Targeting Wnt/β-Catenin Pathway for Developing Therapies for Hair Loss


Abstract
1. Introduction
2. Hair Morphogenesis and Cycling
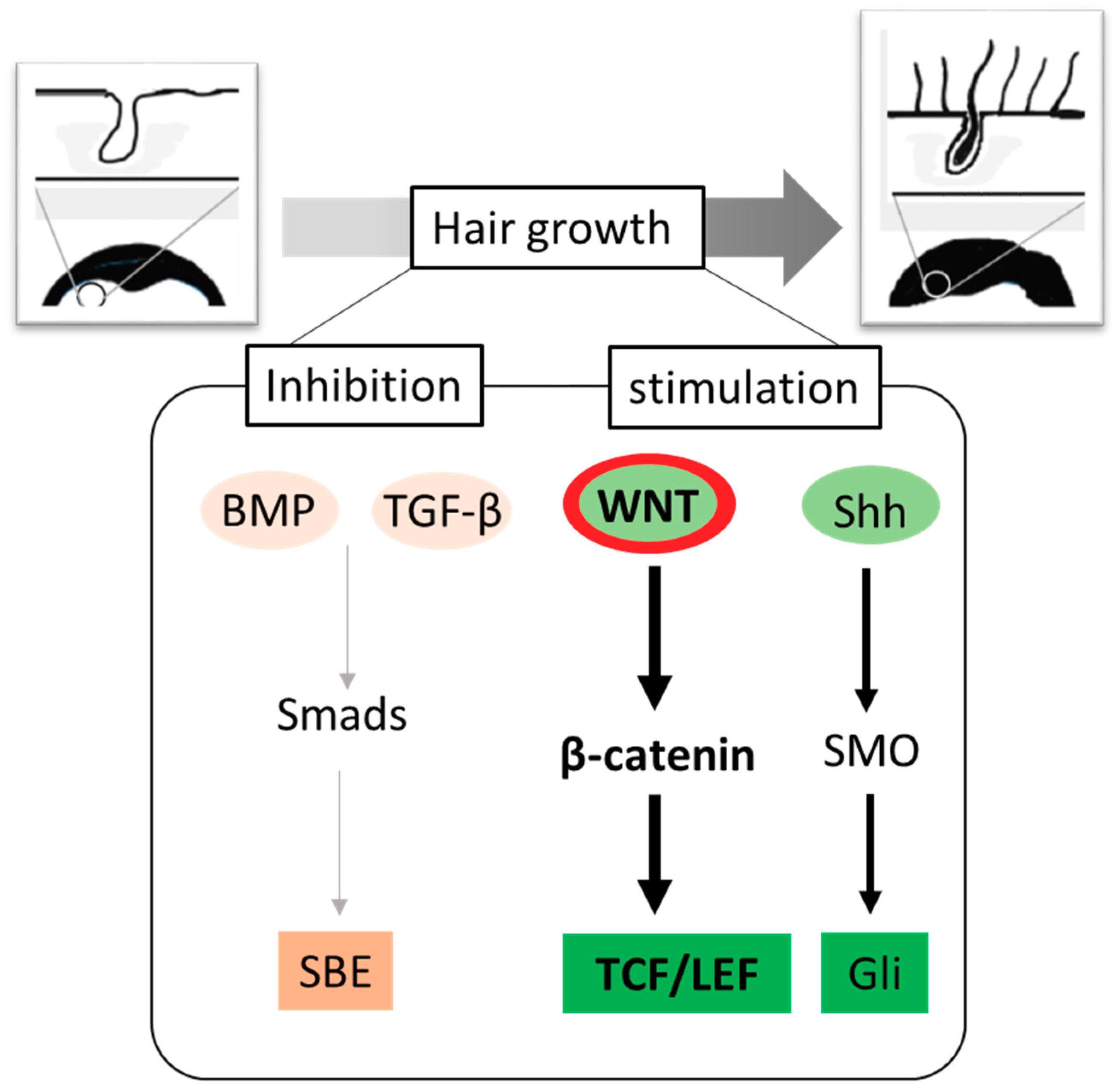
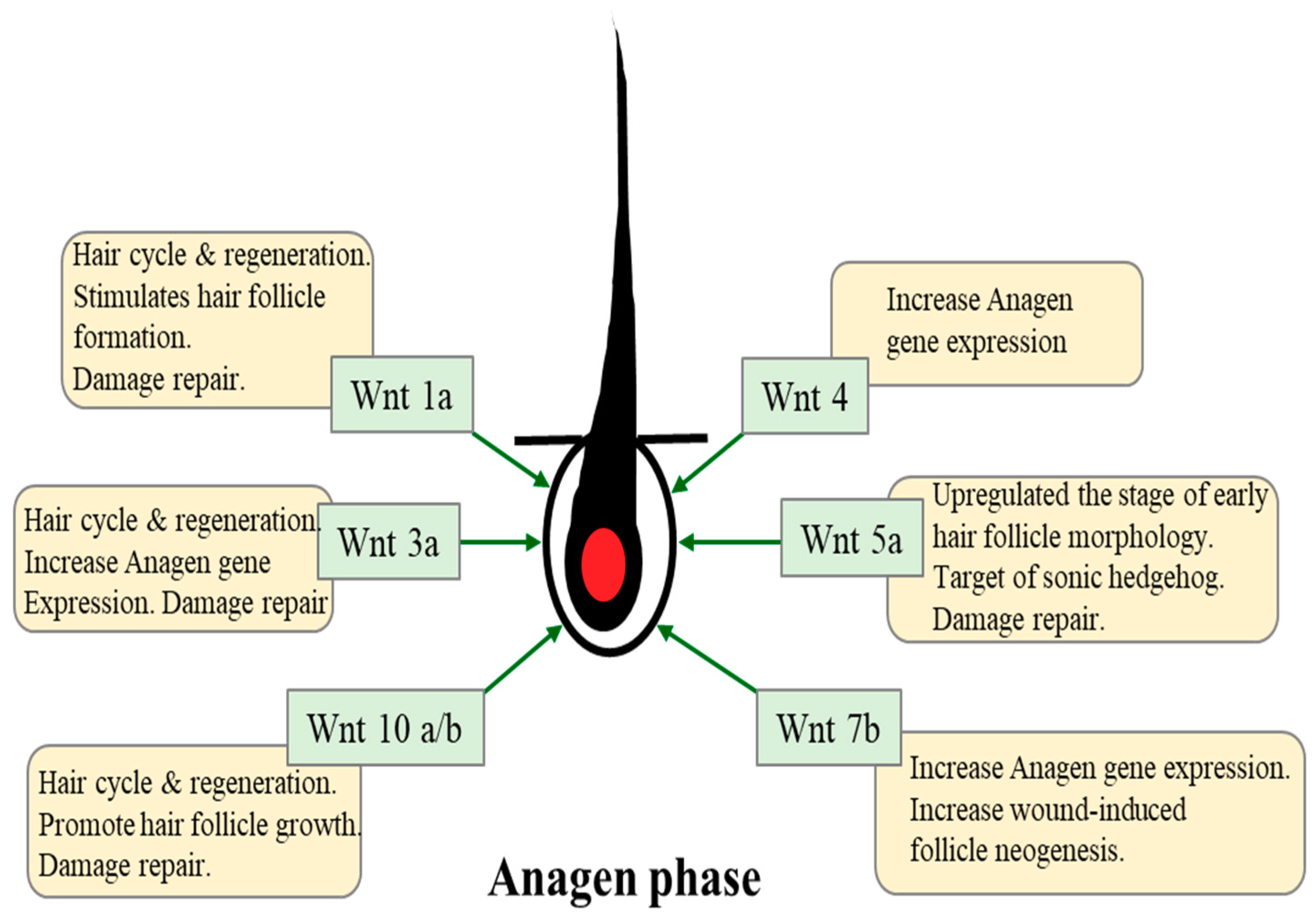
3. Role of Wnt/β-Catenin Signaling in Hair Growth
4. Progress in Developing Therapy for Hair Loss by Targeting Wnt/β-Catenin Signaling
5. Conclusions
Funding
Conflicts of Interest
References
- Mounsey, A.L.; Reed, S.W. Diagnosing and treating hair loss. Am. Fam. Physician 2009, 80, 356–362. [Google Scholar]
- Rothwell, J.T.; Phillips, C.J. Intradermal treatments and mulesing. Aust. Vet. J. 2009, 87, 118–119. [Google Scholar] [CrossRef]
- Price, V.H. Treatment of hair loss. N. Engl. J. Med. 1999, 341, 964–973. [Google Scholar] [CrossRef]
- Abdel-Rahman, S.M.; Farrand, N.; Schuenemann, E.; Stering, T.K.; Preuett, B.; Magie, R.; Campbell, A. The prevalence of infections with Trichophyton tonsurans in schoolchildren: The CAPITIS study. Pediatrics 2010, 125, 966–973. [Google Scholar] [CrossRef]
- Okamoto, M.; Ogawa, Y.; Watanabe, A.; Sugiura, K.; Shimomura, Y.; Aoki, N.; Nagasaka, T.; Tomita, Y.; Muro, Y. Autoantibodies to DFS70/LEDGF are increased in alopecia areata patients. J. Autoimmun. 2004, 23, 257–266. [Google Scholar] [CrossRef]
- Ito, C.; Saitoh, Y.; Fujita, Y.; Yamazaki, Y.; Imamura, T.; Oka, S.; Suzuki, S. Decapeptide with fibroblast growth factor (FGF)-5 partial sequence inhibits hair growth suppressing activity of FGF-5. J. Cell Physiol. 2003, 197, 272–283. [Google Scholar] [CrossRef]
- Adhirajan, N.; Ravi Kumar, T.; Shanmugasundaram, N.; Babu, M. In vivo and in vitro evaluation of hair growth potential of Hibiscus rosa-sinensis Linn. J. Ethnopharmacol. 2003, 88, 235–239. [Google Scholar] [CrossRef]
- Matsuda, H.; Yamazaki, M.; Asanuma, Y.; Kubo, M. Promotion of hair growth by ginseng radix on cultured mouse vibrissal hair follicles. Phytother. Res. 2003, 17, 797–800. [Google Scholar] [CrossRef]
- Rittmaster, R.S. Finasteride. N. Engl. J. Med. 1994, 330, 120–125. [Google Scholar]
- Gentile, P.; Garcovich, S. Systematic Review of Platelet-Rich Plasma Use in Androgenetic Alopecia Compared with Minoxidil((R)), Finasteride((R)), and Adult Stem Cell-Based Therapy. Int. J. Mol. Sci. 2020, 21, 2702. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, Y.; Song, Z.; Hao, F.; Yang, X. Identification of Wnt/beta-catenin signaling pathway in dermal papilla cells of human scalp hair follicles: TCF4 regulates the proliferation and secretory activity of dermal papilla cell. J. Dermatol. 2014, 41, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Blanpain, C.; Fuchs, E. Epidermal homeostasis: A balancing act of stem cells in the skin. Nat. Rev. Mol. Cell Biol. 2009, 10, 207–217. [Google Scholar] [CrossRef]
- Morris, R.J.; Liu, Y.; Marles, L.; Yang, Z.; Trempus, C.; Li, S.; Lin, J.S.; Sawicki, J.A.; Cotsarelis, G. Capturing and profiling adult hair follicle stem cells. Nat. Biotechnol. 2004, 22, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Trempus, C.S.; Morris, R.J.; Bortner, C.D.; Cotsarelis, G.; Faircloth, R.S.; Reece, J.M.; Tennant, R.W. Enrichment for living murine keratinocytes from the hair follicle bulge with the cell surface marker CD34. J. Investig. Dermatol. 2003, 120, 501–511. [Google Scholar] [PubMed]
