Distinct Local and Systemic Molecular Signatures in the Esophageal and Gastric Cancers: Possible Therapy Targets and Biomarkers for Gastric Cancer

 , , , and
, , , and
Abstract
1. Introduction
2. Results
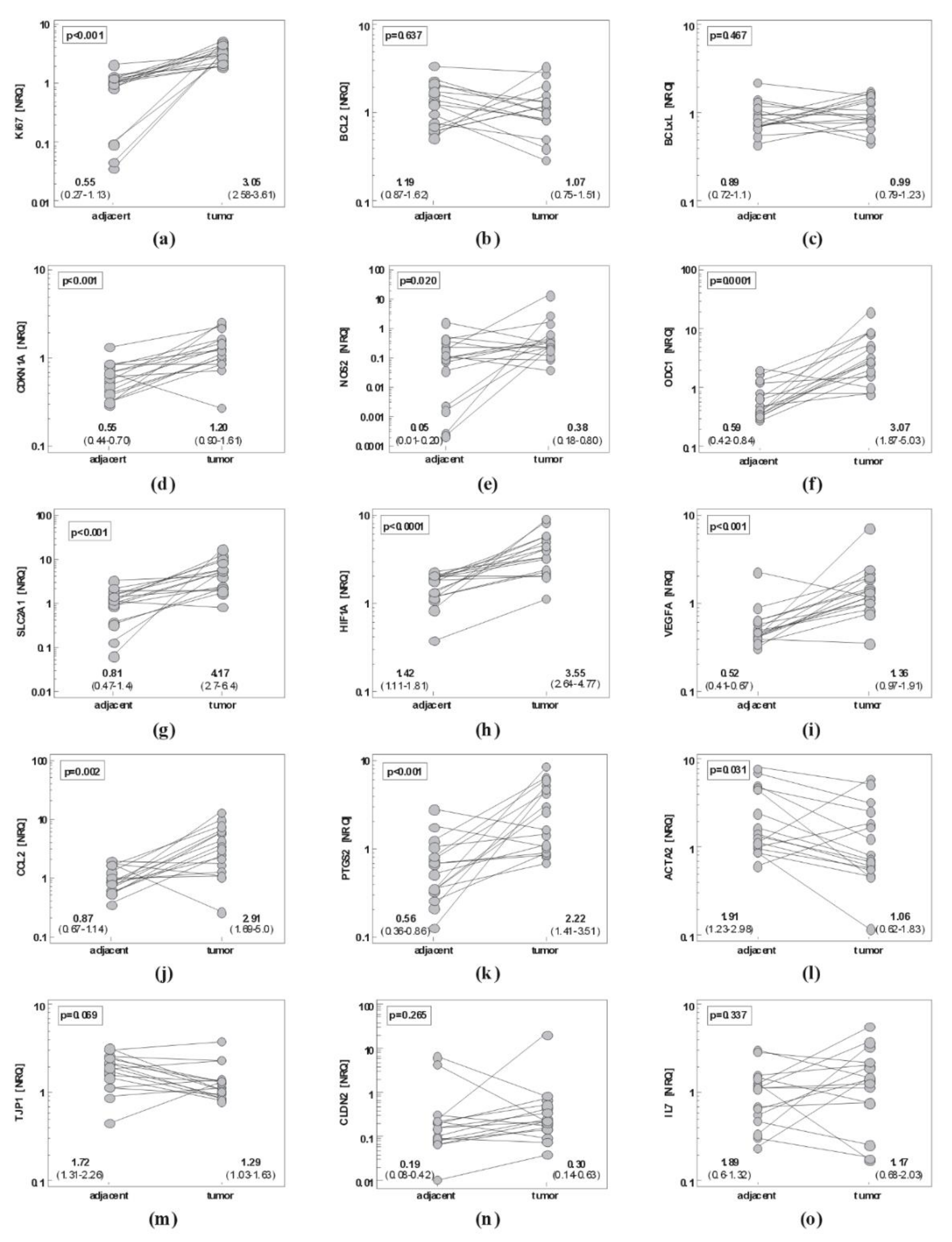
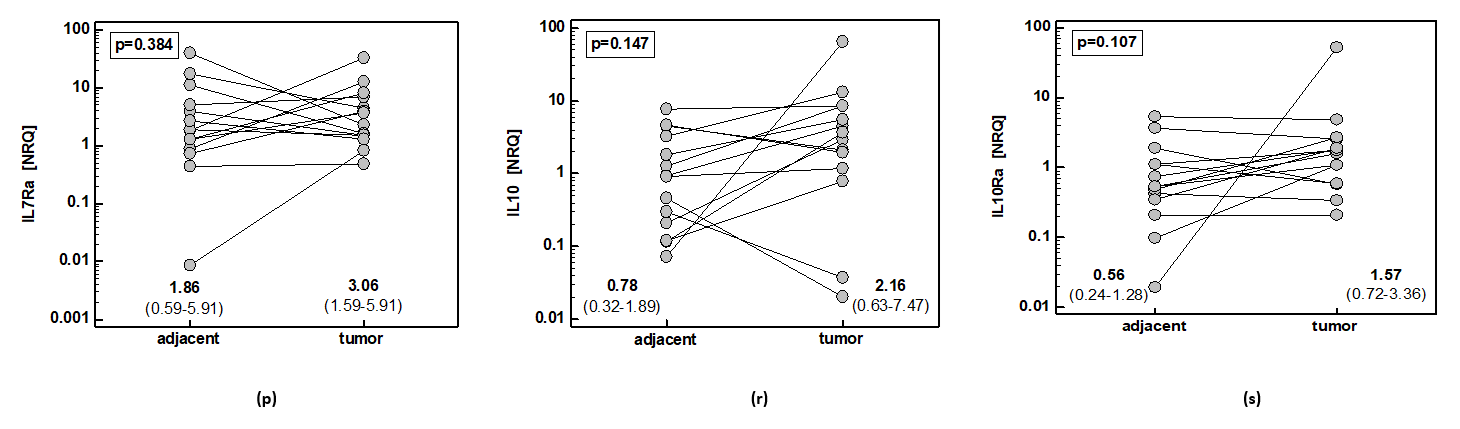
2.1. Local Expression of Cancer-Promoting Mediators
2.1.1. Gastric Cancer
2.1.2. Esophageal Cancer
2.1.3. Comparison of Gene Signatures in Gastric and Esophageal Cancers
Tumor-to-Adjacent Fold-Change in Expression
Gene Expression in Tumors and Non-Cancerous Tissue
Independent Predictors of Gene Expression in Tumors and Non-Cancerous Gastric Tissue
Impact of GC Pathological Stage on Fold Change in Gene Expression
2.2. Systemic Cytokine Signatures in EC and GC
2.2.1. Systemic Concentration of Cytokines, Chemokines, and Growth Factors in Gastric and Esophageal Cancer and Benign Conditions
Cytokine Signatures Distinguishing between GC and Benign Conditions
Cytokine Signatures Distinguishing between GC and EC
2.2.2. Cytokine Signatures Distinguishing between Cardia and Non-Cardia Subtypes of Gastric Cancer
3. Discussion
4. Materials and Methods
4.1. Patients
4.1.1. Study Population—Local Molecular Signatures (Gene Expression)
4.1.2. Study Population—Systemic Cytokine Signatures
4.1.3. Ethical Considerations
4.2. Analytical Methods
4.2.1. Sample Collection
Tissue Samples
Serum Samples
4.2.2. Transcriptional Analysis
Tissue Homogenization
RNA Isolation
cDNA Synthesis
Quantitative (Real-Time) PCR
Expression Calculation and Normalization Strategy
4.2.3. Serum Cytokine Quantification
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AUC | Area under ROC curve |
| EC | Esophageal cancer |
| EMT | Epithelial-mesenchymal transition |
| GC | Gastric cancer |
| GERD | Gastro-esophageal reflux disease |
| IL | Interleukin |
| NRQ | Normalized relative quantity |
| ROC | Receiver operating characteristics curve analysis |
| TNM | Tumor-node-metastasis cancer staging system |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Thrift, A.P.; El-Serag, H.B. Burden of Gastric Cancer. Clin. Gastroenterol. Hepatol. 2020, 18, 534–542. [Google Scholar] [CrossRef] [PubMed]
- GBD 2017 Stomach Cancer Collaborators. The Global, Regional, and National Burden of Stomach Cancer in 195 Countries, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 42–54. [Google Scholar] [CrossRef]
- Rawla, P.; Barsouk, A. Epidemiology of Gastric Cancer: Global Trends, Risk Factors and Prevention. Prz Gastroenterol. 2019, 14, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Meltzer, S.J. Gastric Cancer in the Era of Precision Medicine. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.Y.; Noh, S.H.; Cheong, J.H. Evolution of Gastric Cancer Treatment: From the Golden Age of Surgery to an Era of Precision Medicine. Yonsei Med. J. 2015, 56, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Tustumi, F.; Kimura, C.M.; Takeda, F.R.; Uema, R.H.; Salum, R.A.; Ribeiro-Junior, U.; Cecconello, I. Prognostic Factors and Survival Analysis in Esophageal Carcinoma. Arquivos Brasileiros de Cirurgia Digestiva (São Paulo) 2016, 29, 138–141. [Google Scholar] [CrossRef]
- Aftimos, P.G.; Barthelemy, P.; Awada, A. Molecular Biology in Medical Oncology: Diagnosis, Prognosis, and Precision Medicine. Discov. Med. 2014, 17, 81–91. [Google Scholar]
