A Survey on Tubulin and Arginine Methyltransferase Families Sheds Light on P. lividus Embryo as Model System for Antiproliferative Drug Development


 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
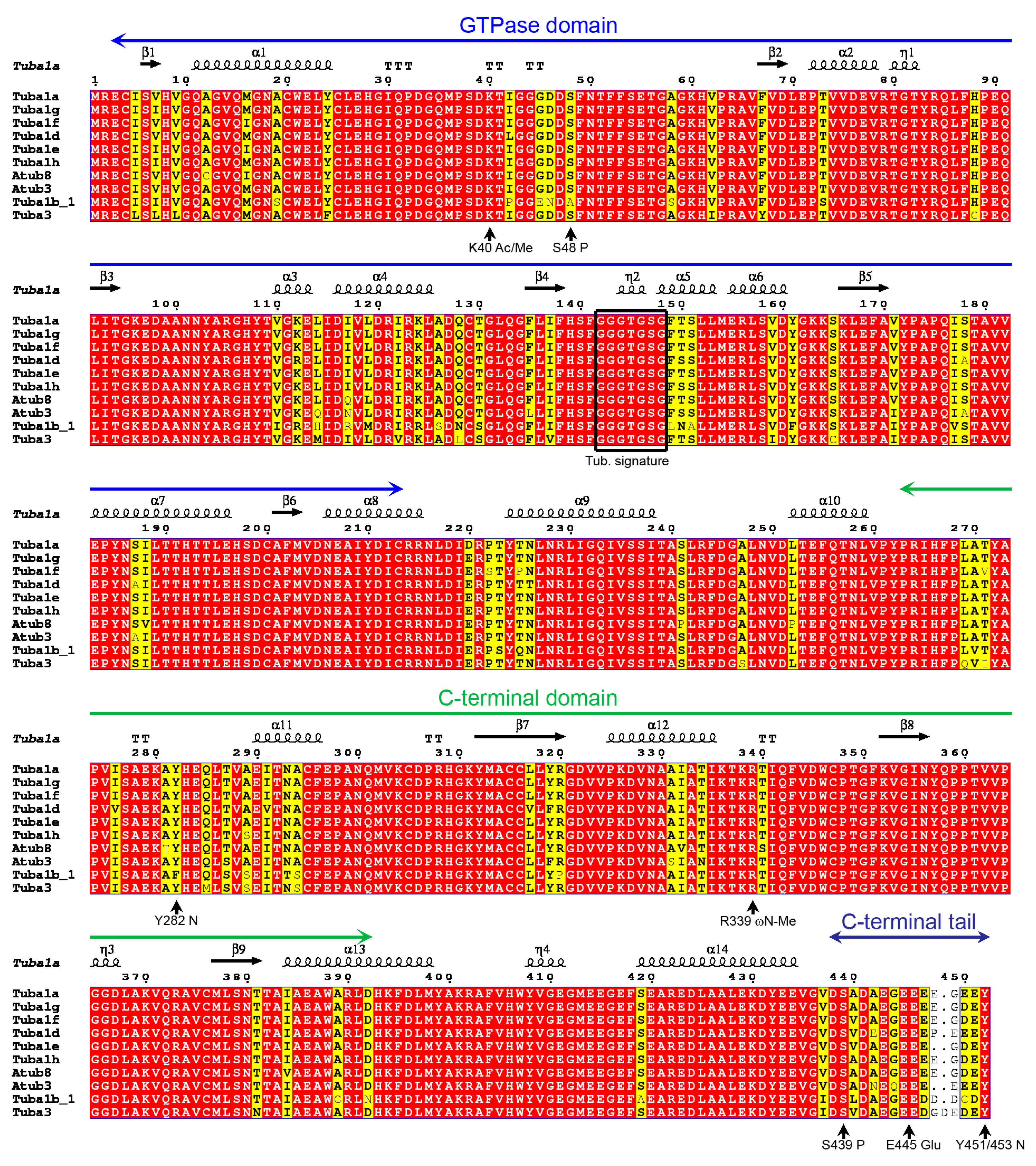
