Comparative Proteomic Analysis of Nodulated and Non-Nodulated Casuarina glauca Sieb. ex Spreng. Grown under Salinity Conditions Using Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS)


 ,
,  ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Identification and Quantification of Casuarina glauca Proteins in Response to Salinity Stress
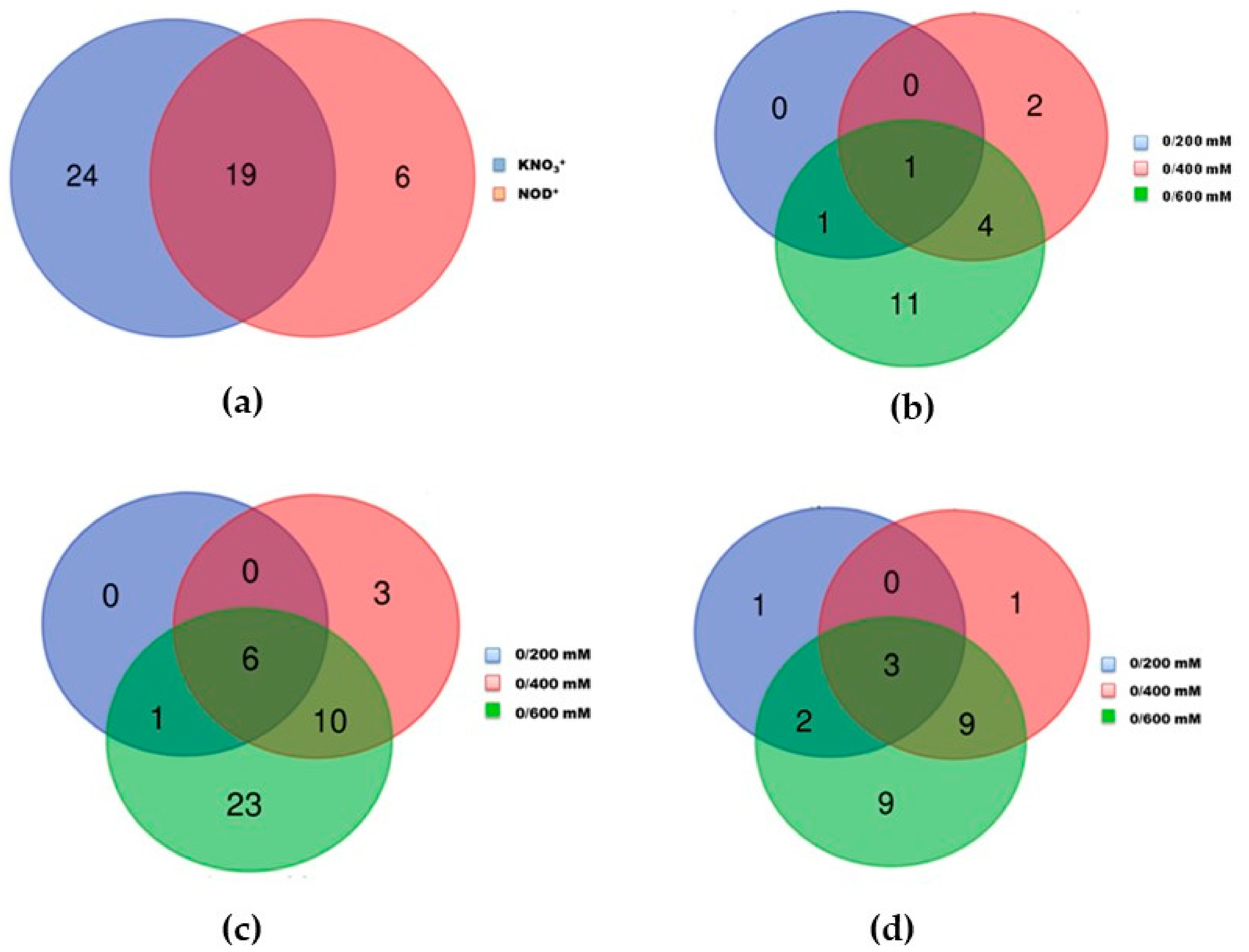
2.2. Differentially Expressed Proteins in Photosynthetic Branchlets of KNO3+ and NOD+ Plants
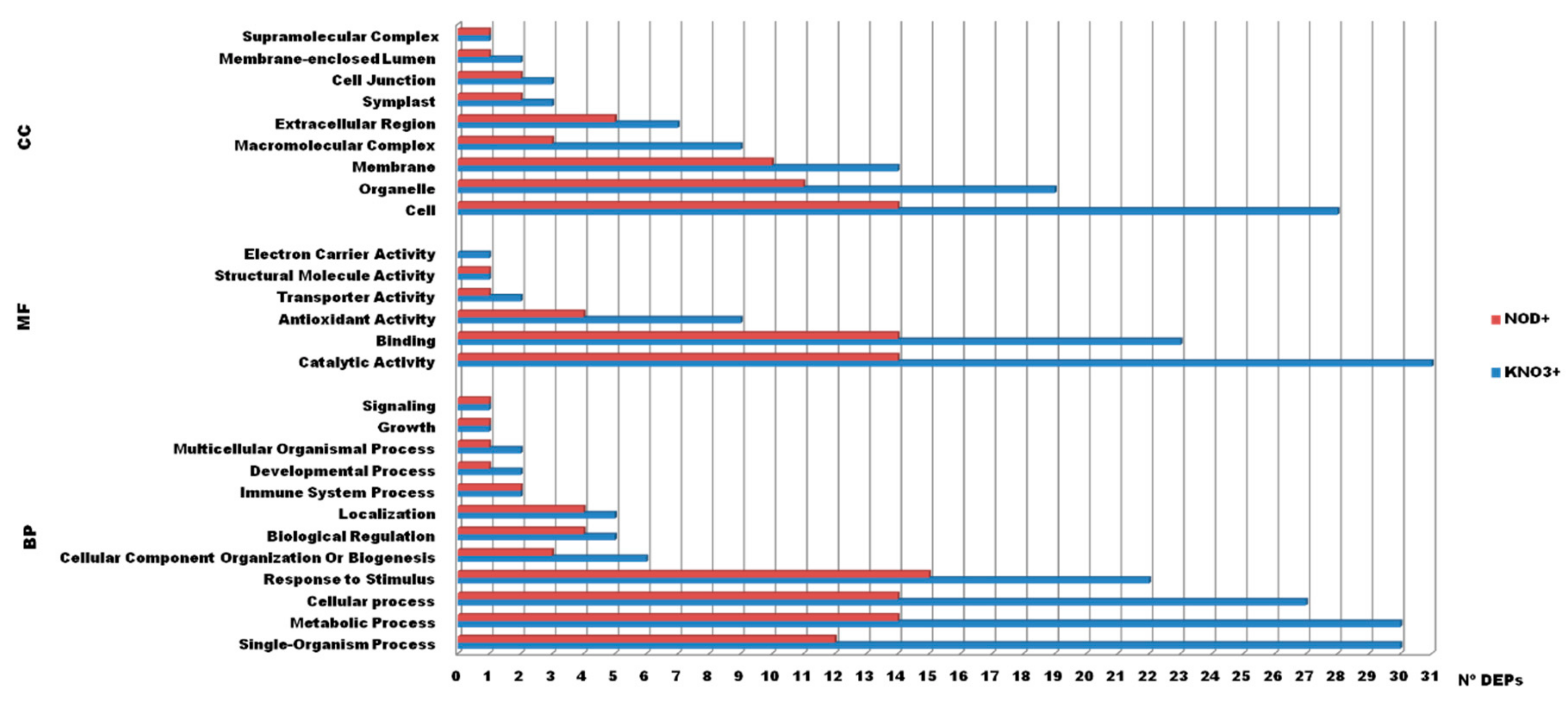
2.3. Functional Annotation and Classification of the Identified Proteins
2.4. Hierarchical Clustering Analysis
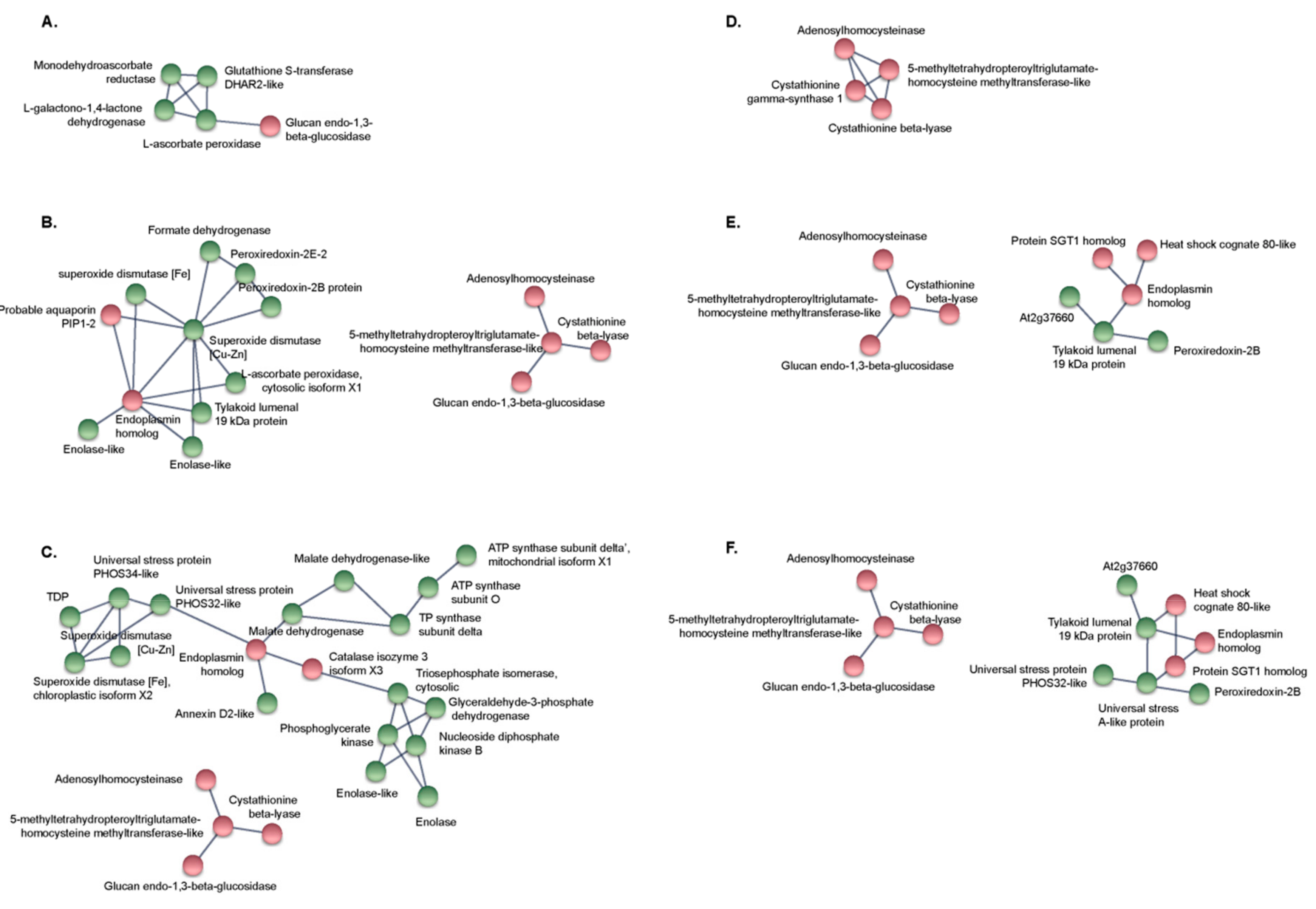
2.5. Protein-Protein Interaction Network
3. Discussion
4. Materials and Methods
4.1. Growing Conditions and Salt Treatment of Casuarina glauca
4.2. Protein Extraction and Quantification
4.3. Sample Preparation for Mass Spectrometry (MS) Analysis
4.4. Protein Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS)
4.4.1. SWATH Acquisition
4.4.2. Protein Identification/Library Generation
4.5. Statistical Analysis
4.6. Functional Annotation and Classification of Identified Proteins
4.7. Hierarchical Cluster Analysis of Protein Abundance and Interaction Network Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DEPs | Differentially Expressed Proteins |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| KNO3+ | Non-nodulated Casuarina glauca supplied with KNO3 |
| NOD+ | C. glauca nodulated by nitrogen-fixing Frankia Thr |
| Short-GeLC-MS/MS | Short Gel, Long Gradient Liquid Chromatography Tandem Mass Spectrometry |
| SWATH-MS | Sequential Window Acquisition of all Theoretical Mass Spectra |
References
- Diem, H.G.; Dommergues, Y.R. Current and potential uses and management of Casuarinaceae in tropics and subtropics. In The Biology of Frankia and Actinorhizal Plants; Schwintzer, C.R., Tjepkma, J.D., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 317–342. [Google Scholar]
- Santos, P.; Fortunato, A.; Graca, I.; Martins, S.; Gouveia, M.; Auguy, F.; Bogusz, D.; Ricardo, C.P.P.; Pawlowski, K.; Ribeiro, A. Characterization of four defense-related genes up-regulated in root nodules of Casuarina glauca. Symbiosis 2010, 50, 27–35. [Google Scholar] [CrossRef]
- Ribeiro, A.; Graça, I.; Pawlowski, K.; Santos, P. Actinorhizal plant defence-related genes in response to symbiotic Frankia. Funct. Plant Biol. 2011, 38, 639–644. [Google Scholar] [CrossRef]