- Patel, A.; Tripathi, G.; Gopalakrishnan, K.; Williams, N.; Arasaradnam, R.P. Field Cancerisation in Colorectal Cancer: A New Frontier or Pastures Past? World J. Gastroenterol. 2015, 21, 3763–3772. [Google Scholar] [CrossRef]
- Sesti, F.; Tsitsilonis, O.E.; Kotsinas, A.; Trougakos, I.P. Oxidative Stress-Mediated Biomolecular Damage and Inflammation in Tumorigenesis. Vivo 2012, 26, 395–402. [Google Scholar]
- Wang, D.H. The Esophageal Squamous Epithelial Cell-Still a Reasonable Candidate for the Barrett’s Esophagus Cell of Origin? Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Clark, D. Molecular Margin of Surgical Resections—Where Do We Go from Here? Cancer 2015, 121, 1914–1916. [Google Scholar] [CrossRef] [PubMed]
- Dakubo, G.D.; Jakupciak, J.P.; Birch-Machin, M.A.; Parr, R.L. Clinical Implications and Utility of Field Cancerization. Cancer Cell Int. 2007, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, K.; Bednarz-Misa, I.; Diakowska, D.; Kapturkiewicz, B.; Gamian, A.; Krzystek-Korpacka, M. Nampt/PBEF/Visfatin Upregulation in Colorectal Tumors, Mirrored in Normal Tissue and Whole Blood of Colorectal Cancer Patients, Is Associated with Metastasis, Hypoxia, IL1β, and Anemia. BioMed. Res. Int. 2015, 2015, 523930. [Google Scholar] [CrossRef]
- Krzystek-Korpacka, M.; Gorska, S.; Diakowska, D.; Kapturkiewicz, B.; Podkowik, M.; Gamian, A.; Bednarz-Misa, I. Midkine Is Pp-regulated in Both Cancerous and Inflamed Bowel, Reflecting Lymph Node Metastasis in Colorectal Cancer and Clinical Activity of Ulcerative Colitis. Cytokine 2017, 89, 68–75. [Google Scholar] [CrossRef]
- Bednarz-Misa, I.; Diakowska, D.; Szczuka, I.; Fortuna, P.; Kubiak, A.; Rosińczuk, J.; Krzystek-Korpacka, M. Interleukins 4 and 13 and Their Receptors Are Differently Expressed in Gastrointestinal Tract Cancers, Depending on the Anatomical Site and Disease Advancement, and Improve Colon Cancer Cell Viability and Motility. Cancers 2020, 12, 1463. [Google Scholar] [CrossRef]
- Bednarz-Misa, I.; Diakowska, D.; Krzystek-Korpacka, M. Local and Systemic IL-7 Concentration in Gastrointestinal Tract Cancers. Medicina 2019, 55, E262. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Wang, J.; Ye, C.; Chen, C.; Xiong, H.; Xie, B.; Zhou, J.; Chen, Y.; Zheng, S.; Wang, L. Glucose Transporter GLUT1 Expression and Clinical Outcome in Solid Tumors: A Systematic Review and Meta-Analysis. Oncotarget 2017, 8, 16875–16886. [Google Scholar] [CrossRef]
- Usuda, K.; Sagawa, M.; Aikawa, H.; Ueno, M.; Tanaka, M.; Machida, Y.; Zhao, X.T.; Ueda, Y.; Higashi, K.; Sakuma, T. Correlation between glucose transporter-1 expression and 18F-fluoro-2-deoxyglucose uptake on positron emission tomography in lung cancer. Gen. Thorac. Cardiovasc. Surg. 2010, 58, 405–410. [Google Scholar] [CrossRef]
- Carvalho, K.C.; Cunha, I.W.; Rocha, R.M.; Ayala, F.R.; Cajaíba, M.M.; Begnami, M.D.; Vilela, R.S.; Paiva, G.R.; Andrade, R.G.; Soares, F.A. GLUT1 Expression in Malignant Tumors and Its Use as an Immunodiagnostic Marker. Clinics 2011, 66, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Kim, H.; Nam, K.; Shin, I. Glut1 Promotes Cell Proliferation, Migration and Invasion by Regulating Epidermal Growth Factor Receptor and Integrin Signaling in Triple-Negative Breast Cancer Cells. BMB Rep. 2017, 50, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Sawayama, H.; Ogata, Y.; Ishimoto, T.; Mima, K.; Hiyoshi, Y.; Iwatsuki, M.; Baba, Y.; Miyamoto, Y.; Yoshida, N.; Baba, H. Glucose Transporter 1 Regulates the Proliferation and Cisplatin Sensitivity of Esophageal Cancer. Cancer Sci. 2019, 110, 1705–1714. [Google Scholar] [CrossRef]