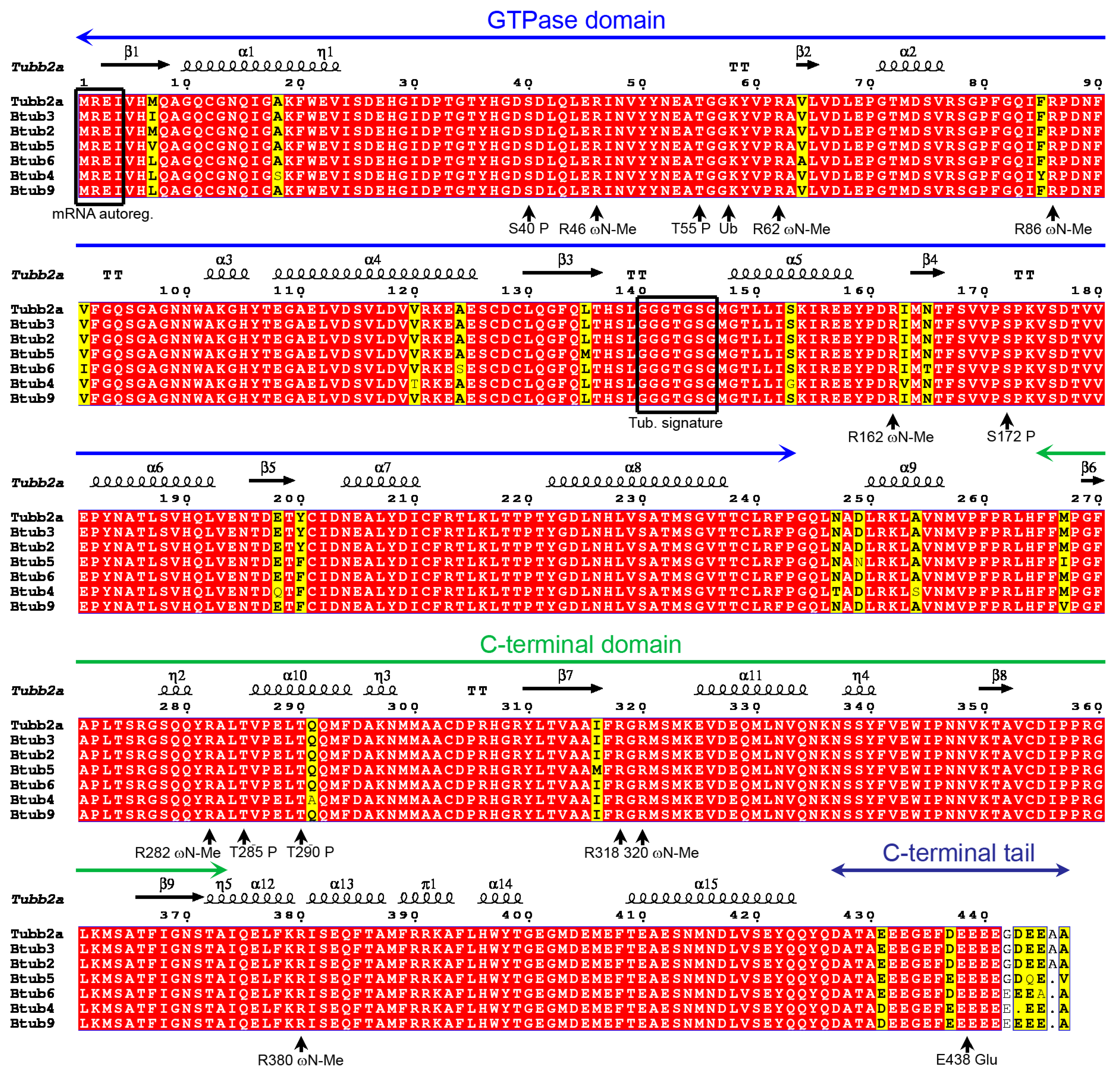
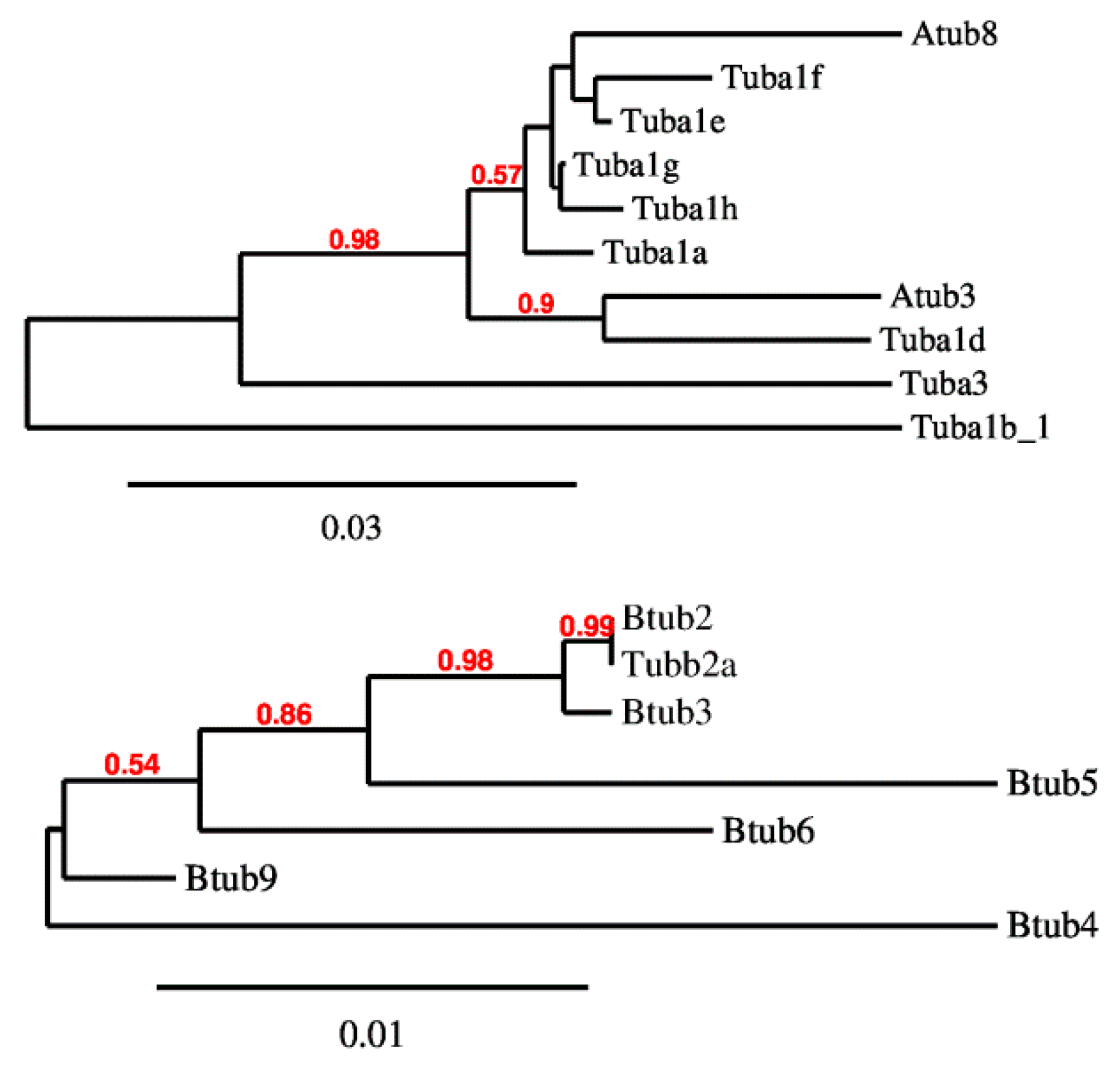
