Two Clubroot-Resistance Genes, Rcr3 and Rcr9wa, Mapped in Brassica rapa Using Bulk Segregant RNA Sequencing

 ,
, 
 , and
, and
Abstract
1. Introduction
2. Results
2.1. Inheritance of CR in B. rapa Line 96-6990-2
2.2. RNA-Seq and Assembling Short Reads from BRS-Seq for Resistance to Pathotype 3H
2.3. Mapping Rcr3 for Resistance to Pathotype 3H Using PPV
2.4. Mapping of Rcr3 Though Kompetitive Allele Specific PCR (KASP) Analysis
2.5. Gene Annotation in the Rcr3 Target Region
2.6. Mapping Rcr9wa for Resistance to Pathotype 5X
3. Discussion
4. Materials and Methods
4.1. Plant Materials for CR Genetic Mapping
4.2. Plant Inoculation with Pathotypes 3H and 5X
4.3. RNA Extraction, Illumina Library Preparation, and Sequencing
4.4. Read Mapping, Variant Analysis, and Genetic Mapping by Analysis of PPV
4.5. SNP Selection, KASP Analysis, and Linkage Analysis
4.6. Identification of Potential Candidates for the Mapped Genes
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AAFC | Agriculture and Agri-Food Canada |
| BC | Back Cross |
| BLAST | Basic Local Alignment Search Tool |
| BSA | Bulk Segregant Analysis |
| BSR-Seq | Bulk Segregant RNA sequencing |
| CC-NBS-LRR | Coiled Coil-Nucleotide Binding Site-Leucine Rich Repeat |
| cDNA | Complementary DNA |
| cM | Centimorgan |
| CR | Clubroot Resistant |
| DEG | Differential Expressed Genes |
| DNA | Deoxyribonucleic Acid |
| ETI | Effector Triggered Immunity |
| Gb | Gigabases |
| GO | Gene Ontology |
| InDels | Insertions and Deletions |
| KASP | Kompetitive Allele Specific PCR |
| Kb | Kilobase |
| MAMPs | Microbial Associated Molecular Patterns |
| MAS | Marker Assisted Selection |
| Mb | Megabase |
| NBS-LRR | Nucleotide Binding Site-Leucine Rich Repeat) |
| NGS | Next Generation Sequencing |
| NS | Non Synonymous |
| PAMPs | Pathogen-Associated Molecular Patterns |
| PCR | Polymerase Chain Reaction |
| PGRC | Plant Genetic Resource Centre |
| PPV | Percent Polymorphic Variants |
| PRRs | Pattern Recognition Receptors |
| PTI | PAMP-Triggered Immunity |
| qPCR | quantitative PCR |
| QTL | Quantitative Trait Locus |
| R bulk | Resistant bulk |
| RQI | RNA quality indicator |
| RNA | Ribonucleic Acid |
| RNA-Seq | RNA sequencing |
| S bulk | Susceptible bulk |
| S | Synonymous |
| SNP | Single Nucleotide Polymorphism |
| TIR-NBS-LRR | Toll Interleukin1 Receptor-Nucleotide Binding Site-Leucine-Rich Repeat |
References
- Dixon, G.R. The Occurrence and Economic Impact of Plasmodiophora brassicae and Clubroot Disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Voorrips, R.E. Plasmodiophora brassicae: Aspects of pathogenesis and resistance in Brassica oleracea. Euphytica 1995, 83, 139–146. [Google Scholar] [CrossRef]
- Gossen, B.D.; Adhikari, K.K.; McDonald, M.R. Effect of seeding date on development of clubroot in short-season Brassica crops. Can. J. Plant Pathol. 2012, 34, 516–523. [Google Scholar] [CrossRef]
