]. Any alteration in the intercellular signals through the plasmodesmata affects normal growth and development [17].
The cells in the PZ divide at a faster rate to form leaves, branches, and floral parts than those in the CZ [15,18]. These functional and cytohistological zones are juxtaposed onto cell layers and govern the patterns of development [19,20]. The PZ cells are differentiated and have distinct identities that help to establish future organs such as leaf primordia [3,21]. Cells in the RZ develop into differentiating stems that support the SAM. The maintenance of SAM and the initiation of an organ through a specific phyllotaxis pattern requires strong signal coordination between the different factors such as hormones and genes [22,23].
The initiation and establishment of the shoot meristem in the embryonic stage of Arabidopsis relies on the expression of many different genes and certain signaling proteins. In plants, the NAC family, i.e., CUP-SHAPED COTYLEDON1 (CUC1), CUC2, and CUC3, are the major transcription factors required to initiate SAM and boundary formation (Table 1) [24,25,26,27]. The double mutant of cuc1 and cuc2 genes generates a fused cup-shaped cotyledon structure devoid of SAM. In Arabidopsis, for the development of SAM during embryogenesis and to sustain its function, the activity of class-1 KNOTTED1-LIKE HOMEOBOX (KNOX) gene SHOOT MERISTEMLESS (STM) is required, which is expressed throughout the SAM but downregulated in leaf developing cells [28,29]. In the organ (leaf) founder cells, the activation of organ-specific transcription factors and auxin accumulation inhibits the expression of the KNOX gene (Figure 1) [27]. Other members of the class-1 KNOX gene family, which are expressed in SAM, include KNAT1/BREVIPEDICELLUS (BP), KNAT2, and KNAT6. STM is a central regulator of SAM organization and development, and stronger alleles, such as the stm-1 mutant, totally failed to establish SAM during embryogenesis [30]. In contrast and under normal conditions, other related genes, such as KNAT1/BP, KNAT2, and KNAT6, have no discernible effect on SAM. In the leaf primordium, the MYB domain transcription factors, ASYMMETRIC LEAVES1 (AS1) from Arabidopsis, ROUGH SHEATH2 (RS2) from maize and PHANTASTICA (PHAN) from Antirrhinum (these are collectively named ARP genes), repress the expression of the KNOX1 gene (Table 1) [31,32]. Furthermore, to maintain the meristematic state of the stem cells, the STM inhibits the expression of AS1 in the shoot apex [28,32]. KNOX1 gene BREVIPEDICELLUS (BP) and KNAT2 gene expression are directly repressed by a repressor complex containing AS1, AS2 [33,34]. STM induces the cytokinin (CK) biosynthesis gene, isopentenyl transferase (AtIPT7) which encodes for the enzyme that contributes to the production of active CK in the SAM layer L1; CK interacts with other systematic signals and controls the meristem size and functions [35].

Table 1.
A list of the key genes involved in leaf growth and differentiation.
The WUSCHEL-related homeobox (WOX) gene family transcription factors are broadly distributed in plant species and belong to the homeobox (HB) superfamily. Its members all possess a conserved DNA-binding homeodomain (HD) with 60 to 66 amino acid residues [54]. It has been reported that the number of WOX genes plays a significant role in a wide range of functions, including the maintenance of stem cells, embryonic development and polarization, and the development of lateral organs. In Arabidopsis, 15 WOX genes, WUS, and WOX1-WOX14 have been identified and well-studied for their function. AtWOX1 plays a significant role in meristem development by regulating S-adenosylmethionine decarboxylase (SAMDC) activity or CLV3 expression [55].
Moreover, the WUSCHEL gene (WUS) is required for the specificity and identity of stem-cells at the SAM [51,54]. WUS expression is crucial for meristem maintenance and shoot development [39], but the exact mechanism in which stem-cells are regulated is not fully understood. In the SAM, WUS, as bifunctional proteins, mainly act as a repressor but also becomes an activator when involved in the regulation of the AGAMOUS (AG) gene [56,57]. WUS directly represses the transcription of the Arabidopsis Response Regulator (ARR-A) genes, which encode for the intracellular inhibitor of cytokinin activity [58]. After the recognition of ARR as targets of WUS, a linkage between phytohormone (cytokinin) and the CLV/WUS stem cell network was established [59,60,61,62]. In Arabidopsis, Type-A ARRs are mostly transcriptional repressors, whereas type-B ARRs promote the cytokinin response and act as transcriptional activators [63]. Type-B ARRs, such as ARR1, ARR10, and ARR12, bind to the cis-element located in the promoter region of WUS and activate its expression. Type-A ARR negatively influences the meristem size, and WUS represses the expression of ARR5, ARR6, ARR7, and ARR15 genes for proper meristem function [58,64]. WUS also protects apical stem cell meristems from differentiation by restricting the auxin signaling pathway via regulation of histone deacetylation [65]. To maintain the integrity of SAM, a high concentration of WUS protein repressed the expression of basic helix-loop-helix (bHLH) transcription factor HECATE1 (HEC1), which is expressed throughout the SAM, except for the OC [66,67]. HEC1 forms protein complexes with other bHLH transcription factors such as HEC2 and HEC3. HEC1 activates type-A AAR7 and ARR15 expressions and represses CLV3 expression. Multiple feedback regulatory mechanisms mediated by transcription factors and hormonal components control WUS expression and meristematic fate in SAM [68,69,70].
It is also reported that WUS expression occurs in the OC, but in addition, it can also control the expression of CLV3 in the CZ. The WUS protein migrates and binds to the CLV3 promoter and thus regulates the expression of CLV3 [29,71]. Besides, computational modeling (Figure 1) shows that the WUS gradient is vital for the maintenance and regulation of the stem cell number [72,73]. Various reports have demonstrated that WUS moves to L2 and L1 via plasmodesmata under a highly regulated fashion, and this movement is required for WUS function and stem cell activity [74]. In Arabidopsis, WUS has been shown to interact with members of the HAIRY MERISTEM (HAM) family protein, GRAS-domain transcription factors [75], and HAM1 and HAM2 expressed in the RZ and lateral edges of PZ where CLV3 expression is reduced. In contrast, HAM1 and HAM2 expressions are not detected in the CZ where CLV3 is highly expressed. WUS activates CLV3 only in the absence of HAMs; the apical-basal gradient of HAMs defines the pattern of CLV3 expression domains [75,76]. The regulation of CLV3 by WUS for the maintenance of stem cells is associated with the WUS gradient into the PZ, where the stem cell progeny is differentiated [18,77,78]. The CLV3-related signaling pathway in the stem cell domain is demonstrated by the diffusion of CLV3 peptide towards the inner layer of the meristem, where at least three receptor complexes recognize it. These receptor complexes include CLV1, CLV2, CORYNE (CRN), RECEPTOR-LIKE PROTEIN KINASE2 (RPK2), and BARELY ANY MERISTEM1 (BAM1/2) and represses the expression of WUS [42,43], which is crucial for the stem cell population [79].
The WUS activated gene is present in the central part of the SAM, and the repressed gene is located in the PZ. Additionally, in Arabidopsis, WUS represses those TFs-coding genes that are involved in differentiation, for example, KANADI (KAN1), (KAN2), AS2, YABBY3 (YAB3), KNAT1/BREVIPEDICELLUS (BP), and BELL1-LIKE HOMEODOMAIN5 (BLH5) [80,81]. The identification of these direct interactions can contribute to the understanding of the molecular network but with limitations to explain the mechanisms by which WUS controls stem cell homeostasis. If the stem cell progenitor is relocated beyond the stem cell niche, they then determine whether to be the part of the main axis or divide into lateral organs such as leaf primordia. The fate of distinction is primarily determined by the auxin influx carrier AUXIN RESISTANT1/like-AUX1 (AUX1/LAX) and the auxin efflux transporter PIN-FORMED (PIN1) (Figure 1). The PIN1 efflux carriers control the orientation of auxin transport into the neighboring cell and the concentration of auxin in the group of cells [82,83,84,85]. The auxin concentration may also vary even in the same group of cells. Several models have postulated phyllotactic patterning, which is mainly based on the interaction between auxin accumulation and distribution of the auxin efflux carrier (PIN1) [5,86]. Moreover, AUX1 is responsible for auxin accumulations, which are mainly in the L1 layers of cells. On the contrary, PIN1 drains auxin toward the base of the leaf primordium by inducing vascular tissue differentiation in the L2 and L3 layers [83,87].
1.2. Gene Functioning in Leaf Initiation
Leaf formation starts with the recruiting of founder cells in the peripheral of SAM by changing the pluripotent cell to a differentiated cell. The first step towards the emergence of leaf primordia is PIN1 mediated formation of auxin maxima in the PZ, and to repress the expression of KNOX genes [88]. This activity is necessary because the KNOX genes actively maintain the undifferentiated state of the cells [88]. A mutation in the KNOX genes changes the indeterminate cell to determinate. Auxin maxima enhance the growth and accelerate differentiation, while STM proteins have the opposite effect on both cell growth and differentiation [89]. The critical role of auxin in SAM is to control organogenesis and self-organization of SAM; auxin also specifies the organ primordium fate in the PZ of SAM [90,91]. Mutation in the auxin biosynthesis, transport, and signaling components affect plant growth and morphology. In Arabidopsis, 23 AUXIN RESPONSE FACTORs (ARFs) have been identified, and ARF works as a transcription factor that binds to the auxin-responsive elements (AuxREs) in the promoter region of the target gene [92]. Among other ARF transcription factors, ARF5 (which is also known as MONOPTEROS (MP)) is present in threshold form from PZ to CZ and plays an important role in gene expression to specify the meristematic and primordium fate [93,94]. The MP activates ARABIDOPSIS HISTIDINE PHOSPHOTRANSFER PROTEIN6 (AHP6) expression and inhibits ARR7/ARR15 to control meristematic fate through the regulation of CK homeostasis [95]. Previously, it was suggested that MP was involved in stem cell regulation by inhibiting the expression of DORNRÖSCHEN/ENHANCER OF SHOOT REGENERATION1 (DRN/ESR1) that activated CLV3 expression in the CZ [96,97].
The auxin-efflux protein PIN1 is expressed in the epidermal cell for auxin accumulation [81], which promotes the formation of pro-vascular tissue. PIN1 is the first marker for mid-vein formation before the leaf primordia bulge out [98,99]. A mutation in PIN1 can cause the irregular distribution of auxin and thus disturb the proper initiation of the leaf primordia [85,100]. In Arabidopsis, the pin1 mutant blocked the floral primordium formation, and in tomato shoot apices treated with polar auxin transport inhibitor, such as N-1-naphthylphthalamic acid (NPA), abolished leaf formation [101,102]. The external application of auxin restored primordium formation in the NPA-inhibitor and pin1 mutant, which indicates the importance of auxin in organ initiation [103]. In the peripheral of the meristems, organ primordia are separated and form a groove at the base that contains small slow-dividing and slow-expanding cells called the boundary domain [19,104]. The boundary domain is defined by several factors, including the NAM/CUC transcription factor family [25,105]. Mutants of CUC genes exhibited a lack of SAM and fused lateral organs, which indicated that this boundary domain contributes to meristem formation and enables the organ separation, as well as providing a particular status of hormones such as low brassinosteroid (BR) and auxin levels [105,106]. The cuc3 loss of function mutant restrains leaf serration, which suggests the role of CUC3 in leaf serration. Moreover, the overexpression of KNOX1 genes in lobed leaf phenotypes was suppressed in the cuc2-3 mutant, which indicates that CUCs act downstream of KNOX-induced alteration in leaf morphology [46,107].
Cytokinin (CK) and gibberellic acid (GA) affect cell division, and KNOX1 can increase the biogenesis of CK by up-regulating the IPT7 genes that further block the GA20-oxidase gene required for GA biosynthesis (Figure 1) [108]. A high GA to CK ratio promotes determinacy; in contrast, a low GA to CK ratio facilitates the indeterminacy of the cells. Additionally, the KNOX-independent genetic pathway involves WUS and CLV3 that control the stem cells’ fates directly by regulating cytokinin-inducible response factors (discussed in the maintenance of SAM). A different pathway attains a low CK and a high GA, while determining how auxin promotes organ growth and is integrated with cell fate allocation by KNOX/AS1 protein is unclear. Furthermore, in the as2 mutant background increases in the transcript level of IPT3 and KIP-RELATED PROTEIN2 (KRP2) and KRP5, and this upregulation is a decline in introducing the double mutant of ett, arf4 in the as2 mutant background. Therefore, it is suggested that the expression of IPT3, KRP2, and KRP5 is negatively regulated by AS1-AS2 through repression of ETT/ARF3 and the ARF4 function in the wild type [109]. In Arabidopsis, KRP2 and KRP5 encode for cyclin-dependent kinase inhibitors (CKIs), which is a key regulator of cell progression. During leaf formation, required cell proliferation is achieved by proper repressive control of KRP2 and KRP5 expression by AS1-AS2 [110].
In Arabidopsis, another pathway involves the MYB transcription factor encoding for ARP genes, which is expressed in the founder cells of the lateral organs, and represses the expression of the KNOX1 gene that tends to promote leaf development (Figure 1) [31]. In Arabidopsis, the epigenetic repression of KNOX genes involves the binding of the AS1-AS2 complex to a specific sequence in the promoter region of the BP gene and inhibits its expression. It was confirmed that AS1-AS2 binds to specific sequences in the BP gene that works as a PRE (polycomb response element); the AS1-AS2 complex recruits the polycomb repressive complex 2 (PRC2), which is a stable silencer of STM regulators [111].
Furthermore, many reports indicate the precise regulatory mechanisms of KNOX1 genes. In this regard, various genes (AS1, AS2, SERRATE (SE), BLADE ON PETIOLE1 (BOP1) and PICKEL (PKL)) were found to be involved in the down-regulation of the KNOX (KNAT1, KNAT2) gene [32,112]. The mutants as1 and as2 have some similar phenotypic characteristics due to the ectopic expression of KNOX1 genes and this expression results in the segmentation of the leaf primordium [32]. Moreover, SE was found to enhance the phenotypic expression of the as1 and as2 mutant. KNOX genes are normally regulated in both pkl and se mutant leaves; however, KNOX target gene GA20ox1 is repressed, suggesting that PKL, SE and KNOX activities cover at least one specific target gene. SE represses a small family of micro-RNA targeted Class III HD-ZIP genes that promote meristem activity, the se mutant showed enhanced response to KNOX activity, indicating elevated level of HD-ZIP III expression, HD-ZIP III gain of function mutant reduced the expression of GA20ox1, mimicking effects of KNOX overexpression [113]. Class 1 KNOX gene overexpression prolongs the proliferation of the leaf cell within the lamina [114]. BOP1 and BOP2 are expressed at the proximal domain of the leaf primordium and activate the expression of the boundary specific LATERAL ORGAN BOUNDARIES (LOB) gene and adaxial expressed AS2 gene [115,116]. The bop1-1 mutant shows a similar effect with as1-1 or as2-2 and stm-1 mutants, which suggests that BOP1 promotes or maintains a developmentally determinate state in leaf cells by regulating class 1 KNOX genes [101]. During leaf development, the expression domain of the KNOX1 gene is also regulated by ARP genes and distinguishes the leaf founder cell from the meristem cell in SAM [33,117].
