The T2 Toxin Produced by Fusarium spp. Impacts Porcine Duodenal Nitric Oxide Synthase (nNOS)-Positive Nervous Structures—The Preliminary Study

 , ,
, , 
Abstract
1. Introduction
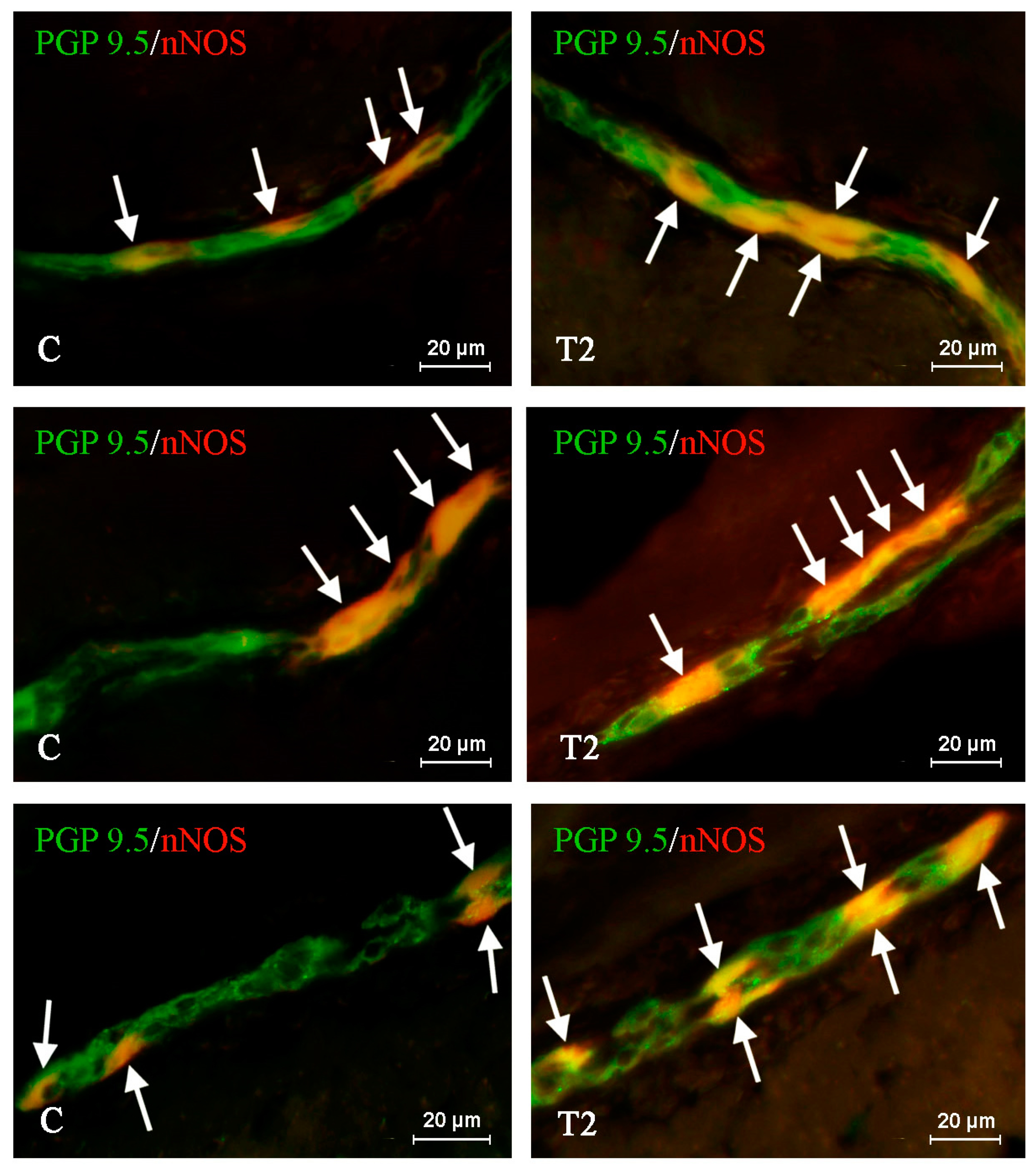
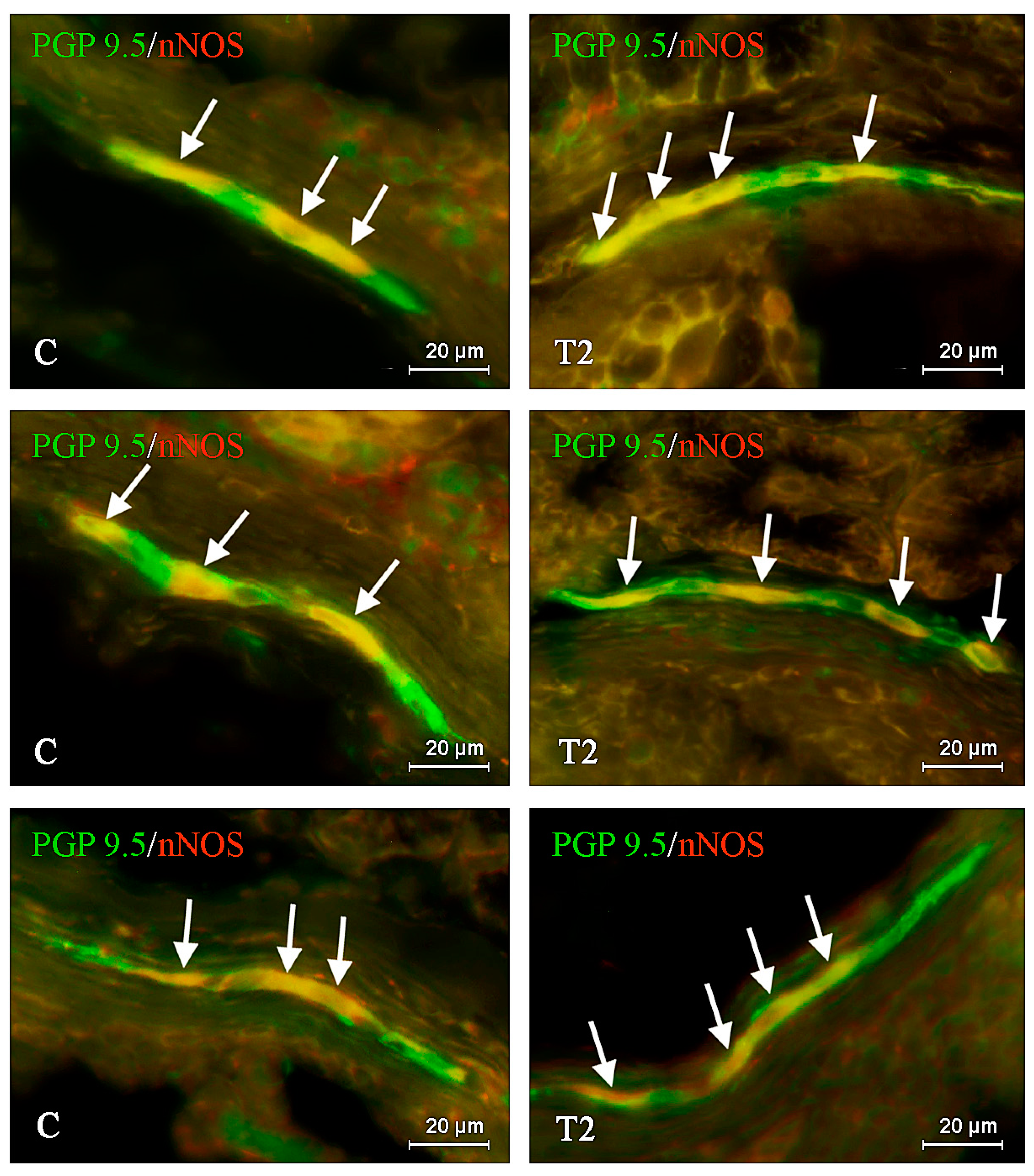
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kuchenbuch, H.S.; Becker, S.; Schulz, M.; Cramer, B.; Humpf, H.U. Thermal stability of T-2 and HT-2 toxins during biscuit- and crunchy muesli-making and roasting. Food Addit. Contam. Part A 2018, 35, 2158–2167. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Liu, S.; Wu, W.; Zhang, H. Comparison of anorectic potencies of type a trichothecenes T-2 toxin, HT-2 toxin, diacetoxyscirpenol, and neosolaniol. Toxins 2018, 10, 179. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; De Smet, D.; De Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef]
- Zhang, J.; Sheng, K.; Wu, W.; Zhang, H. Anorectic responses to T-2 toxin, HT-2 toxin, diacetoxyscirpenol and neosolaniol correspond to plasma elevations of neurotransmitters 5-hydroxytryptamine and substance P. Ecotoxicol. Environ. Saf. 2018, 161, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.; Zhang, H.; Yue, J.; Gu, W.; Gu, C.; Zhang, H.; Wu, W. Anorectic response to the trichothecene T-2 toxin correspond to plasma elevations of the satiety hormone glucose-dependent insulinotropic polypeptide and peptide YY 3-36. Toxicology 2018, 403, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Makowska, K.; Obremski, K.; Gonkowski, S. The impact of T-2 toxin on vasoactive intestinal polypeptide-like immunoreactive (VIP-LI) nerve structures in the wall of the porcine stomach and duodenum. Toxins 2018, 10, 138. [Google Scholar] [CrossRef]
- Makowska, K.; Obremski, K.; Zielonka, Ł.; Gonkowski, S. The influence of low doses of zearalenone and T-2 toxin on calcitonin gene related peptide-like immunoreactive (CGRP-LI) neurons in the ENS of the porcine descending colon. Toxins 2017, 9, 98. [Google Scholar] [CrossRef]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. Adv. Exp. Med. Biol. 2014, 817, 39–71. [Google Scholar] [CrossRef] [PubMed]
- Szymanska, K.; Makowska, K.; Gonkowski, S. The influence of high and low doses of Bisphenol A (BPA) on the enteric nervous system of the porcine ileum. Int. J. Mol. Sci. 2018, 19, 917, . [Google Scholar] [CrossRef]
