Optimization of microRNA Acquirement from Seminal Plasma and Identification of Diminished Seminal microRNA-34b as Indicator of Low Semen Concentration

 , , and
, , and
Abstract
1. Introduction
2. Results
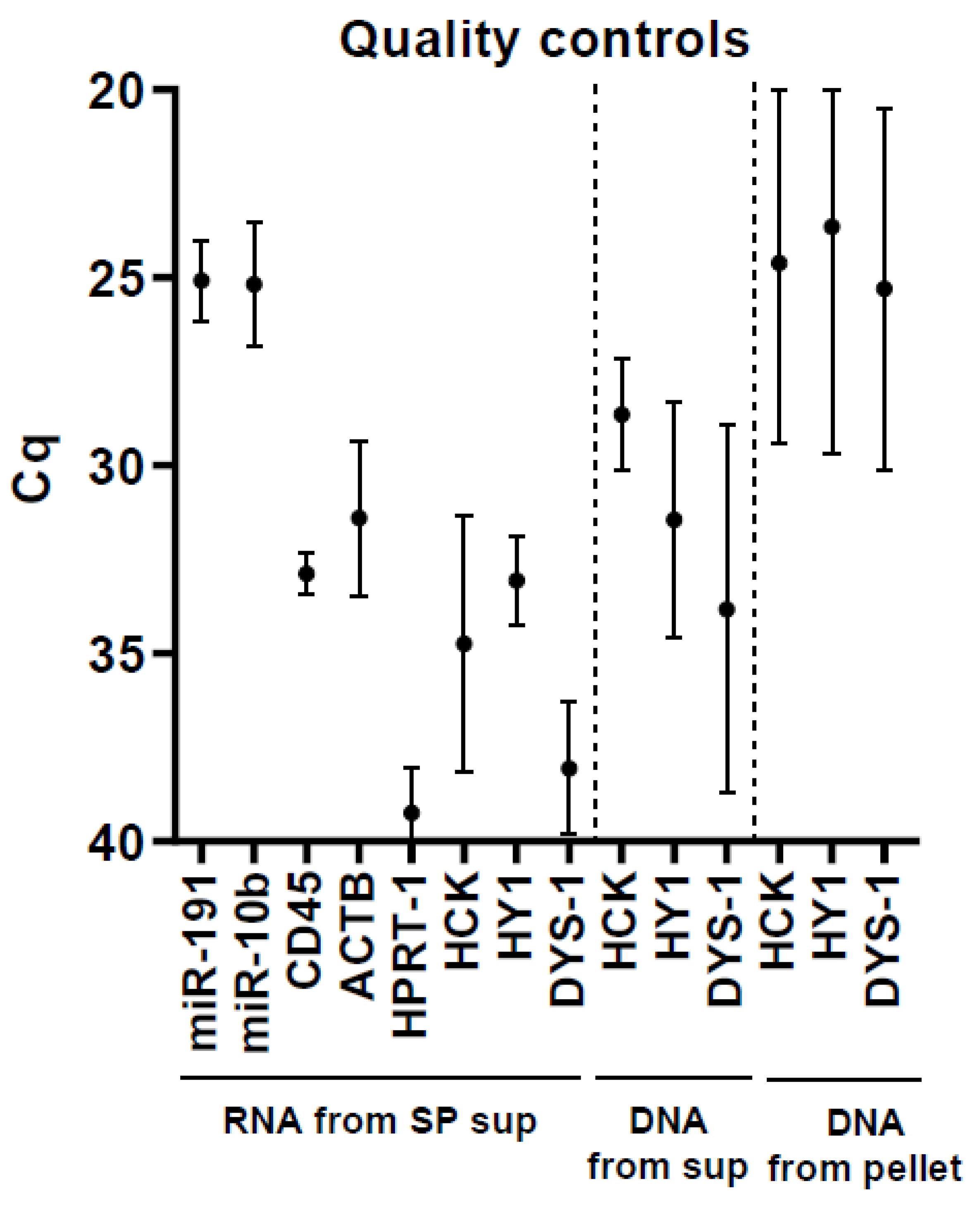
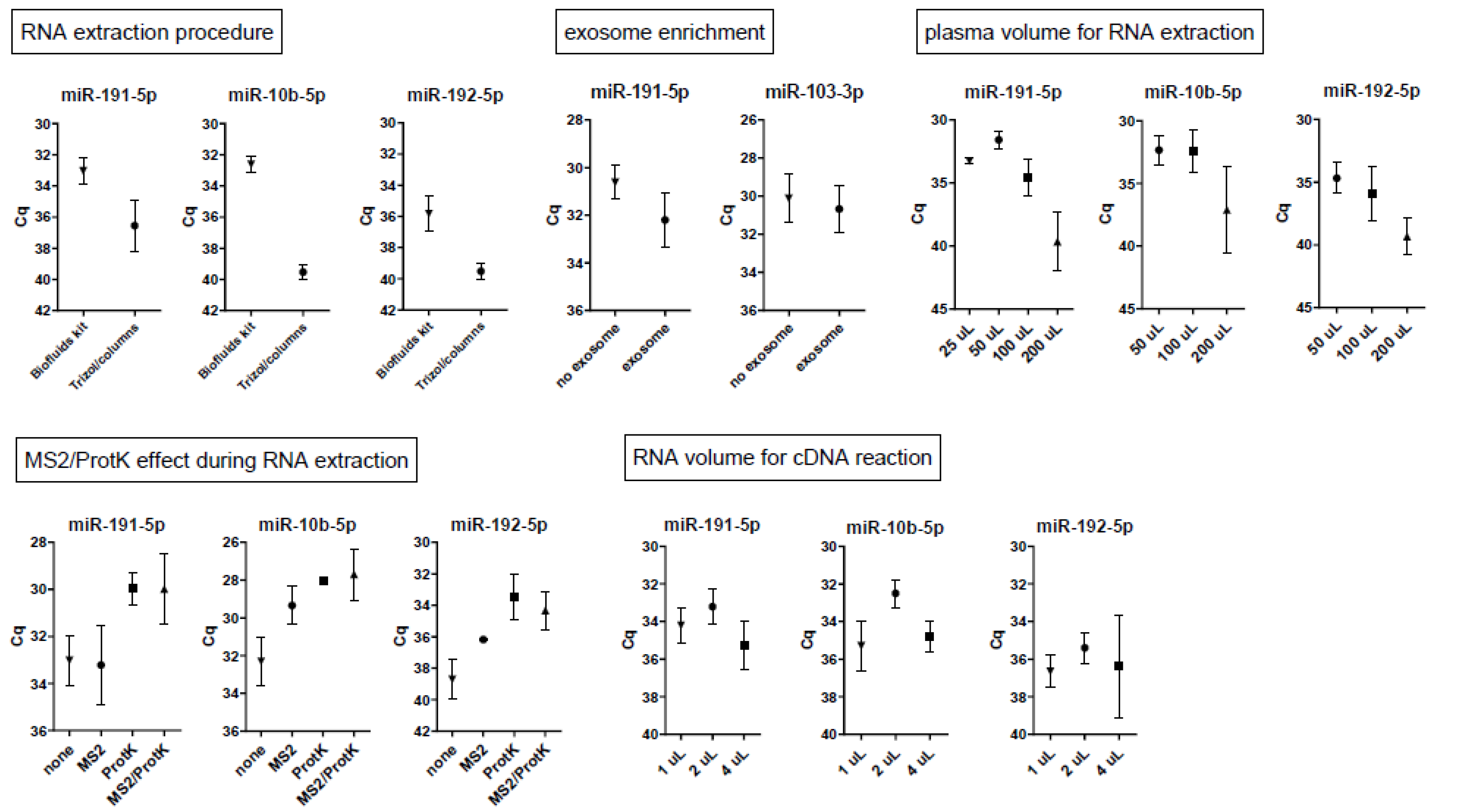
2.1. Optimization of microRNA Acquirement from Seminal Plasma