- Donald, E.C.; Porter, I.J. Clubroot in Australia: The history and impact of Plasmodiophora brassicae in Brassica crops and research efforts directed towards its control. Can. J. Plant Pathol. 2014, 36, 66–84. [Google Scholar] [CrossRef]
- Hwang, S.-F.; Howard, R.J.; Strelkov, S.E.; Gossen, B.D.; Peng, G. Management of clubroot (Plasmodiophora brassicae) on canola (Brassica napus) in western Canada. Can. J. Plant Pathol. 2014, 36, 49–65. [Google Scholar] [CrossRef]
- Tewari, J.P.; Strelkov, S.E.; Orchard, D.; Hartman, M.; Lange, R.M.; Turkington, T.K. Identification of clubroot of crucifers on canola (Brassica napus) in Alberta. Can. J. Plant Pathol. 2005, 27, 143–144. [Google Scholar] [CrossRef]
- Cao, T.; Manolii, V.P.; Strelkov, S.E.; Hwang, S.-F.; Howard, R.J. Virulence and spread of Plasmodiophora brassicae [clubroot] in Alberta, Canada. Can. J. Plant Pathol. 2009, 31, 321–329. [Google Scholar] [CrossRef]
- Dokken-Bouchard, F.L.; Anderson, K.; Bassendowski, K.A.; Bouchard, A.; Brown, B. Survey of canola diseases in Saskatchewan, 2011. Can. Plant Dis. Surv. 2012, 92, 125–129. [Google Scholar]
- Strelkov, S.E.; Manolii, V.; Harding, M.; Hwang, S.F.; Fei, W.; Rong, S.; Burke, D.; Pugh, C.A.; Nielson, J.M.; Barnes, A.; et al. The spread of clubroot on canola in Alberta in 2014. Can. Plant Dis. Surv. 2015, 95, 155–158. [Google Scholar]
- Chittem, K.; Mansouripour, S.; Mendoza, L.E.D.R. First Report of Clubroot on Canola Caused by Plasmodiophora brassicae in North Dakota. Plant Dis. 2014, 98, 1438. [Google Scholar] [CrossRef]
- Peng, G.; Falk, K.C.; Gugel, R.K.; Franke, C.; Yu, F.; James, B.; Strelkov, S.E.; Hwang, S.-F.; McGregor, L. Sources of resistance to Plasmodiophora brassicae (clubroot) pathotypes virulent on canola. Can. J. Plant Pathol. 2014, 36, 89–99. [Google Scholar] [CrossRef]
- Rahman, H.; Peng, G.; Yu, F.; Falk, K.; Kulkarni, M.; Selvaraj, G. Genetics and breeding for clubroot resistance in Canadian spring canola (Brassica napus L.). Can. J. Plant Pathol. 2014, 36, 122–134. [Google Scholar] [CrossRef]
- Wallenhammar, A.-C.; Almquist, C.; Soderstrom, M.; Jónsson, Á. In-field distribution of Plasmodiophora brassicae measured using quantitative real-time PCR. Plant Pathol. 2012, 61, 16–28. [Google Scholar] [CrossRef]
- Strelkov, S.E.; Hwang, S.-F.; Manolii, V.P.; Cao, T.; Feindel, D. Emergence of new virulence phenotypes of Plasmodiophora brassicae on canola (Brassica napus) in Alberta, Canada. Eur. J. Plant Pathol. 2016, 145, 517–529. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- McHale, L.; Tan, X.; Koehl, P.; Michelmore, R. Plant NBS-LRR proteins: Adaptable guards. Genome Biol. 2006, 7, 212. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Kuginuki, Y.; Yoshikawa, H.; Hirai, M. Variation in Virulence of Plasmodiophora brassicae in Japan Tested with Clubroot-resistant Cultivars of Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Eur. J. Plant Pathol. 1999, 105, 327–332. [Google Scholar] [CrossRef]
