Molecular Cloning, Characterization, and Expression Regulation of Acyl-CoA Synthetase 6 Gene and Promoter in Common Carp Cyprinus carpio



Abstract

1. Introduction
2. Results
2.1. Sequence and Phylogenetic Analyses of Common Carp acsl6 cDNA
2.2. Tissue Distribution of the Common Carp acsl6
2.3. The Structural Features of Common Carp acsl6 Gene Promoter
2.4. Demonstration of Binding of Muscle Nucleus Proteins to the Core Regulatory Region by EMSA
2.5. Identification of Cis-Acting Elements in the Core Regulatory Region of Promoter
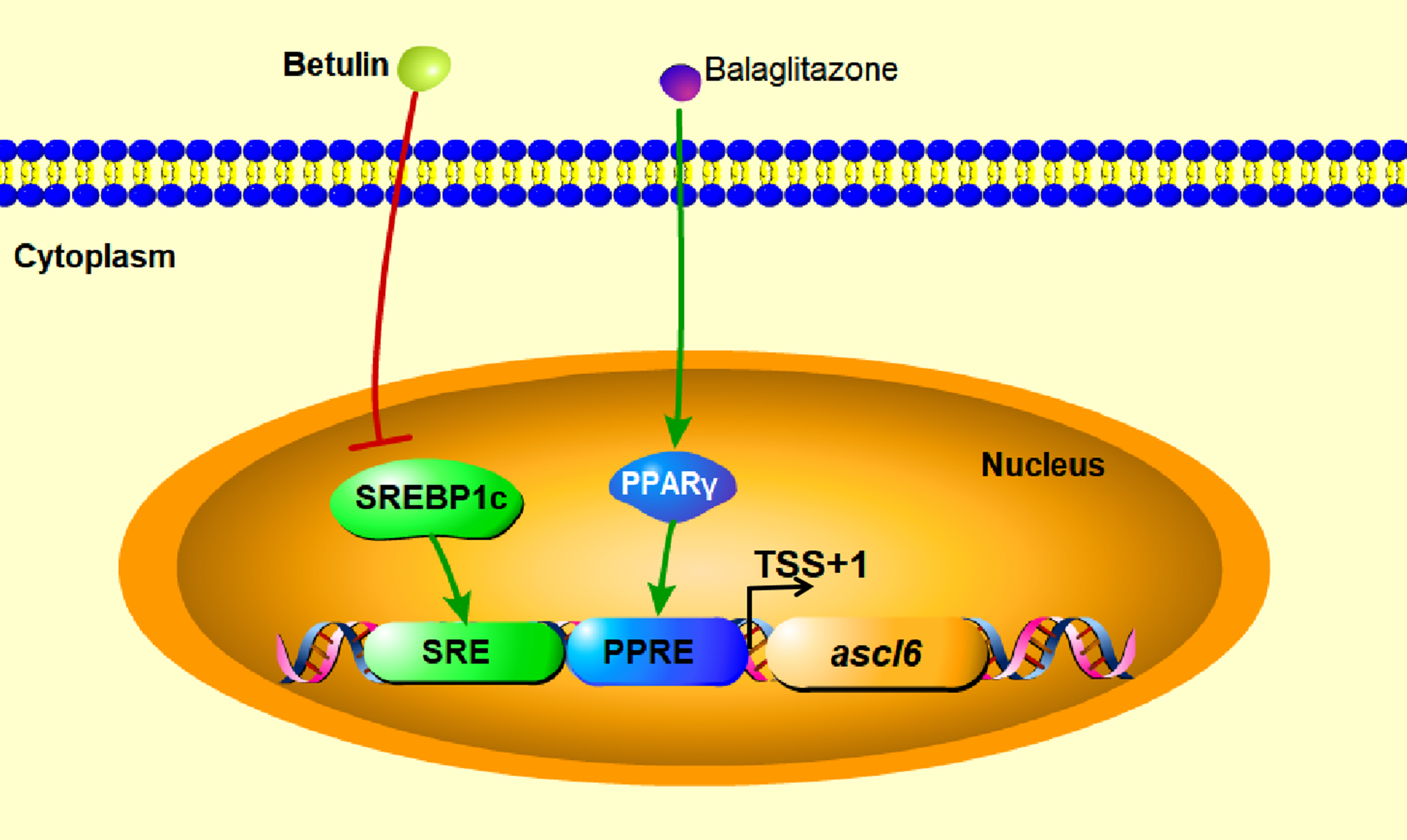
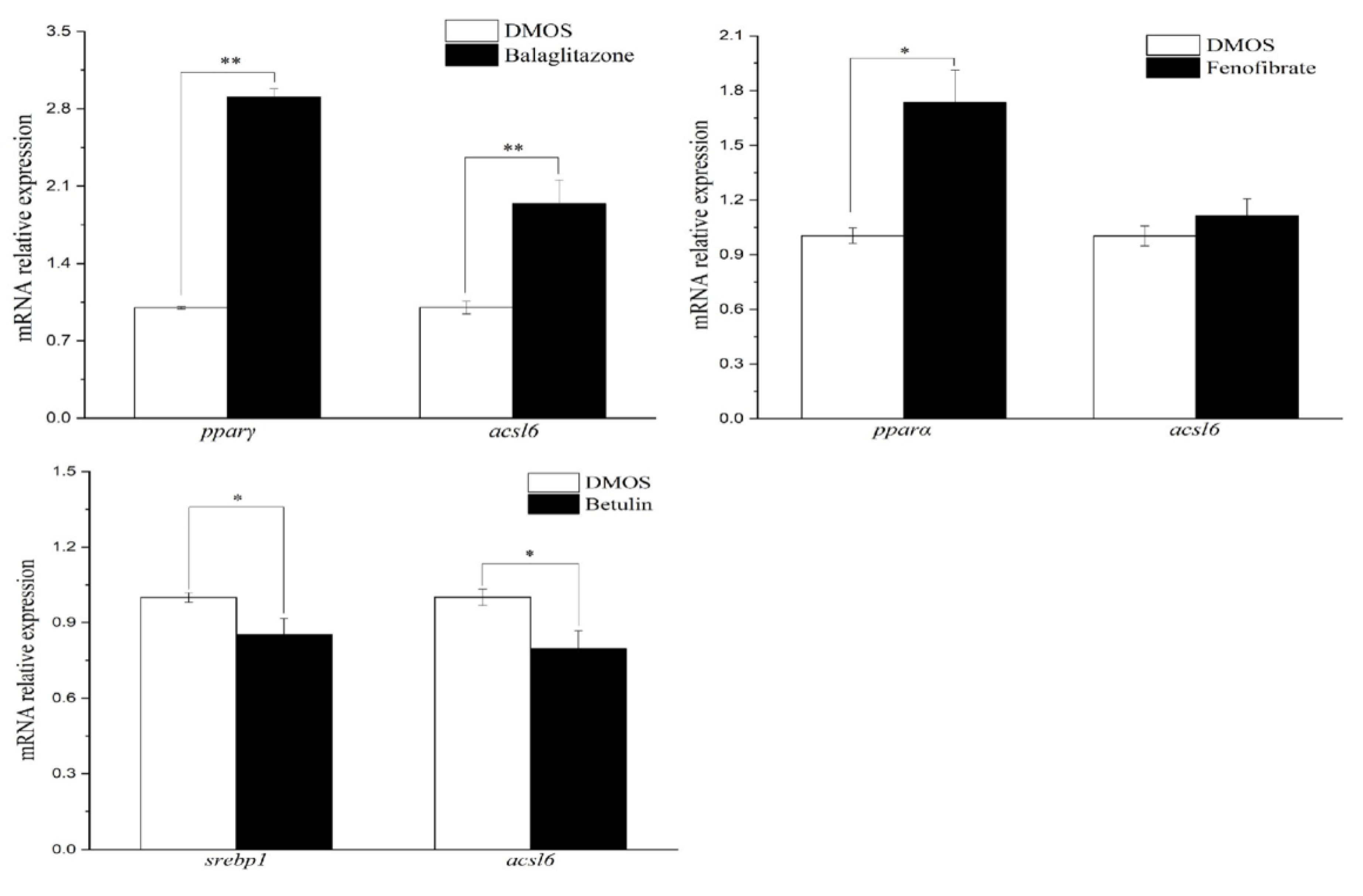
2.6. Effects of TFs Agonists/Antagonist on TFs and acsl6 Expression and Fatty Acid Composition in Muscle
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Fish and Tissue Collection
4.3. Cloning of acsl6 cDNA
4.4. Sequence and Phylogenetic Analysis of acsl6
4.5. Tissue Distribution of acsl6 Transcripts
4.6. Cloning of acsl6 Gene Promoter and Construction of Deletion Mutants
4.7. Site-Directed Mutagenesis of acsl6 Core Promoter
4.8. Cell Culture, Transfection, and Dual Luciferase Assay
4.9. Electrophoretic Mobility Shift Assay (EMSA)
4.10. Intraperitoneal Injection Experiments
4.11. Analysis of Fatty Acids Composition
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALA | α-linolenic acid (18:3n-3) |
| AP1 | activated protein 1 |
| ARA | arachidonic acid (20:4n-6) |
| CDX1 | caudal type homeobox transcription factor 1 |
| C/EBPα | CCAAT enhancer binding protein |
| DHA | docosahexaenoic acid (22:6n-3) |
| EMSA | electrophoresis mobility shift assay |
| EPA | eicosapentaenoic acid (20:5n-3) |
| FAME | fatty acid methyl esters |
| FO | fish oil |
| FOXO1 | forkhead transcription factor 1 |
| GC | Gas Chromatography |
| HEK | 293T cell human embryonic kidney cell line |
| LA | linoleic acid |
| LC-PUFA | long-chain polyunsaturated fatty acids |
| LXR | liver X receptor |
| PPARα | peroxisome proliferator activated receptor α |
| PPARγ | peroxisome proliferator activated receptor γ |
| SP1 | stimulatory protein 1 |
| SRE | sterol regulatory element |
| SREBP1c | sterol-regulatory element binding protein 1c |
| TBP | ATA box binding protein |
| TSS | transcription start site |
| YY1 | Yin Yang 1 |
References