Besides, transcription factors, hormones, and mechanical forces are important for leaf development and morphogenesis. Turgor pressure and cell wall mechanics modulate the direction and rate of cell expansion and affect the pattern of plants. It has been confirmed that the cells in the CZ of SAM have stiffer cell walls compared to the PZ. Organ outgrowth is accompanied by an increase in cell wall elasticity [118]. It has also been shown that auxin regulates cell wall properties, and during leaf initiation, auxin not only reduces stiffness but also affects wall anisotropy through the modulation of the cortical microtubule dynamics [118].
2. Leaf Outgrowth and Expansion
Plants have the ability to grow indeterminately throughout their life and produce many repeated units during their lives. In addition, plants produce certain organs with determinate growth such as leaves, sepals, and petals. In this review, we focused on the size and growth of the Arabidopsis thaliana model plant in which the determined organ (leaf) is regulated by several genetic and environmental factors. The organs of the plants are formed from the reservoir of pluripotent cells, i.e., the meristem. Under the control of cell division and expansion, the leaf primordia achieves its natural size. As the leaf grows, the cell division at the distal end ceases and the expansion process occurs in which the cells at the base are strongly vacuolated rather than proliferated. The base displaces the older cells towards the distal end and eventually, they finally fall out of the proliferation zone. The cells (in a narrow region) are present between the blade and petiole junction, which bidirectionally divides the cells into two parts of the leaf. Furthermore, the dynamic of the cell proliferation regions rapidly appears at the “arrest front” boundary between the proliferation and expansion regions at a constant distance from the base of the leaf for several days before rapidly disappearing [119,120]. Thus, the two main processes of cell proliferation and cell expansion control the final leaf size, whereas any alterations in these processes may affect leaf formation. The timing of the transition from division to expansion within the growth of the leaf lamina is important to determine the final size, shape, flatness, and complexity. The class II TCPs (CINCINNATA-like TCPs) are the key regulators of timing from division to expansion [121,122]. The TCP family consists of plant-specific transcription factors; the CIN-TCPs is the subclass of the TCP family, which has a prominent role in controlling the transition from cell proliferation to expansion during leaf development. During leaf growth, the proliferation phase involves mitotic division interspersed with cytoplasmic growth to increase the primordium size due to the cell number. However, in the expansion phase, the organ increases the size by increasing the cell size [40,123]. There are many genes that positively regulate and control the transition from proliferation to expansion, for example, AINTEGUMENTA (ANT) [124], KLUH/CYP78A5 [125] and GROWTH REGULATING FACTORS (GRFs) [126,127], whereas the negative regulators include CIN-TCPs and DAI [128]. In addition, the PEAPOD1, 2 (PPD1, 2) gene also acts as a negative regulator and changes the dispersed meristematic cells in leaf lamina to stomatal and vascular precursor cells [129]. The promotion of cell proliferation of class I TCPs depends on spatial and temporal expression domains, whereas the CIN-TCP represses cell proliferation (and so the loss of CIN-TCP gene functions increases the proliferation). In Arabidopsis thaliana, there are nine GROWTH REGULATING FACTORS coding genes (GRF1-9), and GRFs delay transition from proliferation to differentiation (Table 1).
TCP4 promotes the expression of miRNA396, which targets seven genes of the GRFs family (Figure 2) [127,130,131]. During the early stage of leaf development, miR396 is expressed in the distal part of the leaf and the expression of GRFs is confined to the proximal regions and promotes cell proliferation. After cell proliferation, miR396 is expressed throughout the organ to decrease GRF expression in the maturing organ [47]. How GRFs stimulate cell proliferation is not fully understood, but GRFs physically interact with GRF-INTERACTING FACTOR1/ANGUSTIFOLIA3 (GIF1/AN3), GIF2, and GIF3 to create a transcriptional module that regulates the leaf size by cell proliferation (Table 1) [41,127,132,133]. Contrary to normal plants, the gif mutant results in smaller and narrower leaves.
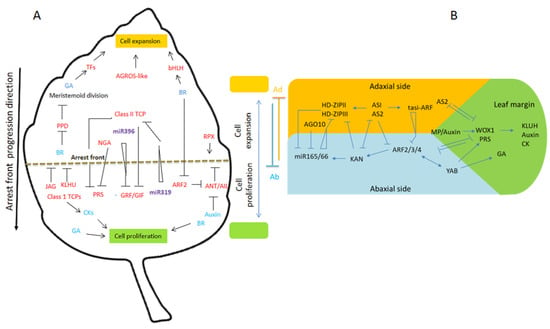
Figure 2.
Regulation of Arabidopsis leaf proliferation and cell expansion transition. (A) the cell cycle arrests the front migration basipetal growth gradient from the apex to the base of the blade petioles, which is a process that is promoted by miR319-TCP modules and repressed by miR393-GRF modules. The gradients of miR319 and miR396 are complementary to TCP and GRF expression. TCP and NGA repressed the blastozone activity. During intercalary growth, the GAs and BRs promote both cell proliferation and expansion (positive and negative regulations are indicated by pointed and T-shaped arrows). Red, genes; blue, hormones; purple, small RNAs. (B) Shortly after its initiation, the young leaf primordium has three regions: adaxial, abaxial, and leaf marginal regions. These regions are determined by the region’s specific transcription factors, such as HD-ZIP III, KANADI, and PRS WOX1, involving multiple negative feedback loop mechanisms. For the sake of clarity, all the interactions are not shown here. Adaxial-abaxial polarity is determined by highly interconnected gene networks. In this network, tasi-ARF limits abaxial determinants and AUXIN RESPONSE FACTORS ARF3/ARF4 to the bottom side. In addition, miR165/166 restricts the expression of adaxial determinants HD-ZIPIII to the top of the leaf (the positive and negative regulations are indicated by pointed and T-shaped arrows, respectively).
GIFs act as a transcriptional coactivator, which is associated with chromatin remodeling machinery. GIF1/AN3 promotes several proliferation-stimulating factors, such as ribosomes, to sustain the high demand of protein that is required in actively proliferating cells and represses the gene that promotes cellular differentiation [41,134]. AN3 lacks a DNA-binding domain, so GRFs help in the recruitment of the AN3-containing chromatin remodeling complex at the promoter region of target genes. During leaf development, the timing of declining GRF-AN3 abundance along the proximo-distal axes link with CIN-TCPs to create the mitotic cycle [40,134]. TCP4 accumulation starts at the tip of the primordium and then covers the whole actively dividing lamina such that TCP activity stops at the tip cells first and is slowly restricted to the base. The miR319 is expressed proximal to the petiole so that miR319-sensitive CIN-TCPs form dynamic spatial gradients and the intensity tapers toward the tips due to inactivation in the distal cell and toward the base [135,136]. In Arabidopsis, the jagged and wavy Dominant (jaw-D) mutant exhibits highly crinkly shaped leaves because of the overexpression of miR319A. The miR319A down-regulates CIN-like TCP genes and their transcript level is dramatically reduced in jaw-D because of the ectopic expression of miR319A (Figure 2) [137,138]. The coordination between the miRNA319-TCP and miR396-GRF modules controls the marginal and overall growth of the leaf through the regulation of cell proliferation [139,140]. The second pathway that controls the leaf lamina outgrowth is the ANT and AINTEGUMENTA LIKE (AIL) gene, which encodes for an AP2/ERF transcription factor. CIN-TCP and ANT have antagonistic effects on the G1/S transition of the cell cycle, and the transcription of CYCLIN-DEPENDENT KINASE INHIBITOR 1 (ICK1) is activated by CIN-TCP that interacts and blocks the activity of G1 cyclins whose transcription is enhanced by ANT [111]. ANT likely functions downstream on auxin, and the auxin-inducible gene ARGOS (an auxin-regulated gene involved in organ size) encodes the ER-localized protein, which is a protein of unknown function. The overexpression or suppression of ARGOS alters the aerial part, such as leaves, flowers, and siliques. The difference in size is mainly due to alterations in cell numbers and the duration of cell proliferation periods. In Arabidopsis, ARGOS-like (ARL) genes, which have some sequence homology to the ARGOS gene, are responsible for the promotion of organ growth via cell expansion. Similarly, the kinase encoded by the TARGET of RAPAMYCIN (TOR) gene is required for cell expansion in the leaves and its overexpression causes an increase in leaf size due to cell expansion [141,142].
ARGOS enhances the expression of AINTEGUMENTA (ANT), another size of the regulator gene, and the change in ANT function has similar effects as ARGOS expression. ANT maintains the expression of D-type cyclin CYCD3;1. The losses in D-type cyclin may cause premature termination of the proliferation phase and change the overall size of the leaf [143]. The ANT also interacts with another family of transcriptional regulators, i.e., ARFs. The ARFs mediate the auxin response and limit the cell size (proliferation) by repressing the ANT and CYCD3;1 activity [144,145].
In addition, when the functions of four NGATHA (NGA) transcription factors are lost, enhanced leaf marginal growth and serration are observed. However, the overexpression of NGAs reduces marginal growth and indicates the redundant roles of NGAs in controlling the switch between leaf marginal expansion and differentiation [140,146]. In Arabidopsis, the transcription factors of NGAs and TCPs terminate the blastozone meristems by inhibiting the expression of the WOX gene [140,147,148]. According to these observations, the leaf blastozone appears by PRESSED FLOWER (PRS) expression and is restricted to the overall marginal regions of young leaves and further restricted to the proximal regions of older leaves [140]. Besides, the genes that encode homeobox transcription factors, WOX genes, PRS/WOX3, and WOX1, redundantly promote leaf blade outgrowth [140,149]. WOX1 and PRS also promote the expression of KLUH, which encodes a cytochrome P450 CYP78A5 monooxygenase that promotes cell proliferation in a non-autonomous manner (Figure 2) [48]. The loss of function mutant kluh produces smaller organs due to the premature arrest of cell proliferation, while the overexpression of KLUH produces larger organs with more cells; and therefore, KLUH promotes organ growth. KLUH is involved in generating mobile factors, and the computational analysis showed that the tissue polarity system specifies the growth patterns in developing leaves [150]. In the early developmental stage, the basic pattern of the growth rate was established across the leaf. Previous studies indicated that leaf meristem activities are controlled by both local regulations and by mobile growth factors that function at the organ level [151].
Besides, in the regulation provided by miRNA-transcription factors, the hormonal control also involved regulating the leaf size and intercalary growth. The most prominent hormones regulating the leaf size are gibberellins (GAs) and brassinosteroids (BRs), which promote leaf growth by cell proliferation and expansion. The GAs increase cell proliferation by repressing cell cycle inhibitors such as KIP-RELATED PROTIEN2 (KRP2) and SIAMESE [152,153]. Overexpression of the BR biosynthesis gene DWARF4 or BR receptor-encoding gene BRASSINOSTEROID INSENSITIVE1 results in larger leaves [128,154]. The BR also represses PPD1 and PPD2, which encodes the transcription factor that limits meristemoid cell proliferation [155]. BRs promote cell expansion via an antagonistic trio of bHLH transcription factors, which are also regulated by GAs, light, and temperature (Figure 2) [156,157].
In the distal region of the leaves, the epidermal and mesophyll cells still have some precursor cells to generate the stomata as well as vascular tissue after cell proliferation ends [158]. These cells can divide the distal region, which is known as dispersed meristematic cells (DMSCs). The DMSCs proliferation is under the control of putative transcription factorPPD1, PPD2. A double mutant of ppd1 and ppd2 produces bell-shaped leaves due to the proliferation of the cell, especially the marginal cells [155].
3. Adaxial/Abaxial Patterning
SAM provides a site for lateral organ formation into the primordium and develops into a flat structure. The lateral organs (leaf) establish polarities along the adaxial/abaxial, mediolateral, and proximodistal axes. The development of adaxial/abaxial patterns requires precise coordination between hundreds of cells throughout primordium development. After the establishment of adaxial/abaxial patterns, they provide a cue for further asymmetric growth. The acquisition and maintenance of adaxial/abaxial polarity are driven by regulatory networks of genes and highly conserved transcription factors [159].
The two sets of transcription factors are expressed at the sides of the leaf, i.e., the upper and bottom that work antagonistically to control the adaxial/abaxial polarities of the leaf. In Arabidopsis thaliana, the members of the HD-ZIP III family (including PHABULOSA (PHB), PHAVOLUTA (PHV), and REVOLUTA (REV)) specify the adaxial cell fate of the leaf (Table 1) [160]. Many transcription factors, i.e., Myb transcription factors PHANTASTICA (PHAN)/AS1 form complexes with the transcription factor AS2 of the LOB to specify the adaxial fate [36,161,162]. The abaxial specification includes three members of the KANADI gene family (KAN1, KAN2, and KAN3) [39,163], four members of the YABBY (YAB) gene family (FILAMENTOUS FLOWER (FIL) [52], YAB3, YAB5, and YAB2)), and two AUXIN RESPONSE FACTORS (ETTIN ETT/ARF3 and ARF4) [53,164]. These regulators, except AS1/PHAN, are specifically expressed in the abaxial or adaxial side and their mutual regulation is very important for proper establishment and maintenance of adaxial-abaxial patterns. In Arabidopsis thaliana, the dominant mutants, phv and phb, form rod-shaped leaves that result in adaxialization of the leaf in the circumference. In fact, the adaxial fate controlling genes, HD-ZIP III, are repressed by miRNA165/166 [165]. Ectopic/constitutive overexpression of miRNA165 and miRNA166 can reduce the transcript level of HD-ZIP III genes [166]. The KANADI gene family requires the abaxial identity of leaves that encode nuclear-localized GARP domain transcription factors [167]. The function of KAN1 disturbs the adaxial/abaxial polarity of the leaf. Ectopic/constitutive expressions of the kan1 mutant with the 35 s promoter produces a narrow cotyledon but no subsequent leaf production [168,169,170]. In fact, PHB is expressed when KAN genes antagonistically regulate HD-ZIP III genes [171]. KAN1 directly represses the expression of AS2, and AS2 indirectly represses the expression of KAN1. Furthermore, AS2 also negatively regulates ETT, KAN2, and YAB5. Likewise, the opposing effect of KAN and HD-ZIP III on the auxin biosynthesis gene has been confirmed by many researchers. A recent study corroborated the finding that the KAN1 allele is responsible for suppressing the expression of PIN1. The ectopic expression of KAN1 reduces the gene expression of pro-cambium cells and PIN1 in pro-vascular cells. APUM23 is a new regulator of leaf polarity, which encodes approximately 20 PUF RNA-binding proteins in Arabidopsis thaliana and interacts with leaf polarity that is required for the maintenance of genes [172,173]. Another gene family, which specifies the abaxial cell fate, is the ARFs, which binds to the promoter element of the auxin response gene and transducer auxin signaling, and their role in adaxial/abaxial polarity is confirmed through mutation and up-regulation patterns of ETT (ARF3) and ARF4 [174,175]. The polarity defects were visualized in ett-1, arf4-1, and ett-1, and art4-2 double mutants. In these mutants, the produced abaxialized leaves were similar to kan1 and kan2 mutants. The direct interaction between KAN and ARF indicates that overlapping patterns control the polarity of the leaf. Both the ETT and ARF4, as a target of TAS3, are derived from trans-acting siRNAs (ta-siRNAs) that up-regulate small interfering RNA (siRNA) [98,176]. According to this experiment, ta-siRNA insensitive ETT or ETT overexpression in the rdr6-15 mutant background showed a defect in leaf morphology. These observations specify TAS3 ta-siRNAs as a negative regulator of abaxial cell fate through targeting of ETT and ARF4 gene expression in small RNAs [98,177].