- Zhang, J.; He, X.C.; Tong, W.G.; Johnson, T.; Wiedemann, L.M.; Mishina, Y.; Feng, J.Q.; Li, L. Bone morphogenetic protein signaling inhibits hair follicle anagen induction by restricting epithelial stem/progenitor cell activation and expansion. Stem Cells 2006, 24, 2826–2839. [Google Scholar] [CrossRef]
- Garza, L.A.; Yang, C.C.; Zhao, T.; Blatt, H.B.; Lee, M.; He, H.; Stanton, D.C.; Carrasco, L.; Spiegel, J.H.; Tobias, J.W.; et al. Bald scalp in men with androgenetic alopecia retains hair follicle stem cells but lacks CD200-rich and CD34-positive hair follicle progenitor cells. J. Clin. Investig. 2011, 121, 613–622. [Google Scholar] [CrossRef]
- Schmidt-Ullrich, R.; Paus, R. Molecular principles of hair follicle induction and morphogenesis. Bioessays 2005, 27, 247–261. [Google Scholar] [CrossRef]
- Houschyar, K.S.; Borrelli, M.R.; Tapking, C.; Popp, D.; Puladi, B.; Ooms, M.; Chelliah, M.P.; Rein, S.; Pforringer, D.; Thor, D.; et al. Molecular Mechanisms of Hair Growth and Regeneration: Current Understanding and Novel Paradigms. Dermatology 2020, 236, 271–280. [Google Scholar] [CrossRef]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef]
- Li, Y.H.; Zhang, K.; Ye, J.X.; Lian, X.H.; Yang, T. Wnt10b promotes growth of hair follicles via a canonical Wnt signalling pathway. Clin. Exp. Dermatol. 2011, 36, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.H.; Xiang, L.J.; Shi, H.X.; Zhang, J.; Jiang, L.P.; Cai, P.T.; Lin, Z.L.; Lin, B.B.; Huang, Y.; Zhang, H.L.; et al. Fibroblast growth factors stimulate hair growth through beta-catenin and Shh expression in C57BL/6 mice. BioMed Res. Int. 2015, 2015, 730139. [Google Scholar] [PubMed]
- Li, Y.H.; Zhang, K.; Yang, K.; Ye, J.X.; Xing, Y.Z.; Guo, H.Y.; Deng, F.; Lian, X.H.; Yang, T. Adenovirus-mediated Wnt10b overexpression induces hair follicle regeneration. J. Investig. Dermatol. 2013, 133, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.H. The secret life of the hair follicle. Trends Genet. 1992, 8, 55–61. [Google Scholar] [CrossRef]
- Huelsken, J.; Vogel, R.; Erdmann, B.; Cotsarelis, G.; Birchmeier, W. beta-Catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell 2001, 105, 533–545. [Google Scholar] [CrossRef]
- Collins, C.A.; Kretzschmar, K.; Watt, F.M. Reprogramming adult dermis to a neonatal state through epidermal activation of beta-catenin. Development 2011, 138, 5189–5199. [Google Scholar] [CrossRef]
- Kundu, J.K.; Mossanda, K.S.; Na, H.K.; Surh, Y.J. Inhibitory effects of the extracts of Sutherlandia frutescens (L.) R. Br. and Harpagophytum procumbens DC. on phorbol ester-induced COX-2 expression in mouse skin: AP-1 and CREB as potential upstream targets. Cancer Lett. 2005, 218, 21–31. [Google Scholar] [CrossRef]
- Bejsovec, A. Wnt signaling: An embarrassment of receptors. Curr. Biol. 2000, 10, R919–R922. [Google Scholar] [CrossRef]
- Wodarz, A.; Nusse, R. Mechanisms of Wnt signaling in development. Annu. Rev. Cell Dev. Biol. 1998, 14, 59–88. [Google Scholar] [CrossRef]