- Shahidi, F.; Ambigaipalan, P. Omega-3 polyunsaturated fatty acids and their health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- Cardoso, C.; Afonso, C.; Bandarra, N.M. Dietary DHA and health: Cognitive function ageing. Nutr. Res. Rev. 2016, 29, 281–294. [Google Scholar] [CrossRef]
- Elagizi, A.; Lavie, C.J.; Marshall, K.; DiNicolantonio, J.J.; O’Keefe, J.H.; Milani, R.V. Omega-3 polyunsaturated fatty acids and cardiovascular health: A comprehensive review. Prog. Cardiovasc. Dis. 2018, 61, 76–85. [Google Scholar] [CrossRef]
- West, L.; Pierce, S.R.; Yin, Y.; Fang, Z.; Zhou, C.; Bae-Jump, V.L. Docosahexaenoic acid (DHA), an omega-3 fatty acid, inhibits ovarian cancer growth and adhesion. Gynecol. Oncol. 2018, 149, 63. [Google Scholar] [CrossRef]
- Setty, B.Y.; Betal, S.G.; Miller, R.E.; Brown, D.S.; Meier, M.; Cahill, M.; Stuart, M.J. Relationship of Omega-3 fatty acids DHA and EPA with the inflammatory biomarker hs-CRP in children with sickle cell anemia. Prostag. Leukotr. Essent. 2019, 146, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Agostoni, C.; Bresson, J.L.; Fairweather, T.S.; Flynn, A.; Golly, I.; Korhonen, H.; Moseley, B. Scientific opinion on the tolerable upper intake level of eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA): EFSA panel on dietetic products, nutrition and allergies (NDA). EFSA. J. 2012, 10, 2815. [Google Scholar]
- Tocher, D.R. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Oboh, A.; Kabeya, N.; Carmona-Antoñanzas, G.; Castro, L.F.C.; Dick, J.R.; Tocher, D.R.; Monroig, Ó. Two alternative pathways for docosahexaenoic acid (DHA, 22: 6n-3) biosynthesis are widespread among teleost fish. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Dong, Y.; Zhao, J.; Chen, J.; Wang, S.; Liu, Y.; Zhang, Q.; You, C.; Monroig, Ó.; Tocher, D.R.; Li, Y. Cloning and characterization of 6/5 fatty acyl desaturase (Fad) gene promoter in the marine teleost Siganus canaliculatus. Gene 2018, 647, 174–180. [Google Scholar] [CrossRef]
- Chen, C.; Wang, S.; Zhang, M.; Chen, B.; You, C.; Xie, D.; Liu, Y.; Zhang, Q.; Zhang, J.; Monroig, Ó.; et al. miR-24 is involved in vertebrate LC-PUFA biosynthesis as demonstrated in marine teleost siganus canaliculatus. BBA-Mol. Cell Biol. 2019, 1864, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, J.; Dong, Y.; Yin, Z.; Li, Y.; Liu, Y.; You, C.; Monroig, Ó.; Tocher, D.R.; Wang, S. Sp1 is Involved in Vertebrate LC-PUFA Biosynthesis by Upregulating the Expression of Liver Desaturase and Elongase Genes. Int. J. Mol. Sci. 2019, 20, 5066. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. New insights into the roles of proteins and lipids in membrane transport of fatty acids. Prostaglandins Leukot. Essent. Fatty Acids 2007, 77, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Hanebutt, F.L.; Demmelmair, H.; Schiessl, B.; Larqué, E.; Koletzko, B. Long-chain polyunsaturated fatty acid (LC-PUFA) transfer across the placenta. Clin. Nutr. 2008, 27, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Schwenk, R.W.; Holloway, G.P.; Luiken, J.J.; Bonen, A.; Glatz, J.F. Fatty acid transport across the cell membrane: Regulation by fatty acid transporters. Prostaglandins Leukot. Essent. Fatty Acids 2010, 82, 149–154. [Google Scholar] [CrossRef]
- Ellis, J.M.; Frahm, J.L.; Li, L.O.; Coleman, R.A. Acyl-coenzyme A synthetases in metabolic control. Curr. Opin. Lipidol. 2010, 21, 212. [Google Scholar] [CrossRef]
- Lopes-Marques, M.; Cunha, I.; Reis-Henriques, M.A.; Santos, M.M.; Castro, L.F.C. Diversity and history of the long-chain acyl-CoA synthetase (Acsl) gene family in vertebrates. BMC Evol. Biol. 2013, 13, 271. [Google Scholar] [CrossRef]
- Fernandez, R.F.; Kim, S.Q.; Zhao, Y.; Foguth, R.M.; Weera, M.M.; Counihan, J.L.; Ellis, J.M. Acyl-CoA synthetase 6 enriches the neuroprotective omega-3 fatty acid DHA in the brain. Proc. Natl. Acad. Sci. USA 2018, 115, 12525–12530. [Google Scholar] [CrossRef]
- Fernandez, R.F.; Kim, S.Q.; Ellis, J.M. Acyl-CoA Synthetase 6 Mediates Brain Docosahexaenoic Acid (DHA) Enrichment and Neuroprotection. FASEB J. 2019, 33, 488. [Google Scholar]
- Ellis, J.M.; Fernandez, R.F.; Zhao, Y.; Counihan, J.L.; Nomura, D.K.; Chester, J.A. Long-Chain Acyl-CoA synthetase 6 deficiency reduces the omega-3 fatty acid DHA in the brain and disrupts motor control. FASEB J. 2018, 32, 521–539. [Google Scholar]
- Shishikura, K.; Kuroha, S.; Matsueda, S.; Iseki, H.; Matsui, T.; Inoue, A.; Arita, M. Acyl-CoA synthetase 6 regulates long-chain polyunsaturated fatty acid composition of membrane phospholipids in spermatids and supports normal spermatogenic processes in mice. FASEB J. 2019, 33, 14194–14203. [Google Scholar] [CrossRef] [PubMed]
