The Microarchitecture of Pancreatic Cancer as Measured by Diffusion-Weighted Magnetic Resonance Imaging Is Altered by T Cells with a Tumor Promoting Th17 Phenotype

 ,
, 
Abstract
1. Introduction
2. Results
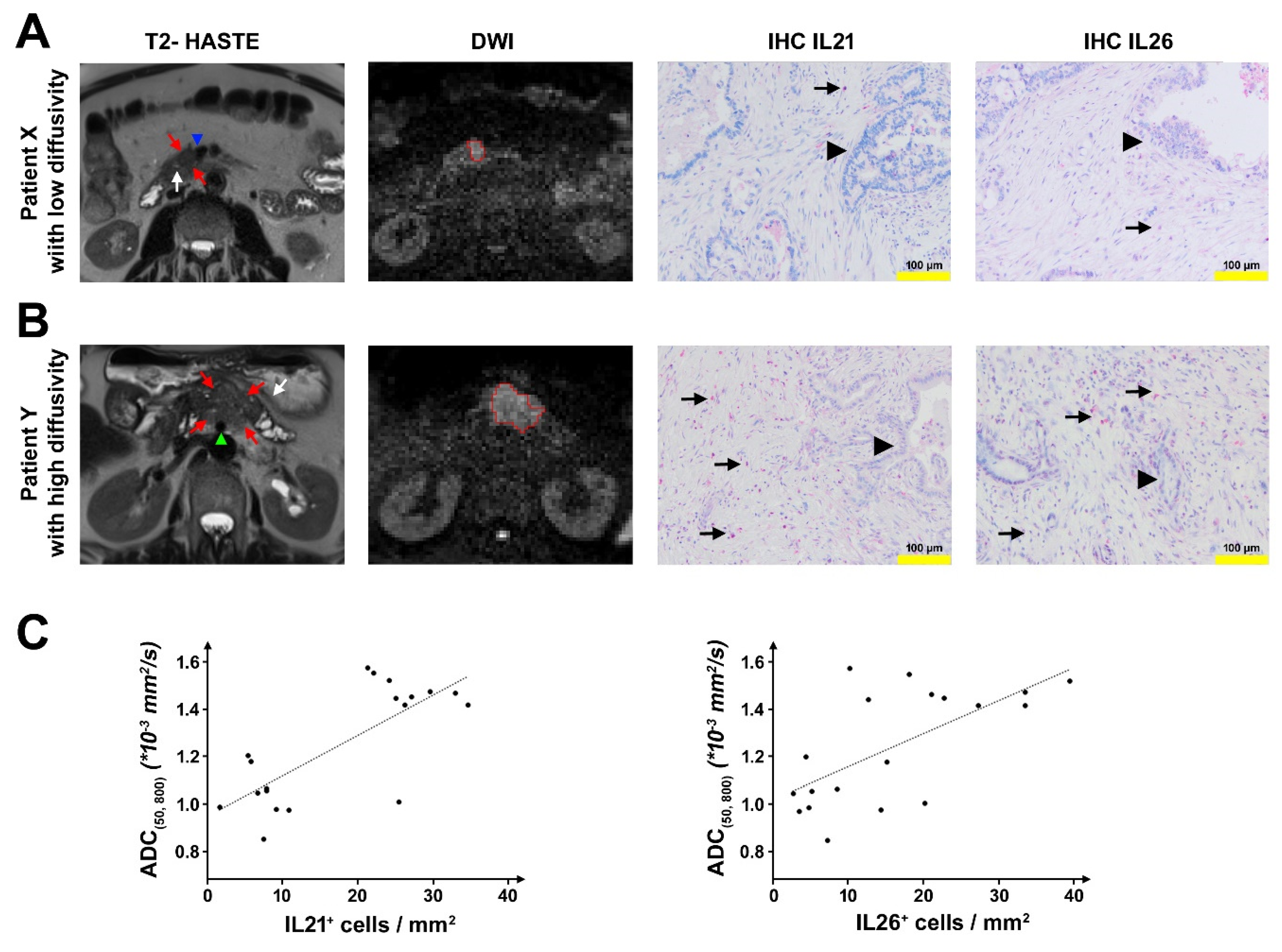
2.1. DW-MRI in Patients with PDAC
2.2. Analysis of Tumor-Infiltrating T Cells in PDAC
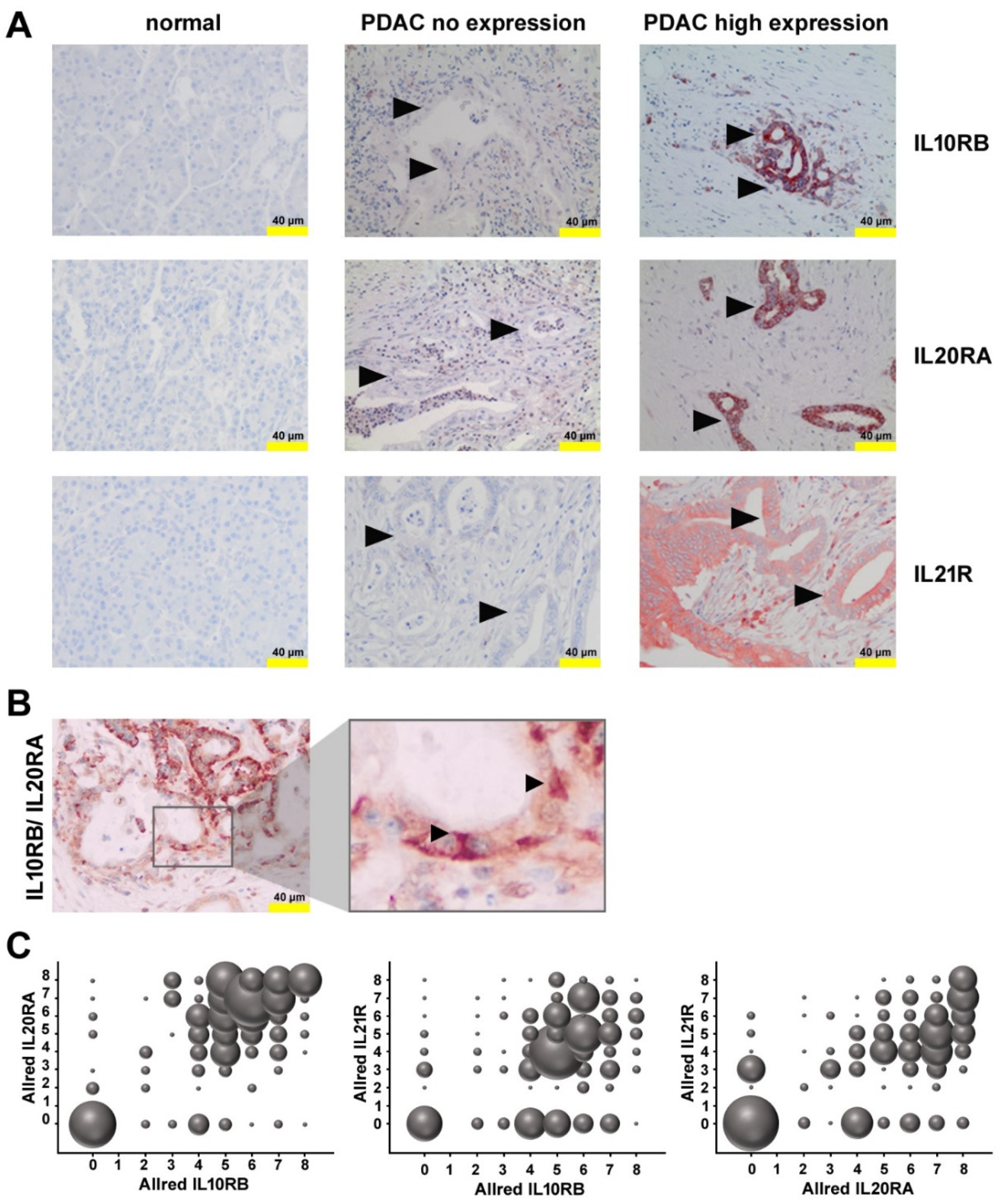
2.3. Expression of Receptors for IL21 and IL26 in PDAC
2.4. Expression of Functional IL10RB and IL20RA on Pancreatic Tumor Cells Lines
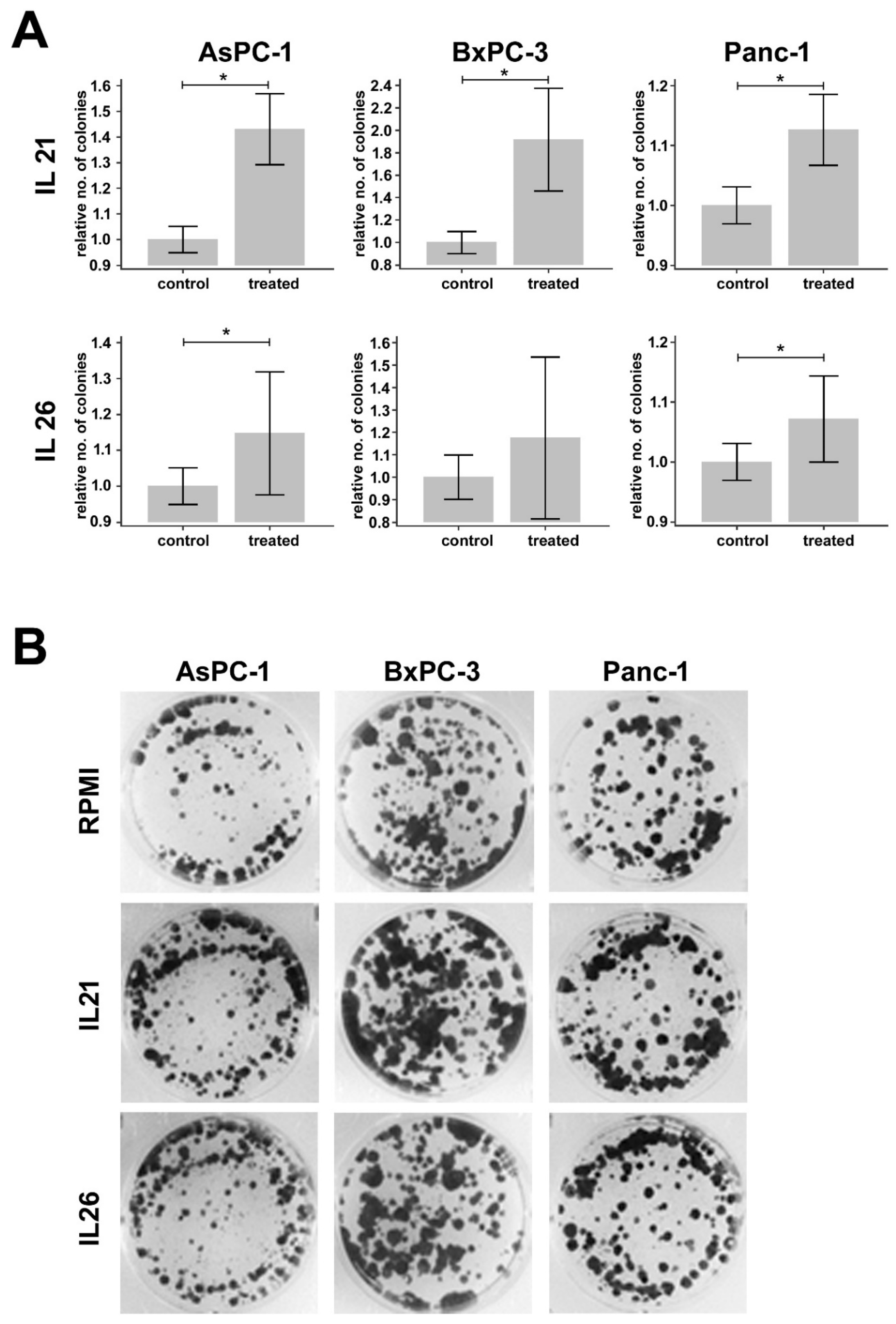
2.5. Effect of IL21 and IL26 on Colony Formation
3. Discussion
4. Materials and Methods
4.1. DW-MRI
4.2. Patient Samples and Immunohistochemistry
4.3. Pancreatic Carcinoma Cell Lines and Cell Culture
4.4. Western Blotting
4.5. Immunofluorescence Staining of IL10RB, IL20RA, and IL21R
4.6. Colony Formation Assay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADC | Apparent diffusion coefficient |
| BCA | Bicinchoninic acid |
| BSA | Bovine serum albumin |
| CD | Cluster of differentiation |
| CT | Computed tomography |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DW | Diffusion-weighted |
| DWI | Diffusion-weighted imaging |
| ERK | Extracellular signal-related kinase |
| FOLFIRINOX | Combination therapy containing folinic acid, fluorouracil, irinotecan, and oxaliplatin |
| GRAPPA | Generalized autocalibrating partial parallel acquisition |
| HASTE | Half-Fourier acquisition single-shot turbo spin echo |
| HE | Hematoxylin-eosin |
| ICC | Intraclass correlation coefficient |
| IHC | Immunohistochemistry |
| IL | Interleukin |
| JAK | Janus Kinase |
| MITK | Medical Imaging Interaction Toolkit |
| MRI | Magnetic resonance imaging |
| NCT | National Center for Tumor Diseases |
