The Effects of Mutual Interaction of Orexin-A and Glucagon-Like Peptide-1 on Reflex Swallowing Induced by SLN Afferents in Rats

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
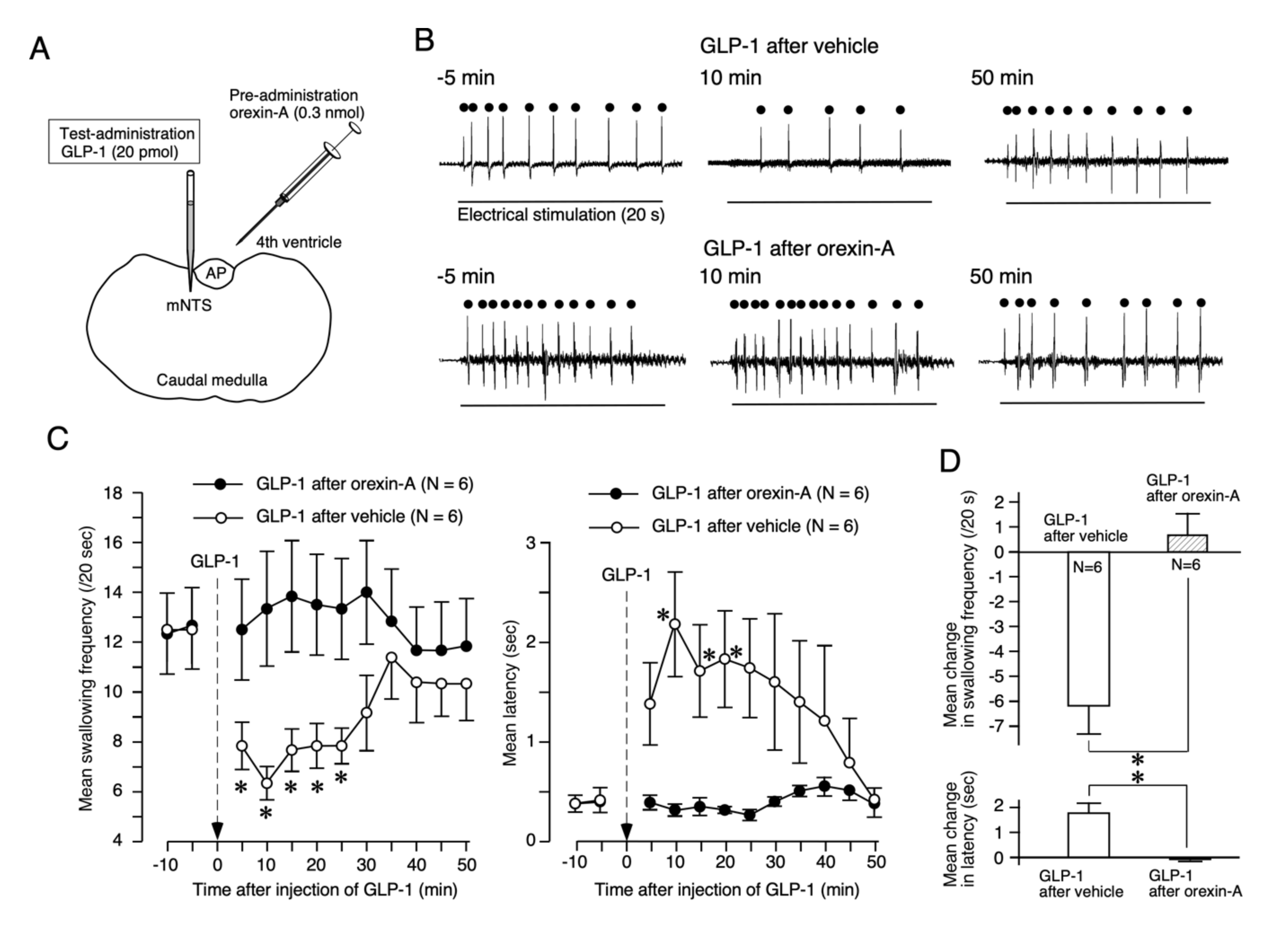
2.1. Orexin-A Suppresses the GLP-1 Response of Reflex Swallowing
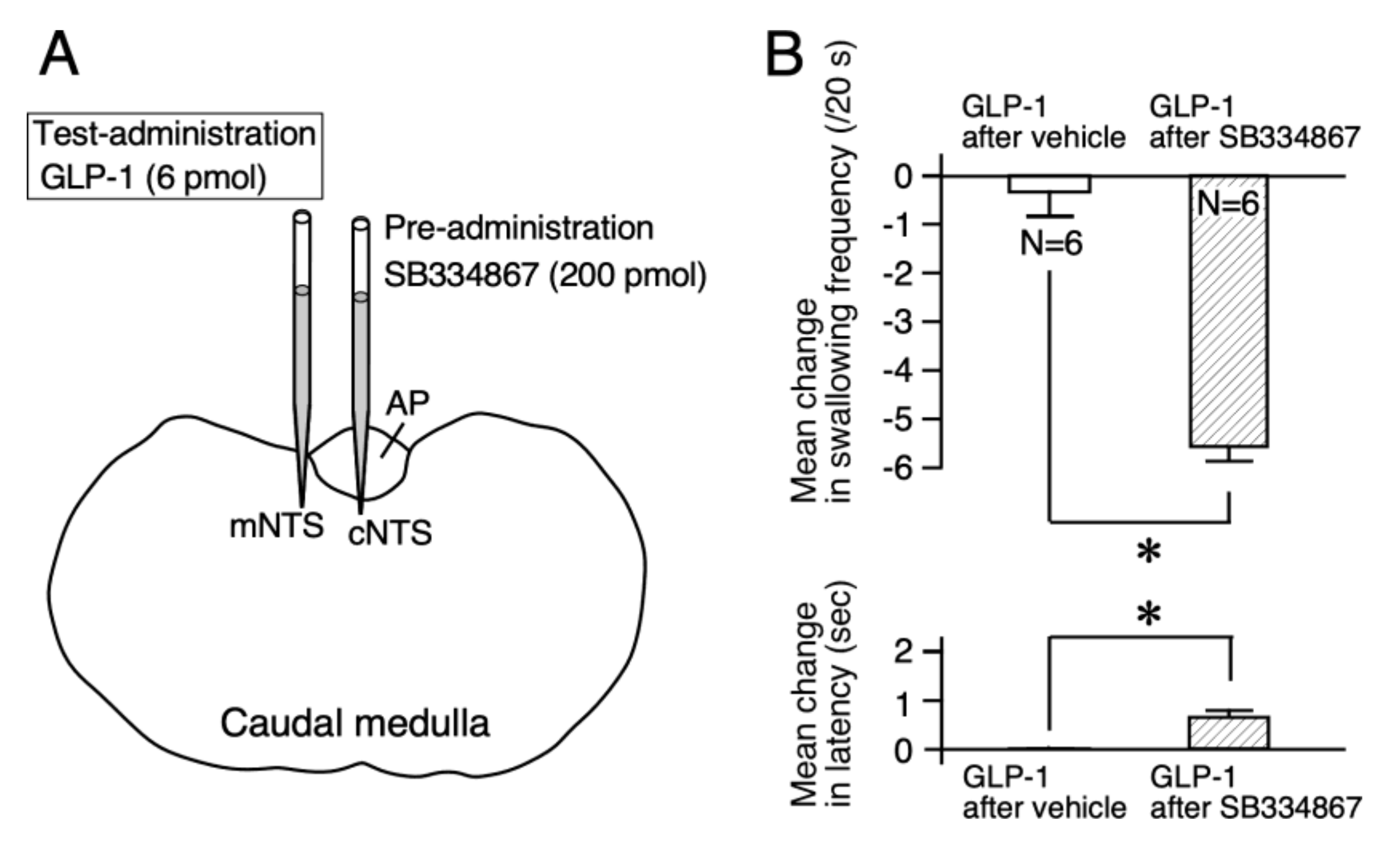
2.2. An Orexin-1 Receptor Antagonist Facilitates the GLP-1 Response of Reflex Swallowing
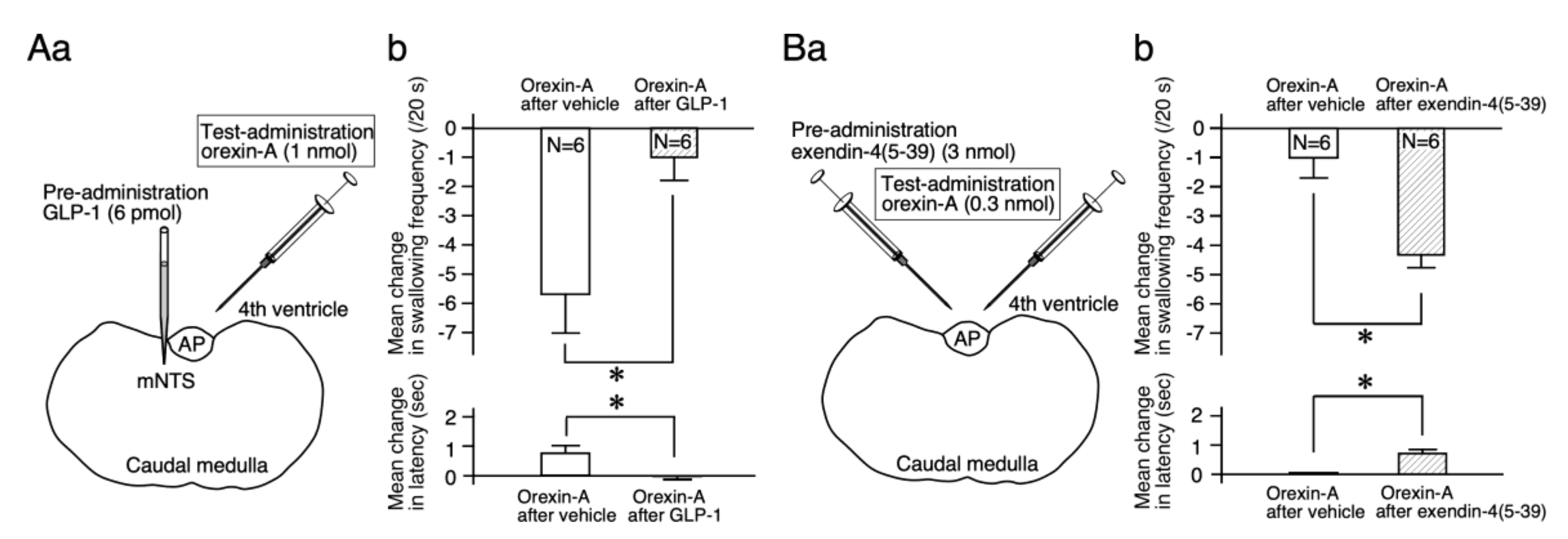
2.3. GLP-1 Suppresses the Orexin-A Effects on Reflex Swallowing
2.4. A GLP-1 Receptor Antagonist Facilitates the Orexin-A Effects on Reflex Swallowing
3. Discussion
4. Materials and Methods
4.1. Animals and Surgery
4.2. Electrical Stimulation and Recording
4.3. Drugs and Administration
4.4. Data Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tsuji, T.; Yamamoto, T.; Tanaka, S.; Bakhshishayan, S.; Kogo, M. Analyses of the facilitatory effect of orexin on eating and masticatory muscle activity in rats. J. Neurophysiol. 2011, 106, 3129–3135. [Google Scholar] [CrossRef] [PubMed]
- Kobashi, M.; Yanagihara, M.; Fujita, M.; Mitoh, Y.; Matsuo, R. Fourth ventricular administration of ghrelin induces relaxation of the proximal stomach in the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R217–R223. [Google Scholar] [CrossRef] [PubMed]
- Kobashi, M.; Furudono, Y.; Matsuo, R.; Yamamoto, T. Central orexin facilitates gastric relaxation and contractility in rats. Neurosci. Lett. 2002, 332, 171–174. [Google Scholar] [CrossRef]
- Koga, T.; Kobashi, M.; Mizutani, M.; Tsukamoto, G.; Matsuo, R. Area postrema mediates gastric motor response induced by apomorphine in rats. Brain Res. 2003, 960, 122–131. [Google Scholar] [CrossRef]
- Kobashi, M.; Xuan, S.Y.; Fujita, M.; Mitoh, Y.; Matsuo, R. Central ghrelin inhibits reflex swallowing elicited by activation of the superior laryngeal nerve in the rat. Regul. Pept. 2010, 160, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Kobashi, M.; Mizutani, S.; Fujita, M.; Mitoh, Y.; Shimatani, Y.; Matsuo, R. Central orexin inhibits reflex swallowing elicited by the superior laryngeal nerve via caudal brainstem in the rat. Physiol. Behav. 2014, 130, 6–12. [Google Scholar] [CrossRef]