The adaxial and abaxial pattern is important for lamina outgrowth and started from the boundary of adaxial/abaxial to the medial/lateral axis. The leaf primordia represent another meristematic zone called the plate meristem or blastzone. Many genes are involved in the medial/lateral specification, but the most important is the YABBY gene family, which encodes for proteins with a zinc finger and a helix-loop-helix domain, and also plays a key role in lamina outgrowth. The Arabidopsis genome contains six YABBY genes (FIL, YABBY2 (YAB2), YAB3, YAB5, CRAB CLAW (CRC) and INNER NO OUTER (INO)) [178,179], and four of these are expressed in the vegetative primordium and (FIL, YAB2, YAB3, and YAB5) and the other two are expressed in floral organs. Consistently, the YABBY expression is localized between the adaxial and abaxial side at the leaf margin and promotes lamina outgrowth. Interestingly, the YABBY gene expression is regulated by the members of the polarity pathway such as the HD-ZIP III, AS, and KANADI pathways [180,181,182,183]. In the kan1 and kan2 double mutants, the YABBY gene is required for ectopic outgrowth. Therefore, for lamina outgrowth, YABBY gene function is integrated with polarity signals [184]. The YABBY gene is important for repression of the genes of SAM in developing leaves and promoting maturation of the leaves. In the double mutants of fil and yab3, class 1 KNOX genes are ectopically expressed [185,186,187]. Four YABBY gene mutants expressed in the leaf primordium show narrow leaves but only limited defects on leaf polarity.
For the WOX genes, at least two subfamily members are essential for lamina outgrowths such as WOX1 and PRS/WOX3. In Arabidopsis, PRS is expressed in the margins of the developing leaf primordium; the prs mutant causes the deletion of stipules at the base of leaf margins without a reduction in leaf width and due to the genetic redundancy with WOX1 [188,189]. The WOX1 gene is expressed along with the adaxial-abaxial juxtaposition and overlaps with PRS at the marginal region of the leaf. The double mutant of wox1/prs showed prominent defects in the lamina outgrowth and redundantly acted to enhance the leaf width. It has been identified that WOX1 expression occurs around the meristem at the boundary between the HD-ZIP III and KAN expression domains. In the kan1 and kan2 double mutants, the expression of WOX1/PRS is enhanced in the abaxial domains of the leaf, which suggest that the KAN gene may function to negatively regulate the expression of WOX1/PRS [190]. Therefore, misexpression of WOX1/PRS may explain the ectopic formation of abaxial margin-like outgrowth that occurs in the kan mutants. The expression of WOX1/PRS is negatively regulated by AS2 in the adaxial domain of the leaf. AS2 expression is also repressed by WOX1/PRS to restrict its expression on the adaxial side of the leaf (Figure 2). Meanwhile, abaxial-specific gene expression is also influenced by WOX1/PRS and these interactions help to restrict WOX1/PRS expression toward the margin domain of the leaf [190,191].
In the marginal regions of the leaf, WOX1/PRS is expressed and enables flattening of the leaf. MP and auxin act together as a positional cue for patterning the WOX1/PRS marginal regions [179]. Abaxial factors, such as KAN, restrict the marginal domain expansion [48], which binds to the same elements as an MP in the WOX1 gene promoter and inhibits their expression [167]. During leaf development, the auxin maxima are first formed at the tips of young primordia and promote distal growth. Previous evidence suggests that auxin works downstream of the leaf polarity genes and enhances lamina outgrowth. In the triple mutant of kan1, kan2 and kan3, there was enhanced lamina outgrowth on the hypocotyl due to ectopic localization of PIN1 proteins [142]. The distribution of auxin and PIN1 is greatly influenced by yabby mutants. The as1 and as2 mutant showed asymmetrical lamina outgrowth due to the asymmetric distribution of auxin. Recent studies revealed that the YUCCA (YUC) gene, which encodes catalyzation, is involved in auxin biosynthesis and plays an important role in lamina outgrowth and leaf margin formation. In Arabidopsis, there are 11 YUC genes that have been identified; the mutant of at least four genes led to loss of marginal characters. Interestingly, YUC gene expression is enhanced in ectopic lamina outgrowth, which is due to ectopic adaxial and abaxial juxtaposition in the as2 rev and kan1 kan2 double mutants [192,193].
4. Leaf Margins Serrations
The leaf is developed from the peripheral zone of the SAM under the control of many intrinsic and extrinsic factors. The leaves are different in size and form, which is a complicated process. The various forms of leaf margins include entire, serrate, and lobe margins. The molecular mechanism of the Arabidopsis leaf indicates that the leaf serrations get more pronounced during plant development. Additionally, miRNA164A, CUC2, PIN1, and DPA4 regulate the leaf serration [45,46,194]. In Arabidopsis thaliana, leaf serration is less prominent in rosette leaves compared to leaves that develop later. Leaf serration is also different in different accessions of Arabidopsis thaliana. The optimal expression of the genes is necessary for continuous growth and development. The miRNAs mainly contribute to the regulation of gene expression [45,195]. In leaf serration, the boundary controlling gene CUC1, CUC2, and CUC3 is required for the maintenance of the shoot apical meristem [45,195]. Furthermore, reports indicate that CUC2 has a very important role in leaf serration. The members of the NAC transcription factors can be expressed in the boundary region and suppress growth. A mutation in this gene at an early stage produces a cup-shaped fused cotyledon. The CUC1 and CUC2 gene expression are controlled by miRNA164A [45,46,196]. The ectopic expression of miRNA164A represses CUC2 gene expression; therefore, the leaf is less serrated (unlike a wild type). The mutations in the mir164a produce deep serration and contrast with the overexpression of miR164A, which causes smooth leaf margins [45]. In overexpression, the CUC2 expression is high and plant leaves are highly serrated (Figure 3). CUC2 promotes PIN1 efflux auxin carriers [4]. The mutant of the pin1 genotypes causes inhibition of the auxin efflux carrier and the leaf has smooth margins [65,197]. The other plant hormones, such as cytokinin (CK), gibberellins (GA), and many others, also have an important role in organogenesis depending on its concentration [198]. JAGGED LATERAL ORGANS (JLO) is a member of LATERAL ORGAN BOUNDARY DOMAIN gene family and is transiently expressed at the site of organ initiation [199,200], promotes the PIN expression for auxin maxima, and resolves in the leaf-meristem boundary during outgrowth [103,200,201].
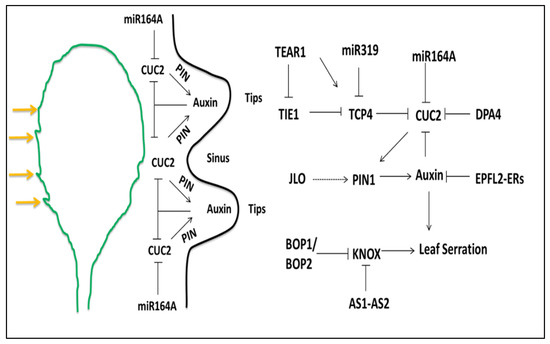
Figure 3.
Schematic depiction of the factors involved in the modification of leaf margins, the genetic interaction between different genes and hormones cause the formation of leaf serration. The CUC2 gene promotes the auxin maxima via PIN1 efflux protein in the epidermal cell. The high concentrations of auxin at the tips block, the expression of CUC2 and thus, the expression of CUC2 gene is restricted to the sinus region of the leaf; miR164A also suppresses the CUC2 expression. The other regulatory factors shown in the figure is discussed in detail in the text (the positive and negative regulations are indicated by pointed and T-shaped arrows, respectively).
In addition, the cin-tcp mutant displays prolonged marginal growth leading to the formation of the lobe and crinkly leaves due to ectopic expression of boundary specific CUC2 genes and meristem-specific KNOX genes [196,202]. CIN-TCPs repress CUC activity through miR164 that targets CUC2. Additionally, TCP4 interacts with CUC2 and CUC3 to prevent their dimerization and transactivation potential in the juvenile stage to inhibit leaf serration [196]. However, in the later stage, the SPL transcription factor destabilized the interaction of TCP-CUC from relieving CUC protein from inhibition. Therefore, the spl mutant loss of function and CIN-TCP gain of function both have the same reduced organ size, which suggests that SPL interferes with CIN-TCP in growth repression [203,204].
The DEVELOPMENT-RELATED PcG TARGET IN THE APEX4 (DPA4) negatively regulates the expression of CUC2 independent of MIR164A and modulates leaf serration [151]. The TCP interactor containing EAR motif protein1 (TIE1) is a transcriptional regulator located in the nucleus. TIE1 recruits co-repressor TOPLESS (TPL)/TOPLESS-RELATED (TPR) in leaf margin morphology development and inhibits the activity of TCP at the protein level. Therefore, over-expression of TIE1 can cause an increase in leaf serration [151,205]. TIE1 and TEAR1 (TIE1-ASSOCIATED RING-TYPE E3 LIGASE1) have a mutual role; a mutation in TEAR1 and its homologous genes increases leaf serration [206]. TIE1 recruits TPL/TPRs to inhibit TCP activity, and TEAR1 restricts the inhibition of TCP through the degradation of TIE1. TIE1 and TEAR1 indirectly affect the leaf margin by regulating TCP (Figure 3).
A peptide in the plant called EPFL2 (Epidermal Patterning Factor-Like) and members of the ERECTA (ER) family are also involved in the morphogenesis of leaf marginal serrations [207]. When the EPFL2 or ER is mutated in Arabidopsis, the leaves become smooth and the auxin is detected throughout the leaf margins. EPFL2 forms a ligand-receptor pair with ERECTA and thereby inhibits the response of auxin at the leaf tooth area, which in turn inhibits the expression of EPFL2 and forms a negative feedback loop [207]. This feedback system maintains the auxin response pattern during leaf margin growth. In addition, current studies have shown that JAGGED (JAG), JLO, and Trifoliate can also regulate leaf margin morphogenesis through the auxin pathway and also affect the KNOX regulatory pathway (Figure 3) [199].
BLADE ON PETIOLE1 (BOP1) and BOP2 belong to BTB family proteins, which form dimmers that function as transcriptional activators. BOP1 and BOP2 are expressed at the base of lateral organs, and BOP expression at the base of the leaves can directly regulate AS2 and inhibit the expression of the KNOX gene [116]. The leaves of bop1 and bop2 double protrusions become larger, and leaf teeth and leaf fins grow at the petioles [208,209]. Therefore, BOP ensures normal leaf morphogenesis by inhibiting KNOX expression at the leaf base and petiole [153,210,211]. High auxin accumulation causes more cell division and growth, and consequently, the teeth regions of the leaf have high auxin maxima [102,212,213]. Auxin represses the expression of the CUC2 gene, and therefore, CUC2 expression is restricted to the sinus region of the leaf [45,46]. CUC2 also suppresses the growth of cells in the sinus region and thereby promotes tip outgrowth. The exact mechanism of the CUC2 and auxin interaction is still not fully understood.
5. Developmental Functions of Micrornas
MicroRNAs are a group of non-coding RNAs that play an important role in diverse cellular pathways and regulate most of the plant and animal biological processes [214]. The transcription and maturation of microRNAs involve a series of complex processes. In the first phase, endogenous genes are transcribed by Pol II or III into long primary miRNAs that consist of several hundred nucleotides. Afterward, single strand pri-miRNAs are folded to form a hairpin-like secondary structure [215]. Pri-miRNAs are processed by endonuclease RNAase III and the mechanism is different in plants and animals [216,217]. Plants lack Drosha homologs and after the pri-miRNAs formation, the RNase III enzyme DICER-LIKE1 (DCL1) regulates the first and second steps (Figure 4). In contrast, in animals, the first step involves Drosha, which cuts miRNAs strands, and in the second step, the pre-miRNAs are processed through Dicer with the aid of HYL1 and SE to form a duplex in the nucleus. The mature microRNAs duplex consists of active and complementary strands. The active strands are called guide strands while the complementary strands are called passenger strands. The guide strands with lower thermodynamic stability and high abundance are loaded into ARGONAUTE (AGO)-associated RNA-induced silencing complexes (RISCs) and target the mRNA transcript [218,219]. The passenger strands of miRNAs become degraded, and the accumulation of passenger strands is lowered by guide strands [220,221]. Many studies have confirmed that miRNAs act as regulatory factors in a large number of biological processes and consist of different numbers of miRNAs in a single species [222,223]. The miRNA biogenesis mutants (dcl1, hyll, se, and hen1) and ago1 mutants produce developmental defects [224]. In this review, we described some of the miRNAs and their function in different stages of plant development, including phase transitions, hormone biosynthesis and signaling, pattern formation, and morphogenesis. During leaf development, different types of miRNAs have a prominent and important role that modulates leaf development in different phases such as establishments, transitions, modifications, and senescence. Transcriptome profiling data showed that several miRNAs are involved in early embryonic development such as miR156, miR166, miR167, miR390, and miR394 [225]. The interaction between two miRNAs affects the maintenance of the meristem and leaf initiation, such as the interplay of LEAF CURLING RESPONSIVENESS (LCR) and the miR394 mediated non-cell-autonomous network, and the module of miR160 and miR165/66 mediated cell-autonomous pathways [165,226]. The protoderm-specific miR394 confers stem cell maintenance by repressing the gene expression of LEAF CURLING RESPONSIVENESS (LCR), which also regulates a local feedback loop mediated by WUS and CLV genes [226].
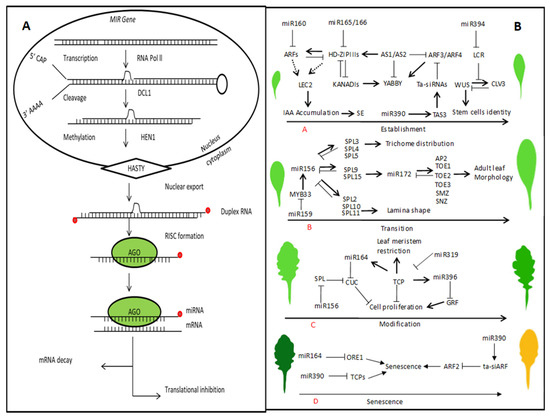
Figure 4.
A schematic pathway for miRNA biosynthesis and the degradation of target mRNA. (A) Primary miRNAs are formed by polymerase II that folds back to form a hairpin structure. Splicing and further processing in the nucleus include the interactive role of different proteins and the CAP-binding proteins CBP20 and CBP80. Further processing occurs by DCL1 and forms miRNA-miRNA* duplexes, which are methylated by HEN1 and transported to the cytoplasm. The mature miRNA is unwound to yield a 22-nt single-strand and incorporated into AGO1-containing RISC (RNA-induced silencing complex) and other regulatory proteins that directly inhibit the translation or cleavage of the target mRNA transcript. (B) Leaf development is a complex biological process with multiple regulatory networks regulated by miRNA and their target genes. The leaf development consists of different stages such as establishment, transition, modification, and senescence. The miRNAs play important roles in each stage. The miR160 and miR165/166 control leaf initiation; leaf polarity is determining by miR165/166 and miR390; leaf morphology is regulating by miR164 and miR319; phase initiation is determined by miR156 and miR172; leaf senescence is determined by miR164 and miR319 (the arrows indicate positive and T-shaped indicate negative regulation).
