Comprehensive Characterization of Secondary Metabolites from Colebrookea oppositifolia (Smith) Leaves from Nepal and Assessment of Cytotoxic Effect and Anti-Nf-κB and AP-1 Activities In Vitro

 ,
,  ,
,  and
and 
Abstract
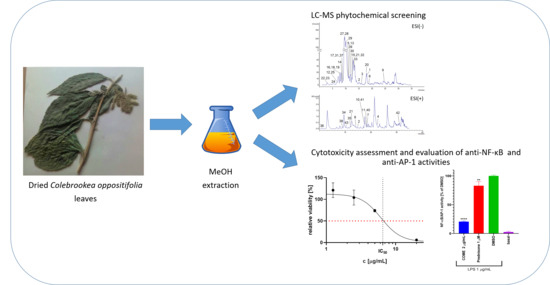
1. Introduction
2. Results
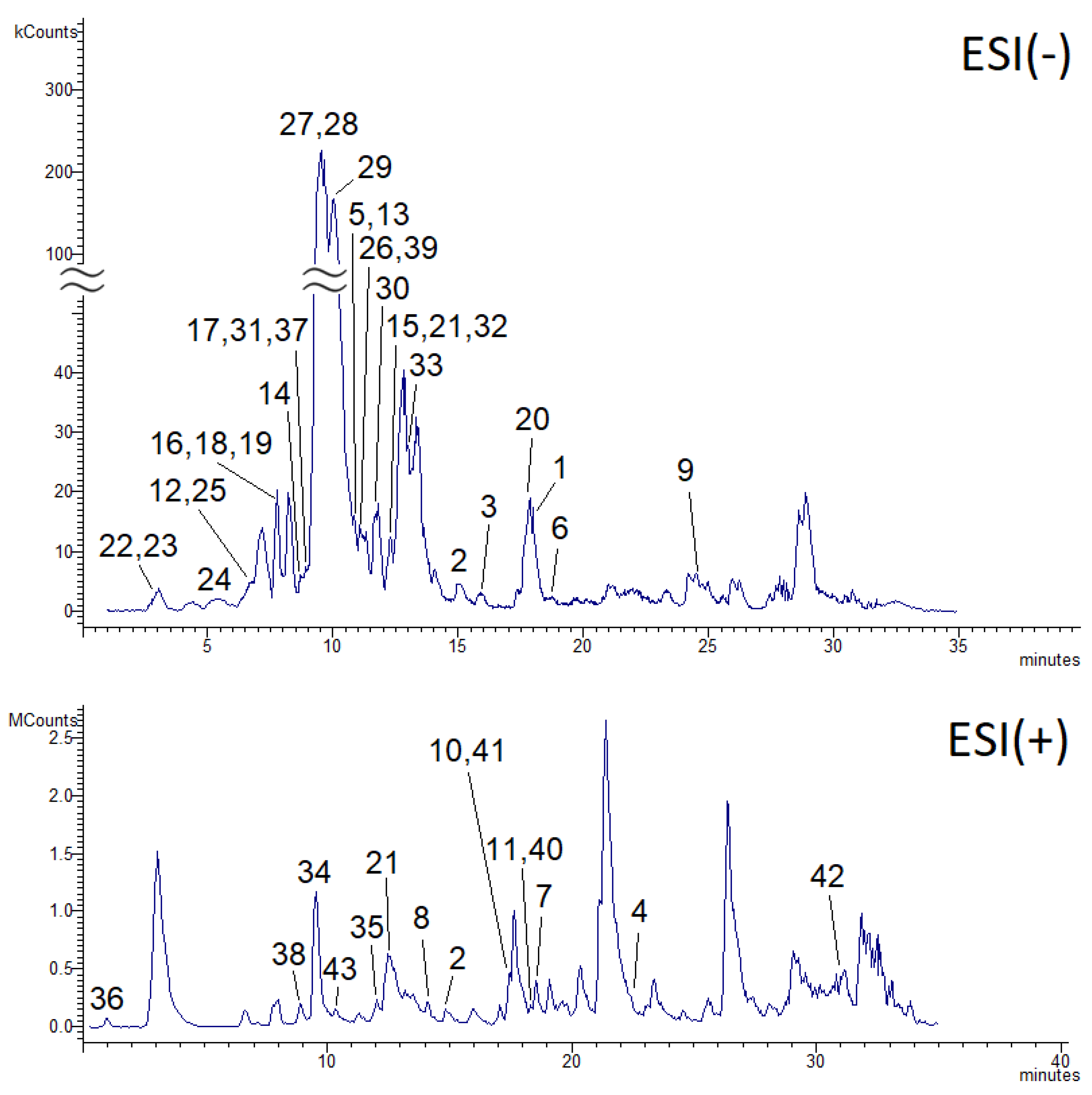
2.1. LC-MS Phytochemical Characterization of C. oppositifolia Methanol Extract
2.2. Cytotoxicity of COME
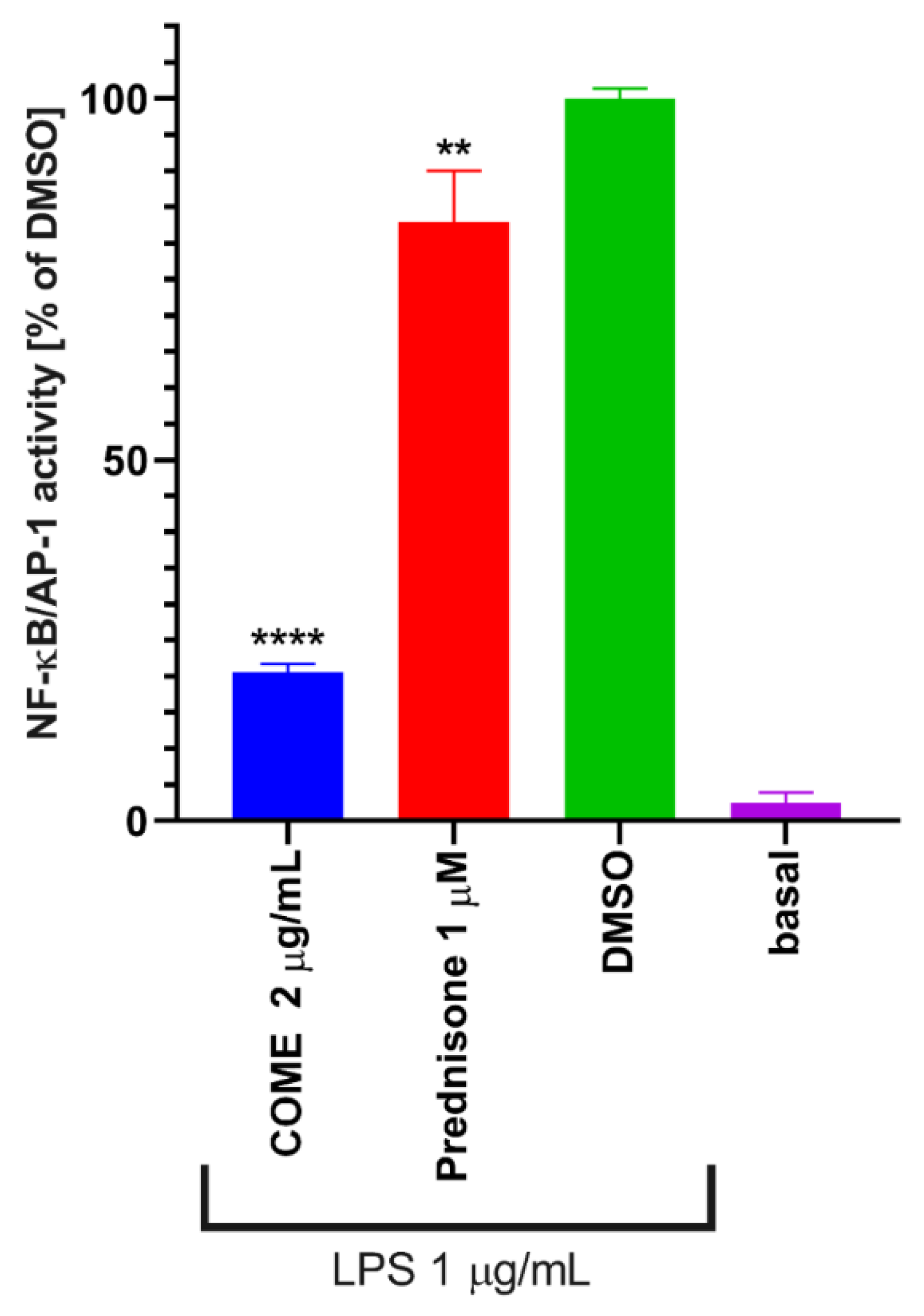
2.3. Inhibition of NF-κB/AP-1 Activities
3. Discussion
4. Materials and Methods
4.1. Plant Collection and Identification
4.2. Extraction of Plant Material
4.3. LC-MSn and UPLC (Ultra-High Performance Liquid Chromatography)-QTOF (Quadrupole-Time of Flight) Analyses of Secondary Metabolites
4.4. Cell Viability Determination
4.5. Determination of NF-κB and AP-1 Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AP-1 | Activator protein-1 |
| COME | Colebrookea Oppositifolia methanol extract |
| DW | Dry weight |
| ESI | Electrospray ionization |
| IC50 | Half maximal inhibitory concentration |
| LC | Liquid Chromatography |
| MS | Mass Spectrometry |
| NF-κB | Nuclear factor-κB |
| QTOF | Quadrupole-Time of Flight |
| R.T. | Retention time |
| UPLC | Ultra-high performance liquid chromatography |
References
- Yadav, D.K. Pharmacognostical, Phytochemical and Pharmacological profile of Colebrookea oppositifolia Smith. J. Drug Deliv. Ther. 2019, 9, 233–237. [Google Scholar] [CrossRef]
- Hansen, B.; Grierson, A.J.C.; Long, D.G. Flora of Buthan. Including a record of plants from Sikkim. Nord. J. Bot. 1989, 8, 584. [Google Scholar] [CrossRef]
- Joshi, A.R.; Joshi, K. Indigenous knowledge and uses of medicinal plants by local communities of the Kali Gandaki Watershed Area, Nepal. J. Ethnopharmacol. 2000, 73, 175–183. [Google Scholar] [CrossRef]
- Acharya, K.P.; Acharya, M. Traditional knowledge on medicinal plants used for the treatment of livestock diseases in Sardikhola VDC, Kaski, Nepal. J. Med. Plants Res. 2010, 4, 235–239. [Google Scholar]
- Viswanatha, G.L.; Venkataranganna, M.V.; Prasad, N.B.L.; Hanumanthappa, S. Chemical characterization and cerebroprotective effect of methanolic root extract of Colebrookea oppositifolia in rats. J. Ethnopharmacol. 2018, 223, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, V.; Yadav, D.K.; Gurudeva, M.; Yoganarasimhan, S. Pharmacognostical studies on the leaves of Colebrookea oppositifolia Smith. Asian J. Trad. Med. 2011, 6, 134–144. [Google Scholar]
- Yang, F.; Li, X.C.; Wang, H.Q.; Yang, C.R. Flavonoid glycosides from Colebrookea oppositifolia. Phytochemistry 1996, 42, 867–869. [Google Scholar] [CrossRef]
- Riaz, T.; Abbasi, M.A.; Shahzadi, T.; Aziz-ur-Rehman; Siddiqui, S.Z.; Ajaib, M. Colebrookia oppositifolia: A valuable source for natural antioxidants. J. Med. Plants Res. 2011, 5, 4180–4187. [Google Scholar]
- Viswanatha, G.L.; Venkataranganna, M.V.; Prasad, N.B.L. Ameliorative potential of Colebrookea oppositifolia methanolic root extract against experimental models of epilepsy: Possible role of GABA mediated mechanism. Biomed. Pharmacother. 2017, 90, 455–465. [Google Scholar] [CrossRef]
- Gupta, R.S.; Yadav, R.K.; Dixit, V.P.; Dobhal, M.P. Antifertility studies of Colebrookia oppositifolia leaf extract in male rats with special reference to testicular cell population dynamics. Fitoterapia 2001, 42, 236–245. [Google Scholar] [CrossRef]