- Gao, J.H.; Guo, L.J.; Huang, Z.Y.; Rao, J.N.; Tang, C.W. Roles of Cellular Polyamines in Mucosal Healing in the Gastrointestinal Tract. J. Physiol. Pharmacol. 2013, 64, 681–693. [Google Scholar] [PubMed]
- Casero, R.A.; Murray Stewart, T.; Pegg, A.E. Polyamine Metabolism and Cancer: Treatments, Challenges and Opportunities. Nat. Rev. Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Meyskens, F.L., Jr.; McLaren, C.E.; Pelot, D.; Fujikawa-Brooks, S.; Carpenter, P.M.; Hawk, E.; Kelloff, G.; Lawson, M.J.; Kidao, J.; McCracken, J.; et al. Difluoromethylornithine Plus Sulindac for the Prevention of Sporadic Colorectal Adenomas: A Randomized Placebo-Controlled, Double-Blind Trial. Cancer Prev. Res. 2008, 1, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Gerner, E.W.; Bruckheimer, E.; Cohen, A. Cancer Pharmacoprevention: Targeting Polyamine Metabolism to Manage Risk Factors for Colon Cancer. J. Biol. Chem. 2018, 293, 18770–18778. [Google Scholar] [CrossRef]
- Danza, K.; Silvestris, N.; Simone, G.; Signorile, M.; Saragoni, L.; Brunetti, O.; Monti, M.; Mazzotta, A.; De Summa, S.; Mangia, A.; et al. Role of miR-27a, miR-181a and miR-20b in gastric cancer hypoxia-induced chemoresistance. Cancer Biol. Ther. 2016, 17, 400–406. [Google Scholar] [CrossRef]
- Itoh, M.; Nishibori, N.; Her, S.; Lee, M.; Morita, K. Chemical Hypoxia-Induced Stimulation of Polyamine Biosynthesis and Ornithine Decarboxylase Gene Transcription in C6 Glioma Cells. J. Mol. Pathophysiol. 2015, 4, 29–37. [Google Scholar] [CrossRef]
- Huang, L.; Wu, R.L.; Xu, A.M. Epithelial-Mesenchymal Transition in Gastric Cancer. Am. J. Transl. Res. 2015, 7, 2141–2158. [Google Scholar]
- Ohtani, S.; Terashima, M.; Satoh, J.; Soeta, N.; Saze, Z.; Kashimura, S.; Ohsuka, F.; Hoshino, Y.; Kogure, M.; Gotoh, M. Expression of Tight-Junction-Associated Proteins in Human Gastric Cancer: Downregulation of Claudin-4 Correlates with Tumor Aggressiveness and Survival. Gastric Cancer 2009, 12, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Ghassemifar, R.; Lai, C.M.; Rakoczy, P.E. VEGF Differentially Regulates Transcription and Translation of ZO-1alpha+ and ZO-1alpha- and Mediates Trans-Epithelial Resistance in Cultured Endothelial and Epithelial Cells. Cell Tissue Res. 2006, 323, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Abu-Farsakh, S.; Wu, T.; Lalonde, A.; Sun, J.; Zhou, Z. High Expression of Claudin-2 in Esophageal Carcinoma and Precancerous Lesions Is Significantly Associated with the Bile Salt Receptors VDR and TGR5. BMC Gastroenterol. 2017, 17, 33. [Google Scholar] [CrossRef]
- Jung, H.; Jun, K.H.; Jung, J.H.; Chin, H.M.; Park, W.B. The Expression of Claudin-1, Claudin-2, Claudin-3, and Claudin-4 in Gastric Cancer Tissue. J. Surg. Res. 2011, 167, e185–e191. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sun, X.; Meng, X. Differences in the Expression Profiles of Claudin Proteins in Human Gastric Carcinoma Compared with Non-Neoplastic Mucosa. Mol. Med. Rep. 2018, 18, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, S.; Anwer, S.; Szászi, K. Claudin-2: Roles beyond Permeability Functions. Int. J. Mol. Sci. 2019, 20, 5655. [Google Scholar] [CrossRef]