- Hale, B.J.; Fernandez, R.F.; Kim, S.Q.; Diaz, V.D.; Jackson, S.N.; Liu, L.; Ellis, J.M. Acyl-CoA synthetase 6 enriches seminiferous tubules with the ω-3 fatty acid docosahexaenoic acid and is required for male fertility in the mouse. J. Biol. Chem. 2019, 294, 14394–14405. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, B.G.; Sampaio, I.H.; Bomfim, L.H.; Queiroz, A.L.; Silveira, L.R.; Souza, A.O.; Neufer, P.D. Long-chain acyl-CoA synthetase 6 regulates lipid synthesis and mitochondrial oxidative capacity in human and rat skeletal muscle. J. Physiol. 2017, 595, 677–693. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Bu, S. Suppression of long chain acyl-CoA synthetase blocks intracellular fatty acid flux and glucose uptake in skeletal myotubes. BBA-Mol. Cell Bio. 2020, 158678. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). The State of World Fisheries and Aquaculture 2016; FAO Fisheries and Aquaculture Department: Rome, Italy, 2016. [Google Scholar]
- Yeganeh, S.; Shabanpour, B.; Shabani, A. Comparison of farmed and wild common carp (Cyprinus carpio): Seasonal variations in chemical composition and fatty acid profile. Czech. J. Food Sci. 2012, 30, 503–511. [Google Scholar] [CrossRef]
- Zhu, C.; Zhu, L.; Huang, H.; Zhou, C.; Zhang, Z.; Zheng, Z. Comparative Analysis of Muscle Nutritional Components of Wild and Cultivated Rock Carp (Procypris rabaudi). J. Nutr. (Chinese) 2017, 39, 203. [Google Scholar] [CrossRef]
- Fujino, T.; Yamamoto, T. Cloning and functional expression of a novel long-chain acyl-CoA synthetase expressed in brain. J. Biochem. 1992, 111, 197–203. [Google Scholar] [CrossRef]
- Malhotra, K.T.; Malhotra, K.; Lubin, B.H.; Kuypers, F.A. Identification and molecular characterization of acyl-CoA synthetase in human erythrocytes and erythroid precursors. Biochem. J. 1999, 344, 135–143. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, H.C.; Cho, Y.Y.; Byun, S.J.; Lim, J.M.; Ryoo, Z.Y. Alternative promotion of the mouse acyl-CoA synthetase 6 (mAcsl6) gene mediates the expression of multiple transcripts with 5′-end heterogeneity: Genetic organization of mAcsl6 variants. Biochem. Biophys. Res. Commun. 2005, 327, 84–93. [Google Scholar] [CrossRef]
- Weedon-Fekjaer, M.S.; Dalen, K.T.; Solaas, K.; Staff, A.C.; Duttaroy, A.K.; Nebb, H.I. Activation of LXR increases acyl-CoA synthetase activity through direct regulation of ACSL3 in human placental trophoblast cells. J. Lipid Res. 2010, 51, 1886–1896. [Google Scholar] [CrossRef]