- Panda, S.K.; Padhi, L.; Leyssen, P.; Liu, M.; Neyts, J.; Luyten, W. Antimicrobial, anthelmintic, and antiviral activity of plants traditionally used for treating infectious disease in the Similipal Biosphere Reserve, Odisha, India. Front. Pharmacol. 2017, 8, 658. [Google Scholar] [CrossRef] [PubMed]
- Chinchansure, A.; Arkile, M.; Shinde, D.; Sarkar, D.; Joshi, S. A New Dinor-cis-Labdane Diterpene and Flavonoids with Antimycobacterium Activity from Colebrookea oppositifolia. Planta Med. Lett. 2016, 3, e20–e24. [Google Scholar] [CrossRef][Green Version]
- Ali, I.; Sharma, P.; Suri, K.A.; Satti, N.K.; Dutt, P.; Afrin, F.; Khan, I.A. In vitro antifungal activities of amphotericin B in combination with acteoside, a phenylethanoid glycoside from Colebrookea oppositifolia. J. Med. Microbiol. 2011, 60, 1326–1336. [Google Scholar] [CrossRef] [PubMed]
- Khullar, M.; Sharma, A.; Wani, A.; Sharma, N.; Sharma, N.; Chandan, B.K.; Kumar, A.; Ahmed, Z. Acteoside ameliorates inflammatory responses through NFkB pathway in alcohol induced hepatic damage. Int. Immunopharmacol. 2019, 69, 109–117. [Google Scholar] [CrossRef]
- Peron, G.; Hošek, J.; Rajbhandary, S.; Pant, D.R.; Dall’Acqua, S. LC-MSn and HR-MS characterization of secondary metabolites from Hypericum japonicum Thunb. ex Murray from Nepalese Himalayan region and assessment of cytotoxic effect and inhibition of NF-κB and AP-1 transcription factors in vitro. J. Pharm. Biomed. Anal. 2019, 174, 663–673. [Google Scholar] [CrossRef]
- Shrestha, S.S.; Sut, S.; Di Marco, S.B.; Zengin, G.; Gandin, V.; De Franco, M.; Pant, D.R.; Mahomoodally, M.F.; Dall’Acqua, S.; Rajbhandary, S. Phytochemical fingerprinting and in vitro bioassays of the ethnomedicinal fern tectaria coadunata (J. Smith) C. Christensen from Central Nepal. Molecules 2019, 24, 4457. [Google Scholar] [CrossRef]
- Shrestha, S.S.; Sut, S.; Ferrarese, I.; Di Marco, S.B.; Zengin, G.; de Franco, M.; Pant, D.R.; Mahomoodally, M.F.; Ferri, N.; Biancorosso, N.; et al. Himalayan nettle girardinia diversifolia as a candidate ingredient for pharmaceutical and nutraceutical applications-phytochemical analysis and in vitro bioassays. Molecules 2020, 25, 1563. [Google Scholar] [CrossRef]
- Bost, J.; Maroon, A.; Maroon, J. Natural anti-inflammatory agents for pain relief. Surg. Neurol. Int. 2010, 1, 80. [Google Scholar] [CrossRef]
- Seca, A.M.L.; Pinto, D.C.G.A. Plant secondary metabolites as anticancer agents: Successes in clinical trials and therapeutic application. Int. J. Mol. Sci. 2018, 19, 263. [Google Scholar] [CrossRef]
- Akhtar, N.; Ihsan-ul-Haq; Mirza, B. Phytochemical analysis and comprehensive evaluation of antimicrobial and antioxidant properties of 61 medicinal plant species. Arab. J. Chem. 2018, 11, 1223–1235. [Google Scholar] [CrossRef]
- Mitreski, I.; Stanoeva, J.P.; Stefova, M.; Stefkov, G.; Kulevanova, S. Polyphenols in representative teucrium species in the flora of R. Macedonia: LC/DAD/ESI-MSn profile and content. Nat. Prod. Commun. 2014, 9, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lin, Z.; Jiang, H.; Tong, L.; Wang, H.; Chen, S. Rapid Identification and Assignation of the Active Ingredients in Fufang Banbianlian Injection Using HPLC-DAD-ESI-IT-TOF-MS. J. Chromatogr. Sci. 2016, 54, 1225–1237. [Google Scholar] [CrossRef]