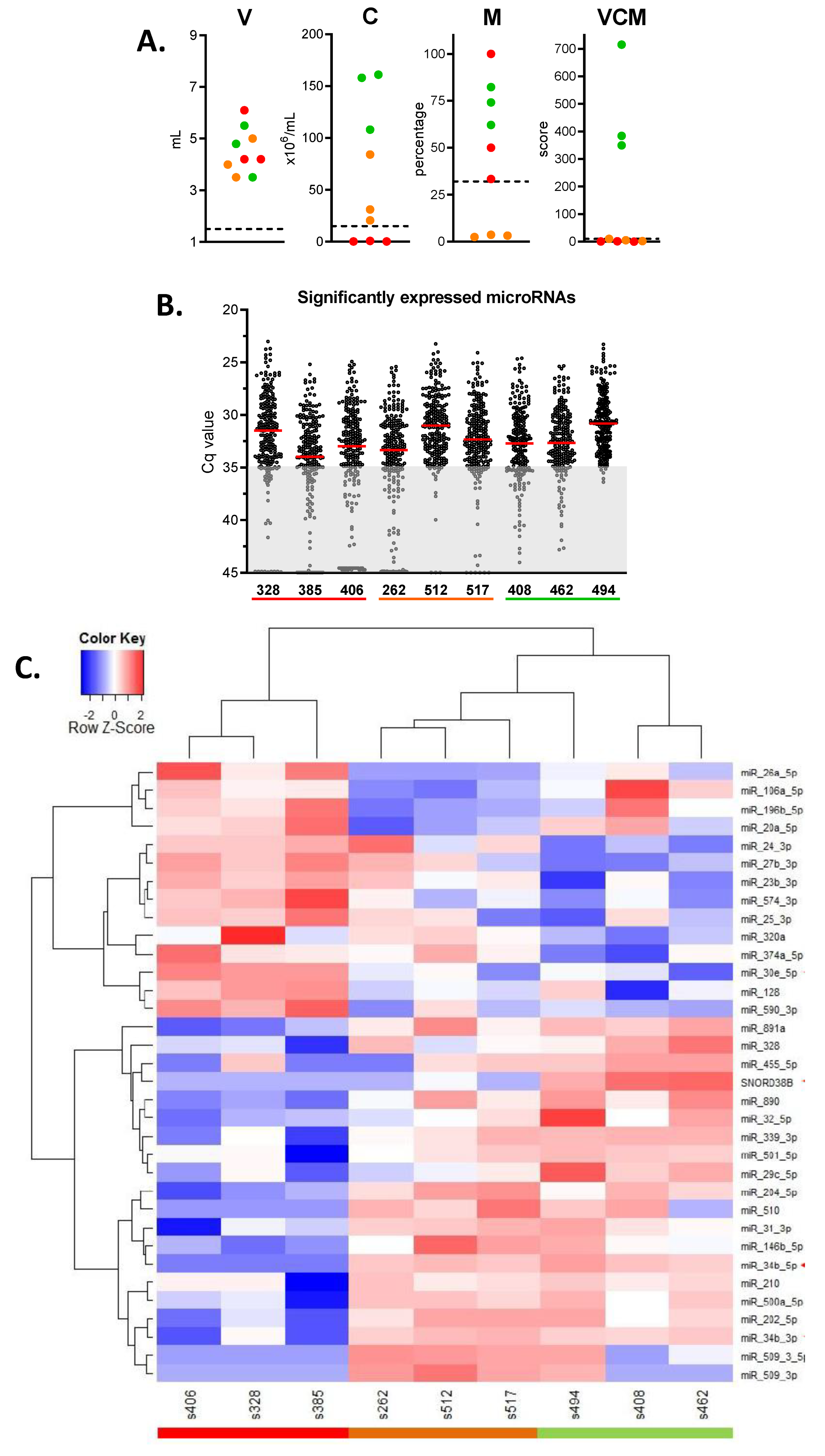
2.2. Identification of microRNAs Related to Semen Quality: Test Phase
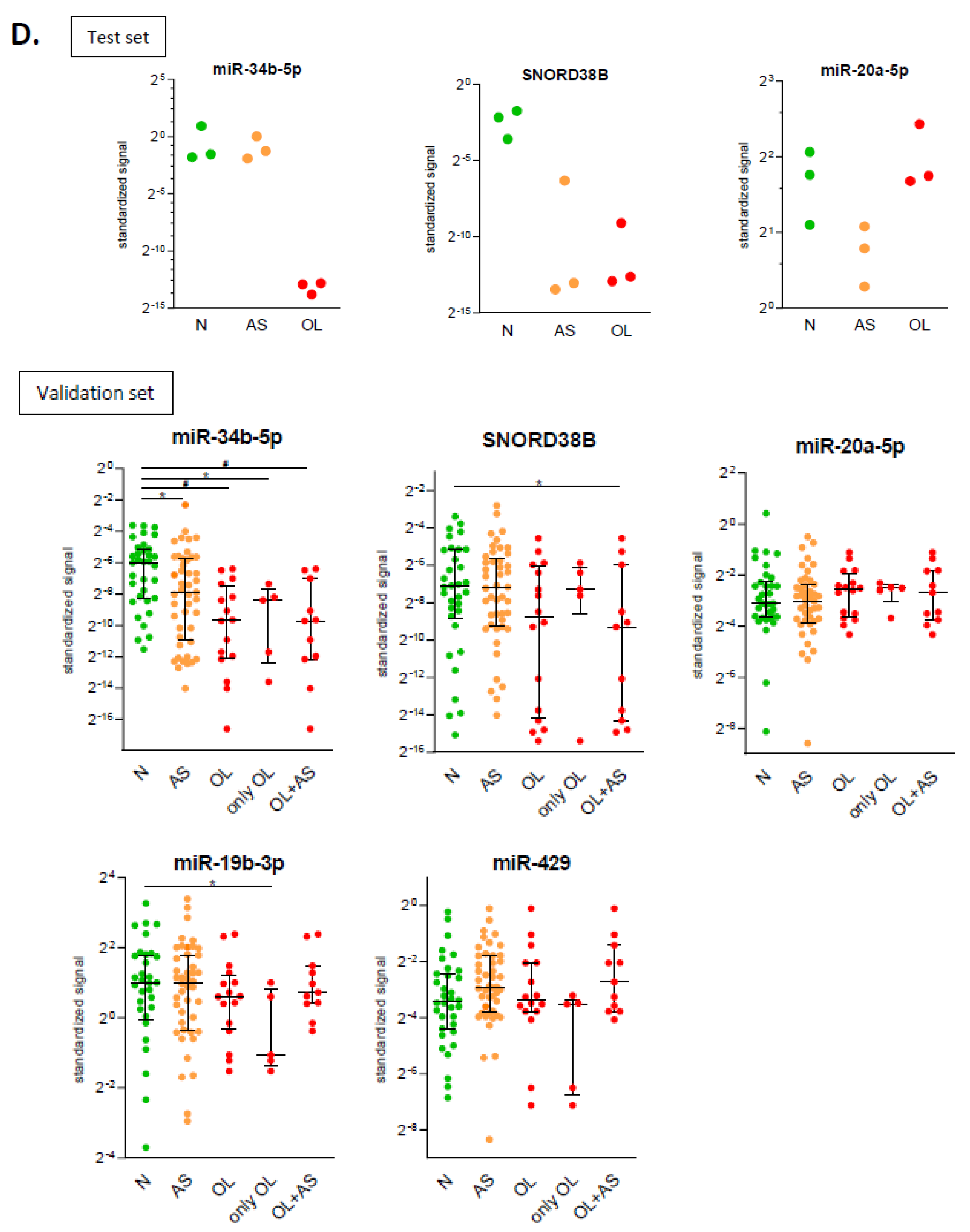
2.3. Identification of microRNAs Related to Semen Quality: Validation Phase
3. Discussion
4. Materials and Methods
4.1. Study Cohort and Processing of Samples
4.2. Optimization of RNA Extraction from Seminal Plasmas
4.3. Complementary DNA Synthesis and PCR Amplification
4.4. Identification of microRNAs Related to Semen Characteristics: Test Phase
4.5. Identification of microRNAs Related to Semen Characteristics: Validation Phase
4.6. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Matzuk, M.M.; Lamb, D.J. Genetic dissection of mammalian fertility pathways. Nat. Cell. Biol. 2002, 4, s41–s49. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination of Human Semen Ans Sperm-Cervical Mucus Interaction; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- WHO Laboratory Manual for the Examination and Processing of Human Semen; WHO Press: Geneva, Switzerland, 2010.