- Felix, B.; Jean, A.; Roman, C. Leptin inhibits swallowing in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R657–R663. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guillebaud, F.; Roussel, G.; Felix, B.; Troadec, J.D.; Dallaporta, M.; Abysique, A. Interaction between nesfatin-1 and oxytocin in the modulation of the swallowing reflex. Brain Res. 2019, 1711, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Kobashi, M.; Mizutani, S.; Fujita, M.; Mitoh, Y.; Shimatani, Y.; Matsuo, R. Central glucagon like peptide-1 inhibits reflex swallowing elicited by the superior laryngeal nerve via caudal brainstem in the rat. Brain Res. 2017, 1671, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Woods, S.C.; Porte, D., Jr.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef]
- Date, Y.; Toshinai, K.; Koda, S.; Miyazato, M.; Shimbara, T.; Tsuruta, T.; Niijima, A.; Kangawa, K.; Nakazato, M. Peripheral interaction of ghrelin with cholecystokinin on feeding regulation. Endocrinology 2005, 146, 3518–3525. [Google Scholar] [CrossRef]
- Asakawa, A.; Inui, A.; Inui, T.; Katsuura, G.; Fujino, M.A.; Kasuga, M. Orexin reverses cholecystokinin-induced reduction in feeding. Diabetes Obes. Metab. 2002, 4, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Parise, E.M.; Lilly, N.; Kay, K.; Dossat, A.M.; Seth, R.; Overton, J.M.; Williams, D.L. Evidence for the role of hindbrain orexin-1 receptors in the control of meal size. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1692–R1699. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, K.; Kaji, T. The acute anorexic effect of liraglutide, a GLP-1 receptor agonist, does not require functional leptin receptor, serotonin, and hypothalamic POMC and CART activities in mice. Diabetes Res. Clin. Pract. 2016, 120, 186–189. [Google Scholar] [CrossRef]
- De Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.; Gautvik, V.T.; Bartlett, F.S.; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef]