- Dhawan, P.; Ahmad, R.; Chaturvedi, R.; Smith, J.J.; Midha, R.; Mittal, M.K.; Krishnan, M.; Chen, X.; Eschrich, S.; Yeatman, T.J.; et al. Claudin-2 Expression Increases Tumorigenicity of Colon Cancer Cells: Role of Epidermal Growth Factor Receptor Activation. Oncogene 2011, 30, 3234–3247. [Google Scholar] [CrossRef]
- Paquet-Fifield, S.; Koh, S.L.; Cheng, L.; Beyit, L.M.; Shembrey, C.; Mølck, C.; Behrenbruch, C.; Papin, M.; Gironella, M.; Guelfi, S.; et al. Tight Junction Protein Claudin-2 Promotes Self-Renewal of Human Colorectal Cancer Stem-like Cells. Cancer Res. 2018, 78, 2925–2938. [Google Scholar] [CrossRef]
- Mima, S.; Takehara, M.; Takada, H.; Nishimura, T.; Hoshino, T.; Mizushima, T. NSAIDs Suppress the Expression of Claudin-2 to Promote Invasion Activity of Cancer Cells. Carcinogenesis 2008, 29, 1994–2000. [Google Scholar] [CrossRef]
- Asplund, J.; Kauppila, J.H.; Mattsson, F.; Lagergren, J. Survival Trends in Gastric Adenocarcinoma: A Population-Based Study in Sweden. Ann. Surg. Oncol. 2018, 25, 2693–2702. [Google Scholar] [CrossRef]
- Matusiewicz, M.; Neubauer, K.; Bednarz-Misa, I.; Gorska, S.; Krzystek-Korpacka, M. Systemic interleukin-9 in inflammatory bowel disease: Association with mucosal healing in ulcerative colitis. World J. Gastroenterol. 2017, 23, 4039–4046. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, K.; Weigmann, B. The Dichotomous Function of Interleukin-9 in Cancer Diseases. J. Mol. Med. 2019, 97, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Krzystek-Korpacka, M.; Diakowska, D.; Kapturkiewicz, B.; Bębenek, M.; Gamian, A. Profiles of Circulating Inflammatory Cytokines in Colorectal Cancer (CRC), High Cancer Risk Conditions, and Health Are Distinct. Possible Implications for CRC Screening and Surveillance. Cancer Lett. 2013, 337, 107–114. [Google Scholar] [CrossRef] [PubMed]
- El-Omar, E.M. The Importance of Interleukin 1β in Helicobacter pylori Associated Disease. Gut 2001, 48, 743–747. [Google Scholar] [CrossRef]
- Rozengurt, E.; Walsh, J.H. Gastrin, CCK, Signaling, and Cancer. Annu. Rev. Physiol. 2001, 63, 49–76. [Google Scholar] [CrossRef]
- Camargo, C.M.; Mera, R.; Correa, P.; Peek, R.M., Jr.; Fontham, E.T.H.; Goodman, K.J.; Piazuelo, B.M.; Sicinschi, L.; Zabaleta, J.; Schneider, B.G. Interleukin-1β and Interleukin-1 Receptor Antagonist Gene Polymorphisms and Gastric Cancer: A Meta-analysis. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1674–1687. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; Gabrilovich, D.; Sotomayor, E.M. Immunosuppressive Strategies that are Mediated by Tumor Cells. Annu. Rev. Immunol. 2007, 25, 267–296. [Google Scholar] [CrossRef]
- Hallett, M.A.; Venmar, K.T.; Fingleton, B. Cytokine Stimulation of Epithelial Cancer Cells: The Similar and Divergent Functions of IL4 and IL13. Cancer Res. 2012, 72, 6338–6343. [Google Scholar] [CrossRef]
- Bankaitis, K.V.; Fingleton, B. Targeting IL4/IL4R for the Treatment of Epithelial Cancer Metastasis. Clin. Exp. Metastasis 2015, 32, 847–856. [Google Scholar] [CrossRef]