- Singh, B.; Jain, S.K.; Bharate, S.B.; Kushwaha, M.; Gupta, A.P.; Vishwakarma, R.A. Simultaneous quantification of five bioactive flavonoids in high altitude plant Actinocarya tibetica by LC-ESI-MS/MS. J. AOAC Int. 2015, 98, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Martucci, M.E.P.; De Vos, R.C.H.; Carollo, C.A.; Gobbo-Neto, L. Metabolomics as a potential chemotaxonomical tool: Application in the genus Vernonia Schreb. PLoS ONE 2014, 9, e93149. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Price, W.E.; Ashton, J.; Tapsell, L.C.; Johnson, S. Identification and characterization of phenolic compounds in hydromethanolic extracts of sorghum wholegrains by LC-ESI-MSn. Food Chem. 2016, 211, 215–226. [Google Scholar] [CrossRef]
- Chen, G.; Li, X.; Saleri, F.; Guo, M. Analysis of flavonoids in Rhamnus davurica and its antiproliferative activities. Molecules 2016, 21, 1275. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Dueñas, M.; Dias, M.I.; Sousa, M.J.; Santos-Buelga, C.; Ferreira, I.C.F.R. Phenolic profiles of in vivo and in vitro grown Coriandrum sativum L. Food Chem. 2012, 132, 841–848. [Google Scholar] [CrossRef]
- Gobbo-Neto, L.; Lopes, N.P. Online identification of chlorogenic acids, sesquiterpene lactones, and flavonoids in the Brazilian arnica Lychnophora ericoides Mart. (Asteraceae) leaves by HPLC-DAD-MS and HPLC-DAD-MS/MS and a validated HPLC-DAD method for their simultaneous analysis. J. Agric. Food Chem. 2008, 56, 1193–1204. [Google Scholar] [CrossRef]
- Spínola, V.; Castilho, P.C. Evaluation of Asteraceae herbal extracts in the management of diabetes and obesity. Contribution of caffeoylquinic acids on the inhibition of digestive enzymes activity and formation of advanced glycation end-products (in vitro). Phytochemistry 2017, 143, 29–35. [Google Scholar] [CrossRef]
- El-Hagrassy, A.M.; Elkhateeb, A.; Hussein, S.R.; Abdel-Hameed, E.S.S.; Marzouk, M.M. LC-ESI-MS profile, antioxidant activity and cytotoxic screening of Oligomeris linifolia (Vahl) Macbr. (Resedaceae). J. Appl. Pharm. Sci. 2017, 7, 43–47. [Google Scholar] [CrossRef][Green Version]
- Ibrahima, R.M.; El-Halawany, A.M.; Saleh, D.O.; El Naggar, E.M.B.; EL-Shabrawy, A.E.R.O.; El-Hawary, S.S. HPLC-DAD-MS/MS profiling of phenolics from securigera securidaca flowers and its anti-hyperglycemic and anti-hyperlipidemic activities. Braz. J. Pharmacogn. 2015, 25, 134–141. [Google Scholar] [CrossRef]
- Ghouti, D.; Rached, W.; Abdallah, M.; Pires, T.C.S.P.; Calhelha, R.C.; Alves, M.J.; Abderrahmane, L.H.; Barros, L.; Ferreira, I.C.F.R. Phenolic profile and in vitro bioactive potential of Saharan Juniperus phoenicea L. and Cotula cinerea (Del) growing in Algeria. Food Funct. 2018, 9, 4664–4672. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, P. A flow-injection mass spectrometry fingerprinting method for authentication and quality assessment of Scutellaria lateriflora-based dietary supplements. Anal. Bioanal. Chem. 2011, 401, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, S.; Galasso, S.; Piccolella, S.; Kretschmer, N.; Pan, S.P.; Nocera, P.; Lettieri, A.; Bauer, R.; Monaco, P. Winter wild fennel leaves as a source of anti-inflammatory and antioxidant polyphenols. Arab. J. Chem. 2018, 11, 513–524. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Kiss, A.K. Determination of the phenolic profile and antioxidant properties of Salvia viridis L. Shoots: A comparison of aqueous and hydroethanolic extracts. Molecules 2018, 23, 1468. [Google Scholar] [CrossRef]