| OS | Overall survival |
| PBS | Phosphate-buffered saline |
| PDAC | Pancreatic ductal adenocarcinoma |
| PFA | Paraformaldehyde |
| RIPA | Radioimmunoprecipitation assay |
| RIS | Radiological information system |
| rs | Spearman rank correlation coefficient |
| RT | Room temperature |
| S | Signal magnitude |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SE2d-EPI | Single-shot two-dimensional echo-planar imaging |
| STAT | Signal transducer and activator of transcription |
| T | Tesla |
| TNF | Tumor necrosis factor |
| VOI | Volume of interest |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA. Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef]
- Coussens, L.M.; Zitvogel, L.; Palucka, A.K. Neutralizing tumor-promoting chronic inflammation: a magic bullet? Science 2013, 339, 286–291. [Google Scholar] [CrossRef]
- Kleeff, J.; Beckhove, P.; Esposito, I.; Herzig, S.; Huber, P.E.; Löhr, J.M.; Friess, H. Pancreatic cancer microenvironment. Int. J. Cancer 2007, 121, 699–705. [Google Scholar] [CrossRef]
- Wente, M.N.; Gaida, M.M.; Mayer, C.; Michalski, C.W.; Haag, N.; Giese, T.; Felix, K.; Bergmann, F.; Giese, N.A.; Friess, H. Expression and potential function of the CXC chemokine CXCL16 in pancreatic ductal adenocarcinoma. Int. J. Oncol. 2008, 33, 297–308. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Swayden, M.; Iovanna, J.; Soubeyran, P. Pancreatic cancer chemo-resistance is driven by tumor phenotype rather than tumor genotype. Heliyon 2018, 4, e01055. [Google Scholar] [CrossRef] [PubMed]
- Sapalidis, K.; Kosmidis, C.; Funtanidou, V.; Katsaounis, A.; Barmpas, A.; Koimtzis, G.; Mantalobas, S.; Alexandrou, V.; Aidoni, Z.; Koulouris, C.; et al. Update on current pancreatic treatments: from molecular pathways to treatment. J. Cancer 2019, 10, 5162–5172. [Google Scholar] [CrossRef] [PubMed]