- Gustafsson, M.; Falt, A.S. Genetic studies on resistance to clubroot in Brassica napus. Ann. Appl. Biol. 1986, 108, 409–415. [Google Scholar] [CrossRef]
- Diederichsen, E.; Beckmann, J.; Schondelmeier, J.; Dreyer, F. Genetics of clubroot resistance in Brassica napus ‘mendel’. Acta Hortic. 2006, 706, 307–312. [Google Scholar] [CrossRef]
- Diederichsen, E.; Frauen, M.; Linders, E.G.A.; Hatakeyama, K.; Hirai, M. Status and Perspectives of Clubroot Resistance Breeding in Crucifer Crops. J. Plant Growth Regul. 2009, 28, 265–281. [Google Scholar] [CrossRef]
- Piao, Z.; Ramchiary, N.; Lim, Y.P. Genetics of Clubroot Resistance in Brassica Species. J. Plant Growth Regul. 2009, 28, 252–264. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Niwa, T.; Kato, T.; Ohara, T.; Kakizaki, T.; Matsumoto, S. The tandem repeated organization of NB-LRR genes in the clubroot-resistant CRb locus in Brassica rapa L. Mol. Genet. Genom. 2017, 292, 397–405. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, X.; Peng, G.; Falk, K.C.; Strelkov, S.E.; Gossen, B.D. Genotyping-by-sequencing reveals three QTL for clubroot resistance to six pathotypes of Plasmodiophora brassicae in Brassica rapa. Sci. Rep. 2017, 7, 4516. [Google Scholar] [CrossRef]
- Matsumoto, E.; Yasui, C.; Ohi, M.; Tsukada, M. Linkage analysis of RFLP markers for clubroot resistance and pigmentation in Chinese cabbage (Brassica rapa ssp. pekinensis). Euphytica 1998, 104, 79–86. [Google Scholar] [CrossRef]
- Matsumoto, E.; Hayashida, N.; Sakamoto, K.; Ohi, M. Behavior of DNA Markers Linked to a Clubroot Resistance Gene in Segregating Populations of Chinese Cabbage (Brassica rapa ssp. pekinensis). J. Jpn. Soc. Hortic. Sci. 2005, 74, 367–373. [Google Scholar] [CrossRef]
- Hayashida, N.; Takabatake, Y.; Nakazawa, N.; Aruga, D.; Nakanishi, H.; Taguchi, G.; Sakamoto, K.; Matsumoto, E. Construction of a Practical SCAR Marker Linked to Clubroot Resistance in Chinese Cabbage, with Intensive Analysis of HC352b Genes. J. Jpn. Soc. Hortic. Sci. 2008, 77, 150–154. [Google Scholar] [CrossRef][Green Version]
- Piao, Z.Y.; Deng, Y.Q.; Choi, S.R.; Park, Y.J.; Lim, Y.P. SCAR and CAPS mapping of CRb, a gene conferring resistance to Plasmodiophora brassicae in Chinese cabbage (Brassica rapa ssp. pekinensis). Theor. Appl. Genet. 2004, 108, 1458–1465. [Google Scholar] [CrossRef]
- Hirai, M.; Harada, T.; Kubo, N.; Tsukada, M.; Suwabe, K.; Matsumoto, S. A novel locus for clubroot resistance in Brassica rapa and its linkage markers. Theor. Appl. Genet. 2004, 108, 639–643. [Google Scholar] [CrossRef]