Small RNAs, such as miR169, miR167, and tasiR-ARFs, target the ARF genes, which play an important role in auxin signaling. During shoot and root development, miR160 regulates the expression of ARF10, ARF16, and ARF17 [227]. However, overexpression of miR160 or miR160-resistance ARFs leads to pleiotropic and developmental defects in all aerial organs [228,229,230]. Besides, miR167 targets ARF6 and ARF8, which redundantly regulates ovule and anther development [231]. During the establishment of leaf boundaries, both miR165/166 and miR390 move between cells and interact with each other. In both Arabidopsis and maize, miR165/166 targets homeodomain-leucine zipper transcription factor genes to establish the abaxial identity of lateral organs. The abundantly produced miR165/166 in the abaxial side of the leaf targets the mRNAs in the adaxial side where these genes specify the adaxial characteristics (Figure 2) [232]. In Arabidopsis, miRNA resistant varieties of PHB and REV genes lead to adaxializtion of the leaves. The tasi-RNAs from the TAS3 locus target ARF3 and ARF4, which results in the abaxial differentiation of lateral organs, vegetative traits, and leaves [39,233,234]. The tasiR-ARFs repress the expression of ARF3 and ARF4, which contributes to adaxial specification in different species (Table 2). However, mutations in ta-siRNA biosynthesis do not exhibit any prominent change in leaf polarity, which is most likely due to the existence of a parallel mechanism to control adaxial polarity [182,235]. To develop a leaf bladeless (lbl) mutant in maize, a homolog of Arabidopsis SGS3 requires ta-siRNA biogenesis that promotes the abaxialization of leaves. The two small RNAs, miR165/166, and tasiR-ARF are expressed in the opposite side of the leaf and establish the abaxial-adaxial axis in leaf development [236,237]. In addition, during further modification, divergent leaf growth polarity is strongly correlated with the miR396-GRF expression gradient. The miR164 regulates the expression of CUC1 and CUC2 transcription factors, which are very important for the proper establishment of organ boundaries, floral patterning, and leaf morphogenesis throughout plant development [45,238].
Table 2.
MicroRNAs and their predicted targets in Arabidopsis.
The most conserved miR156 found in all land plants in Arabidopsis, miR156/157 targets 10 of the SPL TFs, which promote vegetative phase changes and floral transition [263]. SPL3/4/5 determines trichome formation and distribution, SPL9/15 modulates adult leaf morphology, and SPL2/10/11 regulates the lamina shape and acts independently of miR172 (Figure 4) [264]. Overexpression of miR156 prolongs the juvenile stage and extremely delays the flowering stages. In Arabidopsis, miR159 targets MYB33, MYB65, and MYB101 genes that activate gibberellin-responsive genes in the aleurone layer during germination. The overexpression of miR159 is responsible for the delays in flowering [165]. In Arabidopsis, miR172 regulates six of the AP2-domain transcription factors, including [240], TARGET OF EAT1, TOE2, TOE3, SCHLAFMÜTZE (SMZ), SCHNARCHZAPFEN (SNZ), and promotes flowering by repressing TOE1 and TOE2 [264,265]. The miR156 and miR172 act in a linear pathway and coordinate vegetative and floral transition (Table 2) [264]. In a petunia hybrid and Antirrhinum majus, miR169 controls the spatial restriction of the homeotic class C genes that are required for the identities of reproductive organs in the flower [251,266]. In an ectopic expression, miR169 transforms petals into stamens and targets the NF-YA genes that are activators of class C gene expression. In Arabidopsis, miR169 restricts the class C gene expression to delegate the transcription factor gene of APETALA2 (AP2) [253]. In Arabidopsis, miR172 targets the AP2 mRNA, which is uniformly present in all four floral whorls, unlike other floral homeotic genes that are confined to two whorls [253,254]. However, miR172 mediates regulation of AP2 at the translational level and that is why AP2 protein is more concentrated in the outer two floral whorls [215,267]. In Arabidopsis, miR319 targets five TCP genes that mostly control cell division during leaf development [125,268].
Senescence is the last stage of leaf morphogenesis that involves many distinctive actions [103,269]. In Arabidopsis, miR164 and miR319 modulate aging-induced cell death and leaf senescence. The miR164 target ORESAR1 (ORE1) functions as a positive regulator of senescence. The central components of ethylene signaling ETHYLENE INSENSITIVE2 and 3 (EIN2/3) induce ORE1 in an age-dependent manner [270]. During the early stage of leaf development, miR164 down-regulates ORE1 expression but also up-regulates at a later stage by EIN2/3. In addition to ethylene, other plant hormones, such as auxin and JA, also play important roles in controlling plant senescence. ARF2, a negative regulator of auxin responses, is believed to regulate leaf longevity [269,271]. The miR393 targets TIR1, which are the auxin receptor and F-box genes that play an important role in auxin biosynthesis [259]. Furthermore, miR319 targets the TCP4 genes [136,272], and TCP4 has been shown to activate jasmonic acid (JA) biosynthesis gene LIPOXYGENASE2, which increases the level of JA and accelerates leaf senescence [273,274].
6. Conclusions and Future Perspectives
The developmental transition during shoot development in plants is regulated by factors that originate outside and within the shoot apical meristem (SAM). Lateral organ initiation at the shoot apical meristem involves a complex mechanism of hormones and downstream transcriptional regulation that leads to the formation of different organs such as the leaf, shoot, and flower. From vegetative to reproductive transition, the leaf-derived external signal and internal factors that cause the vegetative phase change and that are spatially coordinated remain unclear. This review elucidates the interaction of different factors (genes, miRNAs, and hormones) that maintain the STM cells’ identity. For example, the homeobox gene WUS is expressed at the organizing center (OC) and regulates the boundaries of the STM cell niches. These factors not only maintain the stem cell identity but also help in the initiation, growth, and adaxial/abaxial patterning of leaf development. It has also been suggested that many small RNAs help in early leaf development because of their ability to clear out the transcript when the cell passes from one stage to another. Subtle and complex mechanisms such as leaf development require many levels of control, for instance, buffering and plasticity of the small RNAs is one example of how plants achieve this control. Elaborate signaling and effector networks are also involved in leaf development and further study of these aspects is warranted.
Author Contributions
Conceived of the presented idea, S.A., and N.K.; developed the theory and performed the computations, S.A., and N.K.; Resources, S.A., N.K., and L.X.; Writing—original draft, S.A., and N.K.; Writing—review & editing, S.A., N.K., and L.X. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by project 3180101344, National Natural Science Foundation of China.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Cho, K.H.; Jun, S.E.; Jeong, S.J.; Lee, Y.K.; Kim, G.T. Developmental processes of leaf morphogenesis in arabidopsis. J. Plant Biol. 2007, 50, 282–290. [Google Scholar] [CrossRef]
- Du, F.; Guan, C.; Jiao, Y. Molecular mechanisms of leaf morphogenesis. Molec. Plant 2018, 11, 1117–1134. [Google Scholar] [CrossRef] [PubMed]
- Li, L.C.; Kang, D.M.; Chen, Z.L.; Qu, L.J. Hormonal regulation of leaf morphogenesis in Arabidopsis. J. Integ. Plant Biol. 2007, 49, 75–80. [Google Scholar] [CrossRef]
- Efroni, I.; Eshed, Y.; Lifschitz, E. Morphogenesis of simple and compound leaves: A critical review. Plant Cell 2010, 22, 1019–1032. [Google Scholar] [CrossRef]
- Jiao, K.; Li, X.; Su, S.; Guo, W.; Guo, Y.; Hu, Z.; Shen, Z.; Luo, D. Genetic control of compound leaf development in the mungbean (Vigna radiata L.). Hortic. Res. 2019, 6, 23. [Google Scholar] [CrossRef]
- Barton, M.K. Cell type specifi cation and selfrenewal in the vegetative shoot apical meristem. Curr. Opin. Plant Biol. 1998, 1, 37–42. [Google Scholar] [CrossRef]
- Itoh, J.I.; Kitano, H.; Matsuoka, M.; Nagato, Y. SHOOT ORGANIZATION genes regulate shoot apical meristem organization and the pattern of leaf primordium initiation in rice. Plant Cell 2000, 12, 2161–2174. [Google Scholar] [CrossRef]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.; Jürgens, G.; Laux, T. The stem cell population of Arabidopsis shoot meristems in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 2000, 100, 635–644. [Google Scholar] [CrossRef]
- Deb, Y.; Marti, D.; Frenz, M.; Kuhlemeier, C.; Reinhardt, D. Phyllotaxis involves auxin drainage through leaf primordia. Development 2015, 142, 1992–2001. [Google Scholar] [CrossRef]
- Bhatia, N.; Heisler, M.G. Self-organizing periodicity in development: Organ positioning in plants. Development 2018, 145, dev149336. [Google Scholar] [CrossRef]
- Yonekura, T.; Iwamoto, A.; Fujita, H.; Sugiyama, M. Mathematical model studies of the comprehensive generation of major and minor phyllotactic patterns in plants with a predominant focus on orixate phyllotaxis. PLoS Comput. Biol. 2019, 15, e1007044. [Google Scholar] [CrossRef]
- Traas, J. Phyllotaxis. Development 2013, 140, 249–253. [Google Scholar] [CrossRef]
- Barton, M.K. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev. Biol. 2010, 341, 95–113. [Google Scholar] [CrossRef]
- Kucypera, K.; Lipowczan, M.; Piekarska-Stachowiak, A.; Nakielski, J. A method to generate the surface cell layer of the 3D virtual shoot apex from apical initials. Plant Methods 2017, 13, 110. [Google Scholar] [CrossRef]
- Tax, F.E.; Durbak, A. Meristems in the movies: Live imaging as a tool for decoding intercellular signaling in shoot apical meristems. Plant Cell 2006, 18, 1331–1337. [Google Scholar] [CrossRef]
- Adibi, M.; Yoshida, S.; Weijers, D.; Fleck, C. Centering the organizing center in the Arabidopsis thaliana shoot apical meristem by a combination of cytokinin signaling and self-organization. PLoS ONE 2016, 11, e0147830. [Google Scholar] [CrossRef]
- Bustamante, M.; Matus, J.T.; Riechmann, J.L. Genome-wide analyses for dissecting gene regulatory networks in the shoot apical meristem. J. Exp. Bot. 2016, 67, 1639–1648. [Google Scholar] [CrossRef]
- Fouracre, J.P.; Poethig, R.S. Role for the shoot apical meristem in the specification of juvenile leaf identity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 10168–10177. [Google Scholar] [CrossRef]
- Rinne, P.L.; Kaikuranta, P.M.; Van Der Schoot, C. The shoot apical meristem restores its symplasmic organization during chilling-induced release from dormancy. Plant J. 2001, 26, 249–264. [Google Scholar] [CrossRef]
- Sager, R.; Lee, J.Y. Plasmodesmata in integrated cell signalling: Insights from development and environmental signals and stresses. J. Exp. Bot. 2014, 65, 6337–6358. [Google Scholar] [CrossRef]
- Tian, C.; Wang, Y.; Yu, H.; He, J.; Wang, J.; Shi, B.; Jiao, Y. A gene expression map of shoot domains reveals regulatory mechanisms. Nat. Commum. 2019, 10, 141. [Google Scholar] [CrossRef]
- Reddy, G.V.; Meyerowitz, E.M. Stem-cell homeostasis and growth dynamics can be uncoupled in the Arabidopsis shoot apex. Science 2005, 310, 663–667. [Google Scholar] [CrossRef]
- Yadav, R.K.; Girke, T.; Pasala, S.; Xie, M.; Reddy, G.V. Gene expression map of the Arabidopsis shoot apical meristem stem cell niche. Proc. Natl. Acad. Sci. USA 2009, 106, 4941–4946. [Google Scholar] [CrossRef]
- Shi, B.; Vernoux, T. Patterning at the shoot apical meristem and phyllotaxis. Curr. Top. Dev. Biol. 2019, 131, 81–107. [Google Scholar]
- Ha, C.M.; Jun, J.H.; Fletcher, J.C. Shoot apical meristem form and function. Curr. Top. Dev. Biol. 2010, 91, 103–140. [Google Scholar]
- Vernoux, T.; Besnard, F.; Traas, J. Auxin at the shoot apical meristem. Cold Spring Harb. 2010, 2, a001487. [Google Scholar] [CrossRef]
- Vernoux, T.; Brunoud, G.; Farcot, E.; Morin, V.; Van den Daele, H.; Legrand, J.; Guédon, Y. The auxin signalling network translates dynamic input into robust patterning at the shoot apex. Mol. Syst. Biol. 2011, 7, 508. [Google Scholar] [CrossRef]
- Souer, E.; Van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The No Apical Meristem gene of petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef]
- Hibara, K.I.; Karim, M.R.; Takada, S.; Taoka, K.I.; Furutani, M.; Aida, M.; Tasaka, M. Arabidopsis CUP-SHAPED COTYLEDON3 regulates postembryonic shoot meristem and organ boundary formation. Plant Cell 2006, 18, 2946–2957. [Google Scholar] [CrossRef]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Scofield, S.; Dewitte, W.; Nieuwland, J.; Murray, J.A. The A rabidopsis homeobox gene SHOOT MERISTEMLESS has cellular and meristem-organisational roles with differential requirements for cytokinin and CYCD3 activity. Plant J. 2013, 75, 53–66. [Google Scholar] [CrossRef]
- Clark, S.E.; Jacobsen, S.E.; Levin, J.Z.; Meyerowitz, E.M. The CLAVATA and SHOOT MERISTEMLESS loci competitively regulate meristem activity in Arabidopsis. Development 1996, 122, 1565–1575. [Google Scholar]