- Makowska, K.; Gonkowski, S.; Zielonka, Ł.; Dąbrowski, M.; Całka, J. T2 toxin-induced changes in cocaine- and amphetamine-regulated transcript (CART)-like immunoreactivity in the enteric nervous system within selected fragments of the porcine digestive tract. Neurotox. Res. 2016, 31, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Brehmer, A. Structure of enteric neurons. Adv. Anat. Embryol. Cell Biol. 2006, 186, 1–91. [Google Scholar] [PubMed]
- Timmermans, J.; Barbiers, M.; Scheuermann, D.W.; Stach, W.; Adriaensen, D.; Mayer, B.; De Groodt-Lasseel, M.H. Distribution pattern, neurochemical features and projections of nitrergic neurons in the pig small intestine. Anat. Anz. 1994, 176, 515–525. [Google Scholar] [CrossRef]
- Verma, N.; Schmitz-Spanke, S.; Rettenmeier, A.W. Recent advances in the use of Sus scrofa (pig) as a model system for proteomic studies. Proteomics 2011, 11, 776–793. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, A.; Gonkowski, S.; Nowicki, M.; Calka, J. Inflammatory bowel disease affects density of nitrergic nerve fibers in the mucosal layer of the canine gastrointestinal tract. Can. J. Vet. Res. 2017, 81, 129–136. [Google Scholar] [PubMed]
- Vasina, V.; Barbara, G.; Talamonti, L.; Stanghellini, V.; Corinaldesi, R.; Tonini, M.; De Ponti, F.; De Giorgio, R. Enteric neuroplasticity evoked by inflammation. Auton. Neurosci. 2006, 126, 264–272. [Google Scholar] [CrossRef]
- Makowska, K. Chemically induced inflammation and nerve damage affect the distribution of vasoactive intestinal polypeptide-like immunoreactive (VIP-LI) nervous structures in the descending colon of the domestic pig. Neurogastroenterol. Motil. 2018, 30, 13439. [Google Scholar] [CrossRef]
- Gonkowski, S.; Całka, J. Changes in the somatostatin (SOM)-like immunoreactivity within nervous structures of the porcine descending colon under various pathological factors. Exp. Mol. Pathol. 2010, 88, 416–423. [Google Scholar] [CrossRef]
- Esplugues, J.V. NO as a signalling molecule in the nervous system. Br. J. Pharmacol. 2002, 135, 1079–1095. [Google Scholar] [CrossRef] [PubMed]
- Pidsudko, Z.; WasowiczK; Kaleczyc, J.; Majewski, M.; Lakomy, M. Proliferative enteropathy (PPE)-induced changes in the expression of DBH, VAChT and NOS in the neurons of intramural ganglia of the porcine ileum. Vet. Med. 2008, 53, 533–542. [Google Scholar] [CrossRef]