- Batista-Santos, P.; Duro, N.; Rodrigues, A.P.; Semedo, J.N.; Alves, P.; da Costa, M.; Graça, I.; Pais, I.P.; Scotti-Campos, P.; Lidon, F.C.; et al. Is salt stress tolerance in Casuarina glauca Sieb. ex Spreng. associated with its nitrogen-fixing root-nodule symbiosis? An analysis at the photosynthetic level. Plant Physiol. Biochem. 2015, 96, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.R.; Abdel-Lateif, K.; Bogusz, D.; Franche, C. Influence of salt stress on inoculated Casuarina glauca seedlings. Symbiosis 2016, 70, 29–138. [Google Scholar] [CrossRef]
- Ribeiro-Barros, A.I.; da Costa, M.; Duro, N.; Graça, I.; Batista-Santos, P.; Jorge, T.F.; Lidon, F.C.; Pawlowski, K.; António, C.; Ramalho, J.C. An integrated approach to understand the mechanisms underlying salt stress tolerance in Casuarina glauca and its relation with nitrogen-fixing Frankia Thr. Symbiosis 2016, 70, 111–116. [Google Scholar] [CrossRef]
- Jorge, T.; Duro, N.; da Costa, M.; Florian, A.; Ramalho, J.C.; Ribeiro-Barros, A.; Fernie, A.; António, C. GC-TOF-MS analysis reveals salt stress-responsive metabolites in Casuarina glauca tissues. Metabolomics 2017, 13, 95. [Google Scholar] [CrossRef]
- Jorge, T.; Florêncio, H.; Ribeiro-Barros, A.I.; António, C. Quantification and structural characterization of raffinose family oligosaccharides in Casuarina glauca plant tissues by porous graphitic carbon electrospray quadrupole ion trap mass spectrometry. Int. J. Mass Spectrom. 2017, 413, 127–134. [Google Scholar] [CrossRef]
- Ngom, M.; Oshone, R.; Diagne, N.; Cissoko, M.; Svistoonoff, S.; Tisa, L.S.; Laplaze, L.; Sy, M.O.; Champion, A. Tolerance to environmental stress by the nitrogen-fixing actinobacterium Frankia and its role in actinorhizal plants adaptation. Symbiosis 2016, 70, 17–29. [Google Scholar] [CrossRef]
- Mansour, S.R.; Megahed, M.M. Interaction of soil and different Frankia strains on nodulation and mass production of three Casuarina species. Egypt. J. Microbiol. 2002, 37, 323–342. [Google Scholar]
- Carter, J.L.; Colmer, T.D.; Veneklaas, E.J. Variable tolerance of wetland tree species to combined salinity and waterlogging is related to regulation of ion uptake and production of organic solutes. New Phytol. 2006, 26, 123–134. [Google Scholar] [CrossRef]
- Duro, N.; Batista-Santos, P.; da Costa, M.; Maia, R.; Castro, I.V.; Ramos, M.; Ramalho, J.C.; Pawlowski, K.; Máguas, C.; Ribeiro-Barros, A.I. The impact of salinity on the symbiosis between Casuarina glauca Sieb. ex Spreng. and N2-fixing Frankia bacteria based on the analysis of Nitrogen and Carbon metabolism. Plant Soil 2016, 398, 327–337. [Google Scholar] [CrossRef]