- Zheng, H.; Patterson, L.M.; Berthoud, H.R. Orexin—A projections to the caudal medulla and orexin-induced c-Fos expression, food intake, and autonomic function. J. Comp. Neurol. 2005, 485, 127–142. [Google Scholar] [CrossRef]
- Krowicki, Z.K.; Burmeister, M.A.; Berthoud, H.R.; Scullion, R.T.; Fuchs, K.; Hornby, P.J. Orexins in rat dorsal motor nucleus of the vagus potently stimulate gastric motor function. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G465–G472. [Google Scholar] [CrossRef]
- Takahashi, N.; Okumura, T.; Yamada, H.; Kohgo, Y. Stimulation of gastric acid secretion by centrally administered orexin—A in conscious rats. Biochem. Biophys Res. Comm. 1999, 254, 623–627. [Google Scholar] [CrossRef]
- Burcelin, R.; Da Costa, A.; Drucker, D.; Thorens, B. Glucose competence of the hepatoportal vein sensor requires the presence of an activated glucagon-like peptide-1 receptor. Diabetes 2001, 50, 1720–1728. [Google Scholar] [CrossRef]
- Holst, J.J.; Deacon, C.F. Glucagon-like peptide-1 mediates the therapeutic actions of DPP-IV inhibitors. Diabetologia 2005, 48, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, M.; Nakabayashi, H.; Uchida, K.; Nakagawa, A.; Niijima, A. The hepatic vagal nerve is receptive to incretin hormone glucagon-like peptide-1, but not to glucose-dependent insulinotropic polypeptide, in the portal vein. J. Auton. Nerv. Syst. 1996, 61, 149–154. [Google Scholar] [CrossRef]
- Jin, S.L.; Han, V.K.; Simmons, J.G.; Towle, A.C.; Lauder, J.M.; Lund, P.K. Distribution of glucagonlike peptide I (GLP-I), glucagon, and glicentin in the rat brain: An immunocytochemical study. J. Comp. Neurol. 1988, 271, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Larsen, P.J.; Tang-Christensen, M.; Holst, J.J.; Orskov, C. Distribution of glucagon-like peptide-1 and other preproglucagon-derived peptides in the rat hypothalamus and brainstem. Neuroscience 1997, 77, 257–270. [Google Scholar] [CrossRef]
- Fong, A.Y.; Stornetta, R.L.; Foley, C.M.; Potts, J.T. Immunohistochemical localization of GAD67-expressing neurons and processes in the rat brainstem: Subregional distribution in the nucleus tractus solitarius. J. Comp. Neurol. 2005, 493, 274–290. [Google Scholar] [CrossRef]