- Sanz, M.; De Simón, B.F.; Cadahía, E.; Esteruelas, E.; Muñoz, A.M.; Hernández, T.; Estrella, I.; Pinto, E. LC-DAD/ESI-MS/MS study of phenolic compounds in ash (Fraxinus excelsior L. and F. americana L.) heartwood. Effect of toasting intensity at cooperage. J. Mass Spectrom. 2012, 47, 905–918. [Google Scholar] [CrossRef]
- Cerulli, A.; Napolitano, A.; Masullo, M.; Hošek, J.; Pizza, C.; Piacente, S. Chestnut shells (Italian cultivar “Marrone di Roccadaspide” PGI): Antioxidant activity and chemical investigation with in depth LC-HRMS/MSn rationalization of tannins. Food Res. Int. 2020, 129, 108787. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arráez-Román, D.; Segura-Carretero, A. HPLC-DAD-ESI-MS/MS screening of bioactive components from Rhus coriaria L. (Sumac) fruits. Food Chem. 2015, 166, 179–191. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Gómez-Caravaca, A.M.; Guerra-Hernández, E.; Cerretani, L.; García-Villanova, B.; Verardo, V. Comprehensive metabolite profiling of Solanum tuberosum L. (potato) leaves by HPLC-ESI-QTOF-MS. Food Res. Int. 2018, 112, 390–399. [Google Scholar] [CrossRef]
- Marzouk, M.M.; Hussein, S.R.; Elkhateeb, A.; El-shabrawy, M.; Abdel-Hameed, E.S.S.; Kawashty, S.A. Comparative study of Mentha species growing wild in Egypt: LC-ESI-MS analysis and chemosystematic significance. J. Appl. Pharm. Sci. 2018, 8, 116–122. [Google Scholar] [CrossRef]
- Li, H.; Peng, S.Y.; Yang, D.P.; Bai, B.; Zhu, L.P.; Mu, C.Y.; Tian, Y.J.; Wang, D.M.; Zhao, Z.M. Enantiomeric Neolignans and a Sesquiterpene from Solanum erianthum and Their Absolute Configuration Assignment. Chirality 2016, 28, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.B.; Yue, T.X.; Ai, L.L.; Cheng, Y.F.; Meng, J.F.; Li, M.H.; Zhang, Z.W. Phenolic compound profiles in grape skins of Cabernet Sauvignon, Merlot, Syrah and Marselan Cultivated in the Shacheng Area (China). S. Afr. J. Enol. Vitic. 2016, 37, 132–138. [Google Scholar] [CrossRef][Green Version]
- Mukherjee, P.K.; Mukherjee, K.; Hermans-Lokkerbol, A.C.J.; Verpoorte, R.; Suresh, B. Flavonoid Content of Eupatorium glandulosum and Coolebroke oppositifolia. J. Nat. Remedies 2001, 1, 24. [Google Scholar] [CrossRef]
- Verma, S.K.; Pareek, D.; Singhal, R.; Chauhan, A.K.; Parashar, P.; Dobhal, M.P. Ferulic acid ester from colebrookea oppositifolia. Indian J. Chem. B 2012, 51, 1502–1503. [Google Scholar]
- Ansari, S.; Dobhal, M.P.; Tyagi, R.P.; Joshi, B.C.; Barar, F.S. Chemical investigation and pharmacological screening of the roots of colebrookia oppositifolia Smith. Pharmazie 1982, 37, 70. [Google Scholar]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef]
- Zürn, M.; Tóth, G.; Kraszni, M.; Sólyomváry, A.; Mucsi, Z.; Deme, R.; Rózsa, B.; Fodor, B.; Molnár-Perl, I.; Horváti, K.; et al. Galls of European Fraxinus trees as new and abundant sources of valuable phenylethanoid and coumarin glycosides. Ind. Crop Prod. 2019, 139, 111517. [Google Scholar] [CrossRef]