- Duro, N.; da Costa, M.; Scotti-Campos, P.; Pais, I.P.; Colwell, F.; Rodrigues, A.P.; Semedo, J.N.; Lidon, F.C.; Ramalho, J.C.; Pawlowski, K.; et al. Salt stress tolerance in Casuarina glauca and its relation with nitrogen-fixing Frankia bacteria. In Biological Nitrogen Fixation and Beneficial Plant-Microbe Interactions; González-Andrés, F., James, E., Eds.; Springer: Cham, Switzerland, 2016; pp. 143–151. [Google Scholar]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Stepien, P.; Johnson, G.N. Contrasting responses of photosynthesis to salt stress in the glycophyte Arabidopsis and the halophyte Thellungiella: Role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Zhang, C.J.; Zhao, B.C.; Ge, W.N.; Zhang, Y.F.; Song, Y.; Sun, D.Y.; Guo, Y. An apoplastic h-type thioredoxin is involved in the stress response through regulation of the apoplastic reactive oxygen species in rice. Plant Physiol. 2011, 157, 1884–1899. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Scotti-Campos, P.; Duro, N.; da Costa, M.; Pais, I.P.; Rodrigues, A.P.; Batista-Santos, P.; Semedo, J.N.; Leitão, A.E.; Lidon, F.C.; Pawlowski, K.; et al. Antioxidative ability and membrane integrity in salt-induced responses of Casuarina glauca Sieber ex Spreng. in symbiosis with N2-fixing Frankia Thr or supplemented with mineral nitrogen. J. Plant Physiol. 2016, 196, 60–69. [Google Scholar] [CrossRef]
- Jorge, T.; Tohgeb, T.; Wendenburg, R.; Ramalho, J.C.; Lidon, F.C.; Ribeiro-Barros, A.I.; Fernie, A.R.; António, C. Salt-stress secondary metabolite signatures involved in the ability of Casuarina glauca to mitigate oxidative stress. Environ. Exp. Bot. 2019, 166, 103808. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Banu, N.A.; Hoque, A.; Watanabe-Sugimoto, M.; Matsuoka, K.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Proline and glycinebetaine induce antioxidant defense gene expression and suppress cell death in cultured tobacco cells under salt stress. J. Plant Physiol. 2009, 166, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Banu, M.N.; Hoque, M.A.; Watanable-Sugimoto, M.; Islam, M.M.; Uraji, M.; Matsuoka, K.; Nakamura, Y.; Murata, Y. Proline and glycinebetaine ameliorated NaCl stress via scavenging of hydrogen peroxide and methylglyoxal but not superoxide or nitric oxide in tobacco cultured cells. Biosci. Biotechnol. Biochem. 2010, 74, 2043–2049. [Google Scholar] [CrossRef] [PubMed]
- Ngom, M.; Gray, K.; Diagne, N.; Oshone, R.; Fardoux, J.; Gherbi, H.; Hocher, V.; Svistoonoff, S.; Laplaze, L.; Tisa, L.S.; et al. Symbiotic performance of diverse Frankia strains on salt-stressed Casuarina glauca and Casuarina equisetifolia plants. Front. Plant Sci. 2016, 7, 1331. [Google Scholar] [CrossRef] [PubMed]
- Balbuenam, T.S.; Dias, L.L.C.; Martins, M.L.B.; Chiquieri, T.B.; Santa-Catarina, C.; Floh, E.I.S.; Silveira, V. Challenges in proteome analyses of tropical plants. Braz. J. Plant Physiol. 2011, 23, 91–104. [Google Scholar] [CrossRef]