- Falzone, L.; Salomone, S.; Libra, M. Evolution of Cancer Pharmacological Treatments at the Turn of the Third Millennium. Front. Pharmacol. 2018, 9, 1300. [Google Scholar] [CrossRef]
- Alam, M.S.; Gaida, M.M.; Bergmann, F.; Lasitschka, F.; Giese, T.; Giese, N.A.; Hackert, T.; Hinz, U.; Hussain, S.P.; Kozlov, S.V.; et al. Selective inhibition of the p38 alternative activation pathway in infiltrating T cells inhibits pancreatic cancer progression. Nat. Med. 2015, 21, 1337–1343. [Google Scholar] [CrossRef]
- McAllister, F.; Bailey, J.M.; Alsina, J.; Nirschl, C.J.; Sharma, R.; Fan, H.; Rattigan, Y.; Roeser, J.C.; Lankapalli, R.H.; Zhang, H.; et al. Oncogenic Kras activates a hematopoietic-to-epithelial IL-17 signaling axis in preinvasive pancreatic neoplasia. Cancer Cell 2014, 25, 621–637. [Google Scholar] [CrossRef]
- Linnebacher, A.; Mayer, P.; Marnet, N.; Bergmann, F.; Herpel, E.; Revia, S.; Yin, L.; Liu, L.; Hackert, T.; Giese, T.; et al. Interleukin 21 Receptor/Ligand Interaction Is Linked to Disease Progression in Pancreatic Cancer. Cells 2019, 8, 1104. [Google Scholar] [CrossRef]
- He, W.; Wu, J.; Shi, J.; Huo, Y.-M.; Dai, W.; Geng, J.; Lu, P.; Yang, M.-W.; Fang, Y.; Wang, W.; et al. IL22RA1/STAT3 Signaling Promotes Stemness and Tumorigenicity in Pancreatic Cancer. Cancer Res. 2018, 78, 3293–3305. [Google Scholar] [PubMed]
- Rao, C.V.; Mohammed, A.; Janakiram, N.B.; Li, Q.; Ritchie, R.L.; Lightfoot, S.; Vibhudutta, A.; Steele, V.E. Inhibition of pancreatic intraepithelial neoplasia progression to carcinoma by nitric oxide-releasing aspirin in p48(Cre/+)-LSL-Kras(G12D/+) mice. Neoplasia N. Y. 2012, 14, 778–787. [Google Scholar] [CrossRef] [PubMed]