- Timmermans, M.C.; Hudson, A.; Becraft, P.W.; Nelson, T. ROUGH SHEATH2: A Myb protein that represses knox homeobox genes in maize lateral organ primordia. Science 1999, 284, 151–153. [Google Scholar] [CrossRef]
- Byrne, M.E.; Barley, R.; Curtis, M.; Arroyo, J.M.; Dunham, M.; Hudson, A.; Martienssen, R.A. Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Nature 2000, 408, 967–971. [Google Scholar] [CrossRef]
- Simmons, A.R.; Bergmann, D.C. Transcriptional control of cell fate in the stomatal lineage. Curr. Opin. Plant Biol. 2016, 29, 1–8. [Google Scholar] [CrossRef]
- Sun, X.; Wang, C.; Xiang, N.; Li, X.; Yang, S.; Du, J.; Yang, Y.; Yang, Y. Activation of secondary cell wall biosynthesis by miR319-targeted TCP4 transcription factor. Plant Biotechnol. J. 2017, 15, 1284–1294. [Google Scholar] [CrossRef]
- Lu, Y.; Feng, Z.; Liu, X.; Bian, L.; Xie, H.; Zhang, C.; Liang, J. MiR393 and miR390 synergistically regulate lateral root growth in rice under different conditions. BMC Plant Biol. 2018, 18, 261. [Google Scholar] [CrossRef]
- Iwakawa, H.; Iwasaki, M.; Kojima, S.; Ueno, Y.; Soma, T.; Tanaka, H.; Machida, C. Expression of the ASYMMETRIC LEAVES2 gene in the adaxial domain of Arabidopsis leaves represses cell proliferation in this domain and is critical for the development of properly expanded leaves. Plant J. 2007, 51, 173–184. [Google Scholar] [CrossRef]
- Martin-Trillo, M.; Cubas, P. TCP genes: A family snapshot ten years later. Trends Plant Sci. 2010, 15, 31–39. [Google Scholar] [CrossRef]
- Yuan, S.; Zhao, J.; Li, Z.; Hu, Q.; Yuan, N.; Zhou, M.; Luo, H. MicroRNA396-mediated alteration in plant development and salinity stress response in creeping bentgrass. Hortic. Res. 2019, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yan, A.; Han, H.; Li, T.; Geng, Y.; Liu, X.; Meyerowitz, E.M. HAIRY MERISTEM with WUSCHEL confines CLAVATA3 expression to the outer apical meristem layers. Science (N.Y.) 2018, 361, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Brand, U.; Fletcher, J.C.; Hobe, M.; Meyerowitz, E.M.; Simon, R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 2000, 289, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Fan, Z.; Shan, W.; Yin, X.R.; Kuang, J.F.; Lu, W.J.; Chen, J. Association of BrERF72 with methyl jasmonate-induced leaf senescence of Chinese flowering cabbage through activating JA biosynthesis-related genes. Hortic. Res. 2018, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, B.; Wang, H.; Li, J.; Huang, H.; Xu, L. YUCCA genes are expressed in response to leaf adaxial–abaxial juxtaposition and are required for leaf margin development. Plant Physiol. 2011, 157, 1805–1819. [Google Scholar] [CrossRef]
- Heisler, M.G.; Ohno, C.; Das, P.; Sieber, P.; Reddy, G.V.; Long, J.A.; Meyerowitz, E.M. Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr. Biol. 2005, 15, 1899–1911. [Google Scholar] [CrossRef]
- White, D.W. PEAPOD regulates lamina size and curvature in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 13238–13243. [Google Scholar] [CrossRef]
- Ikeda, M.; Ohme-Takagi, M. TCPs, WUSs, and WINDs: Families of transcription factors that regulate shoot meristem formation, stem cell maintenance, and somatic cell differentiation. Front Plant Sci. 2014, 5, 427. [Google Scholar] [CrossRef]
- Tsiantis, M.; Schneeberger, R.; Golz, J.F.; Freeling, M.; Langdale, J.A. The maize rough sheath2 gene and leaf development programs in monocot and dicot plants. Science 1999, 284, 154–156. [Google Scholar] [CrossRef]
- Kerstetter, R.A.; Laudencia-Chingcuanco, D.; Smith, L.G.; Hake, S. Loss-of-function mutations in the maize homeobox gene, knotted1, are defective in shoot meristem maintenance. Development 1997, 124, 3045–3054. [Google Scholar]
- Murray, J.A.; Jones, A.; Godin, C.; Traas, J. Systems analysis of shoot apical meristem growth and development: Integrating hormonal and mechanical signaling. Plant Cell 2012, 24, 3907–3919. [Google Scholar] [CrossRef] [PubMed]
- Kuchen, E.E.; Fox, S.; De Reuille, P.B.; Kennaway, R.; Bensmihen, S.; Avondo, J.; Coen, E. Generation of leaf shape through early patterns of growth and tissue polarity. Science 2012, 335, 1092–1096. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yong, X.; Yu, J.; Cheng, T.; Wang, J.; Yang, W.; Zhang, Q. Identification of candidate adaxial–abaxial related genes regulating petal expansion during flower opening in Rosa chinensis ‘Old Blush’. Front. Plant Sci. 2019, 10, 1098. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Thomas, J.; Collins, G.; Timmermans, M.C. Direct repression of KNOX loci by the ASYMMETRIC LEAVES1 complex of Arabidopsis. Plant Cell 2008, 20, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Hay, A.; Barkoulas, M.; Tsiantis, M. ASYMMETRIC LEAVES1 and auxin activities converge to repress BREVIPEDICELLUS expression and promote leaf development in Arabidopsis. Development 2006, 133, 3955–3961. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.F.; Schoof, H.; Haecker, A.; Lenhard, M.; Jürgens, G.; Laux, T. Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 1998, 95, 805–815. [Google Scholar] [CrossRef]
- Meng, Y.; Liu, H.; Wang, H.; Liu, Y.; Zhu, B.; Wang, Z.; Hou, Y.; Zhang, P.; Wen, J.; Yang, H.; et al. HEADLESS, a WUSCHEL homolog, uncovers novel aspects of shoot meristem regulation and leaf blade development in Medicago truncatula. J. Exp. Bot. 2019, 70, 149–163. [Google Scholar] [CrossRef]
- Laux, T.; Mayer, K.F.; Berger, J.; Jurgens, G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar]
- Busch, W.; Miotk, A.; Ariel, F.D.; Zhao, Z.; Forner, J.; Daum, G.; Suzaki, T.; Schuster, C.; Schultheiss, S.J.; Leibfried, A.; et al. Transcriptional control of a plant stem cell niche. Dev. Cell. 2010, 18, 841–853. [Google Scholar] [CrossRef]
- Ikeda, M.; Mitsuda, N.; Ohme-Takagi, M. Arabidopsis WUSCHEL is a bifunctional transcription factor that acts as a repressor in stem cell regulation and as an activator in floral patterning. Plant Cell 2009, 21, 3493–3505. [Google Scholar] [CrossRef]
- Leibfried, A.; To, J.P.; Busch, W.; Stehling, S.; Kehle, A.; Demar, M.; Kieber, J.J.; Lohmann, J.U. WUSCHEL controls meristem function by direct regulation of cytokinin-inducible response regulators. Nature 2005, 438, 1172–1175. [Google Scholar] [CrossRef]
- Xie, M.; Chen, H.; Huang, L.; O’Neil, R.C.; Shokhirev, M.N.; Ecker, J.R. A B-ARR-mediated cytokinin transcriptional network directs hormone cross-regulation and shoot development. Nat Commun. 2018, 9, 1604. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.; Honma, T.; Aoyama, T.; Sato, S.; Kato, T.; Tabata, S.; Oka, A. ARR1, a transcription factor for genes immediately responsive to cytokinins. Science 2001, 294, 1519–1521. [Google Scholar] [CrossRef] [PubMed]
- Besnard, F.; Refahi, Y.; Morin, V.; Marteaux, B.; Brunoud, G.; Chambrier, P.; Rozier, F.; Mirabet, V.; Legrand, J.; Lainé, S.; et al. Cytokinin signalling inhibitory fields provide robustness to phyllotaxis. Nature 2014, 505, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Besnard, F.; Rozier, F.; Vernoux, T. The AHP6 cytokinin signaling inhibitor mediates an auxin-cytokinin crosstalk that regulates the timing of organ initiation at the shoot apical meristem. Plant Signal. Behav. 2014, 9, e28788. [Google Scholar] [CrossRef] [PubMed]
- Brandstatter, I.; Kieber, J.J. Two genes with similarity to bacterial response regulators are rapidly and specifically induced by cytokinin in Arabidopsis. Plant Cell 1998, 10, 1009–1020. [Google Scholar] [CrossRef]
- To, J.P.; Haberer, G.; Ferreira, F.J.; Deruere, J.; Mason, M.G.; Schaller, G.E.; Alonso, J.M.; Ecker, J.R.; Kieber, J.J. Type-A Arabidopsis response regulators are partially redundant negative regulators of cytokinin signaling. Plant Cell 2004, 16, 658–671. [Google Scholar] [CrossRef]
- Ma, Y.; Miotk, A.; Šutiković, Z.; Ermakova, O.; Wenzl, C.; Medzihradszky, A.; Gaillochet, C.; Forner, J.; Utan, G.; Brackmann, K.; et al. WUSCHEL acts as an auxin response rheostat to maintain apical stem cells in Arabidopsis. Nat. Commun. 2019, 10, 5093. [Google Scholar] [CrossRef]
- Gremski, K.; Ditta, G.; Yanofsky, M.F. The HECATE genes regulate female reproductive tract development in Arabidopsis thaliana. Development 2007, 134, 3593–3601. [Google Scholar] [CrossRef]
- Schuster, C.; Gaillochet, C.; Medzihradszky, A.; Busch, W.; Daum, G.; Krebs, M.; Kehle, A.; Lohmann, J.U. A regulatory framework for shoot stem cell control integrating metabolic, transcriptional, and phytohormone signals. Dev. Cell 2014, 28, 438–449. [Google Scholar] [CrossRef]
- MacArthur, B.D.; Ma’ayan, A.; Lemischka, I.R. Systems biology of stem cell fate and cellular reprogramming. Nat. Rev. Mol. Cell Biol. 2009, 10, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhao, X.; Hsieh, J.; Wichterle, H.; Impey, S.; Banerjee, S.; Neveu, P.; Kosik, K.S. MicroRNA regulation of neural stem cells and neurogenesis. J. Neurosci. 2010, 30, 14931–14936. [Google Scholar] [CrossRef] [PubMed]
- Heidstra, R.; Sabatini, S. Plant and animal stem cells: Similar yet different. Nat. Rev. Mol. Cell Biol. 2014, 15, 301–312. [Google Scholar] [CrossRef]
- Kinoshita, A.; Betsuyaku, S.; Osakabe, Y.; Mizuno, S.; Nagawa, S.; Stahl, Y.; Simon, R.; Yamaguchi-Shinozaki, K.; Fukuda, H.; Sawa, S. RPK2 is an essential receptor-like kinase that transmits the CLV3 signal in Arabidopsis. Development 2010, 137, 3911–3920. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Xu, Y.; Ng, K.H.; Ito, T. A timing mechanism for stem cell maintenance and differentiation in the Arabidopsis floral meristem. Genes Dev. 2009, 23, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Perales, M.; Gruel, J.; Girke, T.; Jönsson, H.; Reddy, G.V. WUSCHEL protein movement mediates stem cell homeostasis in the Arabidopsis shoot apex. Genes Dev. 2011, 25, 2025–2030. [Google Scholar] [CrossRef]
- Daum, G.; Medzihradszky, A.; Suzaki, T.; Lohmann, J.U. A mechanistic framework for non-cell autonomous stem cell induction in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 14619–14624. [Google Scholar] [CrossRef]
- Gruel, J.; Deichmann, J.; Landrein, B.; Hitchcock, T.; Jönsson, H. The interaction of transcription factors controls the spatial layout of plant aerial stem cell niches. Npj. Syst. Biol. Appl. 2018, 4, 36. [Google Scholar] [CrossRef]
- Lee, C.; Clark, S.E. A WUSCHEL-Independent Stem Cell Specification Pathway Is Repressed by PHB, PHV and CNA in Arabidopsis. PLoS ONE 2015, 10, e0126006. [Google Scholar] [CrossRef]
- Clark, S.E.; Running, M.P.; Meyerowitz, E.M. CLAVATA1, a regulator of meristem and flower development in Arabidopsis. Development 1993, 119, 397–418. [Google Scholar]
- Clark, S.E.; Running, M.P.; Meyerowitz, E.M. CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA1. Development 1995, 121, 2057–2067. [Google Scholar]
- Hu, C.; Zhu, Y.; Cui, Y.; Cheng, K.; Liang, W.; Wei, Z.; Zhu, M.; Yin, H.; Zeng, L.; Xiao, Y.; et al. A group of receptor kinases are essential for CLAVATA signalling to maintain stem cell homeostasis. Nat. Plants 2018, 4, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Perales, M.; Gruel, J.; Ohno, C.; Heisler, M.; Girke, T.; Reddy, G.V. Plant stem cell maintenance involves direct transcriptional repression of differentiation program. Mol. Syst. Biol. 2013, 9, 654. [Google Scholar] [CrossRef] [PubMed]
- Magnani, E.; Jiménez-Gómez, J.M.; Soubigou-Taconnat, L.; Lepiniec, L.; Fiume, E. Profiling the onset of somatic embryogenesis in Arabidopsis. BMC Genom. 2017, 18, 998. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Luo, J. The PIN-FORMED auxin efflux carriers in plants. Int. J. Mol. Sci. 2018, 19, 2759. [Google Scholar] [CrossRef] [PubMed]
- Swarup, R.; Bhosale, R. Developmental roles of AUX1/LAX auxin influx carriers in plants. Front. Plant Sci. 2019, 10, 1306. [Google Scholar] [CrossRef]
- Van Berkel, K.; de Boer, R.J.; Scheres, B.; ten Tusscher, K. Polar auxin transport: Models and mechanisms. Development 2013, 140, 2253–2268. [Google Scholar] [CrossRef]
- Bennett, T.; Hines, G.; van Rongen, M.; Waldie, T.; Sawchuk, M.G.; Scarpella, E.; Leyser, O. Connective auxin transport in the shoot facilitates communication between shoot apices. PLoS Biol. 2016, 14, e1002446. [Google Scholar] [CrossRef]
- Guenot, B.; Bayer, E.; Kierzkowski, D.; Smith, R.S.; Mandel, T.; Žádníková, P.; Kuhlemeier, C. Pin1-independent leaf initiation in Arabidopsis. Plant Physiol. 2012, 159, 1501–1510. [Google Scholar] [CrossRef]
- De Reuille, P.B.; Bohn-Courseau, I.; Ljung, K.; Morin, H.; Carraro, N.; Godin, C.; Traas, J. Computer simulations reveal properties of the cell-cell signaling network at the shoot apex in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 1627–1632. [Google Scholar] [CrossRef]
- Lincoln, C.; Long, J.; Yamaguchi, J.; Serikawa, K.; Hake, S. A Knotted1-like homeobox gene in Arabidopsis is expressed in the vegetative meristem and dramatically alters leaf morphology when overexpressed in transgenic plants. Plant Cell 1994, 6, 1859–1876. [Google Scholar] [PubMed]
- Kierzkowski, D.; Runions, A.; Vuolo, F.; Strauss, S.; Lymbouridou, R.; Routier-Kierzkowska, A.L.; Zhang, Z. A Growth-Based Framework for Leaf Shape Development and Diversity. Cell 2019, 177, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Gälweiler, L.; Guan, C.; Müller, A.; Wisman, E.; Mendgen, K.; Yephremov, A.; Palme, K. Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 1998, 282, 2226–2230. [Google Scholar] [CrossRef] [PubMed]