- Cao, A.; Li, H.; Zhou, Y.; Wu, M.; Liu, J. Long chain acyl-CoA synthetase-3 is a molecular target for peroxisome proliferator-activated receptor δ in HepG2 hepatoma cells. J. Biol. Chem. 2010, 285, 16664–16674. [Google Scholar] [CrossRef] [PubMed]
- Kan, C.F.; Singh, A.B.; Dong, B.; Shende, V.R.; Liu, J. PPARδ activation induces hepatic long-chain acyl-CoA synthetase 4 expression in vivo and in vitro. BBA-Mol. Cell Biol. 2015, 1851, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.B.; Kan, C.F.; Dong, B.; Liu, J. SREBP2 activation induces hepatic long-chain acyl-CoA synthetase 1 (ACSL1) expression in vivo and in vitro through a sterol regulatory element (SRE) motif of the ACSL1 C-promoter. J. Biol. Chem. 2016, 291, 5373–5384. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zan, L.; Li, A.; Cheng, G.; Li, S.; Zhang, Y.; Zhang, Y. Characterization of the promoter region of the bovine long-chain acyl-CoA synthetase 1 gene: Roles of E2F1, Sp1, KLF15, and E2F4. Sci. Rep. 2016, 6, 19661. [Google Scholar] [CrossRef]
- Soupene, E.; Kuypers, F.A. Multiple erythroid isoforms of human long-chain acyl-CoA synthetases are produced by switch of the fatty acid gate domains. BMC Mol. Biol. 2006, 7, 21. [Google Scholar] [CrossRef]
- Soupene, E.; Dinh, N.P.; Siliakus, M.; Kuypers, F.A. Activity of the acyl-CoA synthetase ACSL6 isoforms: Role of the fatty acid Gate-domains. BMC biochem. 2010, 11, 18. [Google Scholar] [CrossRef]
- Hisanaga, Y.; Ago, H.; Nakagawa, N.; Hamada, K.; Ida, K.; Yamamoto, M.; Hori, T.; Arii, Y.; Sugahara, M.; Kuramitsu, S.; et al. Structural basis of the substrate-specific two-step catalysis of long chain fatty acyl-CoA synthetase dimer. J. Biol. Chem. 2004, 279, 31717–31726. [Google Scholar] [CrossRef]
- O’Malley, B.W.; Towle, H.C.; Schwartz, R.J. Regulation of gene expression in eucaryotes. Annu. Rev. Genet. 1997, 11, 239–275. [Google Scholar] [CrossRef]
- Martin, G.; Schoonjans, K.; Lefebvre, A.M.; Staels, B.; Auwerx, J. Coordinate regulation of the expression of the fatty acid transport protein and acyl-CoA synthetase genes by PPARα and PPARγ activators. J. Biol. Chem. 1997, 272, 28210–28217. [Google Scholar] [CrossRef]