- Mostafeezur, R.M.; Zakir, H.M.; Takatsuji, H.; Yamada, Y.; Yamamura, K.; Kitagawa, J. Cannabinoids facilitate the swallowing reflex elicited by the superior laryngeal nerve stimulation in rats. PLoS ONE 2012, 7, e50703. [Google Scholar] [CrossRef]
- Card, J.P.; Johnson, A.L.; Llewellyn-Smith, I.J.; Zheng, H.; Anand, R.; Brierley, D.I.; Trapp, S.; Rinaman, L. GLP-1 neurons form a local synaptic circuit within the rodent nucleus of the solitary tract. J. Comp. Neurol. 2018, 526, 2149–2164. [Google Scholar] [CrossRef]
- Shih, C.D.; Chuang, Y.C. Nitric oxide and GABA mediate bi-directional cardiovascular effects of orexin in the nucleus tractus solitarii of rats. Neuroscience 2007, 149, 625–635. [Google Scholar] [CrossRef]
- Cai, X.J.; Evans, M.L.; Lister, C.A.; Leslie, R.A.; Arch, J.R.; Wilson, S.; Williams, G. Hypoglycemia activates orexin neurons and selectively increases hypothalamic orexin-B levels: Responses inhibited by feeding and possibly mediated by the nucleus of the solitary tract. Diabetes 2001, 50, 105–112. [Google Scholar] [CrossRef]
- Cai, X.J.; Widdowson, P.S.; Harrold, J.; Wilson, S.; Buckingham, R.E.; Arch, J.R.; Tadayyon, M.; Clapham, J.C.; Wilding, J.; Williams, G. Hypothalamic orexin expression: Modulation by blood glucose and feeding. Diabetes 1999, 48, 2132–2137. [Google Scholar] [CrossRef]
- Griffond, B.; Risold, P.Y.; Jacquemard, C.; Colard, C.; Fellmann, D. Insulin-induced hypoglycemia increases preprohypocretin (orexin) mRNA in the rat lateral hypothalamic area. Neurosci. Lett. 1999, 262, 77–80. [Google Scholar] [CrossRef]
- Moriguchi, T.; Sakurai, T.; Nambu, T.; Yanagisawa, M.; Goto, K. Neurons containing orexin in the lateral hypothalamic area of the adult rat brain are activated by insulin-induced acute hypoglycemia. Neurosci. Lett. 1999, 264, 101–104. [Google Scholar] [CrossRef]