- Hellemans, J.; Vandesompele, J. qPCR Data analysis—Unlocking the Secret to Successful Results. In PCR Troubleshooting and Optimization: The Essential Guide, 1st ed.; Kennedy, S., Oswald, N., Eds.; Caister Academic Press: Poole, UK, 2011; pp. 1–13. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics: | EC | GC | p |
|---|---|---|---|
| n | 16 | 15 | − |
| Sex (F/M), n | 6/10 | 4/11 | 0.704 1 |
| Age (yrs.), mean ± SD | 63.3 ± 7 | 66.0 ± 12 | 0.465 2 |
| Stage (I/II/III/IV) | 0/5/9/2 | 2/3/7/3 | 0.404 3 |
| Primary tumor, T (1/2/3/4) | 0/5/8/3 | 1/1/9/4 | 0.279 3 |
| Lymph node metastasis, N (no/yes) | 8/8 | 5/10 | 0.472 1 |
| Distant metastasis, M (no/yes) | 14/2 | 12/3 | 0.654 1 |
| Histological grade, G (1/2/3/x) | 5/7/4/0 | 1/6/7/1 | 0.209 3 |
| Gene | Fold-Change (Tumor-to-Adjacent) | p | |
|---|---|---|---|
| GC | EC | ||
| Ki67 | 1.97 | 5.52 | 0.032 1 |
| BCL2 | 0.35 | 0.90 | 0.078 2 |
| BCLxL | 0.88 | 1.10 | 0.267 1 |
| CDKN1A | 0.62 | 2.18 | <0.001 1 |
| NOS2 | 3.29 | 8.12 | 0.527 1 |
| ODC1 | 0.68 | 5.17 | <0.0001 1 |
| SLC2A1 | 1.66 | 5.15 | 0.017 1 |
| HIF1A | 0.94 | 2.51 | 0.001 1 |
| VEGFA | 0.98 | 2.62 | 0.025 2 |
| CCL2 | 0.52 | 3.34 | <0.0011 |
| PTGS2 | 1.21 | 4.01 | 0.014 1 |
| ACTA2 | 0.71 | 0.56 | 0.564 1 |
| TJP1 | 0.62 | 0.75 | 0.496 1 |
| CLDN2 | 3.98 | 1.61 | 0.128 1 |
| IL7 | 0.98 | 1.32 | 0.642 2 |
| IL7Ra | 1.65 | 1.95 | 0.805 1 |
| IL10 | 2.76 | 3.9 | 0.616 2 |
| IL10Ra | 2.8 | 1.74 | 0.467 2 |
| Gene | Non-Cancerous Tissue (NRQ) | Tumor (NRQ) | ||||
|---|---|---|---|---|---|---|
| GC | EC | p | GC | EC | p | |
| Ki67 | 2.19 | 0.55 | 0.027 1 | 4.32 | 3.05 | 0.347 2 |
| BCL2 | 4.87 | 1.19 | 0.0001 2 | 1.68 | 1.07 | 0.372 2 |
| BCLxL | 3.08 | 0.89 | <0.001 2 | 2.72 | 0.99 | 0.002 2 |
| CDKN1A | 3.41 | 0.55 | <0.0001 2 | 2.1 | 1.2 | 0.038 1 |
| NOS2 | 0.37 | 0.05 | 0.073 1 | 1.23 | 0.38 | 0.100 1 |
| ODC1 | 5.8 | 0.59 | <0.0001 1 | 3.91 | 3.07 | 0.518 1 |
| SLC2A1 | 0.9 | 0.81 | 0.778 1 | 1.5 | 4.17 | 0.011 1 |
| HIF1A | 4.54 | 1.42 | 0.001 2 | 4.27 | 3.55 | 0.609 2 |
| VEGFA | 4.6 | 0.52 | <0.001 2 | 4.52 | 1.36 | 0.004 2 |
| CCL2 | 9.39 | 0.87 | <0.001 2 | 4.93 | 2.91 | 0.243 1 |
| PTGS2 | 2.93 | 0.56 | <0.001 1 | 3.55 | 2.22 | 0.240 1 |
| ACTA2 | 4.57 | 1.91 | 0.008 1 | 3.23 | 1.06 | 0.008 1 |
| TJP1 | 4.44 | 1.72 | <0.001 1 | 2.74 | 1.29 | 0.019 2 |
| CLDN2 | 3.36 | 0.19 | <0.001 1 | 13.4 | 0.3 | <0.001 1 |
| IL7 | 1.93 | 0.89 | 0.095 | 1.9 | 1.17 | 0.345 1 |
| IL7Ra | 1.86 | 0.53 | 0.047 2 | 3.06 | 1.04 | 0.037 1 |
| IL10 | 0.78 | 0.70 | 0.805 2 | 2.16 | 2.73 | 0.695 2 |
| IL10Ra | 0.56 | 0.56 | 0.993 2 | 1.57 | 0.97 | 0.245 2 |
| Explained Variable | Explanatory Variables | ||
|---|---|---|---|
| Entered | Retained | R2 | |
| CLDN2 | BCLxL, SLC2A1, Ki67, NOS2 | Ki67: rp = 0.81, NOS2: rp = 0.80 | 0.827 |
| ACTA2 | CCL2, PTGS2, IL-7, CDKN1A, TJP1 | PTGS2: rp = 0.62 | 0.381 |
| TJP1 | ACTA2, HIF1A, VEGFA, BCLxL, SLC2A1, ODC1, CDKN1A, BCL2, Ki67 | ACTA2: rp = 0.68, HIF1A: rp = 0.67, VEGFA: rp = 0.71 | 0.896 |
| ODC1 | SLC2A1, HIF1A, BCLxL, CDKN1A, BCL2, Ki67, VEGFA, TJP1 | SLC2A1: rp = 0.64, HIF1A: rp = 0.88 | 0.938 |
| SLC2A1 | BCLxL, HIF1A, CDKN1A, ODC1, BCL2, CLDN2, Ki67, VEGFA, TJP1 | ODC1: rp = 0.85 | 0.715 |
| VEGFA | BCLxL, BCL2, CCL2, HIF1A, CDKN1A, ODC1, Ki67, TJP1 | TJP1: rp = 0.84 | 0.712 |
| HIF1A | SLC2A1, BCL2, Ki67, VEGFA, TJP1, BCLxL, CCL2, ODC1, CDKN1A | BCLxL: rp = 0.80, CCL2: rp = 0.90, ODC1: rp = 0.95, CDKN1A: rp = −0.81 | 0.987 |