- Horton, J.D.; Shah, N.A.; Warrington, J.A.; Anderson, N.N.; Park, S.W.; Brown, M.S.; Goldstein, J.L. Combined analysis of oligonucleotide microarray data from transgenic and knockout mice identifies direct SREBP target genes. Proc. Natl. Acad. Sci. USA 2003, 100, 12027–12032. [Google Scholar] [CrossRef]
- Motallebipour, M.; Enroth, S.; Punga, T.; Ameur, A.; Koch, C.; Dunham, I.; Wadelius, C. Novel genes in cell cycle control and lipid metabolism with dynamically regulated binding sites for sterol regulatory element-binding protein 1 and RNA polymerase II in HepG2 cells detected by chromatin immunoprecipitation with microarray detection. FEBS J. 2009, 276, 1878–1890. [Google Scholar] [CrossRef]
- Way, J.M.; Harrington, W.W.; Brown, K.K.; Gottschalk, W.K.; Sundseth, S.S.; Mansfield, T.A.; Kliewer, S.A. Comprehensive messenger ribonucleic acid profiling reveals that peroxisome proliferator-activated receptor γ activation has coordinate effects on gene expression in multiple insulin-sensitive tissues. Endocrinology 2001, 142, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Gerhold, D.L.; Liu, F.; Jiang, G.; Li, Z.; Xu, J.; Lu, M.; Doebber, T.W. Gene expression profile of adipocyte differentiation and its regulation by peroxisome proliferator-activated receptor-γ agonists. Endocrinology 2002, 143, 2106–2118. [Google Scholar] [CrossRef] [PubMed]
- Achouri, Y.; Hegarty, B.D.; Allanic, D.; Becard, D.; Hainault, I.; Ferre, P. Long chain fatty acyl-CoA synthetase 5 expression is induced by insulin and glucose: Involvement of sterol regulatory element-binding protein-1c. Biochimie 2005, 87, 1149–1155. [Google Scholar] [CrossRef]
- Xie, D.; Yu, R.; Chen, F.; Lu, R.; Yang, L.; Meng, X.; Nie, G. Effects of dietary LNA/LA ratio on growth, hepatic fatty acid profiles and △6fad, elovl5 mRNA expression in common carp (Cyprinus carpio). J. Fish (Chinese) 2017, 41, 757–765. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. Then eighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Erijman, A.; Dantes, A.; Bernheim, R.; Shifman, J.M.; Peleg, Y. Transfer-PCR (TPCR): A highway for DNA cloning and protein engineering. J. Struct Biol. 2011, 175, 171–177. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, S.; Chen, J.; Zhang, Q.; Liu, Y.; You, C.; Monroig, Ó.; Tocher, D.R.; Li, Y. Hepatocyte nuclear factor 4α (HNF4α) is a transcription factor of vertebrate fatty acyl desaturase gene as identified in marine teleost Siganus canaliculatus. PLoS ONE 2016, 11, e0160361. [Google Scholar] [CrossRef]