- Bracke, A.; Peeters, K.; Punjabi, U.; Hoogewijs, D.; Dewilde, S. A search for molecular mechanisms underlying male idiopathic infertility. Reprod. Biomed. Online 2018, 36, 327–339. [Google Scholar] [CrossRef]
- Wu, W.; Hu, Z.; Qin, Y.; Dong, J.; Dai, J.; Lu, C.; Zhang, W.; Shen, H.; Xia, Y.; Wang, X. Seminal plasma microRNAs: Potential biomarkers for spermatogenesis status. Mol. Hum. Reprod. 2012, 18, 489–497. [Google Scholar] [CrossRef]
- Vashisht, A.; Gahlay, G.K. Using miRNAs as diagnostic biomarkers for male infertility: Opportunities and challenges. Mol. Hum. Reprod. 2020, 26, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Kiani, M.; Salehi, M.; Mogheiseh, A. MicroRNA expression in infertile men: Its alterations and effects. Zygote 2019, 27, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Z. MicroRNAs as oncogenes and tumor suppressors. N. Engl. J. Med. 2005, 353, 1768–1771. [Google Scholar] [CrossRef]
- Kim, V.N. MicroRNA biogenesis: Coordinated cropping and dicing. Nat. Rev. Mol. Cell Boil. 2005, 6, 376–385. [Google Scholar] [CrossRef]
- Wu, W.; Qin, Y.; Li, Z.; Dong, J.; Dai, J.; Lu, C.; Guo, X.; Zhao, Y.; Zhu, Y.; Zhang, W.; et al. Genome-wide microRNA expression profiling in idiopathic non-obstructive azoospermia: Significant up-regulation of miR-141, miR-429 and miR-7-1-3p. Hum. Reprod. 2013, 28, 1827–1836. [Google Scholar] [CrossRef]
- Wang, C.; Yang, C.; Chen, X.; Yao, B.; Yang, C.; Zhu, C.; Li, L.; Wang, J.; Li, X.; Shao, Y.; et al. Altered Profile of Seminal Plasma MicroRNAs in the Molecular Diagnosis of Male Infertility. Clin. Chem. 2011, 57, 1722–1731. [Google Scholar] [CrossRef]
- Liu, T.; Cheng, W.; Gao, Y.; Wang, H.; Liu, Z. Microarray analysis of microRNA expression patterns in the semen of infertile men with semen abnormalities. Mol. Med. Rep. 2012, 6, 535–542. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Galata, V.; Backes, C.; Keller, A.; Hammadeh, M.; Meese, E. MicroRNA signature in spermatozoa and seminal plasma of proven fertile men and in testicular tissue of men with obstructive azoospermia. Andrology 2019, 52, e13503. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Carrell, D.T.; Jenkins, T.G.; Yeste, M. The role of miRNAs in male human reproduction: A systematic review. Andrology 2019, 8, 7–26. [Google Scholar] [CrossRef]
- McAlexander, M.A.; Phillips, M.J.; Witwer, K.W. Comparison of Methods for miRNA Extraction from Plasma and Quantitative Recovery of RNA from Cerebrospinal Fluid. Front. Genet. 2013, 4, 83. [Google Scholar] [CrossRef]
- Andreasen, D.; Fog, J.U.; Biggs, W.; Salomon, J.; Dahslveen, I.K.; Baker, A.; Mouritzen, P. Improved microRNA quantification in total RNA from clinical samples. Methods 2010, 50, S6–S9. [Google Scholar] [CrossRef]
- Moret, I.; Sánchez-Izquierdo, D.; Iborra, M.; Tortosa, L.; Navarro-Puche, A.; Nos, P.; Cervera, J.; Beltrán, B. Assessing an Improved Protocol for Plasma microRNA Extraction. PLoS ONE 2013, 8, e82753. [Google Scholar] [CrossRef]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The Majority of MicroRNAs Detectable in Serum and Saliva Is Concentrated in Exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef]
- Vojtech, L.; Woo, S.; Hughes, S.M.; Levy, C.; Ballweber, L.; Sauteraud, R.P.; Strobl, J.; Westerberg, K.; Gottardo, R.; Tewari, M.; et al. Exosomes in human semen carry a distinctive repertoire of small non-coding RNAs with potential regulatory functions. Nucleic Acids Res. 2014, 42, 7290–7304. [Google Scholar] [CrossRef]