- Wasinger, V.C.; Cordwell, S.J.; Cerpa-Poljak, A.; Yan, J.X.; Gooley, A.A.; Wilkins, M.R.; Duncan, M.W.; Harris, R.; Williams, K.L.; Humphery-Smith, I. Progress with gene-product mapping of the Mollicutes: Mycoplasma genitalium. Electrophoresis 1995, 16, 1090–1094. [Google Scholar] [CrossRef]
- Cappadona, S.; Baker, P.R.; Cutillas, P.R.; Heck, A.J.R.; van Breukele, B. Current challenges in software solutions for mass spectrometry-based quantitative proteomics. Amino Acids 2012, 43, 1087–1108. [Google Scholar] [CrossRef]
- Smith, R.; Mathis, A.D.; Ventura, D.; Prince, J.T. Proteomics, lipidomics, metabolomics: A mass spectrometry tutorial from a computer scientist’s point of view. BMC Bioinform. 2014, 15, S9. [Google Scholar] [CrossRef]
- Anjo, S.I.; Santa, C.; Manadas, B. Short GeLC-SWATH: A fast and reliable quantitative approach for proteomic screenings. Proteomics 2015, 15, 757–762. [Google Scholar] [CrossRef]
- Gillet, L.C.; Navarro, P.; Tate, S.; Rost, H.; Selevsek, N.; Reiter, L.; Bonner, R.; Aebersold, R. Targeted data extraction of the MS/MS spectra generated by data independent acquisition: A new concept for consistent and accurate proteome analysis. Mol. Cell Proteom. 2012, 11, O111.016717 . [Google Scholar] [CrossRef]
- Liu, Y.; Huttenhain, R.; Surinova, S.; Gillet, L.C.; Mouritsen, J.; Brunner, R.; Navarro, P.; Aebersold, R. Quantitative measurements of N-linked glycoproteins in human plasma by SWATH-MS. Proteomics 2013, 13, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.S.; Chen, J.; Sethi, A.; Li, Q.K.; Chen, L.J.; Collins, B.; Gillet, L.C.; Wollscheid, B.; Zhang, H.; Aebersold, R. Glycoproteomic analysis of prostate cancer tissues by SWATH mass spectrometry discovers N-acylethanolamine acid amidase and protein tyrosine kinase 7 as signatures for tumor aggressiveness. Mol. Cell Proteom. 2014, 13, 1753–1768. [Google Scholar] [CrossRef] [PubMed]
- Haverland, N.A.; Fox, H.S.; Ciborowski, P. Quantitative proteomics by SWATH-MS reveals altered expression of nucleic acid binding and regulatory proteins in HIV-1-infected macrophages. J. Proteome Res. 2014, 13, 2109–2119. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Shilov, I.V.; Seymour, S.L. Non-linear fitting method for determining local false discovery rates from decoy database searches. J. Proteome Res. 2008, 7, 3661–3667. [Google Scholar] [CrossRef] [PubMed]