- Mayorek, N.; Naftali-Shani, N.; Grunewald, M. Diclofenac inhibits tumor growth in a murine model of pancreatic cancer by modulation of VEGF levels and arginase activity. PLoS ONE 2010, 5, e12715. [Google Scholar] [CrossRef] [PubMed]
- Türkbey, B.; Aras, Ö.; Karabulut, N.; Turgut, A.T.; Akpinar, E.; Alibek, S.; Pang, Y.; Ertürk, Ş.M.; El Khouli, R.H.; Bluemke, D.A.; et al. Diffusion-weighted MRI for detecting and monitoring cancer: a review of current applications in body imaging. Diagn. Interv. Radiol. Ank. Turk. 2012, 18, 46–59. [Google Scholar] [CrossRef]
- Fukukura, Y.; Takumi, K.; Kamimura, K.; Shindo, T.; Kumagae, Y.; Tateyama, A.; Nakajo, M. Pancreatic adenocarcinoma: variability of diffusion-weighted MR imaging findings. Radiology 2012, 263, 732–740. [Google Scholar] [CrossRef]
- Klauss, M.; Gaida, M.M.; Lemke, A.; Grünberg, K.; Simon, D.; Wente, M.N.; Delorme, S.; Kauczor, H.-U.; Grenacher, L.; Stieltjes, B. Fibrosis and pancreatic lesions: counterintuitive behavior of the diffusion imaging-derived structural diffusion coefficient d. Investig. Radiol. 2013, 48, 129–133. [Google Scholar]
- Muraoka, N.; Uematsu, H.; Kimura, H.; Imamura, Y.; Fujiwara, Y.; Murakami, M.; Yamaguchi, A.; Itoh, H. Apparent diffusion coefficient in pancreatic cancer: characterization and histopathological correlations. J. Magn. Reson. Imaging JMRI 2008, 27, 1302–1308. [Google Scholar] [CrossRef]
- Heid, I.; Steiger, K.; Trajkovic-Arsic, M.; Settles, M.; Eßwein, M.R.; Erkan, M.; Kleeff, J.; Jäger, C.; Friess, H.; Haller, B.; et al. Co-clinical Assessment of Tumor Cellularity in Pancreatic Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 1461–1470. [Google Scholar] [CrossRef]