| PTGS2 | ACTA2, CDKN1A, BCL2, CCL2 | CCL2: rp = 0.77 | 0.585 |
| CCL2 | ACTA2, HIF1A, CDKN1A, Ki67, PTGS2, BCL2, IL7 | PTGS2: rp = 0.53, BCL2: rp = 0.69, IL7: rp = −0.78 | 0.876 |
| Ki67 | BCLxL, CLDN2, SLC2A1, HIF1A, ODC1, BCL2, VEGFA, TJP1 | BCLxL: rp = 0.70 CLDN2: rp = 0.58 | 0.753 |
| ‘BCLxL | SLC2A1, HIF1A, ODC1, CDKN1A, BCL2, CLDN2, Ki67, VEGFA, TJP1 | HIF1A: rp = 0.90 | 0.808 |
| BCL2 | HIF1A, PTGS2, BCLxL, CCL2, SLC2A1, ODC1, CDKN1A, CLDN2, VEGFA, TPJ1, Ki67 | HIF1A: rp = 0.75, PTGS2: rp = 0.60 | 0.772 |
| CDKN1A | ACTA2, BCLxL, SLC2A1, BCL2, TJP1, PTGS2, CCL2, HIF1A, ODC1, VEGFA | CCL2: rp = 0.89, HIF1A: rp = −0.73, ODC1: rp = 0.59, VEGFA: rp = 0.66 | 0.871 |
| Explained Variable | Explanatory Variables | ||
|---|---|---|---|
| Entered | Retained | R2 | |
| CLDN2 | Ki67, BCLxL | BCLxL: rp = 0.80 | 0.642 |
| ACTA2 | BCL2, CCL2, NOS2, CDKN1A, TJP1 | BCL2: rp = 0.63 | 0.391 |
| TJP1 | CCL2, IL7, NOS2, ODC1, ACTA2, BCL2, HIF1A, CDKN1A, PTGS2 | BCL2: rp = 0.84, HIF1A: rp = 0.71, CDKN1A: rp = 0.91, PTGS2: rp = −0.82 | 0.978 |
| ODC1 | HIF1A, BCLxL, CCL2, SLC2A1, Ki67, CDKN1A, PTGS2, VEGFA, TJP1 | HIF1A: rp = 0.86 | 0.736 |
| SLC2A1 | Ki67, BCLxL, HIF1A, VEGFA, ODC1 | Ki67: rp = 0.80 | 0.632 |
| VEGFA | BCLxL, ODC1, SLC2A1, HIF1A, Ki67, CDKN1A | SLC2A1: rp = 0.77 | 0.589 |
| HIF1A | CCL2, ODC1, BCL2, BCLxL, Ki67, PTGS2, VEGFA, TJP1, SLC2A1, IL7, CDKN1A | CCL2: rp = 0.78, ODC1: rp = 0.76 | 0.896 |
| PTGS2 | BCLxL, CCL2, VEGFA, HIF1A, Ki67, CDKN1A, ODC1, TJP1 | BCLxL: rp = −0.60, CCL2: rp = 0.85, VEGFA: rp = 0.87 | 0.907 |
| CCL2 | HIF1A, BCL2, IL7, ODC1, PTGS2, CDKN1A, ACTA2, TJP1 | HIF1A: rp = 0.74, BCL2: rp = 0.56 | 0.829 |
| Ki67 | BCLxL, ODC1, CLDN2, HIF1A, PTGS2, CDKN1A, VEGFA, SLC2A1 | BCLxL: rp = 0.91, ODC1: rp = 0.82 | 0.940 |
| BCLxL | CLDN2, Ki67, SLC2A1, HIF1A, ODC1, VEGFA | CLDN2: rp = 0.72, Ki67: rp = 0.87 | 0.913 |
| BCL2 | TJP1, CCL2, CLDN2, HIF1A, IL7, CDKN1A, ACTA2 | TJP1: rp = 0.88 | 0.779 |
| CDKN1A | PTGS2, TJP1, BCL2, CCL2, HIF1A, Ki67, NOS2, ODC1, VEGFA, ACTA2 | PTGS2: rp = 0.76, TJP1: rp = 0.89 | 0.915 |
| Gene | TNM 1 | T 1 | N (N0 vs. N1/2) 2 | M 2 | G 1 |
|---|---|---|---|---|---|
| Ki67 | 0.63 3 | 0.50 5 | 0.77 vs. 3.16 3 | ns | ns |
| BCL2 | ns | 0.50 5 | 0.11 vs. 0.62 5 | ns | ns |
| BCLxL | ns | ns | 0.62 vs. 1.05 5 | ns | ns |
| CDKN1A | 0.51 3 | 0.55 3 | ns | 0.50 vs. 1.44 5 | 0.58 3 |
| ODC1 | 0.58 3 | 0.46 5 | 0.32 vs. 0.99 4 | ns | ns |
| HIF1A | 0.56 3 | ns | 0.43 vs. 1.39 4 | ns | ns |
| CCL2 | ns | 0.53 3 | 0.22 vs. 0.8 5 | ns | 0.54 3 |
| PTGS2 | ns | ns | ns | ns | 0.48 5 |
| TJP1 | 0.66 4 | 0.69 4 | 0.3 vs. 0.88 3 | 0.49 vs. 1.59 5 | ns |
| IL10 | ns | ns | ns | ns | 0.56 3 |
| Characteristics: | Benign | EC | GC | p | |
|---|---|---|---|---|---|
| Cardia | Non-Cardia | ||||
| n | 39 | 92 | 32 | 32 | - |
| Sex (F/M), n | 18/21 | 31/61 | 7/25 | 10/21 | 0.195 1 |
| Age (yrs.), mean ± SD | 61.1 ± 13 | 62.4 ± 9 | 62.3 ± 9 | 63.3 ± 11 | 0.820 2 |
| Stage (I/II/III/IV) | 6/23/25/38 | 0/5/6/21 | 1/9/6/16 | 0.260 1 | |
| Primary tumor, T (1/2/3/4) | 8/15/27/42 | 0/1/8/23 | 1/2/12/17 | 0.059 1 | |
| Lymph node metastasis, N (no/yes) | 33/59 | 5/27 | 10/22 | 0.102 1 | |
| Distant metastasis, M (no/yes) | 54/38 | 11/21 | 16/16 | 0.058 1 | |
| Cytokine (pg/mL) | Benign Conditions | Esophageal Cancer | Gastric Cancer | p Value |
|---|---|---|---|---|