- Saito, M.; Kubo, N.; Matsumoto, S.; Suwabe, K.; Tsukada, M.; Hirai, M. Fine mapping of the clubroot resistance gene, Crr3, in Brassica rapa. Theor. Appl. Genet. 2006, 114, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Saito, A.; Hayashida, N.; Taguchi, G.; Matsumoto, E. Mapping of isolate-specific QTLs for clubroot resistance in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Theor. Appl. Genet. 2008, 117, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jing, J.; Zhan, Z.; Zhang, T.; Zhang, C.; Piao, Z. Identification of Novel QTLs for Isolate-Specific Partial Resistance to Plasmodiophora brassicae in Brassica rapa. PLoS ONE 2013, 8, e85307. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Song, T.; Falk, K.C.; Zhang, X.; Liu, X.; Chang, A.; Lahlali, R.; McGregor, L.; Gossen, B.D.; Yu, F.; et al. Fine mapping of Rcr1 and analyses of its effect on transcriptome patterns during infection by Plasmodiophora brassicae. BMC Genom. 2014, 15, 1166. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Peng, G.; Liu, X.; Deora, A.; Falk, K.C.; Gossen, B.D.; McDonald, M.R.; Yu, F. Fine Mapping of a Clubroot Resistance Gene in Chinese Cabbage Using SNP Markers Identified from Bulked Segregant RNA Sequencing. Front. Plant Sci. 2017, 8, 1448. [Google Scholar] [CrossRef]
- Huang, Z.; Peng, G.; Gossen, B.D.; Yu, F. Fine mapping of a clubroot resistance gene from turnip using SNP markers identified from bulked segregant RNA-Seq. Mol. Breed. 2019, 39, 131. [Google Scholar] [CrossRef]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Matsumoto, S.; Hirai, M. Identification of two loci for resistance to clubroot (Plasmodiophora brassicae Woronin) in Brassica rapa L. Theor. Appl. Genet. 2003, 107, 997–1002. [Google Scholar] [CrossRef]
- Laila, R.; Park, J.-I.; Robin, A.H.K.; Natarajan, S.; Vijayakumar, H.; Shirasawa, K.; Isobe, S.; Kim, H.-T.; Nou, I.-S. Mapping of a novel clubroot resistance QTL using ddRAD-seq in Chinese cabbage (Brassica rapa L.). BMC Plant Biol. 2019, 19, 13. [Google Scholar] [CrossRef]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Kondo, M.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Hirai, M.; Matsumoto, S. Simple Sequence Repeat-Based Comparative Genomics Between Brassica rapa and Arabidopsis thaliana: The Genetic Origin of Clubroot Resistance. Genetics 2006, 173, 309–319. [Google Scholar] [CrossRef]
- Suwabe, K.; Suzuki, G.; Nunome, T.; Hatakeyama, K.; Mukai, Y.; Fukuoka, H.; Matsumoto, S. Microstructure of a Brassica rapa genome segment homoeologous to the resistance gene cluster on Arabidopsis chromosome 4. Breed. Sci. 2012, 62, 170–177. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Suwabe, K.; Tomita, R.N.; Kato, T.; Nunome, T.; Fukuoka, H.; Matsumoto, S. Identification and Characterization of Crr1a, a Gene for Resistance to Clubroot Disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PLoS ONE 2013, 8, e54745. [Google Scholar] [CrossRef]
