Mining for Candidate Genes Controlling Secondary Growth of the Carrot Storage Root

 , ,
, , 
Abstract
1. Introduction
2. Results
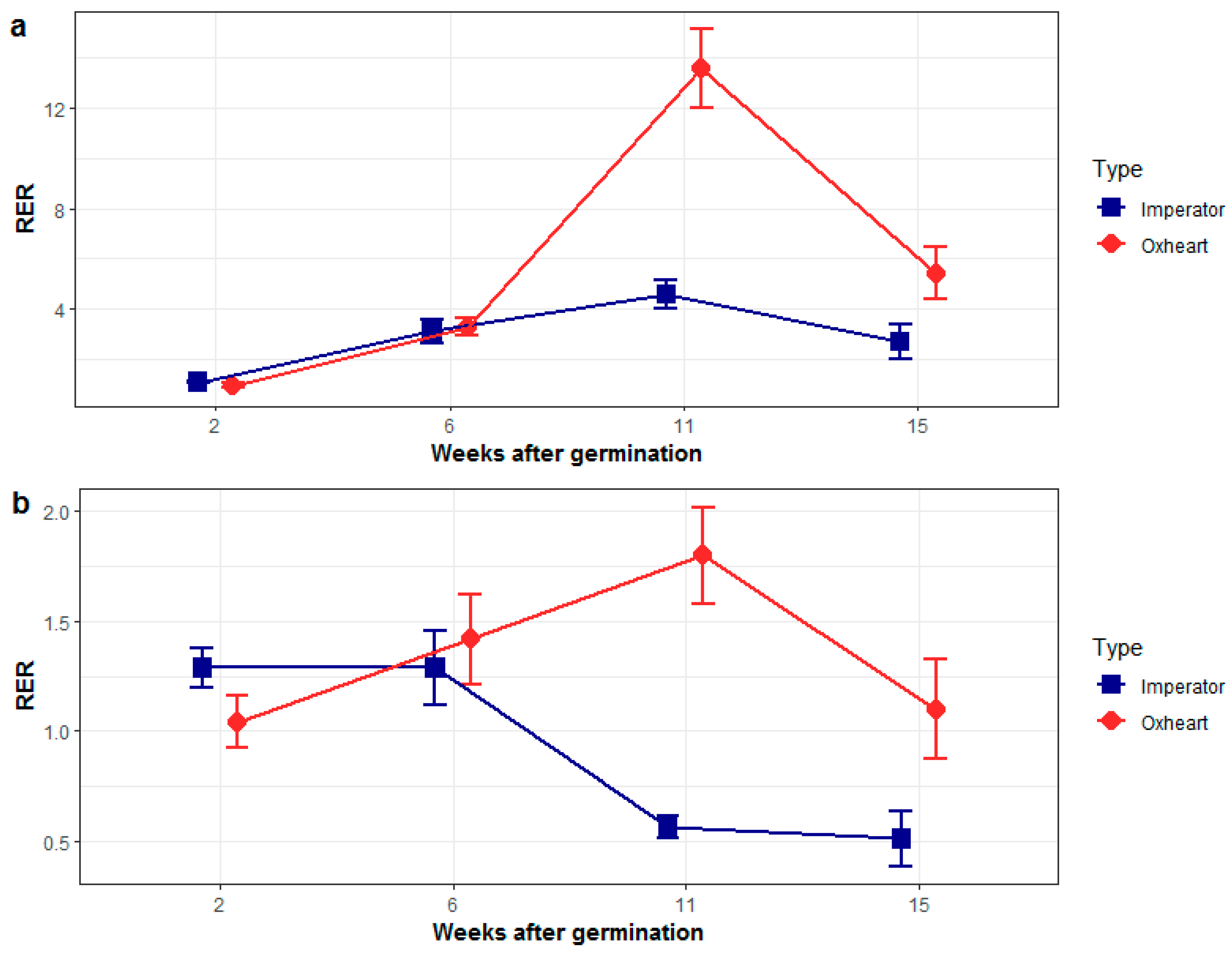
2.1. Phenotyping and Genotyping
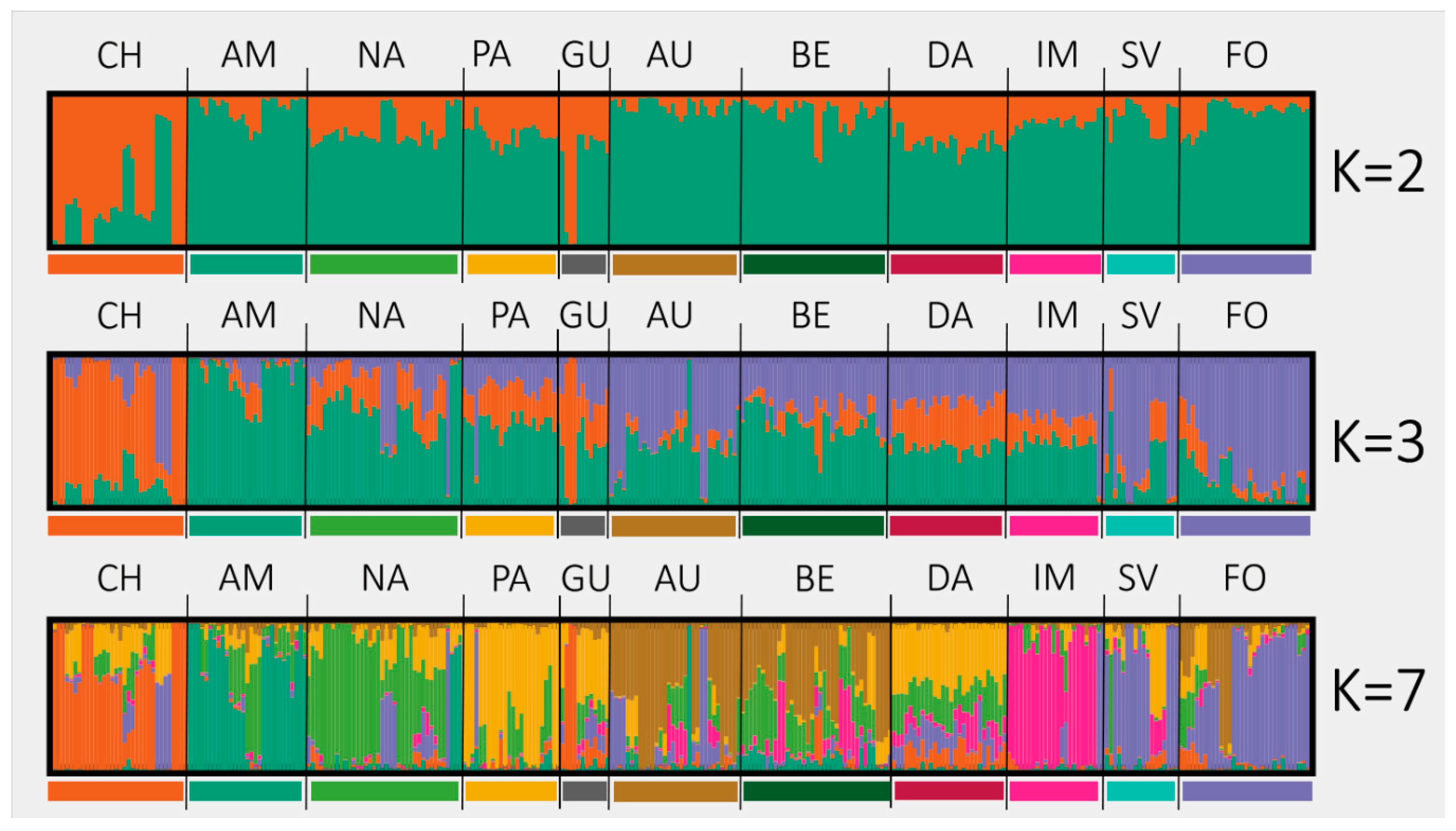
2.2. Population Structure
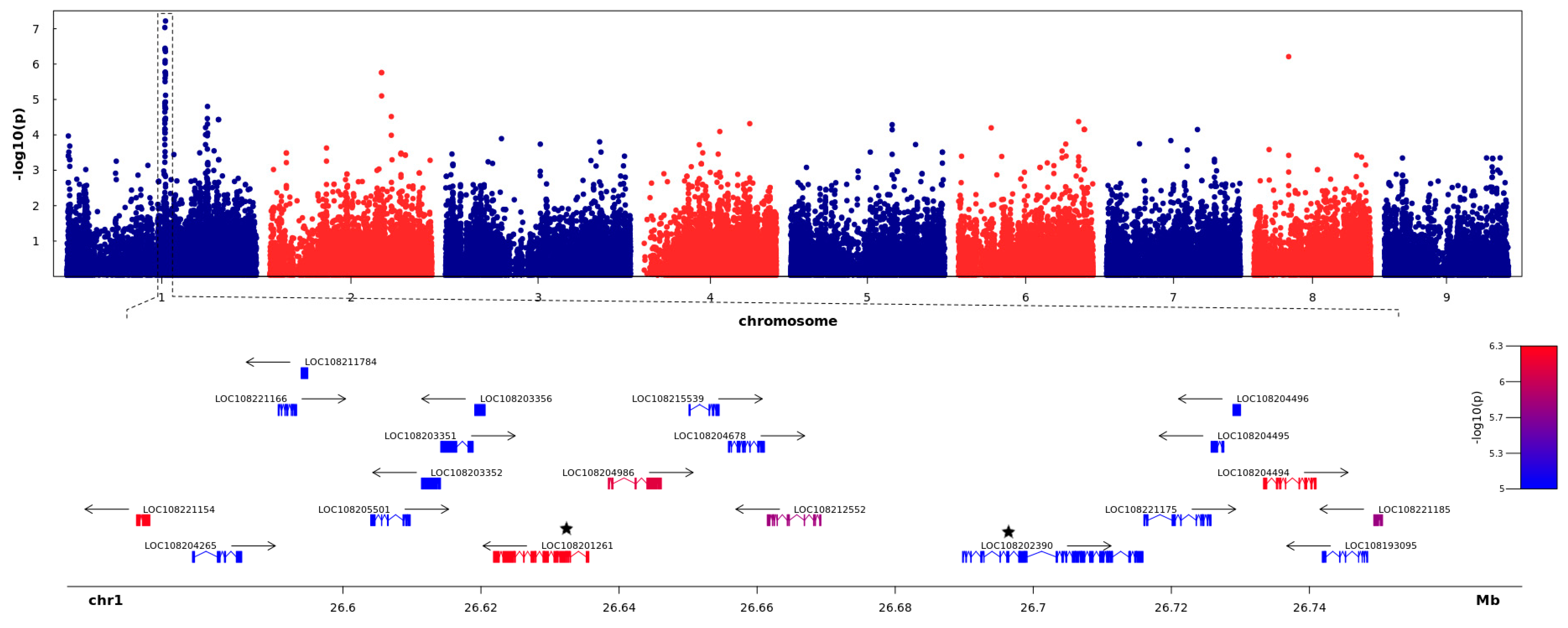
2.3. GWAS
2.4. Identification and Verification of the Candidate Gene
2.5. Genotyping and Expression of Candidate Genes in the Developing Storage Roots
3. Discussion
4. Materials and Methods
4.1. Phenotyping
4.2. Genotyping
4.3. Population Structure
4.4. Association Analysis
4.5. Identification and Validation of Candidate Genes
4.6. Experimental Validation
4.6.1. SNP Genotyping
4.6.2. RT-qPCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BTAF1 | TATA-binding protein-associated factor 1 |
| DCAF1 | DDB1-CUL4 Associated Factor 1 |
| GBS | genotyping-by-sequencing |
| GWAS | genome wide association studies |
| HSD | honestly significant difference |
| LD | linkage disequilibrium |
| QTL | Quantitative traits loci |
| SNP | Single nucleotide polymorphism |
References
- Banga, O. The development of the original European carrot material. Euphytica 1957, 6, 64–76. [Google Scholar] [CrossRef]
- Thompson, R. Some factors affecting carrot root shape and size. Euphytica 1969, 18, 277–285. [Google Scholar] [CrossRef]
- White, J.M. Carrot yield when grown under three soil water concentrations. Hort Sci. 1992, 27, 105–106. [Google Scholar] [CrossRef]
- Evers, A.M.; Tuuri, H.; Hägg, M.; Plaami, S.; Häkkinen, U.; Talvitie, H. Soil forming and plant density effects on carrot yield and internal quality. Plant. Foods Hum. Nutr 1997, 51, 283–294. [Google Scholar] [CrossRef]
- Sakamoto, M.; Suzuki, T. Elevated root-zone temperature modulates growth and quality of hydroponically grown carrots. Agric. Sci. 2015, 6, 749. [Google Scholar] [CrossRef]
- Mbatha, A.N.; Ceronio, G.M.; Coetzer, G.M. Response of carrot (Daucus carota L.) yield and quality to organic fertiliser. S. Afr. J. Plant. Soil 2014, 31, 1–6. [Google Scholar] [CrossRef]