- Barral, M.; Taouli, B.; Guiu, B.; Koh, D.-M.; Luciani, A.; Manfredi, R.; Vilgrain, V.; Hoeffel, C.; Kanematsu, M.; Soyer, P. Diffusion-weighted MR imaging of the pancreas: current status and recommendations. Radiology 2015, 274, 45–63. [Google Scholar] [CrossRef]
- Mayer, P.; Dinkic, C.; Jesenofsky, R.; Klauss, M.; Schirmacher, P.; Dapunt, U.; Hackert, T.; Uhle, F.; Hänsch, G.M.; Gaida, M.M. Changes in the microarchitecture of the pancreatic cancer stroma are linked to neutrophil-dependent reprogramming of stellate cells and reflected by diffusion-weighted magnetic resonance imaging. Theranostics 2018, 8, 13–30. [Google Scholar] [CrossRef]
- Wiggermann, P.; Grützmann, R.; Weissenböck, A.; Kamusella, P.; Dittert, D.-D.; Stroszczynski, C. Apparent diffusion coefficient measurements of the pancreas, pancreas carcinoma, and mass-forming focal pancreatitis. Acta Radiol. Stockh. Swed. 1987 2012, 53, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Lemke, A.; Laun, F.B.; Klauss, M.; Re, T.J.; Simon, D.; Delorme, S.; Schad, L.R.; Stieltjes, B. Differentiation of pancreas carcinoma from healthy pancreatic tissue using multiple b-values: comparison of apparent diffusion coefficient and intravoxel incoherent motion derived parameters. Invest. Radiol. 2009, 44, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Barilla, R.M.; Diskin, B.; Caso, R.C.; Lee, K.B.; Mohan, N.; Buttar, C.; Adam, S.; Sekendiz, Z.; Wang, J.; Salas, R.D.; et al. Specialized dendritic cells induce tumor-promoting IL-10+IL-17+ FoxP3neg regulatory CD4+ T cells in pancreatic carcinoma. Nat. Commun. 2019, 10, 1424. [Google Scholar] [CrossRef] [PubMed]
- Tengvall, S.; Che, K.F.; Lindén, A. Interleukin-26: An Emerging Player in Host Defense and Inflammation. J. Innate Immun. 2016, 8, 15–22. [Google Scholar] [CrossRef]