- Shah, V.H.; Lyford, G.; Gores, G.; Farrugia, G. Nitric oxide in gastrointestinal health and disease. Gastroenterology 2004, 126, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Schleiffer, R.; Raul, F. Nitric oxide and the digestive system in mammals and non-mammalian vertebrates. Comp. Biochem. Physiol. Part A Physiol. 1997, 118, 965–974. [Google Scholar] [CrossRef]
- Kuwahara, A.; Kuramoto, H.; Kadowaki, M. 5-HT activates nitric oxide-generating neurons to stimulate chloride secretion in guinea pig distal colon. Am. J. Physiol. 1998, 275, 829–834. [Google Scholar] [CrossRef]
- Sang, Q.; Young, H. Chemical coding of neurons in the myenteric plexus and external muscle of the small and large intestine of the mouse. Cell Tissue Res. 1996, 284, 39–53. [Google Scholar] [CrossRef]
- Kochar, N.I.; Chandewal, A.V.; Bakal, R.L.; Kochar, P.N. Nitric oxide and the gastrointestinal tract. Int. J. Pharmacol. 2011, 7, 31–39. [Google Scholar] [CrossRef]
- Lin, Z.; Sandgren, K.; Ekblad, E. Increased expression of nitric oxide synthase in cultured neurons from adult rat colonic submucous ganglia. Auton. Neurosci. 2004, 114, 29–38. [Google Scholar] [CrossRef] [PubMed]
- De Fontgalland, D.A.; Wattchow, D.; Costa, M.; Brookes, S.J.H. Immunohistochemical characterization of the innervation of human colonic mesenteric and submucosal blood vessels. Neurogastroenterol. Motil. 2008, 20, 1212–1226. [Google Scholar] [CrossRef]
- Gonkowski, S.; Kamińska, B.; Landowski, P.; Całka, J. Immunohistochemical distribution of cocaine- and amphetamine-regulated transcript peptide-like immunoreactive (CART-LI) nerve fibers and various degree of co-localization with other neuronal factors in the circular muscle layer of human descending colon. Histol. Histopathol. 2012, 28, 851–858. [Google Scholar] [PubMed]
- Takeda, M.; Miyahara, K.; Sueyoshi, R.; Arakawa, A.; Lane, G.J.; Yamataka, A. Innervation of the entire internal anal sphincter in a mouse model of Hirschsprung’s disease: A first report. Pediatr. Surg. Int. 2018, 35, 209–214. [Google Scholar] [CrossRef] [PubMed]
- King, B.F. Purinergic signalling in the enteric nervous system (An overview of current perspectives). Auton. Neurosci. 2015, 191, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.; Du, C.; Ledlow, A.; Bates, J.N.; Conkin, J.L. Nitric oxide: Mediator of nonadrenergic noncholinergicresponses of opossum esophagal muscle. Am. J. Physiol. 1991, 261, 401–406. [Google Scholar] [CrossRef]