- Huang, S.; Zhu, X.; Liu, Y.; Tao, Y.; Feng, G.; He, L.; Guo, X.; Ma, G. Wls is expressed in the epidermis and regulates embryonic hair follicle induction in mice. PLoS ONE 2012, 7, e45904. [Google Scholar] [CrossRef]
- Andl, T.; Reddy, S.T.; Gaddapara, T.; Millar, S.E. WNT signals are required for the initiation of hair follicle development. Dev. Cell 2002, 2, 643–653. [Google Scholar] [CrossRef]
- Zhang, Y.; Tomann, P.; Andl, T.; Gallant, N.M.; Huelsken, J.; Jerchow, B.; Birchmeier, W.; Paus, R.; Piccolo, S.; Mikkola, M.L.; et al. Reciprocal requirements for EDA/EDAR/NF-kappaB and Wnt/beta-catenin signaling pathways in hair follicle induction. Dev. Cell 2009, 17, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, J.; Burgeson, R.E.; Morgan, B.A. Wnt signaling maintains the hair-inducing activity of the dermal papilla. Genes Dev. 2000, 14, 1181–1185. [Google Scholar]
- Noramly, S.; Freeman, A.; Morgan, B.A. beta-catenin signaling can initiate feather bud development. Development 1999, 126, 3509–3521. [Google Scholar] [PubMed]
- Kratochwil, K.; Dull, M.; Farinas, I.; Galceran, J.; Grosschedl, R. Lef1 expression is activated by BMP-4 and regulates inductive tissue interactions in tooth and hair development. Genes Dev. 1996, 10, 1382–1394. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, E.A.; Elgarhy, L.H.; Hasby, E.A.; Mohammad, L. Dickkopf-1 Expression in Androgenetic Alopecia and Alopecia Areata in Male Patients. Am. J. Dermatopathol. 2019, 41, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Bafico, A.; Liu, G.; Yaniv, A.; Gazit, A.; Aaronson, S.A. Novel mechanism of Wnt signalling inhibition mediated by Dickkopf-1 interaction with LRP6/Arrow. Nat. Cell Biol. 2001, 3, 683–686. [Google Scholar] [CrossRef]
- Glinka, A.; Wu, W.; Delius, H.; Monaghan, A.P.; Blumenstock, C.; Niehrs, C. Dickkopf-1 is a member of a new family of secreted proteins and functions in head induction. Nature 1998, 391, 357–362. [Google Scholar] [CrossRef]
- Mao, B.; Wu, W.; Li, Y.; Hoppe, D.; Stannek, P.; Glinka, A.; Niehrs, C. LDL-receptor-related protein 6 is a receptor for Dickkopf proteins. Nature 2001, 411, 321–325. [Google Scholar] [CrossRef]
- Semenov, M.V.; Tamai, K.; Brott, B.K.; Kuhl, M.; Sokol, S.; He, X. Head inducer Dickkopf-1 is a ligand for Wnt coreceptor LRP6. Curr. Biol. 2001, 11, 951–961. [Google Scholar] [CrossRef]
- Zorn, A.M. Wnt signalling: Antagonistic Dickkopfs. Curr. Biol. 2001, 11, R592–R595. [Google Scholar] [CrossRef]
- Kwack, M.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dickkopf 1 promotes regression of hair follicles. J. Investig. Dermatol. 2012, 132, 1554–1560. [Google Scholar] [CrossRef]
- Fu, X.; Jiang, L.; Sun, T.; Yang, Y.; Gu, X.; Sheng, Z. Expression of oncoproteins c-fos and c-jun in hypertrophic scars and chronic dermal ulcers and their regulation of basic fibroblast growth factor. Chin. Med. J. (Engl.) 2001, 114, 852–856. [Google Scholar] [PubMed]
- Chen, D.; Jarrell, A.; Guo, C.; Lang, R.; Atit, R. Dermal beta-catenin activity in response to epidermal Wnt ligands is required for fibroblast proliferation and hair follicle initiation. Development 2012, 139, 1522–1533. [Google Scholar] [CrossRef]