- Banasiak, A.; Biedron, M.; Dolzblasz, A.; Berezowsk, M.A. Ontogenetic changes in auxin biosynthesis and distribution determine the organogenic activity of the shoot apical meristem in pin1 mutants. Int. J. Mol. Sci. 2019, 20, 180. [Google Scholar] [CrossRef]
- Vidaurre, D.P.; Ploense, S.; Krogan, N.T.; Berleth, T. AMP1 and MP antagonistically regulate embryo and meristem development in Arabidopsis. Development 2007, 134, 2561–2567. [Google Scholar] [CrossRef]
- Zhao, Z.; Andersen, S.U.; Ljung, K.; Dolezal, K.; Miotk, A.; Schultheiss, S.J.; Lohmann, J.U. Hormonal control of the shoot stem-cell niche. Nature 2010, 465, 1089–1092. [Google Scholar] [CrossRef]
- Rademacher, E.H.; Möller, B.; Lokerse, A.S.; Llavata-Peris, C.I.; van den Berg, W.; Weijers, D. A cellular expression map of the Arabidopsis AUXIN RESPONSE FACTOR gene family. Plant J. 2011, 68, 597–606. [Google Scholar] [CrossRef]
- Denay, G.; Chahtane, H.; Tichtinsky, G.; Parcy, F. A flower is born: An update on Arabidopsis floral meristem formation. Curr. Opin. Plant Biol. 2017, 35, 15–22. [Google Scholar] [CrossRef]
- Krogan, N.T.; Marcos, D.; Weiner, A.I.; Berleth, T. The auxin response factor MONOPTEROS controls meristem function and organogenesis in both the shoot and root through the direct regulation of PIN genes. New Phytol. 2016, 212, 42–50. [Google Scholar] [CrossRef]
- Luo, L.; Zeng, J.; Wu, H.; Tian, Z.; Zhao, Z. A Molecular Framework for Auxin-Controlled Homeostasis of Shoot Stem Cells in Arabidopsis. Mol. Plant 2018, 11, 819–891. [Google Scholar] [CrossRef]
- Bayer, E.M.; Smith, R.S.; Mandel, T.; Nakayama, N.; Sauer, M.; Prusinkiewicz, P.; Kuhlemeier, C. Integration of transport-based models for phyllotaxis and midvein formation. Genes Dev. 2009, 23, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Capua, Y.; Eshed, Y. Coordination of auxin-triggered leaf initiation by tomato LEAFLESS. Proc. Natl. Acad. Sci. USA 2017, 114, 3246–3251. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Jiao, Y. The Diverse Roles of Auxin in Regulating Leaf Development. Plants 2019, 8, 243. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, D.; Mandel, T.; Kuhlemeier, C. Auxin regulates the initiation and radial position of plant lateral organs. Plant Cell 2000, 12, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, D.; Pesce, E.R.; Stieger, P.; Mandel, T.; Baltensperger, K.; Bennett, M.; Traas, J.; Friml, J.; Kuhlemeier, C. Regulation of phyllotaxis by polar auxin transport. Nature 2003, 426, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Ueda, J.; Komaki, M.K.; Bell, C.J.; Shimura, Y. Requirement of the auxin polar transport system in early stages of Arabidopsis floral bud formation. Plant Cell 1991, 3, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, D. Surface growth at the reproductive shoot apex of Arabidopsis thaliana pin-formed 1 and wild type. J. Exp. Bot. 2004, 55, 1021–1032. [Google Scholar] [CrossRef]
- Gendron, J.M.; Liu, J.S.; Fan, M.; Bai, M.Y.; Wenkel, S.; Springer, P.S.; Barton, M.K.; Wang, Z.Y. Brassinosteroids regulate organ boundary formation in the shoot apical meristem of Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 21152–21157. [Google Scholar] [CrossRef]
- Kawamura, E.; Horiguchi, G.; Tsukaya, H. Mechanisms of leaf tooth formation in Arabidopsis. Plant J. 2010, 62, 429–441. [Google Scholar] [CrossRef]
- Bilsborough, G.D.; Runions, A.; Barkoulas, M.; Jenkins, H.W.; Hasson, A.; Galinha, C.; Laufs, P.; Hay, A.; Prusinkiewicz, P.; Tsiantis, M. Model for the regulation of Arabidopsis thaliana leaf margin development. Proc. Natl. Acad. Sci. USA 2011, 108, 3424–3429. [Google Scholar] [CrossRef] [PubMed]
- Jasinski, S.; Piazza, P.; Craft, J.; Hay, A.; Woolley, L.; Rieu, I.; Phillips, A.; Hedden, P.; Tsiantis, M. KNOX action in Arabidopsis is mediated by coordinate regulation of cytokinin and gibberellin activities. Curr. Biol. 2005, 15, 1560–1565. [Google Scholar] [CrossRef] [PubMed]
- Machida, C.; Nakagawa, A.; Kojima, S.; Takahashi, H.; Machida, Y. The complex of ASYMMETRIC LEAVES (AS) proteins plays a central role in antagonistic interactions of genes for leaf polarity specification in Arabidopsis. Wiley interdisciplinary reviews. Dev. Bio. 2015, 4, 655–671. [Google Scholar] [CrossRef]
- Grigg, S.P.; Canales, C.; Hay, A.; Tsiantis, M. SERRATE coordinates shoot meristem function and leaf axial patterning in Arabidopsis. Nature 2005, 437, 1022–1026. [Google Scholar] [CrossRef]
- Li, Z.; Li, B.; Liu, J.; Guo, Z.; Liu, Y.; Li, Y.; Shen, W.-H.; Huang, Y.; Huang, H.; Zhang, Y.; et al. Transcription factors AS1 and AS2 interact with LHP1 to repress KNOX genes in Arabidopsis. J. Integr. Plant Biol. 2016, 58, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, H.; Ueno, Y.; Semiarti, E.; Onouchi, H.; Kojima, S.; Tsukaya, H.; Hasebe, M.; Soma, T.; Ikezaki, M.; Machida, C.; et al. The ASYMMETRIC LEAVES2 gene of Arabidopsis thaliana, required for formation of a symmetric flat leaf lamina, encodes a member of a novel family of proteins characterized by cysteine repeats and a leucine zipper. Plant Cell Physiol. 2002, 43, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Mei, G.; Hu, Y.; Deng, J.; Zhang, T. LMI1-like and KNOX1 genes coordinately regulate plant leaf development in dicotyledons. Plant Mol. Biol. 2019, 99, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Satterlee, J.W.; Scanlon, M.J. Coordination of Leaf Development across Developmental Axes. Plants 2019, 8, 433. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.M.; Jun, J.H.; Nam, H.G.; Fletcher, J.C. BLADE-ON-PETIOLE 1 and 2 control Arabidopsis lateral organ fate through regulation of LOB domain and adaxial-abaxial polarity genes. Plant Cell 2007, 19, 1809–1825. [Google Scholar] [CrossRef]
- Jun, J.H.; Ha, C.M.; Fletcher, J.C. BLADE-ON-PETIOLE1 coordinates organ determinacy and axial polarity in Arabidopsis by directly activating ASYMMETRIC LEAVES2. Plant Cell 2010, 22, 62–76. [Google Scholar] [CrossRef]
- Furumizu, C.; Alvarez, J.P.; Sakakibara, K.; Bowman, J.L. Antagonistic roles for KNOX1 and KNOX2 genes in patterning the land plant body plan following an ancient gene duplication. PLoS Genet. 2015, 11, e1004980. [Google Scholar] [CrossRef]
- Qi, J.; Wu, B.; Feng, S.; Lu, S.; Guan, C.; Zhang, X.; Qiu, D.; Hu, Y.; Zhou, Y.; Li, C.; et al. Mechanical regulation of organ asymmetry in leaves. Nat. Plants 2017, 3, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Kazama, T.; Ichihashi, Y.; Murata, S.; Tsukaya, H. The mechanism of cell cycle arrest front progression explained by a KLUH/CYP78A5-dependent mobile growth factor in developing leaves of Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Andriankaja, M.; Dhondt, S.; De Bodt, S.; Vanhaeren, H.; Coppens, F.; De Milde, L.; Muhlenbock, P.; Skirycz, A.; Gonzalez, N.; Beemster, G.T.; et al. Exit from proliferation during leaf development in Arabidopsis thaliana: A not-so-gradual process. Dev. Cell 2012, 22, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, M.; Cubas, P. TCP factors: New kids on the signaling block. Curr. Opin. Plant Biol. 2016, 33, 33–41. [Google Scholar] [CrossRef]
- Sablowski, R.; Carnier Dornelas, M. Interplay between cell growth and cell cycle in plants. J. Exp. Bot. 2014, 65, 2703–2714. [Google Scholar] [CrossRef]
- Czesnick, H.; Lenhard, M. Size control in plants–lessons from leaves and flowers. Cold Spring Harb. Perspect. Biol. 2015, 7, a019190. [Google Scholar] [CrossRef]
- Mizukami, Y.; Fischer, R.L. Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 942–947. [Google Scholar] [CrossRef]
- Anastasiou, E.; Kenz, S.; Gerstung, M.; MacLean, D.; Timmer, J.; Fleck, C.; Lenhard, M. Control of plant organ size by KLUH/CYP78A5-dependent intercellular signaling. Dev. Cell 2007, 13, 843–856. [Google Scholar] [CrossRef]
- Lee, B.H.; Ko, J.H.; Lee, S.; Lee, Y.; Pak, J.H.; Kim, J.H. The Arabidopsis GRF-INTERACTING FACTOR gene family performs an overlapping function in determining organ size as well as multiple developmental properties. Plant Physiol. 2009, 151, 655–668. [Google Scholar] [CrossRef]
- Rodriguez, R.E.; Mecchia, M.A.; Debernardi, J.M.; Schommer, C.; Weigel, D.; Palatnik, J.F. Control of cell proliferation in Arabidopsis thaliana by microRNA miR396. Development 2010, 137, 103–112. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, L.; Corke, F.; Smith, C.; Bevan, M.W. Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana. Genes Dev. 2008, 22, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Challa, K.R.; Rath, M.; Nath, U. The CIN-TCP transcription factors promote commitment to differentiation in Arabidopsis leaf pavement cells via both auxin-dependent and independent pathways. PLoS Genet. 2019, 15, e1007988. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, G.; Kim, G.T.; Tsukaya, H. The transcription factor AtGRF5 and the transcription coactivator AN3 regulate cell proliferation in leaf primordia of Arabidopsis thaliana. Plant J. 2005, 43, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kende, H. A transcriptional coactivator, AtGIF1, is involved in regulating leaf growth and morphology in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 13374–13379. [Google Scholar] [CrossRef]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of leaf morphogenesis by microRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef]
- Das Gupta, M.; Nath, U. Divergence in patterns of leaf growth polarity is associated with the expression divergence of miR396. Plant Cell 2015, 27, 2785–2799. [Google Scholar] [CrossRef]
- Vercruyssen, L.; Verkest, A.; Gonzalez, N.; Heyndrickx, K.S.; Eeckhout, D.; Han, S.K.; Jégu, T.; Archacki, R.; Van Leene, J.; Andriankaja, M.; et al. ANGUSTIFOLIA3 binds to SWI/SNF chromatin remodelling complexes to regulate transcription during Arabidopsis leaf development. Plant Cell 2014, 26, 210–229. [Google Scholar] [CrossRef]
- Efroni, I.; Blum, E.; Goldshmidt, A.; Eshed, Y. A protracted and dynamic maturation schedule underlies Arabidopsis leaf development. Plant Cell 2008, 20, 2293–2306. [Google Scholar] [CrossRef]
- Yin, Z.; Li, Y.; Zhu, W.; Fu, X.; Han, X.; Wang, J.; Lin, H.; Ye, W. Identification, Characterization, and Expression Patterns of TCP Genes and microRNA319 in Cotton. Int. J. Mol. Sci. 2018, 19, 3655. [Google Scholar] [CrossRef]
- Nath, U.; Crawford, B.C.; Carpenter, R.; Coen, E. Genetic control of surface curvature. Science 2003, 299, 1404–1407. [Google Scholar] [CrossRef]
- Ori, N.; Cohen, A.R.; Etzioni, A.; Brand, A.; Yanai, O.; Shleizer, S.; Menda, N.; Amsellem, Z.; Efroni, I.; Pekker, I.; et al. Regulation of LANCEOLATE by miR319 is required for compound leaf development in tomato. Nat. Genet. 2007, 39, 787–791. [Google Scholar] [CrossRef]
- Kim, J.H. Biological roles and an evolutionary sketch of the GRF-GIF transcriptional complex in plants. BMB Rep. 2019, 52, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.P.; Goldshmidt, A.; Efroni, I.; Bowman, J.L.; Eshed, Y. The NGATHA distal organ development genes are essential for style specification in Arabidopsis. Plant Cell 2009, 21, 1373–1393. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Poh, H.M.; Chua, N.-H. The Arabidopsis ARGOS-LIKE gene regulates cell expansion during organ growth. Plant J. 2006, 47, 1–9. [Google Scholar] [CrossRef]
- John, F.; Roffler, S.; Wicker, T.; Ringli, C. Plant TOR signaling components. Plant Signal Behav. 2011, 6, 1700–1705. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xie, Q.; Chua, N.H. The Arabidopsis auxin-inducible gene ARGOS controls lateral organ size. Plant Cell 2003, 15, 1951–1961. [Google Scholar] [CrossRef]
- Okushima, Y.; Mitina, I.; Quach, H.L.; Theologis, A. AUXIN RESPONSE FACTOR 2 (ARF2): A pleiotropic developmental regulator. Plant J. 2005, 43, 29–46. [Google Scholar] [CrossRef]
- Schruff, M.C.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R.J. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef]
- Trigueros, M.; Navarrete-Gomez, M.; Sato, S.; Christensen, S.K.; Pelaz, S.; Weigel, D.; Yanofsky, M.F.; Ferrandiz, C. The NGATHA genes direct style development in the Arabidopsis gynoecium. Plant Cell 2009, 21, 1394–1409. [Google Scholar] [CrossRef]
- Martínez-Fernández, I.; Sanchís, S.; Marini, N.; Balanzá, V.; Ballester, P.; Navarrete-Gómez, M.; Oliveira, A.C.; Colombo, L.; Ferrándiz, C. The effect of NGATHA altered activity on auxin signaling pathways within the Arabidopsis gynoecium. Front Plant Sci. 2014, 5, 21. [Google Scholar]
- Bresso, E.G.; Chorostecki, U.; Rodriguez, R.E.; Palatnik, J.F.; Schommer, C. Spatial control of gene expression by miR319-regulated TCP transcription factors in leaf development. Plant Physiol. 2018, 176, 1694–1708. [Google Scholar] [CrossRef] [PubMed]
- Nakata, M.; Matsumoto, N.; Tsugeki, R.; Rikirsch, E.; Laux, T.; Okada, K. Roles of the middle domain-specific WUSCHELRELATED HOMEOBOX genes in early development of leaves in Arabidopsis. Plant Cell 2012, 24, 519–535. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Raman, A.S.; Ream, J.E.; Fujiwara, H.; Cerny, R.E.; Brown, S.M. Overexpression of 20-oxidase confers a gibberellin-overproduction phenotype in Arabidopsis. Plant Physiol. 1998, 118, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Gusti, A.; Cheminant, S.; Alioua, M.; Dhondt, S.; Coppens, F.; Beemster, G.T.; Genschik, P. Gibberellin signalling controls cell proliferation rate in Arabidopsis. Curr. Biol. 2009, 19, 1188–1193. [Google Scholar] [CrossRef]