- Sennels, L.; Bukowski-Wills, J.C.; Rappsilber, J. Improved results in proteomics by use of local and peptide-class specific false discovery rates. BMC Bioinform. 2009, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, A.; Santos, P.; Graça, I.; Gouveia, M.; Martins, S.; Ricardo, C.P.; Pawlowski, K.; Ribeiro, A. Isolation and characterization of cgchi3, a nodule-specific gene from Casuarina glauca encoding a class III chitinase. Physiol. Plant 2007, 130, 418–426. [Google Scholar] [CrossRef]
- Pinheiro, C.; Guerra-Guimarães, L.; David, T.S.; Vieira, A. Proteomics: State of the art to study Mediterranean woody species under stress. Environ. Exp. Bot. 2014, 103, 117–127. [Google Scholar] [CrossRef]
- Santos, P.; Fortunato, A.; Ribeiro, A.; Pawlowski, K. Chitinases in root nodules. Plant Biotechnol. 2008, 25, 299–307. [Google Scholar] [CrossRef]
- Graça, I.; Liang, J.; Guilherme, M.; Tavares, P.; Ferreira-Pinto, M.; Melo, A.; Ribeiro-Barros, A.I.; Pereira, A. Cloning, overexpression and functional characterization of a class III chitinase from Casuarina glauca nodules. Symbiosis 2016, 70, 139–148. [Google Scholar] [CrossRef]
- Graça, I.; Guilherme, M.; Tavares, P.; Pereira, A.S.; Ribeiro-Barros, A.I. Functional characterization of a chitinase class III (CgCHI3) and a glutathione S-transferase (CgGST) involved in Casuarina glauca-Frankia symbiosis. In Biological Nitrogen Fixation and Beneficial Plant-Microbe Interactions; González-Andrés, F., James, E., Eds.; Springer: Cham, Switzerland, 2016; pp. 97–107. [Google Scholar]
- Drechsler, N.; Zheng, Y.; Bohner, A.; Nobmann, B.; von Wirén, N.; Kunze, R.; Rausch, C. Nitrate-dependent control of shoot K+ homeostasis by the nitrate transporter1/peptide transporter family member NPF7.3/NRT1.5 and the stelar K+ outward rectifier SKOR in Arabidopsis. Plant Physiol. 2015, 169, 2832–2847. [Google Scholar]
- Yashvardhini, N.; Bhattacharya, S.; Chaudhuri, S.; Sengupta, D.N. Molecular characterization of the 14-3-3 gene family in rice and its expression studies under abiotic stress. Planta 2018, 247, 229–253. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Jin, X.; Sun, X.; Gao, T.; Chen, X.; She, Y.; Jiang, T.; Chen, S.; Dai, S. Hydrogen peroxide response in leaves of poplar (Populus simonii × Populus nigra) revealed from physiological and proteomic analyses. Int. J. Mol. Sci. 2018, 18, 2085. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Ko, K.; Chang, W.L.; Kuo, W.C.; Chen, G.H.; Lin, T.P. Increased glutathione contributes to stress tolerance and global translational changes in Arabidopsis. Plant J. 2015, 83, 926–939. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, M.; Xia, Z. The SULTR gene family in maize (Zea mays L.): Gene cloning and expression analyses under sulfate starvation and abiotic stress. J. Plant Physiol. 2018, 220, 24–33. [Google Scholar] [CrossRef]