- Myung, P.S.; Takeo, M.; Ito, M.; Atit, R.P. Epithelial Wnt ligand secretion is required for adult hair follicle growth and regeneration. J. Investig. Dermatol. 2013, 133, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Enshell-Seijffers, D.; Lindon, C.; Kashiwagi, M.; Morgan, B.A. beta-catenin activity in the dermal papilla regulates morphogenesis and regeneration of hair. Dev. Cell 2010, 18, 633–642. [Google Scholar] [CrossRef]
- Dong, L.; Hao, H.; Xia, L.; Liu, J.; Ti, D.; Tong, C.; Hou, Q.; Han, Q.; Zhao, Y.; Liu, H.; et al. Treatment of MSCs with Wnt1a-conditioned medium activates DP cells and promotes hair follicle regrowth. Sci. Rep. 2014, 4, 5432. [Google Scholar] [CrossRef] [PubMed]
- Ouji, Y.; Yoshikawa, M.; Moriya, K.; Ishizaka, S. Effects of Wnt-10b on hair shaft growth in hair follicle cultures. Biochem. Biophys. Res. Commun. 2007, 359, 516–522. [Google Scholar] [CrossRef]
- Rajendran, R.L.; Gangadaran, P.; Seo, C.H.; Kwack, M.H.; Oh, J.M.; Lee, H.W.; Gopal, A.; Sung, Y.K.; Jeong, S.Y.; Lee, S.W.; et al. Macrophage-Derived Extracellular Vesicle Promotes Hair Growth. Cells 2020, 9, 856. [Google Scholar] [CrossRef]
- Xing, Y.Z.; Wang, R.M.; Yang, K.; Guo, H.Y.; Deng, F.; Li, Y.H.; Ye, J.X.; He, L.; Lian, X.H.; Yang, T. Adenovirus-mediated Wnt5a expression inhibits the telogen-to-anagen transition of hair follicles in mice. Int. J. Med. Sci. 2013, 10, 908–914. [Google Scholar] [CrossRef][Green Version]
- Xing, L.; Dai, Z.; Jabbari, A.; Cerise, J.E.; Higgins, C.A.; Gong, W.; de Jong, A.; Harel, S.; DeStefano, G.M.; Rothman, L.; et al. Alopecia areata is driven by cytotoxic T lymphocytes and is reversed by JAK inhibition. Nat. Med. 2014, 20, 1043–1049. [Google Scholar] [CrossRef]
- Kitagawa, T.; Matsuda, K.; Inui, S.; Takenaka, H.; Katoh, N.; Itami, S.; Kishimoto, S.; Kawata, M. Keratinocyte growth inhibition through the modification of Wnt signaling by androgen in balding dermal papilla cells. J. Clin. Endocrinol. Metab. 2009, 94, 1288–1294. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, Y.; Xing, Y.; Xu, W.; Guo, H.; Deng, F.; Ma, X.; Li, Y. The balance of Bmp6 and Wnt10b regulates the telogen-anagen transition of hair follicles. Cell Commun. Signal 2019, 17, 16. [Google Scholar] [CrossRef]
- Lin, C.M.; Yuan, Y.P.; Chen, X.C.; Li, H.H.; Cai, B.Z.; Liu, Y.; Zhang, H.; Li, Y.; Huang, K. Expression of Wnt/beta-catenin signaling, stem-cell markers and proliferating cell markers in rat whisker hair follicles. J. Mol. Histol. 2015, 46, 233–240. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, K.; Xu, M.; Andl, T.; Millar, S.E.; Boyce, S.; Zhang, Y. Activating beta-catenin signaling in CD133-positive dermal papilla cells increases hair inductivity. FEBS J. 2016, 283, 2823–2835. [Google Scholar] [CrossRef]
- Liu, S.; Leask, A. CCN2 modulates hair follicle cycling in mice. Mol. Biol. Cell 2013, 24, 3939–3944. [Google Scholar] [CrossRef]
- Zhang, H.; Nan, W.; Wang, S.; Zhang, T.; Si, H.; Yang, F.; Li, G. Epidermal Growth Factor Promotes Proliferation and Migration of Follicular Outer Root Sheath Cells via Wnt/beta-Catenin Signaling. Cell Physiol. Biochem. 2016, 39, 360–370. [Google Scholar] [CrossRef]