- Pesce, M.; Franceschelli, S.; Ferrone, A.; De Lutiis, M.A.; Patruno, A.; Grilli, A.; Felaco, M.; Speranza, L. Verbascoside down-regulates some pro-inflammatory signal transduction pathways by increasing the activity of tyrosine phosphatase SHP-1 in the U937 cell line. J. Cell. Mol. Med. 2015, 19, 1548–1556. [Google Scholar] [CrossRef]
- Ma, H.; Qin, S.; Zhao, S. Osteoarthritis is Prevented in Rats by Verbascoside via Nuclear Factor kappa B (NF-kB) Pathway Downregulation. Med. Sci. Monit. 2020, 26, e921276. [Google Scholar] [CrossRef]
- Ye, N.; Ding, Y.; Wild, C.; Shen, Q.; Zhou, J. Small molecule inhibitors targeting activator protein 1 (AP-1). J. Med. Chem. 2014, 57, 6930–6948. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.H.C.N.; Fraga, C.A.M. NF-κB-IKKβ Pathway as a Target for Drug Development: Realities, Challenges and Perspectives. Curr. Drug Targets 2018, 19, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Horng, T. Transcriptional Control of the Inflammatory Response. Nat. Rev. Immunol. 2009, 9, 692–703. [Google Scholar] [CrossRef]
- Ha, S.K.; Moon, E.; Kim, S.Y. Chrysin suppresses LPS-stimulated proinflammatory responses by blocking NF-κB and JNK activations in microglia cells. Neurosci. Lett. 2010, 485, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gong, X.B.; Huang, L.G.; Wang, Z.X.; Wan, R.Z.; Zhang, P.; Zhang, Q.Y.; Chen, Z.; Zhang, B.S. Diosmetin exerts anti-oxidative, anti-inflammatory and antiapoptotic effects to protect against endotoxin-induced acute hepatic failure in mice. Oncotarget 2017, 8, 30723–30733. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Teng, H.; Jia, Z.; Battino, M.; Miron, A.; Yu, Z.; Cao, H.; Xiao, J. Intracellular signaling pathways of inflammation modulated by dietary flavonoids: The most recent evidence. Crit. Rev. Food Sci. Nutr. 2018, 58, 2908–2924. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Meng, F.; Reece, E.A.; Zhao, Z. Modulation of nuclear factor-κB signaling and reduction of neural tube defects by quercetin-3-glucoside in embryos of diabetic mice. Am. J. Obstet. Gynecol. 2018, 219, 197.e1–197.e8. [Google Scholar] [CrossRef]
- Sun, L.R.; Zhou, W.; Zhang, H.M.; Guo, Q.S.; Yang, W.; Li, B.J.; Sun, Z.H.; Gao, S.H.; Cui, R.J. Modulation of Multiple Signaling Pathways of the Plant-Derived Natural Products in Cancer. Front. Oncol. 2019, 9, 1153. [Google Scholar] [CrossRef]
- Åkesson, C.; Lindgren, H.; Pero, R.W.; Leanderson, T.; Ivars, F. Quinic acid is a biologically active component of the Uncaria tomentosa extract C-Med 100®. Int. Immunopharmacol. 2005, 5, 219–229. [Google Scholar] [CrossRef]
- Nagasaka, R.; Chotimarkorn, C.; Shafiqul, I.M.; Hori, M.; Ozaki, H.; Ushio, H. Anti-inflammatory effects of hydroxycinnamic acid derivatives. Biochem. Biophys. Res. Commun. 2007, 358, 615–619. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Role of chlorogenic acids in controlling oxidative and inflammatory stress conditions. Nutrients 2016, 8, 16. [Google Scholar] [CrossRef]