- Choe, S.; Fujioka, S.; Noguchi, T.; Takatsuto, S.; Yoshida, S.; Feldmann, K.A. Overexpression of DWARF4 in the brassinosteroid biosynthetic pathway results in increased vegetative growth and seed yield in Arabidopsis. Plant J. 2001, 26, 573–582. [Google Scholar] [CrossRef]
- Zhiponova, M.K.; Vanhoutte, I.; Boudolf, V.; Betti, C.; Dhondt, S.; Coppens, F.; Mylle, E.; Maes, S.; Gonzalez-Garcia, M.P.; Cano- Delgado, A.I.; et al. Brassinosteroid production and signalling differentially control cell division and expansion in the leaf. New Phytol. 2013, 197, 490–502. [Google Scholar] [CrossRef]
- Gonzalez, N.; Pauwels, L.; Baekelandt, A.; De Milde, L.; Van Leene, J.; Besbrugge, N.; Heyndrickx, K.S.; Cuellar Perez, A.; Durand, A.N.; De Clercq, R.; et al. A repressor protein complex regulates leaf growth in Arabidopsis. Plant Cell 2015, 27, 2273–2287. [Google Scholar] [CrossRef]
- Ikeda, M.; Fujiwara, S.; Mitsuda, N.; Ohme-Takagi, M. A triantagonistic basic helix–loop–helix system regulates cell elongation in Arabidopsis. Plant Cell 2012, 24, 4483–4497. [Google Scholar] [CrossRef]
- Bai, M.Y.; Fan, M.; Oh, E.; Wang, Z.Y. A triple helix-loop-helix/basic helix-loop-helix cascade controls cell elongation downstream of multiple hormonal and environmental signaling pathways in Arabidopsis. Plant Cell 2012, 24, 4917–4929. [Google Scholar] [CrossRef]
- Moon, J.; Hake, S. How a leaf gets its shape? Curr. Opin. Plant Biol. 2011, 14, 24–30. [Google Scholar] [CrossRef]
- Ochando, I.; González-Reig, S.; Ripoll, J.J.; Vera, A.; Martínez-Laborda, A. Alteration of the shoot radial pattern in Arabidopsis thaliana by a gain-of-function allele of the class III HD-Zip gene INCURVATA4. Int. J. Dev. Biol. 2008, 52, 953–961. [Google Scholar] [CrossRef]
- Waites, R.; Selvadurai, H.R.; Oliver, I.R.; Hudson, A. The PHANTASTICA gene encodes a MYB transcription factor involved in growth and dorsoventrality of lateral organs in Antirrhinum. Cell 1998, 93, 779–789. [Google Scholar] [CrossRef]
- Pekker, I.; John, P.A.; Yuval, E. Auxin response factors mediate Arabidopsis organ asymmetry via modulation of KANADI activity. Plant Cell 2005, 17, 2899–2910. [Google Scholar] [CrossRef] [PubMed]
- Sarojam, R.; Sappl, P.G.; Goldshmidt, A.; Efroni, I.; Floyd, S.K.; Eshed, Y.; Bowman, J.L. Differentiating Arabidopsis shoots from leaves by combined YABBY activities. Plant Cell 2010, 22, 2113–2130. [Google Scholar] [CrossRef]
- Kumaran, M.K.; Bowman, J.L.; Sundaresan, V. YABBY polarity genes mediate the repression of KNOX homeobox genes in Arabidopsis. Plant Cell 2002, 14, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Husbands, A.Y.; Benkovics, A.H.; Nogueira, F.T.; Lodha, M.; Timmermans, M.C. The ASYMMETRIC LEAVES complex employs multiple modes of regulation to affect adaxial-abaxial patterning and leaf complexity. Plant Cell 2015, 27, 3321–3335. [Google Scholar] [CrossRef]
- Williams, L.; Grigg, S.P.; Xie, M.; Christensen, S.; Fletcher, J.C. Regulation of Arabidopsis shoot apical meristem and lateral organ formation by microRNA miR166 g and its AtHD-ZIP target genes. Development 2005, 132, 3657–3668. [Google Scholar] [CrossRef]
- Singh, A.; Singh, S.; Panigrahi, K.C.; Reski, R.; Sarkar, A.K. Balanced activity of microRNA166/165 and its target transcripts from the class III homeodomain-leucine zipper family regulates root growth in Arabidopsis thaliana. Plant Cell Rep. 2014, 33, 945–953. [Google Scholar] [CrossRef]
- Yu, T.; Guan, C.; Wang, J.; Sajjad, M.; Ma, L.; Jiao, Y. Dynamic patterns of gene expression during leaf initiation. J. Genet. Genom. 2017, 44, 599–601. [Google Scholar] [CrossRef]
- Eshed, Y.; Baum, S.F.; Perea, J.V.; Bowman, J.L. Establishment of polarity in lateral organs of plants. Curr. Biol. 2001, 11, 1251–1260. [Google Scholar] [CrossRef]
- Kerstetter, R.A.; Bollman, K.; Taylor, R.A.; Bomblies, K.; Poethig, R.S. KANADI regulates organ polarity in Arabidopsis. Nature 2001, 411, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial patterning of Arabidopsis shoots by class III HD-ZIP and KANADI genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Roy, S.; Singh, S.; Das, S.S.; Gautam, V.; Yadav, S.; Sarkar, A.K. Phytohormonal crosstalk modulates the expression of miR166/165s, target Class III HD-ZIPs, and KANADI genes during root growth in Arabidopsis thaliana. Sci. Rep. 2017, 7, 3408. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, N.; Kim, H.B.; Park, N.I.; Kim, H.S.; Kim, Y.K.; Park, Y.I.; Choi, S.B. APUM23, a nucleolar Puf domain protein, is involved in pre-ribosomal RNA processing and normal growth patterning in Arabidopsis. Plant J. 2010, 64, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Ohbayashi, I.; Takahashi, H.; Kojima, S.; Ishibashi, N.; Keta, S.; Sugiyama, M. A genetic link between epigenetic repressor AS1-AS2 and a putative small subunit processome in leaf polarity establishment of Arabidopsis. Biol. Open 2016, 5, 942–954. [Google Scholar] [CrossRef] [PubMed]
- Luong, T.Q.; Keta, S.; Asai, T.; Kojima, S.; Nakagawa, A.; Micol, J.L.; Machida, C. A genetic link between epigenetic repressor AS1-AS2 and DNA replication factors in establishment of adaxial-abaxial leaf polarity of Arabidopsis. Plant Biotechnol. (Tokyo) 2018, 35, 39–49. [Google Scholar] [CrossRef]
- de Felippes, F.F.; Marchais, A.; Sarazin, A.; Oberlin, S.; Voinnet, O. A single miR390 targeting event is sufficient for triggering TAS3-tasiRNA biogenesis in Arabidopsis. Nucleic Acids Res. 2017, 45, 5539–5554. [Google Scholar] [CrossRef]
- Zhu, H.; Hu, F.; Wang, R.; Zhou, X.; Sze, S.H.; Liou, L.W.; Zhang, X. Arabidopsis Argonaute10 specifically sequesters miR166/165 to regulate shoot apical meristem development. Cell 2011, 145, 242–256. [Google Scholar] [CrossRef]
- Xia, R.; Xu, J.; Meyers, B.C. The emergence, evolution, and diversification of the miR390–TAS3–ARF pathway in land plants. Plant Cell 2017, 29, 1232–1247. [Google Scholar] [CrossRef]
- Sawa, S.S.; Watanabe, K.; Goto, K.; Kanaya, E.; Morita, E.H.; Okada, K. FILAMENTOUS FLOWER, a meristem and organ identity gene of Arabidopsis, encodes a protein with a zinc finger and HMG-related domain. Gene Dev. 1999, 13, 1079–1088. [Google Scholar] [CrossRef]
- Guan, C.; Wu, B.; Yu, T.; Wang, Q.; Krogan, N.T.; Liu, X.; Jiao, Y. Spatial Auxin Signaling Controls Leaf Flattening in Arabidopsis. Curr. Biol. CB 2017, 27, 2940–2950. [Google Scholar] [CrossRef]
- Siegfried, K.R.; Eshed, Y.; Baum, S.F.; Otsuga, D.; Drews, G.N.; Bowman, J.L. Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 1999, 126, 4117–4128. [Google Scholar] [PubMed]
- Li, H.; Xu, L.; Wang, H.; Yuan, Z.; Cao, X.; Yang, Z.; Zhang, D.; Xu, Y.; Huang, H. The putative RNA-dependent RNA polymerase RDR6 acts synergistically with ASYMMETRIC LEAVES1 and 2 to repress BREVIPEDICELLUS and microRNA165/166 in Arabidopsis leaf development. Plant Cell 2005, 17, 2157–2171. [Google Scholar] [CrossRef]
- Garcia, D.; Collier, S.A.; Byrne, M.E.; Martienssen, R.A. Specification of leaf polarity in Arabidopsis via the trans-acting siRNA pathway. Curr. Biol. 2006, 16, 933–938. [Google Scholar] [CrossRef]
- Eshed, Y.; Izhaki, A.; Baum, S.F.; Floyd, S.K.; Bowman, J.L. Asymmetric leaf development and blade expansion in Arabidopsis are mediated by KANADI and YABBY activities. Development 2004, 131, 2997–3006. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, A.; Yockteng, R.; Schnable, J.; Alvarez-Buylla, E.R.; Freeling, M.; Specht, C.D. Co-option of the polarity gene network shapes filament morphology in angiosperms. Sci. Rep. 2015, 4, 6194. [Google Scholar] [CrossRef] [PubMed]
- Takahiro, Y.; Akira, N.; Hirokazu, T. Leaf adaxial–abaxial polarity specification and lamina outgrowth: Evolution and development. Plant Cell Physiol. 2012, 53, 1180–1194. [Google Scholar] [CrossRef]
- Floyd, S.K.; Bowman, J.L. The ancestral developmental tool kit of land plants. Int. J. Plant Sci. 2007, 168, 1–35. [Google Scholar] [CrossRef]
- Floyd, S.K.; Bowman, J.L. Gene expression patterns in seed plant shoot meristems and leaves: Homoplasy or homology? J. Plant Res. 2010, 123, 43–55. [Google Scholar] [CrossRef]
- Nardmann, J.; Ji, J.; Werr, W.; Scanlon, M.J. The maize duplicate genes narrow sheath1 and narrow sheath2 encode a conserved homeobox gene function in a lateral domain of shoot apical meristems. Development 2004, 131, 2827–2839. [Google Scholar] [CrossRef]
- Aichinger, E.; Kornet, N.; Friedrich, T.; Laux, T. Plant stem cell niches. Annu. Rev. Plant Biol. 2012, 63, 615–636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tucker, E.; Hermann, M.; Laux, T. A Molecular Framework for the Embryonic Initiation of Shoot Meristem Stem Cells. Dev. Cell 2017, 40, 264–277. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Floyd, S.K. Patterning and polarity in seed plant shoots. Annu. Rev. Plant. Biol. 2008, 59, 67–88. [Google Scholar] [CrossRef] [PubMed]
- Fujino, K.; Matsuda, Y.; Ozawa, K.; Nishimura, T.; Koshiba, T.; Fraaije, M.W.; Sekiguchi, H. NARROW LEAF 7 controls leaf shape mediated by auxin in rice. Mol. Genet. Genom. 2008, 279, 499–507. [Google Scholar] [CrossRef]
- Kong, X.; Huang, G.; Xiong, Y.; Zhao, C.; Wang, J.; Song, X.; Zuo, K. IBR5 Regulates Leaf Serrations Development via Modulation of the Expression of PIN1. Int. J. Mol. Sci. 2019, 20, 4429. [Google Scholar] [CrossRef]
- Nikovics, K.; Blein, T.; Peaucelle, A.; Ishida, T.; Morin, H.; Aida, M.; Laufs, P. The balance between the MIR164A and CUC2 genes controls leaf margin serration in Arabidopsis. Plant Cell 2006, 18, 2929–2945. [Google Scholar] [CrossRef]
- Maugarny-Calès, A.; Cortizo, M.; Adroher, B.; Borrega, N.; Gonçalves, B.; Brunoud, G.; Laufs, P. Dissecting the pathways coordinating patterning and growth by plant boundary domains. PLoS Genet. 2019, 15, e1007913. [Google Scholar] [CrossRef]
- Hasson, A.; Plessis, A.; Blein, T.; Adroher, B.; Grigg, S.; Tsiantis, M.; Laufs, P. Evolution and diverse roles of the CUP-SHAPED COTYLEDON genes in Arabidopsis leaf development. Plant Cell 2011, 110, 54–68. [Google Scholar] [CrossRef]
- Sharma, A.; Zheng, B. Molecular Responses during Plant Grafting and Its Regulation by Auxins, Cytokinins, and Gibberellins. Biomolecules 2019, 9, 397. [Google Scholar] [CrossRef]
- Rast-Somssich, M.I.; Žádníková, P.; Schmid, S.; Kieffer, M.; Kepinski, S.; Simon, R. The Arabidopsis JAGGED LATERAL ORGANS (JLO) gene sensitizes plants to auxin. J. Exp. Bot. 2017, 68, 2741–2755. [Google Scholar] [CrossRef]
- Rast, M.I.; Simon, R. Arabidopsis JAGGED LATERAL ORGANS acts with ASYMMETRIC LEAVES2 to coordinate KNOX and PIN expression in shoot and root meristems. Plant Cell 2012, 24, 2917–2933. [Google Scholar] [CrossRef] [PubMed]
- Žádníková, P.; Simon, R. How boundaries control plant development. Curr. Opin. Plant Biol. 2014, 17, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Hasson, A.; Belcram, K.; Cortizo, M.; Morin, H.; Nikovics, K.; Vialette-Guiraud, A.; Takeda, S.; Aida, M.; Laufs, P.; et al. A conserved role for CUP-SHAPED COTYLEDON genes during ovule development. Plant J. 2015, 83, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Engelhorn, J.; Reimer, J.J.; Leuz, I.; Göbel, U.; Huettel, B.; Farrona, S.; Turck, F. DEVELOPMENT-RELATED PcG TARGET IN THE APEX 4 controls leaf margin architecture in Arabidopsis thaliana. Development 2012, 139, 2566–2575. [Google Scholar] [CrossRef]
- Yang, Y.; Nicolas, M.; Zhang, J.; Yu, H.; Guo, D.; Yuan, R.; Qin, G. Correction: The TIE1 transcriptional repressor controls shoot branching by directly repressing BRANCHED1 in Arabidopsis. PLoS Genet. 2018, 14, e1007565. [Google Scholar] [CrossRef]
- Tao, Q.; Guo, D.; Wei, B.; Zhang, F.; Pang, C.; Jiang, H.; Zhang, J.; Wei, T.; Gu, H.; Qu, L.J.; et al. The TIE1 transcriptional repressor links TCP transcription factors with TOPLESS/ TOPLESS-RELATED corepressors and modulates leaf development in Arabidopsis. Plant Cell 2013, 25, 421–437. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, B.; Yuan, R.; Wang, J.; Ding, M.; Chen, Z.; Yu, H.; Qin, G. The Arabidopsis RING-type E3 ligase TEAR1 controls leaf development by targeting the TIE1 transcrip-tional repressor for degradation. Plant Cell 2017, 29, 243–259. [Google Scholar] [CrossRef]
- Tameshige, T.; Okamoto, S.; Lee, J.S.; Aida, M.; Tasaka, M.; Torii, K.U.; Uchida, N. A secreted peptide and its receptors shape the auxin response pattern and leaf margin morphogenesis. Curr. Biol. 2016, 26, 2478–2485. [Google Scholar] [CrossRef]
- Toriba, T.; Tokunaga, H.; Shiga, T.; Nie, F.; Naramoto, S.; Honda, E.; Kyozuka, J. BLADE-ON-PETIOLE genes temporally and developmentally regulate the sheath to blade ratio of rice leaves. Nat. Commun. 2019, 10, 619. [Google Scholar] [CrossRef]
- Norberg, M.; Holmlund, M.; Nilsson, O. The BLADE ON PETIOLE genes act redundantly to control the growth and development of lateral organs. Development 2005, 132, 2203–2213. [Google Scholar] [CrossRef]
- Sakamoto, T.; Sotta, N.; Suzuki, T.; Fujiwara, T.; Matsunaga, S. The 26S proteasome is required for the maintenance of root apical meristem by modulating auxin and cytokinin responses under high-boron stress. Front. Plant Sci. 2019, 10, 590. [Google Scholar] [CrossRef] [PubMed]