- Paranjape, S.A.; Vavaiya, K.K.; Kale, A.Y.; Briski, K.P. Habituation of insulin-induced hypoglycemic transcription activation of lateral hypothalamic orexin-A-containing neurons to recurring exposure. Regul. Pept. 2006, 135, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, C.; Goke, R.; Richter, G.; Fehmann, H.C.; Arnold, R.; Goke, B. Glucagon-like peptide-1 and glucose-dependent insulin-releasing polypeptide plasma levels in response to nutrients. Digestion 1995, 56, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Orskov, C.; Wettergren, A.; Holst, J.J. Secretion of the incretin hormones glucagon-like peptide-1 and gastric inhibitory polypeptide correlates with insulin secretion in normal man throughout the day. Scand. J. Gastroenterol. 1996, 31, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Vrang, N.; Phifer, C.B.; Corkern, M.M.; Berthoud, H.R. Gastric distension induces c-Fos in medullary GLP-1/2-containing neurons. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R470–R478. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: Burlington, NJ, USA, 2007. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobashi, M.; Shimatani, Y.; Fujita, M.; Mitoh, Y.; Yoshida, R.; Matsuo, R. The Effects of Mutual Interaction of Orexin-A and Glucagon-Like Peptide-1 on Reflex Swallowing Induced by SLN Afferents in Rats. Int. J. Mol. Sci. 2020, 21, 4422. https://doi.org/10.3390/ijms21124422
Kobashi M, Shimatani Y, Fujita M, Mitoh Y, Yoshida R, Matsuo R. The Effects of Mutual Interaction of Orexin-A and Glucagon-Like Peptide-1 on Reflex Swallowing Induced by SLN Afferents in Rats. International Journal of Molecular Sciences. 2020; 21(12):4422. https://doi.org/10.3390/ijms21124422
Chicago/Turabian StyleKobashi, Motoi, Yuichi Shimatani, Masako Fujita, Yoshihiro Mitoh, Ryusuke Yoshida, and Ryuji Matsuo. 2020. "The Effects of Mutual Interaction of Orexin-A and Glucagon-Like Peptide-1 on Reflex Swallowing Induced by SLN Afferents in Rats" International Journal of Molecular Sciences 21, no. 12: 4422. https://doi.org/10.3390/ijms21124422
APA StyleKobashi, M., Shimatani, Y., Fujita, M., Mitoh, Y., Yoshida, R., & Matsuo, R. (2020). The Effects of Mutual Interaction of Orexin-A and Glucagon-Like Peptide-1 on Reflex Swallowing Induced by SLN Afferents in Rats. International Journal of Molecular Sciences, 21(12), 4422. https://doi.org/10.3390/ijms21124422