- Sarna, S.K.; Otterson, M.F.; Ryan, R.P.; Cowles, V.E. Nitric oxide regulates migrating motor complex cycling and its postprandial disruption. Am. J. Physiol. 1993, 265, 759–766. [Google Scholar] [CrossRef]
- Watkins, D.J.; Besner, G.E. The role of the intestinal microcirculation in necrotizing enterocolitis. Semin. Pediatr. Surg. 2013, 22, 83–87. [Google Scholar] [CrossRef]
- Lanas, A. Role of nitric oxide in the gastrointestinal tract. Arthritis Res. Ther. 2008, 10, S4. [Google Scholar] [CrossRef]
- Beck, M.; Schlabrakowski, A.; Schrödl, F.; Neuhuber, W.; Brehmer, A. ChAT and NOS in human myenteric neurons: Co-existence and co-absence. Cell Tissue Res. 2009, 338, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Stanek, A.; Gadowska-Cicha, A.; Gawron, K.; Wielkoszyński, T.; Adamek, B.; Cieślar, G.; Wiczkowski, A.; Sieron, A. Role of nitric oxide in physiology and pathology of the gastrointestinal tract. Mini Rev. Med. Chem. 2008, 8, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.; Suzuki, T.; Murota, M.; Takahashi, Y.; Takeguchi, N. Nitric oxide-induced Cl− secretion in isolated rat colon is mediated by the release of thromboxane A2. J. Physiol. 2002, 543, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Baraona, E.; Shoichet, L.; Navder, K.; Lieber, C.S. Mediation by nitric oxide of the stimulatory effects of ethanol on blood flow. Life Sci. 2002, 70, 2987–2995. [Google Scholar] [CrossRef]
- Szymanska, K.; Gonkowski, S. Neurochemical characterization of the enteric neurons within the porcine jejunum in physiological conditions and under the influence of bisphenol A (BPA). Neurogastroenterol. Motil. 2019, 31, e13580. [Google Scholar] [CrossRef]
- Sugata, H.; Ueno, T.; Shimosegawa, T.; Yoshimura, T. Direct detection of nitric oxide and its roles in maintaining gastric mucosal integrity following ethanol-induced injury in rats. Free. Radic. Res. 2003, 37, 159–169. [Google Scholar] [CrossRef]
- Ancha, H.; Ojeas, H.; Tedesco, D.; Ward, A.; Harty, R.F. Somatostatin-induced gastric protection against ethanol: Involvement of nitric oxide and effects on gastric mucosal blood flow. Regul. Pept. 2003, 110, 107–113. [Google Scholar] [CrossRef]
- Le, Q.L.; Zhang, J.; Xu, Q.Z.; Gao, H.Y. Role of nitric oxide in omeprazole protection of the gastric mucosa in rats. Di Yi Jun Yi Da Xue Xue Bao 2001, 21, 926–928. [Google Scholar] [PubMed]