- Ouyang, L.; Dan, Y.; Shao, Z.; Yang, S.; Yang, C.; Liu, G.; Zhou, W.; Duan, D. Effect of umbelliferone on adjuvant-induced arthritis in rats by MAPK/NF-κB pathway. Drug Des. Devel. Ther. 2019, 13, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yin, P.; Gong, P.; Lv, A.; Zhang, Z.; Liu, F. 8-Methoxypsoralen protects bovine mammary epithelial cells against lipopolysaccharide-induced inflammatory injury via suppressing JAK/STAT and NF-κB pathway. Microbiol. Immunol. 2019, 63, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.L.; Chen, P.N.; Chu, S.C.; Kuo, D.Y.; Kuo, W.H.; Chen, J.Y.; Hsieh, Y.S. Peonidin 3-glucoside inhibits lung cancer metastasis by downregulation of proteinases activities and MAPK pathway. Nutr. Cancer 2010, 11, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Ndhlala, A.R.; Ghebrehiwot, H.M.; Ncube, B.; Aremu, A.O.; Gruz, J.; Šubrtová, M.; Doležal, K.; du Plooy, C.P.; Abdelgadir, H.A.; Van Staden, J. Antimicrobial, anthelmintic activities and characterisation of functional phenolic acids of Achyranthes aspera Linn.: A medicinal plant used for the treatment of wounds and ringworm in east Africa. Front. Pharmacol. 2015, 6, 274. [Google Scholar] [CrossRef]
- D’Acquisto, F.; May, M.J.; Ghosh, S. Inhibition of Nuclear Factor Kappa B (NF-B): An Emerging Theme in Anti-Inflammatory Therapies. Mol. Interv. 2002, 2, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Mainardi, T.; Kapoor, S.; Bielory, L. Complementary and alternative medicine: Herbs, phytochemicals and vitamins and their immunologic effects. J. Allergy Clin. Immunol. 2009, 123, 283–294. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nº | Experimental m/z | Theoretical m/z | ppm | Molecular Formula | ESI(−)/ ESI(+) | MS2 # | MS3 | R.T. (min) | Tentative Identification | Chemical Class | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 253.0494 | 253.0501 | 2.9 | C15H10O4 | − | 161 145 143 121 | 18.2 | Chrysin | Flavonoid | [12] | |
| 2 | 299.0548 | 299.0556 | 2.8 | C16H11O6 | − | 284 | 256 227 | 14.9 | Diosmetin a | Flavonoid | [21] |
| 3 | 299.0546 | 299.0556 | 3.5 | C16H11O6 | − | 284 | 255 256 227 211 | 15.9 | 4′-Hydroxywogonin | Flavonoid | [22] |
| 4 | 299.0909 | 299.0919 | 3.5 | C17H15O5 | + | 284 | 256 | 23.4 | Mosloflavone | Flavonoid | [23] |
| 5 | 301.0341 | 301.0348 | 2.5 | C15H9O7 | − | 179 151 | 10.7 | Quercetin * | Flavonoid | - | |
| 6 | 313.0701 | 313.0712 | 3.7 | C17H13O6 | − | 298 299 | 19.0 | Ladanein | Flavonoid | [12] | |
| 7 | 313.1070 | 313.1076 | 2.0 | C18H17O5 | + | 298 283 269 252 | 18.4 | Retusin dimethylether | Flavonoid | Metlin | |
| 8 | 331.0824 | 331.0818 | 1.9 | C17H15O7 | + | 316 | 301 288 | 14.1 | 3,7-dimethoxy-5,3′,4′-trihydroxyflavone | Flavonoid | [24] |
| 9 | 335.0417 | 335.0403 | 4.4 | C15H11O9 | − | 291 272 263 193 | 24.6 | 6-Hydroxyampelopsin | Flavonoid | [25] | |
| 10 | 345.0961 | 345.0974 | 4.0 | C18H17O7 | + | 330 312 284 | 17.5 | Eupatorin | Flavonoid | Metlin | |
| 11 | 403.1380 | 403.1392 | 3.2 | C21H23O8 | + | 373 355 342 327 | 358 345 330 301 | 18.1 | Irigenin trimethylether | Flavonoid | Metlin |
| 12 | 461.1073 | 461.1084 | 2.5 | C22H21O11 | − | 299 301 284 | 6.9 | Diosmetin 7-O-glucoside | Flavonoid | [26] | |
| 13 | 463.0885 | 463.0877 | 1.8 | C21H19O12 | − | 301 | 10.6 | Quercetin-O-glucoside | Flavonoid | [27] | |
| 14 | 577.1544 | 577.1557 | 2.4 | C27H29O14 | − | 487 457 367 337 | 8.7 | 6,8-di-C-β-glucopyranosylchrysin | Flavonoid | [28] | |
| 15 | 593.1500 | 593.1507 | 1.3 | C27H29O15 | − | 503 473 383 353 325 | 12.5 | Apigenin-6,8-di-C-hexoside (vicenin-2) | Flavonoid | [29] | |
| 16 | 595.1651 | 595.1663 | 2.1 | C27H31O15 | − | 505 475 385 355 271 | 8.0 | Naringenin-6,8-di-C-hexoside | Flavonoid | [30] | |