- Zhang, H.; Nan, W.; Wang, S.; Si, H.; Li, G. Balance between fibroblast growth factor 10 and secreted frizzled-relate protein-1 controls the development of hair follicle by competitively regulating beta-catenin signaling. Biomed. Pharmacother. 2018, 103, 1531–1537. [Google Scholar] [CrossRef]
- Premanand, A.; Rajkumari, B.R. In silico analysis of gene expression data from bald frontal and haired occipital scalp to identify candidate genes in male androgenetic alopecia. Arch. Dermatol. Res. 2019, 311, 815–824. [Google Scholar] [CrossRef]
- Huang, W.Y.; Lai, S.F.; Chiu, H.Y.; Chang, M.; Plikus, M.V.; Chan, C.C.; Chen, Y.T.; Tsao, P.N.; Yang, T.L.; Lee, H.S.; et al. Mobilizing Transit-Amplifying Cell-Derived Ectopic Progenitors Prevents Hair Loss from Chemotherapy or Radiation Therapy. Cancer Res. 2017, 77, 6083–6096. [Google Scholar] [CrossRef]
- Ahmed, N.S.; Ghatak, S.; El Masry, M.S.; Gnyawali, S.C.; Roy, S.; Amer, M.; Everts, H.; Sen, C.K.; Khanna, S. Epidermal E-Cadherin Dependent beta-Catenin Pathway Is Phytochemical Inducible and Accelerates Anagen Hair Cycling. Mol. Ther. 2017, 25, 2502–2512. [Google Scholar] [CrossRef]
- Zhou, Y.; Tang, G.; Li, X.; Sun, W.; Liang, Y.; Gan, D.; Liu, G.; Song, W.; Wang, Z. Study on the chemical constituents of nuts oil from Prunus mira Koehne and the mechanism of promoting hair growth. J. Ethnopharmacol. 2020, 10, 112831. [Google Scholar] [CrossRef] [PubMed]
- Truong, V.L.; Bak, M.J.; Lee, C.; Jun, M.; Jeong, W.S. Hair Regenerative Mechanisms of Red Ginseng Oil and Its Major Components in the Testosterone-Induced Delay of Anagen Entry in C57BL/6 Mice. Molecules 2017, 22, 1505. [Google Scholar] [CrossRef] [PubMed]
- Beoy, L.A.; Woei, W.J.; Hay, Y.K. Effects of tocotrienol supplementation on hair growth in human volunteers. Trop. Life Sci. Res. 2010, 21, 91–99. [Google Scholar] [PubMed]
- Bejaoui, M.; Villareal, M.O.; Isoda, H. beta-catenin-mediated hair growth induction effect of 3,4,5-tri-O-caffeoylquinic acid. Aging (Albany NY) 2019, 11, 4216–4237. [Google Scholar] [CrossRef]
- Lee, Y.R.; Bae, S.; Kim, J.Y.; Lee, J.; Cho, D.H.; Kim, H.S.; An, I.S.; An, S. Monoterpenoid Loliolide Regulates Hair Follicle Inductivity of Human Dermal Papilla Cells by Activating the Akt/beta-Catenin Signaling Pathway. J. Microbiol. Biotechnol. 2019, 29, 1830–1840. [Google Scholar] [CrossRef]
- Woo, H.; Lee, S.; Kim, S.; Park, D.; Jung, E. Effect of sinapic acid on hair growth promoting in human hair follicle dermal papilla cells via Akt activation. Arch. Dermatol. Res. 2017, 309, 381–388. [Google Scholar] [CrossRef]
- Kim, D.Y.; Choi, B.Y. Costunolide-A Bioactive Sesquiterpene Lactone with Diverse Therapeutic Potential. Int. J. Mol. Sci. 2019, 20, 2926. [Google Scholar] [CrossRef]
- Kim, Y.E.; Choi, H.C.; Nam, G.; Choi, B.Y. Costunolide promotes the proliferation of human hair follicle dermal papilla cells and induces hair growth in C57BL/6 mice. J. Cosmet. Dermatol. 2019, 18, 414–421. [Google Scholar] [CrossRef]
- Xing, F.; Yi, W.J.; Miao, F.; Su, M.Y.; Lei, T.C. Baicalin increases hair follicle development by increasing canonical Wnt/betacatenin signaling and activating dermal papillar cells in mice. Int. J. Mol. Med. 2018, 41, 2079–2085. [Google Scholar]