- Li, Y.; Monroig, Ó.; Zhang, L.; Wang, S.; Zheng, X.; Dick, J.R.; You, C.; Tocher, D.R. Vertebrate fatty acyl desaturase with Δ4 activity. P. Natl. Acad. Sci. USA. 2010, 107(39), 16840–16845. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Fatty Acids | Injected Treatments | |||
|---|---|---|---|---|
| DMSO | Fenofibrate | Balaglitazone | Betulin | |
| 16:0 | 13.10 ± 0.68 | 11.92 ± 1.78 | 12.22 ± 2.32 | 14.73 ± 1.51 |
| 18:0 | 1.54 ± 0.05 | 1.11 ± 0.07 * | 1.05 ± 0.05 * | 1.93 ± 0.07 * |
| 20:0 | 0.28 ± 0.03 | 0.21 ± 0.02 | 0.25 ± 0.02 | 0.23 ± 0.01 |
| 16:1 | 1.99 ± 0.18 | 2.10 ± 0.37 | 2.13 ± 0.17 | 2.31 ± 0.29 |
| 18:1 | 25.13 ± 1.44 | 21.81 ± 3.11 | 23.25 ± 3.15 | 27.49 ± 4.06 |
| 18:2n-6 | 32.92 ± 3.54 | 33.92 ± 3.51 | 31.97 ± 2.13 | 34.11 ± 2.34 |
| 18:3n-6 | 0.23 ± 0.05 | 0.28 ± 0.10 | 0.27 ± 0.04 | 0.13 ± 0.01 * |
| 20:4n-6 (ARA) | 1.54 ± 0.03 | 1.55 ± 0.22 | 1.63 ± 0.06 | 1.38 ± 0.10 * |
| 18:3n-3 | 5.21 ± 0.02 | 4.93 ± 0.02 | 4.85 ± 0.01 | 5.16 ± 0.02 |
| 18:4n-3 | 0.43 ± 0.10 | 0.40 ± 0.04 | 0.44 ± 0.03 | 0.35 ± 0.01 |
| 20:5n-3 (EPA) | 0.43 ± 0.03 | 0.41 ± 0.02 | 0.48 ± 0.03 | 0.39 ± 0.01 |
| 22:6n-3 (DHA) | 2.27 ± 0.10 | 2.28 ± 0.13 | 2.53 ± 0.07 * | 1.81 ± 0.11 * |
| Subject | Primer Name | Primer Sequence (5′–3′) |
|---|---|---|
| First fragment cloning | acsl6-F1 | ATGCAGTTTCAGGATTGGTTACGT |
| acsl6-R1 | CTATACGGATGATTTCCTGTACATC | |
| 3′RACE | acsl6-F2 | CAGGTGAAGGACTTATATTTGCAC |
| acsl6-F3 | GATGTACAGGAAATCATCCGTATAG | |
| 5′RACE | acsl6-R2 | GCGGAGCGTCTACAGCTCTGATC |
| acsl6-R3 | ACGTAACCAATCCTGAAACTGCAT | |
| PCR for 5′ flanking sequence cloning | SD1-F | CTTACGCGTGCTAGCCCGGGCTCGAGGCCCAGAATTTGCACAGATTACGTG |
| SD2-F | GCTCTTACGCGTGCTAGCCCGGGCTCGAGTGTGAAACATTATGTAAGTTGAATACTTTTAC | |
| SD3-F | GCTCTTACGCGTGCTAGCCCGGGCTCGAGTGATGAAGACGTATTCTTAATTTGCTGG | |
| SD4-F | GCTCTTACGCGTGCTAGCCCGGGCTCGAGACTTATAGTTTGAATCAGATTTTATTTTCCAT | |
| SD-R | ACCAACAGTACCGGAATGCCAAGCTTCTTGCCATCTCCTCCAGTGAAAC | |
| EMSA probes | EMF1 (5′-biotin labeled) | CATTGGCTGATACATCATCATCAT |
| EMR1 (5′-biotin labeled) | CATTTCACCTTTAACCACAGACA | |
| EMF2 | ATTGGCTGATACATCATCATCAT | |
| EMR2 | ATTTCACCTTTAACCACAGACA | |