- Wójcik, A.M.; Nodine, M.D.; Gaj, M.D. miR160 and miR166/165 Contribute to the LEC2-Mediated Auxin Response Involved in the Somatic Embryogenesis Induction in Arabidopsis. Front. Plant Sci. 2017, 8, 2024. [Google Scholar] [CrossRef] [PubMed]
- Saini, K.; Markakis, M.N.; Zdanio, M.; Balcerowicz, D.M.; Beeckman, T.; De Veylder, L.; Vissenberg, K. Alteration in auxin homeostasis and signaling by overexpression of PINOID kinase causes leaf growth defects in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1009. [Google Scholar] [CrossRef] [PubMed]
- Kasprzewska, A.; Carter, R.; Swarup, R.; Bennett, M.; Monk, N.; Hobbs, J.K.; Fleming, A. Auxin influx importers modulate serration along the leaf margin. Plant J. 2015, 83, 705–718. [Google Scholar] [CrossRef]
- Stefano, P.; Ivana, M.; Arianna, G.; Letizia, Z.; Angelo, G.; Rosella, C.; Andrea, G. Identification of microRNAs and relative target genes in Moringa oleifera leaf and callus. Sci. Rep. 2019, 9, 1–14. [Google Scholar]
- Wang, J.; Mei, J.; Ren, G. Plant microRNAs: Biogenesis, homeostasis and degradation. Front. Plant Sci. 2019, 10, 360. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Du, T.; Zamore, P.D. microPrimer: The biogenesis and function of microRNA. Development 2005, 132, 4645–4652. [Google Scholar] [CrossRef]
- Rogers, K.; Chen, X. Biogenesis, turnover, and mode of action of plant MicroRNAs. Plant Cell 2013, 25, 2383–2399. [Google Scholar] [CrossRef]
- Iwakawa, H.O.; Tomari, Y. The functions of MicroRNAs: mRNA decay and translational repression. Trends Cell Biol. 2015, 25, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Bologna, N.G.; Mateos, J.L.; Bresso, E.G.; Palatnik, J.F. A loop-to-base processing mechanism underlies the biogenesis of plant microRNAs miR319 and miR159. EMBO J. 2009, 28, 3646–3656. [Google Scholar] [CrossRef]
- Michlewski, G.; Cáceres, J.F. Post-transcriptional control of miRNA biogenesis. RNA 2019, 25, 1–16. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Shao, C.; Chen, M. Toward microRNA-mediated gene regulatory networks in plants. Brief Bioinform. 2011, 12, 645–659. [Google Scholar] [CrossRef]
- Yu, Y.; Ji, L.; Le, B.H.; Zhai, J.; Chen, J.; Luscher, E.; Gao, L.; Liu, C.; Cao, X.; Mo, B.; et al. ARGONAUTE10 promotes the degradation of miR165/6 through the SDN1 and SDN2 exonucleases in Arabidopsis. PLoS Biol. 2017, 15, e2001272. [Google Scholar] [CrossRef]
- Armenta-Medina, A.; Lepe-Soltero, D.; Xiang, D.Q.; Datla, R.; Abreu-Goodger, C.; Gillmor, C.S. Arabidopsis thaliana miRNAs promote embryo pattern formation beginning in the zygote. Dev. Biol. 2017, 431, 145–151. [Google Scholar] [CrossRef]
- Knauer, S.; Holt, A.L.; Rubio-Somoza, I.; Tucker, E.J.; Hinze, A.; Pisch, M.; Javelle, M.; Timmermans, M.C.; Tucker, M.R.; Laux, T. A protodermal miR394 signal defines a region of stem cell competence in the Arabidopsis shoot meristem. Dev. Cell 2013, 24, 125–132. [Google Scholar] [CrossRef]
- Pulido, A.; Laufs, P. Co-ordination of developmental processes by small RNAs during leaf development. J. Exp. Bot. 2010, 61, 1277–1291. [Google Scholar] [CrossRef]
- Yang, T.; Wang, Y.; Teotia, S.; Wang, Z.; Shi, C.; Sun, H.; Tang, G. The interaction between miR160 and miR165/166 in the control of leaf development and drought tolerance in Arabidopsis. Sci. Rep. 2019, 9, 2832. [Google Scholar] [CrossRef]
- Mallory, A.C.; Bartel, D.P.; Bartel, B. MicroRNA-directed regulation of Arabidopsis AUXIN RESPONSE FACTOR17 is essential for proper development and modulates expression of early auxin response genes. Plant Cell 2005, 17, 1360–1375. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.P.; Montgomery, T.A.; Fahlgren, N.; Kasschau, K.D.; Nonogaki, H.; Carrington, J.C. Repression of AUXIN RESPONSE FACTOR10 by microRNA160 is critical for seed germination and post-germination stages. Plant J. 2007, 52, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Chen, J.; Zhou, J.; Yu, H.; Ge, C.; Zhang, M.; Zhao, Y. An essential role for miRNA167 in maternal control of embryonic and seed development. Plant Physiol. 2019, 180, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Ding, N.; Fan, W.; Yan, J.; Gu, Y.; Tang, X.; Li, R.; Tang, G. Functional plasticity of miR165/166 in plant development revealed by small tandem target mimic. Plant Sci. 2015, 233, 11–21. [Google Scholar] [CrossRef]
- Fahlgren, N.; Montgomery, T.A.; Howell, M.D.; Allen, E.; Dvorak, S.K.; Alexander, A.L.; Carrington, J.C. Regulation of AUXIN RESPONSE FACTOR3 by TAS3 ta-siRNA affects developmental timing and patterning in Arabidopsis. Curr. Biol. 2006, 16, 939–944. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Y.; Liu, Y.; Li, D.; Zhao, Y.; Li, Z.; Liu, Z. Overexpression of OsAGO1b Induces Adaxially Rolled Leaves by Affecting Leaf Abaxial Sclerenchymatous Cell Development in Rice. Rice 2019, 12, 60. [Google Scholar] [CrossRef]
- Kidner, C.A.; Martienssen, R.A. Spatially restricted microRNA directs leaf polarity through ARGONAUTE1. Nature 2004, 428, 81–84. [Google Scholar] [CrossRef]
- Nagasaki, H.; Itoh, J.I.; Hayashi, K.; Hibara, K.I.; Satoh-Nagasawa, N.; Nosaka, M.; Nagato, Y. The small interfering RNA production pathway is required for shoot meristem initiation in rice. Proc. Natl. Acad. Sci. USA 2007, 104, 14867–14871. [Google Scholar] [CrossRef]
- Xu, M.; Hu, T.; Zhao, J.; Park, M.Y.; Earley, K.W.; Wu, G.; Yang, L.; Poethig, R.S. Developmental functions of miR156-regulated SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) genes in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006263. [Google Scholar] [CrossRef]
- Scanlon, M.J.; Schneeberger, R.G.; Freeling, M. The maize mutant narrow sheath fails to establish leaf margin identity in a meristematic domain. Development 1996, 122, 1683–1691. [Google Scholar]
- Merelo, P.; Ram, H.; Caggiano, M.P.; Ohno, C.; Ott, F.; Straub, D.; Heisler, M.G. Regulation of MIR165/166 by class II and class III homeodomain leucine zipper proteins establishes leaf polarity. Proc. Natl. Acad. Sci. USA 2016, 113, 11973–11978. [Google Scholar] [CrossRef] [PubMed]
- Waites, R.; Hudson, A. phantastica: A gene required for dorsoventrality of leaves in Antirrhinum majus. Development 1995, 121, 2143–2154. [Google Scholar]
- Chuck, G.; Whipple, C.; Jackson, D.; Hake, S. The maize SBP-box transcription factor encoded by tasselsheath4 regulates bract development and the establishment of meristem boundaries. Development 2010, 137, 1243–1250. [Google Scholar] [CrossRef]
- Achard, P.; Herr, A.; Baulcombe, D.C.; Harberd, N.P. Modulation of floral development by a gibberellin-regulated microRNA. Development 2004, 131, 3357–3365. [Google Scholar] [CrossRef]
- Damodharan, S.; Zhao, D.; Arazi, T. A common miRNA160-based mechanism regulates ovary patterning, floral organ abscission and lamina outgrowth in tomato. Plant J. 2016, 86, 458–471. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Kasschau, K.D.; Carrington, J.C. Negative feedback regulation of Dicer-Like1 in Arabidopsis by microRNA-guided mRNA degradation. Curr. Biol. 2003, 13, 784–789. [Google Scholar] [CrossRef]
- Raman, S.; Greb, T.; Peaucelle, A.; Blein, T.; Laufs, P.; Theres, K. Interplay of miR164, CUP-SHAPED COTYLEDON genes and LATERAL SUPPRESSOR controls axillary meristem formation in Arabidopsis thaliana. Plant J. 2008, 55, 65–76. [Google Scholar] [CrossRef]
- Busch, B.L.; Schmitz, G.; Rossmann, S.; Piron, F.; Ding, J.; Bendahmane, A.; Theres, K. Shoot branching and leaf dissection in tomato are regulated by homologous gene modules. Plant Cell 2011, 23, 3595–3609. [Google Scholar] [CrossRef]
- Li, X.; Qian, Q.; Fu, Z.; Wang, Y.; Xiong, G.; Zeng, D.; Wang, X.; Liu, X.; Teng, S.; Hiroshi, F.; et al. Control of tillering in rice. Nature 2003, 422, 618–621. [Google Scholar] [CrossRef]
- Huijser, P.; Schmid, M. The control of developmental phase transitions in plants. Development 2011, 138, 4117–4129. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yao, X.; Pi, L.; Wang, H.; Cui, X.; Huang, H. The ARGONAUTE10 gene modulates shoot apical meristem maintenance and establishment of leaf polarity by repressing miR165/166 in Arabidopsis. Plant J. 2009, 58, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Aukerman, M.J.; Sakai, H. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2- like target genes. Plant Cell 2003, 15, 2730–2741. [Google Scholar] [CrossRef] [PubMed]
- Cartolano, M.; Castillo, R.; Efremova, N.; Kuckenberg, M.; Zethof, J.; Gerats, T.; Vandenbussche, M. A conserved microRNA module exerts homeotic control over Petunia hybrida and Antirrhinum majus floral organ identity. Nat. Genet. 2007, 39, 901. [Google Scholar] [CrossRef] [PubMed]
- Marin, E.; Jouannet, V.; Herz, A.; Lokerse, A.S.; Weijers, D.; Vaucheret, H.; Nussaume, L.; Crespi, M.D.; Maizel, A. miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. Plant Cell 2010, 22, 1104–1117. [Google Scholar] [CrossRef]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. Plant Cell 2005, 17, 61–76. [Google Scholar] [CrossRef]
- Vaucheret, H. AGO1 Homeostasis Involves Differential Production of 21-nt and 22-nt miR168 Species by MIR168a and MIR168b. PLoS ONE 2009, 4, e6442. [Google Scholar] [CrossRef]
- Ji, L.; Liu, X.; Yan, J.; Wang, W.; Yumul, R.E.; Kim, Y.J.; Dinh, T.T.; Liu, J.; Cui, X.; Zheng, B.; et al. ARGONAUTE10 and ARGONAUTE1 regulate the termination of floral stem cells through two microRNAs in Arabidopsis. PLoS Genet. 2011, 7, e1001358. [Google Scholar] [CrossRef]
- Lim, P.O.; Kim, H.J.; Nam, H.G. Leaf senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef]
- Jung, J.H.; Lee, S.; Yun, J.; Lee, M.; Park, C.M. The miR172 target TOE3 represses AGAMOUS expression during Arabidopsis floral patterning. Plant Sci. 2014, 215, 29–38. [Google Scholar] [CrossRef]
- Palatnik, J.F.; Wollmann, H.; Schommer, C.; Schwab, R.; Boisbouvier, J.; Rodriguez, R.; Warthmann, N.; Allen, E.; Dezulian, T.; Huson, D.; et al. Sequence and expression differences underlie functional specialization of Arabidopsis microRNAs miR159 and miR319. Dev. Cell 2007, 13, 115–125. [Google Scholar] [CrossRef]
- Montgomery, T.A.; Howell, M.D.; Cuperus, J.T.; Li, D.; Hansen, J.E.; Alexander, A.L.; Chapman, E.J.; Fahlgren, N.; Allen, E.; Carrington, J.C. Specificity of ARGONAUTE7–miR390 interaction and dual functionality in TAS3 trans-acting siRNA formation. Cell 2008, 133, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Laufs, P.; Peaucelle, A.; Morin, H.; Traas, J. MicroRNA regulation of the CUC genes is required for boundary size control in Arabidopsis meristems. Development 2004, 131, 4311–4322. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.S.; Li, J.; Stahle, M.I.; Dubroué, A.; Gubler, F.; Millar, A.A. Genetic analysis reveals functional redundancy and the major target genes of the Arabidopsis miR159 family. Proc. Natl. Acad. Sci. USA 2007, 104, 16371–16376. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Schwab, R.; Czech, B.; Mica, E.; Weigel, D. Dual effects of miR156-targeted SPL genes and CYP78A5/KLUH on plastochron length and organ size in Arabidopsis thaliana. Plant Cell 2008, 20, 1231–1243. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.K.; Xu, Y.M.; Shi, M.; Lai, Y.M.; Wu, X.; Wang, H.S.; Zhu, Z.; Poethig, R.S.; Wu, G. Repression of miR156 by miR159 regulates the timing of the juvenile-to-adult transition in Arabidopsis. Plant Cell 2017, 29, 1293–1304. [Google Scholar] [CrossRef]
- Chen, X. Small RNAs and their roles in plant development. Annu. Rev. Cell Dev. Biol. 2009, 25, 21–44. [Google Scholar] [CrossRef]
- Chen, X. A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development. Science 2004, 303, 2022–2025. [Google Scholar] [CrossRef]
- Wollmann, H.; Mica, E.; Todesco, M.; Long, J.A.; Weigel, D. On reconciling the interactions between APETALA2, miR172 and AGAMOUS with the ABC model of flower development. Development 2010, 137, 3633–3642. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ping, X.; Cao, Y.; Jian, H.; Gao, Y.; Wang, J.; Liu, L. Genome-wide exploration and characterization of miR172/euAP2 genes in Brassica napus L. for likely role in flower organ development. BMC Plant Biol. 2019, 19, 336. [Google Scholar] [CrossRef]
- Koyama, T.; Furutani, M.; Tasaka, M.; Ohme-Takagi, M. TCP transcription factors control the morphology of shoot lateral organs via negative regulation of the expression of boundary-specific genes in Arabidopsis. Plant Cell 2007, 19, 473–484. [Google Scholar] [CrossRef]
- Li, Z.; Peng, J.; Wen, X.; Guo, H. Ethylene-insensitive3 is a senescence associated gene that accelerates age-dependent leaf senescence by directly repressing miR164 transcription in Arabidopsis. Plant Cell 2013, 25, 3311–3328. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.M.; Nagpal, P.; Young, J.C.; Hagen, G.; Guilfoyle, T.J.; Reed, J.W. AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 2005, 132, 4563–4574. [Google Scholar] [CrossRef] [PubMed]
- Parry, G.; Calderon-Villalobos, L.I.; Prigge, M.; Peret, B.; Dharmasiri, S.; Itoh, H.; Lechner, E.; Gray, W.M.; Bennett, M.; Estelle, M. Complex regulation of the TIR1/AFB family of auxin receptors. Proc. Natl. Acad. Sci. USA 2009, 106, 22540–22545. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Chen, R. PHANTASTICA regulates leaf polarity and petiole identity in Medicago truncatula. Plant Signal. Behav. 2014, 9, e28121. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).