- Ueno, H.; Matsumoto, E.; Aruga, D.; Kitagawa, S.; Matsumura, H.; Hayashida, N. Molecular characterization of the CRa gene conferring clubroot resistance in Brassica rapa. Plant Mol. Biol. 2012, 80, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Marone, D.; Russo, M.A.; Laidò, G.; De Leonardis, A.M.; Mastrangelo, A.M. Plant Nucleotide Binding Site–Leucine-Rich Repeat (NBS-LRR) Genes: Active Guardians in Host Defense Responses. Int. J. Mol. Sci. 2013, 14, 7302–7326. [Google Scholar] [CrossRef]
- Hasan, M.J.; Strelkov, S.E.; Howard, R.J.; Rahman, H. Screening of Brassica germplasm for resistance to Plasmodiophora brassicae pathotypes prevalent in Canada for broadening diversity in clubroot resistance. Can. J. Plant Sci. 2012, 92, 501–515. [Google Scholar] [CrossRef]
- Hasan, M.J.; Rahman, H. Genetics and molecular mapping of resistance to Plasmodiophora brassicae pathotypes 2, 3, 5, 6, and 8 in rutabaga (Brassica napus var. napobrassica). Genome 2016, 59, 805–815. [Google Scholar] [CrossRef]
- Strelkov, S.E.; Hwang, S.-F.; Manolii, V.P.; Cao, T.; Fredua-Agyeman, R.; Harding, M.W.; Peng, G.; Gossen, B.D.; McDonald, M.R.; Feindel, D. Virulence and pathotype classification of Plasmodiophora brassicae populations collected from clubroot resistant canola (Brassica napus) in Canada. Can. J. Plant Pathol. 2018, 40, 284–298. [Google Scholar] [CrossRef]
- Dakouri, A.; Zhang, X.; Peng, G.; Falk, K.C.; Gossen, B.D.; Strelkov, S.E.; Yu, F. Analysis of genome-wide variants through bulked segregant RNA sequencing reveals a major gene for resistance to Plasmodiophora brassicae in Brassica oleracea. Sci. Rep. 2018, 8, 17657. [Google Scholar] [CrossRef]
- Dean, A. On a chromosome far, far away: LCRs and gene expression. Trends Genet. 2006, 22, 38–45. [Google Scholar] [CrossRef]
- Baulcombe, D. Reaping Benefits of Crop Research. Science 2010, 327, 761. [Google Scholar] [CrossRef] [PubMed]
- Van Nimwegen, K.; Van Soest, R.A.; Veltman, J.A.; Nelen, M.R.; Van Der Wilt, G.J.; Vissers, L.E.; Grutters, J.P. Is the $1000 Genome as Near as We Think? A Cost Analysis of Next-Generation Sequencing. Clin. Chem. 2016, 62, 1458–1464. [Google Scholar] [CrossRef]
- Mammadov, J.; Aggarwal, R.; Buyyarapu, R.; Kumpatla, S. SNP Markers and Their Impact on Plant Breeding. Int. J. Plant Genom. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Takano, S.; Matsuda, S.; Kinoshita, N.; Shimoda, N.; Sato, T.; Kato, K. Genome-wide single nucleotide polymorphisms and insertion–deletions of Oryza sativa L. subsp. japonica cultivars grown near the northern limit of rice cultivation. Mol. Breed. 2014, 34, 1007–1021. [Google Scholar] [CrossRef]
- Sathya, B.; Dharshini, A.P.; Kumar, G.R. NGS meta data analysis for identification of SNP and INDEL patterns in human airway transcriptome: A preliminary indicator for lung cancer. Appl. Transl. Genom. 2014, 4, 4–9. [Google Scholar] [CrossRef]