- Cheon, H.I.; Bae, S.; Ahn, K.J. Flavonoid Silibinin Increases Hair-Inductive Property Via Akt and Wnt/beta-Catenin Signaling Activation in 3-Dimensional-Spheroid Cultured Human Dermal Papilla Cells. J. Microbiol. Biotechnol. 2019, 29, 321–329. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, Q.; Nan, W.; Wang, Y.; Wang, S.; Yang, F.; Li, G. Ginkgolide B and bilobalide promote the growth and increase beta-catenin expression in hair follicle dermal papilla cells of American minks. Biofactors 2019, 45, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, H.; Jing, J.; Yu, L.; Wu, X.; Lu, Z. Morroniside regulates hair growth and cycle transition via activation of the Wnt/beta-catenin signaling pathway. Sci. Rep. 2018, 8, 13785. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Zhang, N.; Park, D.K. Topical application of Polygonum multiflorum extract induces hair growth of resting hair follicles through upregulating Shh and beta-catenin expression in C57BL/6 mice. J. Ethnopharmacol. 2011, 135, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, M.; Lin, P.; He, Y.; Yu, J.; Zhao, R. Hair Growth Promotion Activity and Its Mechanism of Polygonum multiflorum. Evid. Based Complement. Alternat. Med. 2015, 2015, 517901. [Google Scholar] [CrossRef]
- Park, P.J.; Moon, B.S.; Lee, S.H.; Kim, S.N.; Kim, A.R.; Kim, H.J.; Park, W.S.; Choi, K.Y.; Cho, E.G.; Lee, T.R. Hair growth-promoting effect of Aconiti Ciliare Tuber extract mediated by the activation of Wnt/beta-catenin signaling. Life Sci. 2012, 91, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.N.; Park, D.K.; Park, H.J. Hair growth-promoting activity of hot water extract of Thuja orientalis. BMC Complement. Altern. Med. 2013, 13, 9. [Google Scholar] [CrossRef]
- Lee, E.Y.; Choi, E.J.; Kim, J.A.; Hwang, Y.L.; Kim, C.D.; Lee, M.H.; Roh, S.S.; Kim, Y.H.; Han, I.; Kang, S. Malva verticillata seed extracts upregulate the Wnt pathway in human dermal papilla cells. Int. J. Cosmet. Sci. 2016, 38, 148–154. [Google Scholar] [CrossRef]
- Shu, Z.M.; Shu, X.D.; Li, H.Q.; Sun, Y.; Shan, H.; Sun, X.Y.; Du, R.H.; Lu, M.; Xiao, M.; Ding, J.H.; et al. Ginkgolide B Protects Against Ischemic Stroke Via Modulating Microglia Polarization in Mice. CNS Neurosci. Ther. 2016, 22, 729–739. [Google Scholar] [CrossRef]
- Kim, Y.M.; Kwon, S.J.; Jang, H.J.; Seo, Y.K. Rice bran mineral extract increases the expression of anagen-related molecules in human dermal papilla through wnt/catenin pathway. Food Nutr. Res. 2017, 61, 1412792. [Google Scholar] [CrossRef][Green Version]
- Kim, Y.E.; Choi, H.C.; Lee, I.C.; Yuk, D.Y.; Lee, H.; Choi, B.Y. 3-Deoxysappanchalcone Promotes Proliferation of Human Hair Follicle Dermal Papilla Cells and Hair Growth in C57BL/6 Mice by Modulating WNT/beta-Catenin and STAT Signaling. Biomol. Ther. (Seoul) 2016, 24, 572–580. [Google Scholar] [CrossRef]
- Legrand, J.M.D.; Roy, E.; Ellis, J.J.; Francois, M.; Brooks, A.J.; Khosrotehrani, K. STAT5 Activation in the Dermal Papilla Is Important for Hair Follicle Growth Phase Induction. J. Investig. Dermatol. 2016, 136, 1781–1791. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Garcovich, S. Advances in Regenerative Stem Cell Therapy in Androgenic Alopecia and Hair Loss: Wnt pathway, Growth-Factor, and Mesenchymal Stem Cell Signaling Impact Analysis on Cell Growth and Hair Follicle Development. Cells 2019, 8, 466. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Compound/Extract (Structure) | Experimental Model | Experimental Findings | Ref |