- West, S.D.; Helmer, K.S.; Chang, L.K.; Cui, Y.; Greeley, G.H.; Mercer, D.W. Cholecystokinin secretagogue-induced gastroprotection: Role of nitric oxide and blood flow. Am. J. Physiol. 2003, 284, G399–G410. [Google Scholar] [CrossRef]
- Yu, F.F.; Lin, X.L.; Yang, L.; Liu, H.; Wang, X.; Fang, H.; Lammi, Z.J.; Guo, X. Comparison of T-2 toxin and HT-2 toxin distributed in the skeletal system with that in other tissues of rats by acute toxicity test. Biomed. Environ. Sci. 2017, 30, 851–854. [Google Scholar] [PubMed]
- Lutsky, I.I.; Mor, N. Alimentary toxic aleukia (septic angina, endemic panmyelotoxicosis, alimentary hemorrhagic aleukia): T-2 toxin-induced intoxication of cats. Am. J. Pathol. 1981, 104, 189–191. [Google Scholar] [PubMed]
- Moriez, R.; Abdo, H.; Chaumette, T.; Faure, M.; Lardeux, B.; Neunlist, M. Neuroplasticity and neuroprotection in enteric neurons: Role of epithelial cells. Biochem. Biophys. Res. Commun. 2009, 382, 577–582. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2017, 15, 252–259. [Google Scholar] [CrossRef]
- Miller, M.J.S.; Clark, D. Nitric oxide synthase inhibition can initiate or prevent gut inflammation: Role of enzyme source. Inflamm. Res. 1994, 41, C231–C232. [Google Scholar] [CrossRef]
- Yasukawa, K.; Tokuda, H.; Tun, X.; Utsumi, H.; Yamada, K.I. The detrimental effect of nitric oxide on tissue is associated with inflammatory events in the vascular endothelium and neutrophils in mice with dextran sodium sulfate-induced colitis. Free. Radic. Res. 2012, 46, 1427–1436. [Google Scholar] [CrossRef]
- Tan, L.; Bornstein, J.C.; Anderson, C. Neurochemical and morphological phenotypes of vagal afferent neurons innervating the adult mouse jejunum. Neurogastroenterol. Motil. 2009, 21, 994–1001. [Google Scholar] [CrossRef]
- Filpa, V.; Carpanese, E.; Marchet, S.; Pirrone, C.; Conti, A.; Rainero, A.; Moro, E.; Chiaravalli, A.M.; Zucchi, I.; Moriondo, A.; et al. Nitric oxide regulates homeoprotein OTX1 and OTX2 expression in the rat myenteric plexus after intestinal ischemia-reperfusion injury. Am. J. Physiol. 2017, 312, G374–G389. [Google Scholar] [CrossRef]
- Rivera, L.R.; Poole, D.P.; Thacker, M.; Furness, J.B. The involvement of nitric oxide synthase neurons in enteric neuropathies. Neurogastroenterol. Motil. 2011, 23, 980–988. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA) Scientific opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J. 2014, 12, 3916.