- Scheben, A.; Batley, J.; Edwards, D. Genotyping-by-sequencing approaches to characterize crop genomes: Choosing the right tool for the right application. Plant Biotechnol. J. 2017, 15, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Li, X.; Zhu, C.; Yeh, C.-T.; Wu, W.; Takacs, E.M.; Petsch, K.A.; Tian, F.; Bai, G.; Buckler, E.S.; Muehlbauer, G.J.; et al. Genic and nongenic contributions to natural variation of quantitative traits in maize. Genome Res. 2012, 22, 2436–2444. [Google Scholar] [CrossRef]
- Pennisi, E. The Human Genome. Science 2001, 291, 1177–1180. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, X.; Huang, Z.; Chu, M.; Song, T.; Falk, K.C.; Deora, A.; Chen, Q.; Zhang, Y.; McGregor, L.; et al. Identification of Genome-Wide Variants and Discovery of Variants Associated with Brassica rapa Clubroot Resistance Gene Rcr1 through Bulked Segregant RNA Sequencing. PLoS ONE 2016, 11, e0153218. [Google Scholar] [CrossRef]
- Wang, X.; The Brassica rapa Genome Sequencing Project Consortium; Wang, H.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.-H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef]
- Chang, A.; Lamara, M.; Wei, Y.; Hu, H.; Parkin, I.A.; Gossen, B.D.; Peng, G.; Yu, F. Clubroot resistance gene Rcr6 in Brassica nigra resides in a genomic region homologous to chromosome A08 in B. rapa. BMC Plant Biol. 2019, 19, 224. [Google Scholar] [CrossRef]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef]
- Trick, M.; Adamski, N.M.; Mugford, S.G.; Jiang, C.-C.; Febrer, M.; Uauy, C. Combining SNP discovery from next-generation sequencing data with bulked segregant analysis (BSA) to fine-map genes in polyploid wheat. BMC Plant Biol. 2012, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Wit, F. The use of inbred lines in turnips breeding. Qualitas Plantarum et Materiae Vegetabiles 1966, 13, 305–310. [Google Scholar] [CrossRef]
- Van ooijen, J.V.; Voorrips, R.E. Join Map ® 3.0, Software for the Calculation of Genetic Linkage Maps, Plant Research International: Wageningen, The Netherlands, 2001.
- Kosambi, D.D. The estimation of map distances from recombination values. Ann. Eugen. 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parents and Crosses | Type | Clubroot Rating (0-3) | No. of Plants and Analysis | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | Total | R | S | χ2 | P | ||
| Pathotype 3H | ||||||||||
| 96-6990-2 | R parent | 14 | 0 | 0 | 0 | 14 | 14 | 0 | − | − |
| ACDC | S parent | 0 | 0 | 0 | 14 | 14 | 0 | 14 | − | − |
| ACDC × 96-6990-2 | F1 | 7 | 0 | 0 | 0 | 7 | 7 | 0 | − | − |
| 96-6990-2 × ACDC | F1 | 7 | 0 | 0 | 0 | 7 | 7 | 0 | − | − |
| ACDC × F1 | BC1 | 118 | 0 | 0 | 122 | 240 | 118 | 122 | 0.06 | 0.81 |
| Pathotype 5X | ||||||||||
| 96-6990-2 | R parent | 7 | 0 | 0 | 0 | 7 | 7 | 0 | − | − |
| ACDC | S parent | 0 | 0 | 0 | 2 | 7 | 0 | 7 | − | − |
| ACDC × 96-6990-2 | F1 | 14 | 0 | 0 | 0 | 14 | 14 | 0 | − | − |
| 96-6990-2 × ACDC | F1 | 14 | 0 | 0 | 0 | 14 | 14 | 0 | − | − |
| ACDC × F1 | BC1 | 77 | 0 | 0 | 60 | 137 | 77 | 60 | 2.10 | 0.15 |