- Müller, S.M.; Wang, S.; Telman, W.; Liebthal, M.; Schnitzer, H.; Viehhauser, A.; Sticht, C.; Delatorre, C.; Wirtz, M.; Hell, R.; et al. The redox-sensitive module of cyclophilin 20-3, 2-cysteine peroxiredoxin and cysteine synthase integrates sulfur metabolism and oxylipin signaling in the high light acclimation response. Plant J. 2017, 91, 995–1014. [Google Scholar] [CrossRef]
- Wasternack, C.; Feussner, I. The oxylipin pathways: Biochemistry and function. Annu. Rev. Plant Biol. 2018, 69, 363–386. [Google Scholar] [CrossRef]
- Curien, G.; Giustini, C.; Montillet, J.L.; Mas-Y-Mas, S.; Cobessi, D.; Ferrer, J.L.; Matringe, M.; Grechkin, A.; Rolland, N. The chloroplast membrane associated ceQORH putative quinone oxidoreductase reduces long-chain, stress-related oxidized lipids. Phytochemistry 2016, 122, 45–55. [Google Scholar] [CrossRef]
- Nohzadeh Malakshah, S.; Habibi Rezaei, M.; Heidari, M.; Salekdeh, G.H. Proteomics reveals new salt responsive proteins associated with rice plasma membrane. Biosci. Biotechnol. Biochem. 2007, 71, 2144–2154. [Google Scholar] [CrossRef]
- Wang, J.; Meng, Y.; Li, B.; Ma, X.; Lai, Y.; Si, E.; Yang, K.; Xu, X.; Shang, X.; Wang, H.; et al. Physiological and proteomic analyses of salt stress response in the halophyte Halogeton glomeratus. Plant Cell Environ. 2015, 38, 655–666. [Google Scholar] [CrossRef]
- Zhang, H.; Ni, Z.; Chen, Q.; Guo, Z.; Gao, W.; Su, X.; Qu, Y. Proteomic responses of drought-tolerant and drought-sensitive cotton varieties to drought stress. Mol. Genet. Genom. 2016, 291, 1293–1303. [Google Scholar] [CrossRef]
- Alekseeva, A.A.; Savin, S.S.; Tishkov, V.I. NAD+-dependent formate dehydrogenase from plants. Acta Nat. 2011, 3, 38–54. [Google Scholar] [CrossRef]
- Liu, J.J.; Zamani, A.; Ekramoddoullah, A.K. Expression profiling of a complex thaumatin-like protein family in western white pine. Planta 2010, 231, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Charron, J.B.; Ouellet, F.; Pelletier, M.; Danyluk, J.; Chauve, C.; Sarhan, F. Identification, expression, and evolutionary analyses of plant lipocalins. Plant Physiol. 2005, 139, 2017–2028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, Y.; Xiao, H.; Zheng, Y.; Yue, B. Genome-wide identification, evolution, and expression analysis of RNA-binding glycine-rich protein family in maize. J. Integr. Plant Biol. 2014, 56, 1020–1031. [Google Scholar] [CrossRef]
- Konopka-Postupolska, D.; Clark, D. Annexins as overlooked regulators of membrane trafficking in plant cells. Int. J. Mol. Sci. 2017, 18, 863. [Google Scholar] [CrossRef]
- Takahashi, H.; Wang, S.; Hayashi, S.; Wakasa, Y.; Takaiwa, F. Cis-element of the rice PDIL2-3 promoter is responsible for inducing the endoplasmic reticulum stress response. J. Biosci. Bioeng. 2014, 117, 620–623. [Google Scholar] [CrossRef]