- Jagosz, B. Combining ability of carrot (Daucus carota L.) lines and heritability of yield and its quality components. Folia Hortic. 2012, 24, 115–122. [Google Scholar] [CrossRef]
- Turner, S.D.; Ellison, S.; Senalik, D.A.; Simon, P.W.; Spalding, E.; Miller, N. An automated image analysis pipeline enables genetic studies of shoot and root morphology in carrot (Daucus carota L.). Front. Plant. Sci 2018, 9, 1703. [Google Scholar] [CrossRef]
- Ellison, S. Carrot Domestication. In The Carrot Genome; Simon, P.W., Iorizzo, M., Grzebelus, D., Baranski, R., Eds.; Springer: Cham, Switzerland, 2019; pp. 77–91. [Google Scholar] [CrossRef]
- Grzebelus, D.; Iorizzo, M.; Senalik, D.; Ellison, S.; Cavagnaro, P.; Macko-Podgorni, A.; Simon, P.W. Diversity, genetic mapping, and signatures of domestication in the carrot (Daucus carota L.) genome, as revealed by Diversity Arrays Technology (DArT) markers. Mol. Breed. 2014, 33, 625–637. [Google Scholar] [CrossRef]
- Ellison, S.L.; Luby, C.H.; Corak, K.E.; Coe, K.M.; Senalik, D.; Iorizzo, M.; Dawson, J.C. Carotenoid Presence Is Associated with the Or Gene in Domesticated Carrot. Genetics 2018, 210, 1497–1508. [Google Scholar] [CrossRef]
- Macko-Podgórni, A.; Iorizzo, M.; Smółka, K.; Simon, P.W.; Grzebelus, D. Conversion of a diversity arrays technology marker differentiating wild and cultivated carrots to a co-dominant cleaved amplified polymorphic site marker. Acta Biochim. Pol. 2014, 61. [Google Scholar]
- Macko-Podgórni, A.; Machaj, G.; Stelmach, K.; Senalik, D.; Grzebelus, E.; Iorizzo, M.; Grzebelus, D. Characterization of a genomic region under selection in cultivated carrot (Daucus carota subsp. sativus) reveals a candidate domestication gene. Front. Plant. Sci. 2017, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Machaj, G.; Bostan, H.; Macko-Podgórni, A.; Iorizzo, M.; Grzebelus, D. Comparative transcriptomics of root development in wild and cultivated carrots. Genes 2018, 9, 431. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Bradbury, P.J.; Brown, P.J.; Hung, H.; Sun, Q.; Flint-Garcia, S.; Buckler, E.S. Genome-wide association study of leaf architecture in the maize nested association mapping population. Nat. Genet. 2011, 43, 159. [Google Scholar] [CrossRef]