| IL-1β | 0.15 (0.15–0.15) 2,3 | 0.15 (0.15–1.66) 1,3 | 1.12 (0.15–1.55) 1,2 | <0.0001 |
| IL-1ra | 339.3 (116–630) 2,3 | 119.4 (71–518) 1,3 | 67.8 (42–167) 1,2 | <0.0001 |
| IL-4 | 3.52 (2.4–4.2) 3 | 2.97 (2.0–4.0) 3 | 4.50 (3.3–5.0) 1,2 | <0.0001 |
| IL-5 | 5.43 (4.1–7.5) | 5.65 (3.8–7.0) | 4.68 (3.0–8.1) | 0.270 |
| IL-6 | 5.31 (1–11) 2,3 | 8.84 (15.5–14) 1 | 8.12 (5.4–15) 1 | 0.010 |
| IL-7 | 8.02 (5.5–10) | 8.05 (6–11) | 7.06 (4.8–9.5) | 0.091 |
| IL-8 | 43.5 (20–50) | 33.6 (17–49) | 21.7 (13–49) | 0.058 |
| IL-9 | 58.5 (12–70) | 14.9 (7.5–69) 3 | 52.6 (38–67) 2 | 0.054 |
| IL-10 | 4.15 (2.3–8.1) | 4.83 (3.1–7.9) | 4.33 (2.6–8.2) | 0.817 |
| IL-12(p70) | 54.7 (28–81) 3 | 37.6 (18–70) 3 | 17.9 (6.9–57) 1,2 | <0.001 |
| IL-13 | 10.35 (5.6–14) 3 | 11.04 (7.6–16) 3 | 6.4 (4.3–9.8) 1,2 | <0.0001 |
| IFNγ | 25.2 (16–34) 2,3 | 31.8 (19–47) 1,3 | 56.5 (28–74) 1,2 | <0.0001 |
| IP-10 | 925 (625–1244) | 745 (490–1124) | 831 (649–1097) | 0.326 |
| EOX1 | 135.5 (110–184) | 138.8 (92–195) | 150.5 (104–181) | 0.888 |
| FGF2 | 23.9 (13–30) | 16.6 (8.6–28) 3 | 28.3 (18–36) 2 | 0.002 |
| G-CSF | 33.9 (31–46) 2,3 | 43.6 (34–65) 1 | 41.0 (34–52) 1 | 0.024 |
| GM-CSF | 3.05 (0.68–7.28) 2 | 6.39 (1.65–12.6) 1,3 | 2.61 (0.06–8.89) 2 | 0.008 |
| MCP-1 | 66.1 (42–76) 3 | 54.3 (27–71) 3 | 24.1 (15–61) 1,2 | <0.0001 |
| MIP-1α | 1.49 (0.2–2.7) 2 | 2.38 (1.4–3.4) 1 | 1.84 (1.5–2.7) | 0.019 |
| MIP-1β | 74.5 (54–96) | 53.5 (32–85) | 74 (42–103) | 0.058 |
| PDGF-BB | 1436 (1171–1970) 3 | 1584 (1021–2406) 3 | 1909 (1245–2757) 1,2 | 0.032 |
| RANTES | 19,865 (912–21,922) 2,3 | 1893 (640–21,105) 1 | 2992 (1898–5199) 1 | 0.025 |
| TNFα | 30.1 (25–34) | 27.0 (22–32) | 30.6 (23–39) | 0.137 |
| VEGF-A | 43.4 (28–103) | 50.2 (26–88) | 48.8 (23–118) | 0.955 |
| Cytokine | AUC (95%CI), p | Sens. and Spec. | J Index | Cut-Off |
|---|---|---|---|---|
| IL-1β | 0.75 (0.65–0.83), p < 0.001 | 70.3% and 84.6% | 0.549 | >0.15 pg/mL |
| IL-1ra | 0.76 (0.66–0.84), p < 0.001 | 81.2% and 61.5% | 0.428 | ≤238.1 pg/mL |
| IL-4 | 0.68 (0.58–0.76), p = 0.001 | 57.8% and 76.9% | 0.347 | >4.15 pg/mL |
| IL-6 | 0.67 (0.57–0.76), p = 0.003 | 75.0% and 53.8% | 0.289 | >5.53 pg/mL |
| IL12(p70) | 0.71 (0.61–0.79), p < 0.001 | 62.5% and 74.4% | 0.369 | ≤29.3 pg/mL |
| IL-13 | 0.66 (0.56–0.75), p = 0.003 | 79.7% and 51.3% | 0.310 | ≤10.3 pg/mL |
| IFNγ | 0.76 (0.66–0.84), p < 0.001 | 54.7% and 94.9% | 0.496 | >54.01 pg/mL |
| G-CSF | 0.62 (0.52–0.71), p = 0.043 | 82.8% and 41.0% | 0.238 | >32.3 pg/mL |
| MCP-1 | 0.72 (0.62–0.80), p < 0.001 | 62.5% and 76.9% | 0.394 | ≤44.4 pg/mL |
| PDGF-BB | 0.64 (0.54–0.73), p = 0.012 | 53.1% and 74.4% | 0.275 | >1891 pg/mL |
| RANTES | 0.67 (0.57–0.76), p = 0.007 | 78.1% and 66.7% | 0.448 | ≤6005 pg/mL |
| Panel 1 1 | 0.87 (0.79–0.93), p < 0.001 | 71.9% and 94.9% | 0.668 | >0.742 3 |
| Panel 2 2 | 0.89 (0.82–0.95), p < 0.001 | 82.8% and 92.3% | 0.751 | >0.595 3 |
| Symbol | Gene Name | Accession No. | Primer Sequence 5′→3′ | Amp. Size (bp) |
|---|---|---|---|---|
| IL71 | Interleukin 7 | NM_000880.4 | F: gacagcatgaaagaaattggtagc R: caacttgcgagcagcacggaat | 117 |
| IL7Ra1 | Interleukin 7 receptor alpha | NM_002185.5 | F: atcgcagcactcactgacctgt R: tcaggcactttacctccacgag | 101 |
| IL101 | Interleukin 10 | NM_000572.3 | F: tctccgagatgccttcagcaga R: tcagacaaggcttggcaaccca | 126 |