| 17 | 609.1462 | 609.1456 | 1.0 | C27H29O16 | − | 301 | 9.2 | Rutin * | Flavonoid | - | |
| 18 | 639.1554 | 639.1561 | 1.2 | C28H31O17 | − | 477 459 315 | 8.1 | Isorhamnetin-O-sophoroside | Flavonoid | [31] | |
| 19 | 641.1334 | 641.1354 | 3.3 | C27H29O18 | − | 479 463 317 | 8.0 | Myricetin-dihexoside | Flavonoid | [32] | |
| 20 | 723.1698 | 723.1714 | 2.4 | C39H31O14 | − | 577 559 453 269 | 17.8 | Anisofolin a | Flavonoid | [11] | |
| 21 | 891.1591 | 891.1620 | 3.5 | C42H35O22 | − | 445 | 269 | 12.3 | Baicalin (dimer) b | Flavonoid | [33] |
| 22 | 191.0547 | 191.0556 | 5.0 | C7H11O6 | − | 173 | 3.0 | Quinic acid * | Phenolic acid | - | |
| 23 | 193.0499 | 193.0501 | 1.1 | C10H9O4 | - | 134 | 3.2 | Ferulic acid * | Phenolic acid | - | |
| 24 | 353.0866 | 353.0873 | 2.1 | C16H17O9 | - | 191 179 173 | 5.6 | Chlorogenic acid isomer 1 | Phenolic acid | [25] | |
| 25 | 353.0865 | 353.0873 | 2.4 | C16H17O9 | - | 191 179 173 | 7.2 | Chlorogenic acid isomer 2 | Phenolic acid | [25] | |
| 26 | 549.1964 | 549.1972 | 1.5 | C27H33O12 | - | 387 505 489 | 225 207 163 | 11.1 | 12-O-(6′-caffeoylhexosyl)jasmonic acid | Phenolic acid | [34] |
| 27 | 623.1958 | 623.1976 | 3.1 | C29H35O15 | - | 461 315 297 | 135 | 9.8 | Acteoside (verbascoside) | Phenolic acid | [35] |
| 28 | 623.1961 | 623.1976 | 2.5 | C29H35O15 | - | 461 315 297 | 135 | 9.9 | Isoacteoside (isoverbascoside) | Phenolic acid | [35] |
| 29 | 637.1788 | 637.1769 | 3.2 | C29H33O16 | - | 475 | 367 329 312 | 10.4 | β-oxoacteoside | Phenolic acid | [35] |
| 30 | 651.2263 | 651.2289 | 4.2 | C31H39O15 | - | 505 475 457 329 | 11.7 | Martynoside | Phenolic acid | [35] | |
| 31 | 653.2066 | 653.2082 | 2.6 | C30H37O16 | - | 621 491 459 | 9.3 | β-methoxylverbascoside | Phenolic acid | [36] | |
| 32 | 951.0719 | 951.0739 | 2.2 | C41H27O27 | - | 789 475 | 12.6 | HHDP-valoneoyl-glucose isomer 1 | Phenolic acid | [37] | |
| 33 | 951.0718 | 951.0739 | 2.2 | C41H27O27 | - | 789 475 | 13.3 | HHDP-valoneoyl-glucose isomer 2 | Phenolic acid | [37] | |
| 34 | 163.0391 | 163.0395 | 2.6 | C9H7O3 | + | 145 | 118 117 | 9.9 | Umbelliferone | Coumarin | [38] |
| 35 | 217.0497 | 217.0501 | 1.9 | C12H9O4 | + | 202 189 173 161 | 174 146 | 12.3 | Methoxsalen | Coumarin | Metlin |
| 36 | 132.0810 | 132.0813 | 2.4 | C9H10N | + | 91 | 1.0 | 3-methylindole | Other | Metlin | |
| 37 | 161.0441 | 161.0450 | 5.9 | C6H9O5 | - | 117 | 9.4 | Hydroxymethylglutaric acid | Other | [39] | |
| 38 | 201.1020 | 201.1028 | 4.2 | C12H13N2O | + | 160 | 8.9 | Harmalol | Other | Metlin | |
| 39 | 387.1644 | 387.1655 | 3.0 | C18H27O9 | - | 225 207 163 | 11.1 | 12-Hydroxyjasmonic acid glucoside | Other | [40] | |
| 40 | 403.1751 | 403.1756 | 1.3 | C22H27O7 | + | 373 342 | 18.3 | (7S,8R,7′E)-4-hydroxy-3,5,5′,9′-tetramethoxy-4′,7-epoxy-8,3′-neo-lign-7′-en-9-ol | Other | [41] | |
| 41 | 403.1750 | 403.1756 | 1.6 | C22H27O7 | + | 373 342 | 17.9 | (7S,8R,7′E)-4-hydroxy-3,5,5′,9′-tetramethoxy-4′,7-epoxy-8,3′-neo-lign-7′-en-9-ol isomer | Other | [41] | |
| 42 | 417.3355 | 417.3368 | 3.3 | C27H45O3 | + | 400 226 212 | 31.1 | Neotigogenin | Other | Metlin | |
| 43 | 463.1233 | 463.1240 | 1.6 | C22H23O11 | + | 301 258 | 10.3 | Peonidin-3-glucoside | Other | [42] |
| ESI(−)/ ESI(+) | Nº | [M-H]−/ [M+H]+ | Compound | mg·g−1 DW * |
|---|---|---|---|---|