| Chrom. | Ref. Genome Length (Mb) | Length of Accumulated Sequence (Mb) | Reads per Strand (×106) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Top | Bottom | Total | |||||||
| R | S | R | S | R | S | R | S | ||
| A01 | 26.9 | 782.1 | 703.1 | 5.2 | 4.7 | 5.2 | 4.6 | 10.4 | 9.3 |
| A02 | 27.0 | 676.3 | 607.9 | 4.6 | 4.1 | 4.4 | 4.0 | 9.0 | 8.1 |
| A03 | 31.9 | 1127.5 | 1012.4 | 7.5 | 6.7 | 7.4 | 6.7 | 14.9 | 13.4 |
| A04 | 19.3 | 601.4 | 548.9 | 4.1 | 3.7 | 3.9 | 3.6 | 8.0 | 7.3 |
| A05 | 25.4 | 733.6 | 668.1 | 4.9 | 4.5 | 4.8 | 4.4 | 9.7 | 8.9 |
| A06 | 25.3 | 912.2 | 799.2 | 6.1 | 5.3 | 6.0 | 5.3 | 12.1 | 10.6 |
| A07 | 25.9 | 805.1 | 732.0 | 5.4 | 4.9 | 5.3 | 4.8 | 10.7 | 9.7 |
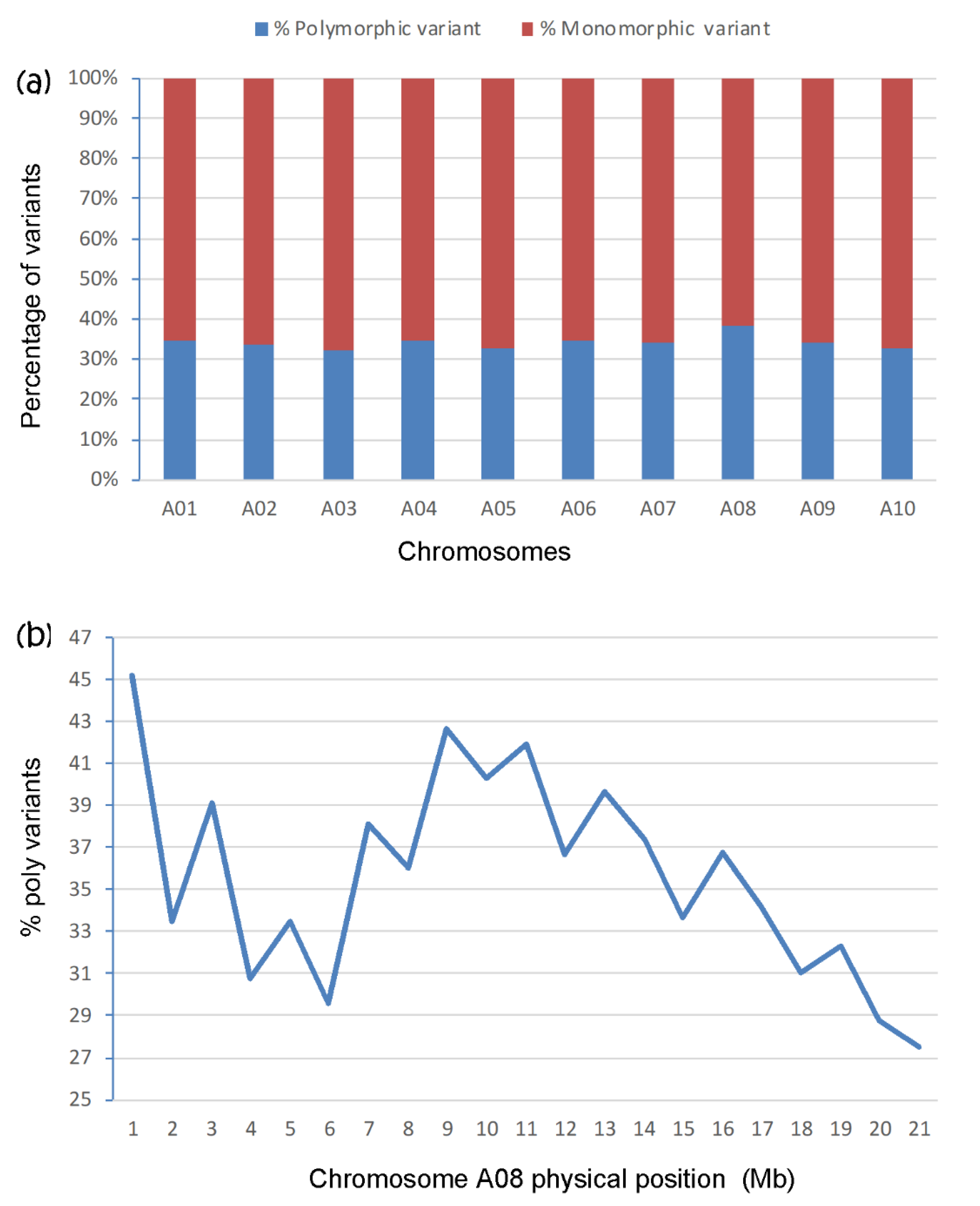
| A08 | 20.9 | 724.6 | 669.3 | 4.8 | 4.5 | 4.8 | 4.4 | 9.6 | 8.9 |
| A09 | 39.0 | 1186.2 | 1077.9 | 7.9 | 7.2 | 7.8 | 7.1 | 15.7 | 14.3 |
| A10 | 16.4 | 563.2 | 513.5 | 3.8 | 3.4 | 3.7 | 3.4 | 7.5 | 6.8 |
| Total | 257.9 | 8112.1 | 7332.3 | 54.2 | 49.0 | 53.3 | 48.2 | 107.5 | 97.1 |
| Chrom. | SNPs | InDels | Total | |||
|---|---|---|---|---|---|---|
| (×1000) | (×1000) | (×1000) | ||||
| R | S | R | S | R | S | |
| A01 | 57.5 | 56.1 | 6.1 | 6.0 | 63.6 | 62.0 |
| A02 | 53.8 | 52.1 | 5.6 | 5.4 | 59.4 | 57.5 |
| A03 | 87.1 | 86.5 | 9.1 | 9.0 | 96.2 | 95.5 |
| A04 | 37.9 | 36.2 | 3.9 | 3.8 | 41.8 | 40.0 |
| A05 | 58.0 | 57.0 | 6.2 | 6.1 | 64.2 | 63.1 |
| A06 | 60.4 | 60.5 | 7.0 | 6.8 | 67.4 | 67.4 |
| A07 | 58.2 | 56.4 | 6.0 | 5.9 | 64.2 | 62.3 |
| A08 | 48.0 | 42.8 | 5.2 | 4.9 | 53.2 | 47.7 |
| A09 | 80.4 | 79.4 | 14.4 | 14.2 | 94.8 | 93.5 |
| A10 | 44.4 | 44.0 | 4.8 | 4.7 | 49.2 | 48.7 |
| Total | 585.8 | 571.0 | 68.3 | 66.6 | 654.1 | 637.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karim, M.M.; Dakouri, A.; Zhang, Y.; Chen, Q.; Peng, G.; Strelkov, S.E.; Gossen, B.D.; Yu, F. Two Clubroot-Resistance Genes, Rcr3 and Rcr9wa, Mapped in Brassica rapa Using Bulk Segregant RNA Sequencing. Int. J. Mol. Sci. 2020, 21, 5033. https://doi.org/10.3390/ijms21145033
Karim MM, Dakouri A, Zhang Y, Chen Q, Peng G, Strelkov SE, Gossen BD, Yu F. Two Clubroot-Resistance Genes, Rcr3 and Rcr9wa, Mapped in Brassica rapa Using Bulk Segregant RNA Sequencing. International Journal of Molecular Sciences. 2020; 21(14):5033. https://doi.org/10.3390/ijms21145033
Chicago/Turabian StyleKarim, Md. Masud, Abdulsalam Dakouri, Yan Zhang, Qilin Chen, Gary Peng, Stephen E. Strelkov, Bruce D. Gossen, and Fengqun Yu. 2020. "Two Clubroot-Resistance Genes, Rcr3 and Rcr9wa, Mapped in Brassica rapa Using Bulk Segregant RNA Sequencing" International Journal of Molecular Sciences 21, no. 14: 5033. https://doi.org/10.3390/ijms21145033
APA StyleKarim, M. M., Dakouri, A., Zhang, Y., Chen, Q., Peng, G., Strelkov, S. E., Gossen, B. D., & Yu, F. (2020). Two Clubroot-Resistance Genes, Rcr3 and Rcr9wa, Mapped in Brassica rapa Using Bulk Segregant RNA Sequencing. International Journal of Molecular Sciences, 21(14), 5033. https://doi.org/10.3390/ijms21145033