- Beyer, S.; Daba, S.; Tyagi, P.; Bockelman, H.; Brown-Guedira, G.; Mohammadi, M. Loci and candidate genes controlling root traits in wheat seedlings—A wheat root GWAS. Funct. Integr. Genom. 2019, 19, 91–107. [Google Scholar] [CrossRef]
- Huang, X.; Yang, S.; Gong, J.; Zhao, Y.; Feng, Q.; Gong, H.; Chen, N. Genomic analysis of hybrid rice varieties reveals numerous superior alleles that contribute to heterosis. Nat. Comm. 2015, 6, 6258. [Google Scholar] [CrossRef]
- Keilwagen, J.; Lehnert, H.; Berner, T.; Budahn, H.; Nothnagel, T.; Ulrich, D.; Dunemann, F. The Terpene Synthase Gene Family of Carrot (Daucus carota L.): Identification of QTLs and Candidate Genes Associated with Terpenoid Volatile Compounds. Front. Plant. Sci. 2017, 8, 1930. [Google Scholar] [CrossRef]
- Jourdan, M.; Gagné, S.; Dubois-Laurent, C.; Maghraoui, M.; Huet, S.; Suel, A.; Geoffriau, E. Carotenoid content and root color of cultivated carrot: A candidate-gene association study using an original broad unstructured population. PLoS ONE 2015, 10, e0116674. [Google Scholar] [CrossRef]
- Iorizzo, M.; Ellison, S.; Senalik, D.; Zeng, P.; Satapoomin, P.; Huang, J.; Yildiz, M. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657. [Google Scholar] [CrossRef]
- Whitney, S.M.; Birch, R.; Kelso, C.; Beck, J.L.; Kapralov, M.V. Improving recombinant Rubisco biogenesis, plant photosynthesis and growth by coexpressing its ancillary RAF1 chaperone. Proc. Natl. Acad. Sci. USA 2015, 112, 3564–3569. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Beavis, W.; Berardini, T.Z.; Chen, G.; Dixon, D.; Doyle, A.; Miller, N. The Arabidopsis Information Resource (TAIR): A model organism database providing a centralized, curated gateway to Arabidopsis biology, research materials and community. Nucleic Acids Res. 2003, 31, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Perilli, S.; Di Mambro, R.; Sabatini, S. Growth and development of the root apical meristem. Curr. Opin. Plant. Biol. 2012, 15, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Elo, A.; Immanen, J.; Nieminen, K.; Helariutta, Y. Stem cell function during plant vascular development. In Seminars in Cell & Developmental Biology; Davey, J., Ed.; Academic Press: London, UK, 2009; Volume 20, pp. 1097–1106. [Google Scholar] [CrossRef]