| − | 1 | 253 | Chrysin | 0.53 ± 0.01 |
| 2 | 299 | Diosmetin | 0.75 ± 0.02 | |
| 3 | 299 | 4′-Hydroxywogonin | 0.61 ± 0.01 | |
| 5 | 301 | Quercetin | 0.46 ± 0.00 | |
| 6 | 313 | Ladanein | 0.53 ± 0.01 | |
| 9 | 335 | 6-Hydroxyampelopsin | 0.54 ± 0.01 | |
| 12 | 461 | Diosmetin 7-O-glucoside | 0.47 ± 0.00 | |
| 13 | 463 | Quercetin-O-glucoside | 0.66 ± 0.00 | |
| 14 | 577 | 6,8-di-C-β-glucopyranosylchrysin | 0.66 ± 0.02 | |
| 15 | 593 | Apigenin-6,8-di-C-hexoside | 0.54 ± 0.01 | |
| 16 | 595 | Naringenin-6,8-di-C-hexoside | 0.56 ± 0.00 | |
| 17 | 609 | Rutin | 0.51 ± 0.01 | |
| 18 | 639 | Isorhamnetin-O-sophoroside | 0.53 ± 0.01 | |
| 19 | 641 | Myricetin-dihexoside | 0.65 ± 0.01 | |
| 20 | 723 | Anisofolin a | 1.87 ± 0.01 | |
| 21 | 891 | Baicalin (dimer) | 0.62 ± 0.00 | |
| + | 4 | 299 | Mosloflavone | 0.51 ± 0.01 |
| 7 | 313 | Retusin dimethylether | 0.23 ± 0.01 | |
| 8 | 331 | 3,7-dimethoxy-5,3′,4′-trihydroxyflavone | 0.30 ± 0.02 | |
| 10 | 345 | Eupatorin | 0.51 ± 0.01 | |
| 11 | 403 | Irigenin trimethylether | 0.21 ± 0.01 | |
| Total | 12.23 ± 0.03 |
| Nº | [M-H]- | Compound | mg·g−1 DW * |
|---|---|---|---|
| 22 | 191 | Quinic acid | 0.13 ± 0.01 |
| 23 | 193 | Ferulic acid | 0.02 ± 0.01 |
| 24 | 353 | Chlorogenic acid isomer 1 | 0.74 ± 0.06 |
| 25 | 353 | Chlorogenic acid isomer 2 | 0.10 ± 0.01 |
| 26 | 549 | 12-O-(6′-caffeoylhexosyl)jasmonic acid | 0.48 ± 0.08 |
| 27 + 28 | 623 | Acteoside + Isoacteoside | 24.71 ± 0.10 |
| 29 | 637 | β-oxoacteoside | 0.81 ± 0.03 |
| 30 | 651 | Martynoside | 0.54 ± 0.07 |
| 31 | 653 | β-methoxylverbascoside | 0.99 ± 0.04 |
| 32 | 951 | HHDP-valoneoyl-glucose isomer 1 | 2.17 ± 0.07 |
| 33 | 951 | HHDP-valoneoyl-glucose isomer 2 | 1.64 ± 0.05 |
| Total | 32.33 ± 0.51 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peron, G.; Hošek, J.; Prasad Phuyal, G.; Raj Kandel, D.; Adhikari, R.; Dall’Acqua, S. Comprehensive Characterization of Secondary Metabolites from Colebrookea oppositifolia (Smith) Leaves from Nepal and Assessment of Cytotoxic Effect and Anti-Nf-κB and AP-1 Activities In Vitro. Int. J. Mol. Sci. 2020, 21, 4897. https://doi.org/10.3390/ijms21144897
Peron G, Hošek J, Prasad Phuyal G, Raj Kandel D, Adhikari R, Dall’Acqua S. Comprehensive Characterization of Secondary Metabolites from Colebrookea oppositifolia (Smith) Leaves from Nepal and Assessment of Cytotoxic Effect and Anti-Nf-κB and AP-1 Activities In Vitro. International Journal of Molecular Sciences. 2020; 21(14):4897. https://doi.org/10.3390/ijms21144897
Chicago/Turabian StylePeron, Gregorio, Jan Hošek, Ganga Prasad Phuyal, Dharma Raj Kandel, Rameshwar Adhikari, and Stefano Dall’Acqua. 2020. "Comprehensive Characterization of Secondary Metabolites from Colebrookea oppositifolia (Smith) Leaves from Nepal and Assessment of Cytotoxic Effect and Anti-Nf-κB and AP-1 Activities In Vitro" International Journal of Molecular Sciences 21, no. 14: 4897. https://doi.org/10.3390/ijms21144897
APA StylePeron, G., Hošek, J., Prasad Phuyal, G., Raj Kandel, D., Adhikari, R., & Dall’Acqua, S. (2020). Comprehensive Characterization of Secondary Metabolites from Colebrookea oppositifolia (Smith) Leaves from Nepal and Assessment of Cytotoxic Effect and Anti-Nf-κB and AP-1 Activities In Vitro. International Journal of Molecular Sciences, 21(14), 4897. https://doi.org/10.3390/ijms21144897