| RT-PCR and qPCR | RT-acsl6-F | ATGCAGTTTCAGGATTGGTTACGT |
| RT-acsl6-R | TGCTATCTGCAGTCCTCTCTGGAA | |
| Q-acsl6-F | GATCAGAGCTGTAGACGCTCCGCT | |
| Q-acsl6-R | CCTCAGTGTAAGAGATCCACTGAT | |
| pparγ-F | AGACATGGTGGACACGCAGACGTT | |
| pparγ-R | CGCTCTCCTGCATCCTGTAGTTCT | |
| pparα-F | GGGAAAGAGCAGCACGAG | |
| pparα-R | GCGTGCTTTGGCTTTGTT | |
| srebp1-F | CGTCTGCTTCACTTCACTACTC | |
| srebp1-R | GGACCAGTCTTCATCCACAAA | |
| 18S rRNA | CTGAGAAACGGCTACCATTC | |
| 18S rRNA | GCCTCGAAAGAGACCTGTATTG |
| TFs | Position | Predicted Element | Mutation Site |
|---|---|---|---|
| FOXO1 | −733 | TGGTGTTTTTC | TGGTGTTTTTC→× |
| SP1 | −672 | CATTGGCTG | CATTGGCTG→× |
| AP1 | −658 | TCATCATCATC | TCATCATCATC→× |
| CDX1 | −545 | ATTTATA | ATTTATA→× |
| YY1 | −485 | GCCATTTA | GCCATTTA→× |
| −453 | AAAAATGGCACA | AAAAATGGCACA→× | |
| C/EBPα | −474 | CTTTGCAAT | CTTTGCAAT→× |
| TBP | −404 | CTATTAAAAGGGCCA | CTATTAAAAGGGCCA→× |
| SREBP-1 | −343 | GTGTTGTGATG | GTGTTGTGATG→× |
| PPARγ:RXRα | −325 | TCTGTGGTTAAAGGTGAAAT | TCTGTGGTTAAAGGTGAAAT→× |
| PPARα:RXRα | −323 | TGTGGTTAAAGGTGAA | TGTGGTTAAAGGTGAA→× |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, D.; He, Z.; Dong, Y.; Gong, Z.; Nie, G.; Li, Y. Molecular Cloning, Characterization, and Expression Regulation of Acyl-CoA Synthetase 6 Gene and Promoter in Common Carp Cyprinus carpio. Int. J. Mol. Sci. 2020, 21, 4736. https://doi.org/10.3390/ijms21134736
Xie D, He Z, Dong Y, Gong Z, Nie G, Li Y. Molecular Cloning, Characterization, and Expression Regulation of Acyl-CoA Synthetase 6 Gene and Promoter in Common Carp Cyprinus carpio. International Journal of Molecular Sciences. 2020; 21(13):4736. https://doi.org/10.3390/ijms21134736
Chicago/Turabian StyleXie, Dizhi, Zijie He, Yewei Dong, Zhiyuan Gong, Guoxing Nie, and Yuanyou Li. 2020. "Molecular Cloning, Characterization, and Expression Regulation of Acyl-CoA Synthetase 6 Gene and Promoter in Common Carp Cyprinus carpio" International Journal of Molecular Sciences 21, no. 13: 4736. https://doi.org/10.3390/ijms21134736
APA StyleXie, D., He, Z., Dong, Y., Gong, Z., Nie, G., & Li, Y. (2020). Molecular Cloning, Characterization, and Expression Regulation of Acyl-CoA Synthetase 6 Gene and Promoter in Common Carp Cyprinus carpio. International Journal of Molecular Sciences, 21(13), 4736. https://doi.org/10.3390/ijms21134736