- Wang, G.L.; Jia, X.L.; Xu, Z.S.; Wang, F.; Xiong, A.S. Sequencing, assembly, annotation, and gene expression: Novel insights into the hormonal control of carrot root development revealed by a high-throughput transcriptome. Mol. Genet. Genom. 2015, 290, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, W.; Li, M.; Xu, Z.; Wang, F.; Xiong, A. Expression profiles of genes involved in jasmonic acid biosynthesis and signaling during growth and development of carrot. Acta Biochim. Biophys. Sin. 2016, 48, 795–803. [Google Scholar] [CrossRef]
- Wang, G.L.; Sun, S.; Xing, G.M.; Wu, X.J.; Wang, F.; Xiong, A.S. Morphological characteristics, anatomical structure, and gene expression: Novel insights into cytokinin accumulation during carrot growth and development. PLoS ONE 2015, 10, e0134166. [Google Scholar] [CrossRef]
- Que, F.; Wang, G.L.; Li, T.; Wang, Y.H.; Xu, Z.S.; Xiong, A.S. Genome-wide identification, expansion, and evolution analysis of homeobox genes and their expression profiles during root development in carrot. Funct. Integr. Genom. 2018, 18, 685–700. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, S.; Chen, F.; Chen, H.; Wang, J.; McCall, C.; Deng, X.W. Arabidopsis DDB1-CUL4 ASSOCIATED FACTOR1 forms a nuclear E3 ubiquitin ligase with DDB1 and CUL4 that is involved in multiple plant developmental processes. Plant. Cell 2008, 20, 1437–1455. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, S.; Terzaghi, W.; Deng, X.W. A new family of plant E3 ubiquitin ligases. Plant. Signal. Behav. 2008, 3, 1049–1052. [Google Scholar] [CrossRef][Green Version]
- Seo, K.I.; Lee, J.H.; Nezames, C.D.; Zhong, S.; Song, E.; Byun, M.O.; Deng, X.W. ABD1 is an Arabidopsis DCAF substrate receptor for CUL4-DDB1–based E3 ligases that acts as a negative regulator of abscisic acid signaling. Plant. Cell 2014, 26, 695–711. [Google Scholar] [CrossRef]