- Hör, S.; Pirzer, H.; Dumoutier, L.; Bauer, F.; Wittmann, S.; Sticht, H.; Renauld, J.-C.; de Waal Malefyt, R.; Fickenscher, H. The T-cell lymphokine interleukin-26 targets epithelial cells through the interleukin-20 receptor 1 and interleukin-10 receptor 2 chains. J. Biol. Chem. 2004, 279, 33343–33351. [Google Scholar] [CrossRef]
- You, W.; Tang, Q.; Zhang, C.; Wu, J.; Gu, C.; Wu, Z.; Li, X. IL-26 promotes the proliferation and survival of human gastric cancer cells by regulating the balance of STAT1 and STAT3 activation. PLoS ONE 2013, 8, e63588. [Google Scholar] [CrossRef]
- Mürtz, P.; Sprinkart, A.M.; Reick, M.; Pieper, C.C.; Schievelkamp, A.-H.; König, R.; Schild, H.H.; Willinek, W.A.; Kukuk, G.M. Accurate IVIM model-based liver lesion characterisation can be achieved with only three b-value DWI. Eur. Radiol. 2018, 28, 4418–4428. [Google Scholar] [CrossRef]
- Allred, D.C.; Clark, G.M.; Elledge, R.; Fuqua, S.A.; Brown, R.W.; Chamness, G.C.; Osborne, C.K.; McGuire, W.L. Association of p53 protein expression with tumor cell proliferation rate and clinical outcome in node-negative breast cancer. J. Natl. Cancer Inst. 1993, 85, 200–206. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort 1 (n = 19) | Cohort 2 (n = 199) | |
|---|---|---|
| Sex | ||
| Female | 12 | 98 |
| Male | 7 | 101 |
| Mean Age ± SD [years] | 66.2 ± 8.1 | 64.1 ± 9.8 |
| Location of primary tumor | ||
| Pancreatic head | 14 | 192 |
| Pancreatic body and/ or tail | 5 | 7 |
| Type of surgery | ||
| Pylorus-preserving Whipple | 6 | 167 |
| Classical Whipple | 4 | 21 |
| Left pancreatic resection | 3 | 5 |
| Total pancreatectomy | 6 | 6 |
| Extent of primary tumor (T) | ||
| T1 | 0 | 23 |
| T2 | 0 | 155 |
| T3 | 19 | 21 |
| T4 | 0 | 0 |
| Lymph node metastases (N) | ||
| N0 | 5 | 32 |
| N1 | 4 | 70 |
| N2 | 10 | 97 |