- Li, H.; Huang, S.; Guo, C.; Guan, H.; Xiong, C. Cell-Free Seminal mRNA and MicroRNA Exist in Different Forms. PLoS ONE 2012, 7, e34566. [Google Scholar] [CrossRef]
- Tomlinson, M.J.; Kessopoulou, E.; Barratt, C.L. The diagnostic and prognostic value of traditional semen parameters. J. Androl. 1999, 20, 588–593. [Google Scholar]
- Bonde, J.P.E.; Ernst, E.; Jensen, M.B.; Hjollund, N.H.; Kolstad, H.; Hviid, T.V.F.; Giwercman, A.; Skakkebæk, N.E.; Henriksen, T.B.; Olsen, J. Relation between semen quality and fertility: A population-based study of 430 first-pregnancy planners. Lancet 1998, 352, 1172–1177. [Google Scholar] [CrossRef]
- Eikmans, M.; Rekers, N.V.; Anholts, J.D.H.; Heidt, S.; Claas, F.H.J. Blood cell mRNAs and microRNAs: Optimized protocols for extraction and preservation. Blood 2013, 121, e81–e89. [Google Scholar] [CrossRef]
- Mestdagh, P.; Hartmann, N.; Baeriswyl, L.; Andreasen, D.; Bernard, N.; Chen, C.; Cheo, D.; D’Andrade, P.; DeMayo, M.; Dennis, L.; et al. Evaluation of quantitative miRNA expression platforms in the microRNA quality control (miRQC) study. Nat. Methods 2014, 11, 809–815. [Google Scholar] [CrossRef]
- Lu, L.-F.; Liston, A. MicroRNA in the immune system, microRNA as an immune system. Immunology 2009, 127, 291–298. [Google Scholar] [CrossRef]
- Anglicheau, D.; Muthukumar, T.; Suthanthiran, M. MicroRNAs: Small RNAs with big effects. Transplant 2010, 90, 105–112. [Google Scholar] [CrossRef]
- Robertson, S.A.; Guerin, L.R.; Moldenhauer, L.M.; Hayball, J.D. Activating T regulatory cells for tolerance in early pregnancy—The contribution of seminal fluid. J. Reprod. Immunol. 2009, 83, 109–116. [Google Scholar] [CrossRef]
- Robertson, S.A.; Sharkey, D.J. The role of semen in induction of maternal immune tolerance to pregnancy. Semin. Immunol. 2001, 13, 243–254. [Google Scholar] [CrossRef]
- Craenmehr, M.; Van Der Keur, C.; Anholts, J.; Kapsenberg, J.; Van Der Westerlaken, L.; Van Kooten, C.; Claas, F.H.; Heidt, S.; Eikmans, M. Effect of seminal plasma on dendritic cell differentiation in vitro depends on the serum source in the culture medium. J. Reprod. Immunol. 2020, 137, 103076. [Google Scholar] [CrossRef]
- Turner, M.; Galloway, A.; Vigorito, E. Noncoding RNA and its associated proteins as regulatory elements of the immune system. Nat. Immunol. 2014, 15, 484–491. [Google Scholar] [CrossRef]
- Baltimore, D.; Boldin, M.; O’Connell, R.M.; Rao, D.S.; Taganov, K.D. MicroRNAs: New regulators of immune cell development and function. Nat. Immunol. 2008, 9, 839–845. [Google Scholar] [CrossRef]
- Yadav, R.P.; Kotaja, N. Small RNAs in spermatogenesis. Mol. Cell. Endocrinol. 2014, 382, 498–508. [Google Scholar] [CrossRef]
- Kotaja, N. MicroRNAs and spermatogenesis. Fertil. Steril. 2014, 101, 1552–1562. [Google Scholar] [CrossRef]
- Hilz, S.; Modzelewski, A.J.; Cohen, P.E.; Grimson, A. The roles of microRNAs and siRNAs in mammalian spermatogenesis. Development 2016, 143, 3061–3073. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, E.; Liu, H.; Rotunno, M.; Koshiol, J.; Marincola, F.M.; Landi, M.T.; McShane, L.M. Evaluation of normalization methods for two-channel microRNA microarrays. J. Transl. Med. 2010, 8, 69. [Google Scholar] [CrossRef]