- Yu, F.; Wu, Y.; Xie, Q. Ubiquitin–proteasome system in ABA signaling: From perception to action. Mol. Plant. 2016, 9, 21–33. [Google Scholar] [CrossRef]
- Zhang, J.; Hafeez, M.T.; Di, D.; Wu, L.; Zhang, L. Precise control of ABA signaling through post-translational protein modification. Plant. Growth Regul. 2019, 88, 99–111. [Google Scholar] [CrossRef]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The role and regulation of ABI5 (ABA-Insensitive 5) in plant development, abiotic stress responses and phytohormone crosstalk. Front. Plant. Sci. 2016, 7, 1884. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Gemenet, D.C.; Villordon, A. Root system architecture and abiotic stress tolerance: Current knowledge in root and tuber crops. Front. Plant. Sci. 2016, 7, 1584. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Zhu, M.; Yu, J.; Han, R.; Tang, C.; Xu, T.; Li, Z. RNA-Seq and iTRAQ reveal multiple pathways involved in storage root formation and development in sweet potato (Ipomoea batatas L.). BMC Plant. Biol. 2019, 19, 136. [Google Scholar] [CrossRef]
- Koster, M.J.; Snel, B.; Timmers, H.T.M. Genesis of chromatin and transcription dynamics in the origin of species. Cell 2015, 161, 724–736. [Google Scholar] [CrossRef]
- Klejman, M.P.; Zhao, X.; van Schaik, F.M.; Herr, W.; Timmers, H.T.M. Mutational analysis of BTAF1–TBP interaction: BTAF1 can rescue DNA-binding defective TBP mutants. Nucleic Acids Res. 2005, 33, 5426–5436. [Google Scholar] [CrossRef]
- Tamaki, H.; Konishi, M.; Daimon, Y.; Aida, M.; Tasaka, M.; Sugiyama, M. Identification of novel meristem factors involved in shoot regeneration through the analysis of temperature-sensitive mutants of Arabidopsis. Plant. J. 2009, 57, 1027–1039. [Google Scholar] [CrossRef]
- Ćosić, T.; Raspor, M.; Savić, J.; Cingel, A.; Matekalo, D.; Zdravković-Korać, S.; Ninković, S. Expression profiles of organogenesis-related genes over the time course of one-step de novo shoot organogenesis from intact seedlings of kohlrabi. J. Plant. Physiol. 2019, 232, 257–269. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 26 May 2020).