{kind=link}
{kind=link}
{kind=link}
| Part of the Intestinal Wall | C Group | T2 Group | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pig 1 | Pig 2 | Pig 3 | Pig 4 | Pig 5 | Pig 1 | Pig 2 | Pig 3 | Pig 4 | Pig 5 | ||
| CML | A | 21.925 | 24.2 | 28.15 | 25.15 | 24.2 | 23.95 | 22.9 | 23.675 | 22.925 | 24.2 |
| average ± SEM | 24.73 ± 1.008 | 23.53 ± 0.265 | |||||||||
| MP | B | 502/186 | 511/214 | 506/202 | 503/186 | 512/182 | 503/250 | 511/243 | 512/246 | 507/212 | 506/222 |
| C | 37.05% | 41.88% | 39.92% | 36.98% | 35.55% | 49.7% | 47.55% | 48.05% | 41.81% | 43.87% | |
| average ± SEM | 38.28 ± 1.147% * | 46.20 ± 1.453% * | |||||||||
| OSP | B | 504/174 | 504/193 | 507/208 | 509/209 | 505/187 | 505/230 | 502/233 | 506/235 | 507/212 | 506/222 |
| C | 34.52% | 38.29% | 41,03% | 41,06% | 37,03% | 45,54% | 46,41% | 46,44% | 44,05% | 44,49% | |
| average ± SEM | 38.39 ± 1.244% * | 45.39 ± 0.488% * | |||||||||
| ISP | B | 502/185 | 501/184 | 503/191 | 506/164 | 507/166 | 500/220 | 504/222 | 505/218 | 501/226 | 500/220 |
| C | 36.85% | 36.73% | 37.97% | 32.41% | 32.74% | 44.00% | 44.05% | 43.17% | 45.11% | 44.00% | |
| average ± SEM | 35.34 ± 1.151% * | 44.07 ± 0.308% * | |||||||||
| ML | A | 7.45 | 9.675 | 11.225 | 10.25 | 9.675 | 8.3 | 9.825 | 9.675 | 8.75 | 9.225 |
| average ± SEM | 9.66 ± 0.62 | 9.16 ± 0.285 | |||||||||
| C Group | T2 Group | |
|---|---|---|
| MP | 2619.13 ± 142.09 | 2524.08 ± 123.87 |
| OSP | 1879.69 ± 95.45 | 1634.08 ± 92.95 |
| ISP | 795.83 ± 46.2 | 695.81 ± 40.42 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rychlik, A.; Gonkowski, S.; Kaczmar, E.; Obremski, K.; Calka, J.; Makowska, K. The T2 Toxin Produced by Fusarium spp. Impacts Porcine Duodenal Nitric Oxide Synthase (nNOS)-Positive Nervous Structures—The Preliminary Study. Int. J. Mol. Sci. 2020, 21, 5118. https://doi.org/10.3390/ijms21145118
Rychlik A, Gonkowski S, Kaczmar E, Obremski K, Calka J, Makowska K. The T2 Toxin Produced by Fusarium spp. Impacts Porcine Duodenal Nitric Oxide Synthase (nNOS)-Positive Nervous Structures—The Preliminary Study. International Journal of Molecular Sciences. 2020; 21(14):5118. https://doi.org/10.3390/ijms21145118
Chicago/Turabian StyleRychlik, Andrzej, Slawomir Gonkowski, Ewa Kaczmar, Kazimierz Obremski, Jaroslaw Calka, and Krystyna Makowska. 2020. "The T2 Toxin Produced by Fusarium spp. Impacts Porcine Duodenal Nitric Oxide Synthase (nNOS)-Positive Nervous Structures—The Preliminary Study" International Journal of Molecular Sciences 21, no. 14: 5118. https://doi.org/10.3390/ijms21145118
APA StyleRychlik, A., Gonkowski, S., Kaczmar, E., Obremski, K., Calka, J., & Makowska, K. (2020). The T2 Toxin Produced by Fusarium spp. Impacts Porcine Duodenal Nitric Oxide Synthase (nNOS)-Positive Nervous Structures—The Preliminary Study. International Journal of Molecular Sciences, 21(14), 5118. https://doi.org/10.3390/ijms21145118