- Mestdagh, P.; Van Vlierberghe, P.; De Weer, A.; Muth, D.; Westermann, F.; Speleman, F.; Vandesompele, J. A novel and universal method for microRNA RT-qPCR data normalization. Genome Boil. 2009, 10, R64. [Google Scholar] [CrossRef]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef]
- Occhipinti, G.; Giulietti, M.; Principato, G.; Piva, F. The choice of endogenous controls in exosomal microRNA assessments from biofluids. Tumor Boil. 2016, 37, 11657–11665. [Google Scholar] [CrossRef]
- Zhang, H.-T.; Zhang, Z.; Hong, K.; Tang, W.-H.; Liu, D.-F.; Mao, J.-M.; Yang, Y.-Z.; Lin, H.-C.; Jiang, H. Altered microRNA profiles of testicular biopsies from patients with nonobstructive azoospermia. Asian J. Androl. 2019, 22, 100–105. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Hammadeh, M.; Backes, C.; Fischer, U.; Leidinger, P.; Lubbad, A.M.; Keller, A.; Meese, E. Panel of five microRNAs as potential biomarkers for the diagnosis and assessment of male infertility. Fertil. Steril. 2014, 102, 989–997.e1. [Google Scholar] [CrossRef]
- Muñoz, X.; Mata, A.; Bassas, L.; Larriba, S. Altered miRNA Signature of Developing Germ-cells in Infertile Patients Relates to the Severity of Spermatogenic Failure and Persists in Spermatozoa. Sci. Rep. 2015, 5, 17991. [Google Scholar] [CrossRef]
- Cui, L.; Fang, L.; Shi, B.; Qiu, S.; Ye, Y. Spermatozoa micro ribonucleic acid–34c level is correlated with intracytoplasmic sperm injection outcomes. Fertil. Steril. 2015, 104, 312–317.e1. [Google Scholar] [CrossRef]
- Comazzetto, S.; Di Giacomo, M.; Rasmussen, K.D.; Much, C.; Azzi, C.; Perlas, E.; Morgan, M.; O’Carroll, D. Oligoasthenoteratozoospermia and Infertility in Mice Deficient for miR-34b/c and miR-449 Loci. PLoS Genet. 2014, 10, e1004597. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Tang, C.; Zhang, Y.; Wu, J.; Bao, J.; Zheng, H.; Xu, C.; Yan, W. mir-34b/c and mir-449a/b/c are required for spermatogenesis, but not for the first cleavage division in mice. Boil. Open 2015, 4, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Chen, Y.; Yang, R.; Bai, Y.; Li, C.; Li, H.; Xiong, C. miR-424/322 is downregulated in the semen of patients with severe DNA damage and may regulate sperm DNA damage. Reprod. Fertil. Dev. 2015, 28, 1598–1607. [Google Scholar] [CrossRef]
- Meuleman, T.; Snaterse, G.; Van Beelen, E.; Anholts, J.; Pilgram, G.; Van Der Westerlaken, L.; Eikmans, M.; Claas, F. The immunomodulating effect of seminal plasma on T cells. J. Reprod. Immunol. 2015, 110, 109–116. [Google Scholar] [CrossRef]
- Barratt, C.L.; Björndahl, L.; Menkveld, R.; Mortimer, D. ESHRE special interest group for andrology basic semen analysis course: A continued focus on accuracy, quality, efficiency and clinical relevance. Hum. Reprod. 2011, 26, 3207–3212. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | |
|---|---|---|
| CD45 | 5′-GGTCGTCAAACAAAAACTTCCC-3′ | 5′-TGAGATCCATCCCTGCAGTG-3′ |
| HPRT-1 | 5′-AGATGGTCAAGGTCGCAAGC-3′ | 5′-TCAAGGGCATATCCTACAACAAAC-3′ |
| ACTB | 5′-ACCACACCTTCTACAATGAG-3′ | 5′-TAGCACAGCCTGGATAGC-3′ |
| HCK | 5′-TATTAGCACCATCCATAGGAGGCTT-3′ | 5′-CTTCCGCTCCACTTTCCCTAAC-3′ |
| HY | 5′-TGGCGATTAAGTCAAATTCGC-3′ | 5′-CCCCCTAGTACCCTGACAATGTATT-3′ |
| DYS-1 | 5′-TCCTGCTTATCCAAATTCACCAT-3′ | 5′-ACTTCCCTCTGACATTACCTGATAATTG-3′ |
| N vs. Low-C | N vs. Low-M | Low-C vs. Low-M | K-W | Average Cq | |
|---|---|---|---|---|---|
| miR-23b-3p | x | x | - | x | 28.6 |
| miR-27b-3p | x | x | - | x | 28.9 |
| miR-99b-5p | x | x | - | x | 29.8 |
| miR-152 | x | x | - | x | 30.8 |
| SNORD38b 2 | x | x | - | x | 33.7 |