- Briard, M.; Le Clerc, M.; Grzebelus, D.; Senalik, D.; Simon, P.W. Modified protocols for rapid carrot genomic DNA extraction and AFLP analysis using silver staining or radioisotopes. Plant. Mol. Biol. Rep. 2000, 18, 235–241. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A High Capacity Genotyping by Sequencing Analysis Pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; McVean, G. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Browning, S.R.; Browning, B.L. Rapid and accurate haplotype phasing and missing data inference for whole genome association studies by use of localized haplotype clustering. Am. J. Hum. Genet. 2007, 81, 1084–1097. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.A.; Donnelly, P. Association mapping in structured populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Stelmach, K.; Macko-Podgórni, A.; Machaj, G.; Grzebelus, D. Miniature inverted repeat transposable element insertions provide a source of intron length polymorphism markers in the carrot (Daucus carota L.). Front. Plant. Sci. 2017, 8, 725. [Google Scholar] [CrossRef] [PubMed]
- Frichot, E.; François, O. LEA: An R package for landscape and ecological association studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Phanstiel, D.H.; Boyle, A.P.; Araya, C.L.; Snyder, M.P.; Sushi, R. Flexible, quantitative and integrative genomic visualizations for publication-quality multi-panel figures. Bioinformatics 2014, 30, 2808–2810. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Bowman, M.J.; Willis, D.K.; Simon, P.W. Transcript abundance of phytoene synthase 1 and phytoene synthase 2 is associated with natural variation of storage root carotenoid pigmentation in carrot. J. Am. Soc. Hortic Sci. 2014, 139, 63–68. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Mean [mm] | Range [mm] | SD | CV% |
|---|---|---|---|---|
| shoulder | 41.73 | 19.42–73.76 | 11.06 | 26.51 |
| length | 213.86 | 60.00–365.00 | 57.99 | 27.11 |
| collar | 15.98 | 5.08–30.98 | 5.30 | 33.19 |
| tip | 21.64 | 5.25–46.04 | 8.17 | 37.79 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macko-Podgórni, A.; Stelmach, K.; Kwolek, K.; Machaj, G.; Ellison, S.; Senalik, D.A.; Simon, P.W.; Grzebelus, D. Mining for Candidate Genes Controlling Secondary Growth of the Carrot Storage Root. Int. J. Mol. Sci. 2020, 21, 4263. https://doi.org/10.3390/ijms21124263
Macko-Podgórni A, Stelmach K, Kwolek K, Machaj G, Ellison S, Senalik DA, Simon PW, Grzebelus D. Mining for Candidate Genes Controlling Secondary Growth of the Carrot Storage Root. International Journal of Molecular Sciences. 2020; 21(12):4263. https://doi.org/10.3390/ijms21124263
Chicago/Turabian StyleMacko-Podgórni, Alicja, Katarzyna Stelmach, Kornelia Kwolek, Gabriela Machaj, Shelby Ellison, Douglas A. Senalik, Philipp W. Simon, and Dariusz Grzebelus. 2020. "Mining for Candidate Genes Controlling Secondary Growth of the Carrot Storage Root" International Journal of Molecular Sciences 21, no. 12: 4263. https://doi.org/10.3390/ijms21124263
APA StyleMacko-Podgórni, A., Stelmach, K., Kwolek, K., Machaj, G., Ellison, S., Senalik, D. A., Simon, P. W., & Grzebelus, D. (2020). Mining for Candidate Genes Controlling Secondary Growth of the Carrot Storage Root. International Journal of Molecular Sciences, 21(12), 4263. https://doi.org/10.3390/ijms21124263