|---|---|---|---|
Tocotrienol | Incubation of HaCaT keratinocytes with 1 μM for 24 h Topical application (5 mg/cm2 skin) for 21 days in male C57BL/6 mice Human trial: 35 volunteers, tocotrienol capsule (50 mg) twice daily for 8 months | Reduced expression of E-cadherin. TCF3 activation by increasing nuclear translocation of β-catenin. The db/db mice showed an increase in the number of anagen hair follicles. Inhibited the expression of epidermal E-cadherin, Oct4, Sox9, Klf4, c-Myc, and Nanog. Increased 34% in hair counts. (Baseline 284.8 ± 111.3 vs. after treatment 383.1 ±120.9) | [60,63] |
3,4,5-tri-O-caffeoylquinic acid | Treatment of human DPCs with 10 μM for 48 h Topical application (1% aqueous solution) in C3H mice | Significantly enhanced the 60% ATP content. Enhanced the hair growth by approximately 40%, 80%, and 120% at Days 14, 20, and 30 after treatment. Activation of Wnt/β-catenin signaling pathway. | [64] |
Loliolide | Human DPCs treated with 20 μg/mL for 48 h | Increased spheroid size. Increased growth factor (IGF-1, VEGF.) Hair induction through the activation of AKT-mediated Wnt signaling. | [65] |
Sinapic acid | Human hair follicle DPCs incubated with 10, 50, and 100 μM of the compound | Increased hair count to the levels of 118.3%, 119.2%, and 150.5%. Activation of β-catenin, p-GSK-3β and growth factor (IGF-1, VEGF). | [66] |
Costunolide | Human hair follicle DPCs treated with 3 μM for 48 h. C57BL/6 mice treated topically with 200 µL of costunolide (3 µmol/L) twice daily for 15 days, | Promoted the cell proliferation through WNT-β-catenin, hedgehog-Gli activity and inhibition of the TGF-β1-Smad pathway. Promoted significant hair growth in mice. The diameter and depth of hair follicles are significantly higher. | [67,68] |
Baicalin | Skin and epidermal cells were isolated from the skin of a C57BL/6 mouse grafted in a 1:1 ratio by incising a wound on the back of a Balb/c-nu mouse and treated with 50 μmol or 100 μmol for 28days. | Significantly increased TCF/LEF reporter activity. Increased ALP activity and stimulates the expression of Wnt3a, Wnt5a, frizzled 7 and disheveled 2. Activation of Wnt/β-catenin signaling and increased of ALP, IGF-1 and VEGF. Significantly increased hair shafts. | [69] |
Silibinin | Human hair follicle DPCs treated with 10 μM for 48 h | Increase of 13.53% in the viability of cells, enhances the formation of DP spheroids in vitro, Activates the Wnt/β-catenin signaling pathway and increased the expression of Wnt5a and Lef1. Elevated the phosphorylation of Akt. | [70] |
Ginkgo biloba extract | Human hair follicle DPCs treated with 0, 25, 50, and 100 μM for 48 h. | Increased cell growth on time and dose dependent activities of Akt, ERK1/2, and β-catenin. | [71] |