| miR-204-5p | x | - | x | x | 31.7 |
| miR-221-3p | x | - | x | x | 32.1 |
| miR-34b-5p 2 | x | - | x | x | 32.3 |
| miR-146b-5p | x | - | x | x | 32.6 |
| miR-510 | x | - | x | x | 37.9 |
| miR-100-5p | - | x | x | x | 31.1 |
| miR-509-3p | - | x | x | x | 39.9 |
| miR-891a | x | - | - | - | 27.2 |
| miR-888-5p | x | - | - | - | 28.2 |
| miR-25-3p | x | - | - | - | 29.5 |
| miR-574-3p | x | - | - | x | 30.3 |
| miR-374b-5p | x | - | - | - | 30.3 |
| miR-30d-5p | x | - | - | - | 31.4 |
| miR-598 | x | - | - | - | 33.6 |
| miR-32-5p | x | - | - | x | 33.6 |
| miR-16-1-3p | x | - | - | x | 33.9 |
| miR-501-5p | x | - | - | - | 34.1 |
| miR-590-3p | x | - | - | - | 34.7 |
| miR-339-3p | x | - | - | x | 35.2 |
| miR-34c-3p | x | - | - | - | 36.7 |
| miR-99a-5p | - | x | - | - | 26.4 |
| miR-106a-5p | - | x | - | - | 27.8 |
| miR-24-3p | - | x | - | - | 28.4 |
| let-7d-3p | - | x | - | - | 31.6 |
| miR-376c-3p | - | x | - | - | 33.9 |
| miR-328 | - | x | - | - | 34.1 |
| miR-665 | - | x | - | x | 36.1 |
| miR-142-3p | - | x | - | - | 37.0 |
| miR-455-5p | - | x | - | - | 37.8 |
| miR-26a-5p | - | - | x | - | 27.1 |
| miR-20a-5p | - | - | x | - | 28.8 |
| miR-205-5p | - | - | x | - | 28.1 |
| miR-222-3p | - | - | x | - | 29.4 |
| miR-196b-5p | - | - | x | x | 30.5 |
| miR-532-5p | - | - | x | - | 32.1 |
| miR-892a | - | - | x | - | 32.1 |
| miR-18b-5p | - | - | x | - | 33.0 |
| miR-202-5p | - | - | x | x | 34.7 |
| miR-455-3p | - | - | x | - | 35.3 |
| miR-500a-5p | - | - | x | x | 35.3 |
| miR-502-3p | - | - | x | - | 35.8 |
| miR-31-3p | - | - | x | - | 35.9 |
| miR-181b-5p | - | - | x | x | 35.9 |
| miR-34b-3p | - | - | x | x | 36.2 |
| miR-202-3p | - | - | x | x | 36.8 |
| miR-502-5p | - | - | x | - | 37.4 |
| miR-449a | - | - | x | - | 37.9 |
| miR-335-3p | - | - | x | x | 38.9 |
| miR-509-3-5p | - | - | x | x | 39.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eikmans, M.; D. H. Anholts, J.; Blijleven, L.; Meuleman, T.; van Beelen, E.; van der Hoorn, M.-L.P.; Claas, F.H.J. Optimization of microRNA Acquirement from Seminal Plasma and Identification of Diminished Seminal microRNA-34b as Indicator of Low Semen Concentration. Int. J. Mol. Sci. 2020, 21, 4089. https://doi.org/10.3390/ijms21114089
Eikmans M, D. H. Anholts J, Blijleven L, Meuleman T, van Beelen E, van der Hoorn M-LP, Claas FHJ. Optimization of microRNA Acquirement from Seminal Plasma and Identification of Diminished Seminal microRNA-34b as Indicator of Low Semen Concentration. International Journal of Molecular Sciences. 2020; 21(11):4089. https://doi.org/10.3390/ijms21114089
Chicago/Turabian StyleEikmans, Michael, Jacqueline D. H. Anholts, Laura Blijleven, Tess Meuleman, Els van Beelen, Marie-Louise P. van der Hoorn, and Frans H. J. Claas. 2020. "Optimization of microRNA Acquirement from Seminal Plasma and Identification of Diminished Seminal microRNA-34b as Indicator of Low Semen Concentration" International Journal of Molecular Sciences 21, no. 11: 4089. https://doi.org/10.3390/ijms21114089
APA StyleEikmans, M., D. H. Anholts, J., Blijleven, L., Meuleman, T., van Beelen, E., van der Hoorn, M.-L. P., & Claas, F. H. J. (2020). Optimization of microRNA Acquirement from Seminal Plasma and Identification of Diminished Seminal microRNA-34b as Indicator of Low Semen Concentration. International Journal of Molecular Sciences, 21(11), 4089. https://doi.org/10.3390/ijms21114089