| IL10Ra1 | Interleukin 10 receptor alpha | NM_001558.4 | F: gccgaaagaagctacccagtgt R: ggtccaagttcttcagctctgg | 153 |
| ACTA21 | Alpha smooth muscle actin | NM_001141945.2 | F: ctatgcctctggacgcacaact R: cagatccagacgcatgatggca | 115 |
| BCL21 | B-cell lymphoma 2 | NM_000633.3 | F: atcgccctgtggatgactgagt R: gccaggagaaatcaaacagaggc | 127 |
| BCLxL1 | B-cell lymphoma-extra large | NM_001317919.2 | F: gccacttacctgaatgaccacc R: aaccagcggttgaagcgttcct | 131 |
| CCL21 | Monocyte chemoattractant protein 1 (MCP1) | NM_002982.4 | F: agaatcaccagcagcaagtgtcc R: tcctgaacccacttctgcttgg | 98 |
| CDKN1A1 | Cyclin Dependent Kinase Inhibitor 1A (p21CIP1/WAF1) | NM_001220777.2 | F: aggtggacctggagactctcag R: tcctcttggagaagatcagccg | 95 |
| GAPDH2 | Glyceraldehyde-3-phosphate dehydrogenase | NM_001256799.3 | F: tagattattctctgatttggtcgtattgg R: gctcctggaagatggtgatgg | 223 |
| CLDN21 | Claudin 2 | NM_020384.4 | F: gtgacagcagttggcttctcca R: ggagattgcactggatgtcacc | 153 |
| SLC2A11 | Glucose transporter 1 (GLUT1) | NM_006516.4 | F: ttgcaggcttctccaactggac R: cagaaccaggagcacagtgaag | 113 |
| HIF1A1 | Hypoxia-inducible factor 1α | NM_181054.3 | F: tatgagccagaagaacttttaggc R: cacctcttttggcaagcatcctg | 145 |
| Ki671 | Proliferation marker Ki67 | NM_001145966.2 | F: gaaagagtggcaacctgccttc R: gcaccaagttttactacatctgcc | 151 |
| NOS21 | Inducible nitric oxide synthase | NM_000625.4 | F: gctctacacctccaatgtgacc R: ctgccgagatttgagcctcatg | 136 |
| ODC11 | Ornithine decarboxylase | NM_001287189.2 | F: ccaaagcagtctgtcgtctcag R: cagagattgcctgcacgaaggt | 162 |
| PTGS21 | Prostaglandin-endoperoxide synthase 2 (COX2) | NM_000963.4 | F: cggtgaaactctggctagacag R: gcaaaccgtagatgctcaggga | 156 |
| TJP11 | Tight junction protein 1 | NM_001355014.2 | F: gtccagaatctcggaaaagtgcc R: ctttcagcgcaccataccaacc | 132 |
| VEGFA1 | Vascular endothelial growth factor A | NM_001025366.3 | F: ttgccttgctgctctacctcca R: gatggcagtagctgcgctgata | 126 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bednarz-Misa, I.; Fortuna, P.; Diakowska, D.; Jamrozik, N.; Krzystek-Korpacka, M. Distinct Local and Systemic Molecular Signatures in the Esophageal and Gastric Cancers: Possible Therapy Targets and Biomarkers for Gastric Cancer. Int. J. Mol. Sci. 2020, 21, 4509. https://doi.org/10.3390/ijms21124509
Bednarz-Misa I, Fortuna P, Diakowska D, Jamrozik N, Krzystek-Korpacka M. Distinct Local and Systemic Molecular Signatures in the Esophageal and Gastric Cancers: Possible Therapy Targets and Biomarkers for Gastric Cancer. International Journal of Molecular Sciences. 2020; 21(12):4509. https://doi.org/10.3390/ijms21124509
Chicago/Turabian StyleBednarz-Misa, Iwona, Paulina Fortuna, Dorota Diakowska, Natalia Jamrozik, and Małgorzata Krzystek-Korpacka. 2020. "Distinct Local and Systemic Molecular Signatures in the Esophageal and Gastric Cancers: Possible Therapy Targets and Biomarkers for Gastric Cancer" International Journal of Molecular Sciences 21, no. 12: 4509. https://doi.org/10.3390/ijms21124509
APA StyleBednarz-Misa, I., Fortuna, P., Diakowska, D., Jamrozik, N., & Krzystek-Korpacka, M. (2020). Distinct Local and Systemic Molecular Signatures in the Esophageal and Gastric Cancers: Possible Therapy Targets and Biomarkers for Gastric Cancer. International Journal of Molecular Sciences, 21(12), 4509. https://doi.org/10.3390/ijms21124509