- Kayum, M.A.; Park, J.I.; Nath, U.K.; Saha, G.; Biswas, M.K.; Kim, H.T.; Nou, I.S. Genome-wide characterization and expression profiling of PDI family gene reveals function as abiotic and biotic stress tolerance in Chinese cabbage (Brassica rapa ssp. pekinensis). BMC Genom. 2017, 18, 885. [Google Scholar] [CrossRef]
- Damaris, R.N.; Li, M.; Liu, Y.; Chen, X.; Murage, H.; Yang, P. A proteomic analysis of salt stress response in seedlings of two African rice cultivars. Biochim. Biophys. Acta 2016, 1864, 1570–1578. [Google Scholar] [CrossRef]
- Zhou, S.; Chen, Q.; Sun, Y.; Li, Y. Histone H2B monoubiquitination regulates salt stress-induced microtubule depolymerization in Arabidopsis. Plant Cell Environ. 2017, 40, 1512–1530. [Google Scholar] [CrossRef]
- Hwang, D.H.; Kim, S.T.; Kim, S.G.; Kang, K.Y. Comprehensive analysis of the expression of twenty-seven beta-1, 3-glucanase genes in rice (Oryza sativa L.). Mol. Cells 2007, 23, 207–214. [Google Scholar]
- Su, Y.; Wang, Z.; Liu, F.; Li, Z.; Peng, Q.; Guo, J.; Xu, L.; Que, Y. Isolation and characterization of ScGluD2, a new sugarcane beta-1,3-glucanase D family gene induced by Sporisorium scitamineum, ABA, H2O2, NaCl, and CdCl2 Stresses. Front. Plant Sci. 2016, 7, 1348. [Google Scholar] [CrossRef] [PubMed]
- Azri, W.; Barhoumi, Z.; Chibani, F.; Borji, M.; Bessrour, M.; Mliki, A. Proteomic responses in shoots of the facultative halophyte Aeluropus littoralis (Poaceae) under NaCl salt stress. Funct. Plant Biol. 2016, 43, 1028–1104. [Google Scholar] [CrossRef]
- Qiao, G.; Zhang, X.; Jiang, J.; Liu, M.; Han, X.; Yang, H.; Renying, Z. Comparative proteomic analysis of responses to salt stress in Chinese willow (Salix matsudana Koidz). Plant Mol. Biol. Rep. 2014, 32, 814–827. [Google Scholar] [CrossRef]
- Ramalho, J.C.; DaMatta, F.M.; Rodrigues, A.P.; Scotti-Campos, P.; Pais, I.; Batista-Santos, P.; Partelli, F.L.; Ribeiro, A.; Lidon, F.C.; Leitão, A.E. Cold impact and acclimation response of Coffea spp. plants. Theor. Exp. Plant Phys. 2014, 26, 5–18. [Google Scholar] [CrossRef]
- Girgis, M.G.Z.; Ishac, Y.Z.; El-Haddad, M.; Saleh, E.A.; Diem, H.G.; Dommergues, Y.R. First report on isolation and culture of effective Casuarina-compatible strains of Frankia from Egypt. In Advances in Casuarina Research and Utilization, Proceedings of the Second International Casuarina Workshop, Cairo, Egypt, 15–20 January 1990; El-Lakany, M.H., Turnbull, J.W., Brewbaker, J.L., Eds.; American University: Cairo, Egypt, 1990; pp. 156–164. [Google Scholar]