| Distant metastases (M) | ||
| M0/Mx | 19 | 190 |
| M1 | 0 | 9 |
| Tumor Grading (G) | ||
| G1 | 0 | 5 |
| G2 | 11 | 126 |
| G3 | 8 | 68 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayer, P.; Linnebacher, A.; Glennemeier-Marke, H.; Marnet, N.; Bergmann, F.; Hackert, T.; Klauss, M.; Poth, T.; Gaida, M.M. The Microarchitecture of Pancreatic Cancer as Measured by Diffusion-Weighted Magnetic Resonance Imaging Is Altered by T Cells with a Tumor Promoting Th17 Phenotype. Int. J. Mol. Sci. 2020, 21, 346. https://doi.org/10.3390/ijms21010346
Mayer P, Linnebacher A, Glennemeier-Marke H, Marnet N, Bergmann F, Hackert T, Klauss M, Poth T, Gaida MM. The Microarchitecture of Pancreatic Cancer as Measured by Diffusion-Weighted Magnetic Resonance Imaging Is Altered by T Cells with a Tumor Promoting Th17 Phenotype. International Journal of Molecular Sciences. 2020; 21(1):346. https://doi.org/10.3390/ijms21010346
Chicago/Turabian StyleMayer, Philipp, Alica Linnebacher, Hannah Glennemeier-Marke, Nicole Marnet, Frank Bergmann, Thilo Hackert, Miriam Klauss, Tanja Poth, and Matthias M. Gaida. 2020. "The Microarchitecture of Pancreatic Cancer as Measured by Diffusion-Weighted Magnetic Resonance Imaging Is Altered by T Cells with a Tumor Promoting Th17 Phenotype" International Journal of Molecular Sciences 21, no. 1: 346. https://doi.org/10.3390/ijms21010346
APA StyleMayer, P., Linnebacher, A., Glennemeier-Marke, H., Marnet, N., Bergmann, F., Hackert, T., Klauss, M., Poth, T., & Gaida, M. M. (2020). The Microarchitecture of Pancreatic Cancer as Measured by Diffusion-Weighted Magnetic Resonance Imaging Is Altered by T Cells with a Tumor Promoting Th17 Phenotype. International Journal of Molecular Sciences, 21(1), 346. https://doi.org/10.3390/ijms21010346