Morroniside | Outer root sheath cell (ORSC) and DP cell incubated 10 µM for 72 h. C57BL/6 mice were injected into mouse HFs on post-depilation day: 100 µM in 100 μL (100 μL over 3 or 6 days) | Significantly increased proliferation by 1.3-fold and increased ORSC migration. Upregulated Wnt10b, β-catenin and lef1 and regulates cell growth and development partly through the Wnt/β-catenin signaling pathway. Accelerated hair cycling from telogen to anagen Delay catagen progression. | [72] |
polygonum multiflorum extract | C57BL6/N mics shaved on the dorsal skin were treated with a dose of 4.7 mg per 12 cm2 as topically for 1–4 weeks. | Hair length was significantly longer, promotes hair growth by inducing anagen phase in resting hair follicles. Induction of β-catenin and Sonic hedgehog (Shh). | [73,74] |
| Prunus mira Koehne 2,3,5,4-tetrahydroxystilbene-2-O-β-d-gluoside (TSG) Seed(nut) oil (include α-tocopherol, Vitamin E, β-sitosterol, linoleic acid, and Oleic acid.) | Crude oil 60.26 mg (cm2·day)−1 and 1/2 diluted in C57BL/6 mice were administered once every 7 days and observed on 21 days. | Increasing Wnt 10 b, β-catenin mRNA and protein expression, and GSK-3 β protein expression. Significant increase in hair length, hair growth rate, and hair weight. | [61] |
| Red ginseng oil (RGO) | Topical treatment with 10% RGO for 28 days on C57BL/6 mice | Restored hair regenerative. Upregulated Wnt/β-catenin and Shh/Gli pathways-mediated expression of genes such as β-catenin, Lef-1, Sonic hedgehog, Smoothened, Gli-1, Cyclin D1, and Cyclin E. | [62] |
| Aconitie ciliare tuber Natural Plant extracts | Human hair follicle DPCs and rat vibrissa dermal papilla cells (RvDP) treated with 5–50 µg/mL for 24 h. After dorsal skin, C57BL/6 mice were treated with topical treatment at 10 mg/mL for 5 weeks. | Significant cell growth at 10 µg/mL. Activates the Wnt/β-catenin signaling pathway by enhancing β-catenin transcription. Induced early telogen-to-anagen phase conversion of hair follicles. | [75] |
| Thuja orientalis Natural Plant extracts (Platycladus orientalis) | C57BL6/N mice shaved on the dorsal skin were treated with 5.05 mg/cm2/day as topically for 21 days | Promotes hair growth, hair length and number of hair follicles. Induction of β-catenin and Shh. | [76] |
| Malva verticilata Natural Plant extracts (Chinese mallow) | Human hair follicle DPCs treated with 20 µg/mL for 72 h. | Increased cell proliferation. Increased β-catenin and increased transcription levels (IGF-1, KGF, VEGF and HGF). | [77] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, B.Y. Targeting Wnt/β-Catenin Pathway for Developing Therapies for Hair Loss. Int. J. Mol. Sci. 2020, 21, 4915. https://doi.org/10.3390/ijms21144915
Choi BY. Targeting Wnt/β-Catenin Pathway for Developing Therapies for Hair Loss. International Journal of Molecular Sciences. 2020; 21(14):4915. https://doi.org/10.3390/ijms21144915
Chicago/Turabian StyleChoi, Bu Young. 2020. "Targeting Wnt/β-Catenin Pathway for Developing Therapies for Hair Loss" International Journal of Molecular Sciences 21, no. 14: 4915. https://doi.org/10.3390/ijms21144915
APA StyleChoi, B. Y. (2020). Targeting Wnt/β-Catenin Pathway for Developing Therapies for Hair Loss. International Journal of Molecular Sciences, 21(14), 4915. https://doi.org/10.3390/ijms21144915