- Bertolde, F.Z.; Almeida, A.A.; Pirovani, C.P. Analysis of gene expression and proteomic profiles of clonal genotypes from Theobroma cacao subjected to soil flooding. PLoS ONE 2014, 9, e108705. [Google Scholar] [CrossRef] [PubMed]
- Manadas, B.; Santos, A.R.; Szabadfi, K.; Gomes, J.R.; Garbis, S.D.; Fountoulakis, M.; Duarte, C.B. BDNF-induced changes in the expression of the translation machinery in hippocampal neurons: Protein levels and dendritic mRNA. J. Proteome Res. 2009, 8, 4536–4552. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2go: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S. Blast2GO: A comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genom. 2008, 2008, 619832. [Google Scholar] [CrossRef]
- Caraux, G.; Pinloche, S. PermutMatrix: A graphical environment to arrange gene expression profiles in optimal linear order. Bioinformatics 2005, 21, 1280–1281. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graça, I.; Mendes, V.M.; Marques, I.; Duro, N.; da Costa, M.; Ramalho, J.C.; Pawlowski, K.; Manadas, B.; Pinto Ricardo, C.P.; Ribeiro-Barros, A.I. Comparative Proteomic Analysis of Nodulated and Non-Nodulated Casuarina glauca Sieb. ex Spreng. Grown under Salinity Conditions Using Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS). Int. J. Mol. Sci. 2020, 21, 78. https://doi.org/10.3390/ijms21010078
Graça I, Mendes VM, Marques I, Duro N, da Costa M, Ramalho JC, Pawlowski K, Manadas B, Pinto Ricardo CP, Ribeiro-Barros AI. Comparative Proteomic Analysis of Nodulated and Non-Nodulated Casuarina glauca Sieb. ex Spreng. Grown under Salinity Conditions Using Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS). International Journal of Molecular Sciences. 2020; 21(1):78. https://doi.org/10.3390/ijms21010078
Chicago/Turabian StyleGraça, Inês, Vera M. Mendes, Isabel Marques, Nuno Duro, Mário da Costa, José C. Ramalho, Katharina Pawlowski, Bruno Manadas, Cândido P. Pinto Ricardo, and Ana I. Ribeiro-Barros. 2020. "Comparative Proteomic Analysis of Nodulated and Non-Nodulated Casuarina glauca Sieb. ex Spreng. Grown under Salinity Conditions Using Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS)" International Journal of Molecular Sciences 21, no. 1: 78. https://doi.org/10.3390/ijms21010078
APA StyleGraça, I., Mendes, V. M., Marques, I., Duro, N., da Costa, M., Ramalho, J. C., Pawlowski, K., Manadas, B., Pinto Ricardo, C. P., & Ribeiro-Barros, A. I. (2020). Comparative Proteomic Analysis of Nodulated and Non-Nodulated Casuarina glauca Sieb. ex Spreng. Grown under Salinity Conditions Using Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS). International Journal of Molecular Sciences, 21(1), 78. https://doi.org/10.3390/ijms21010078