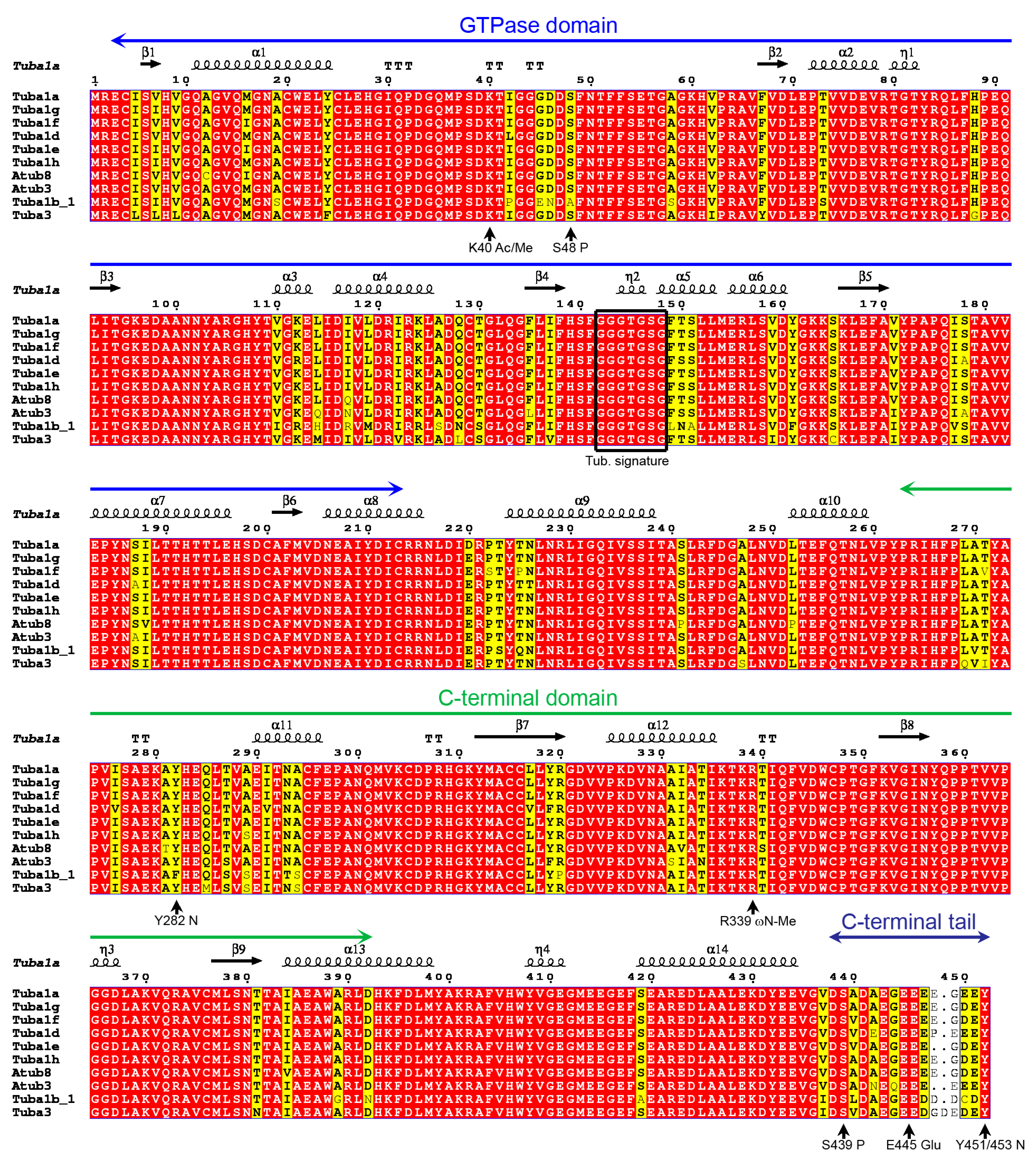
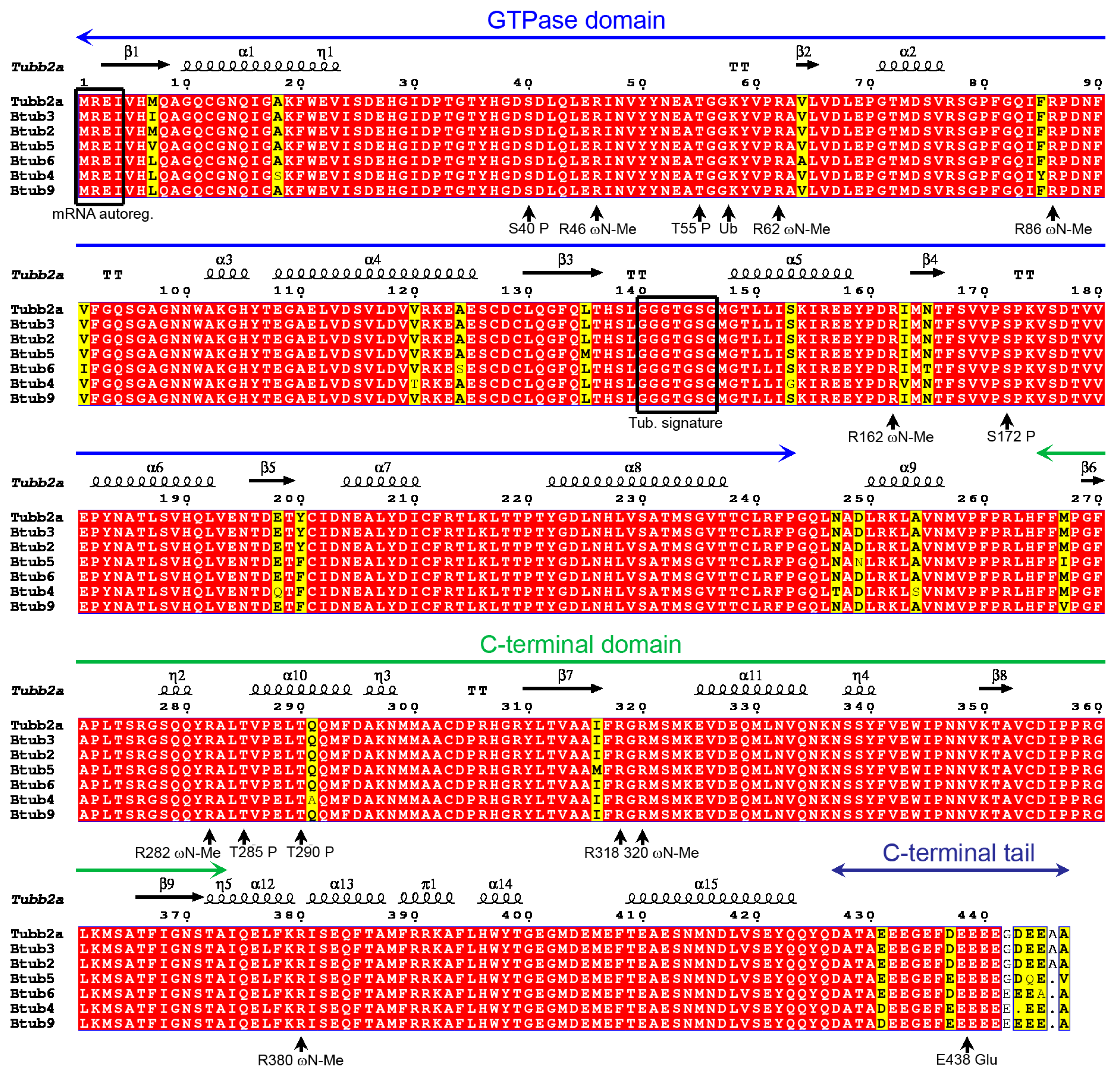
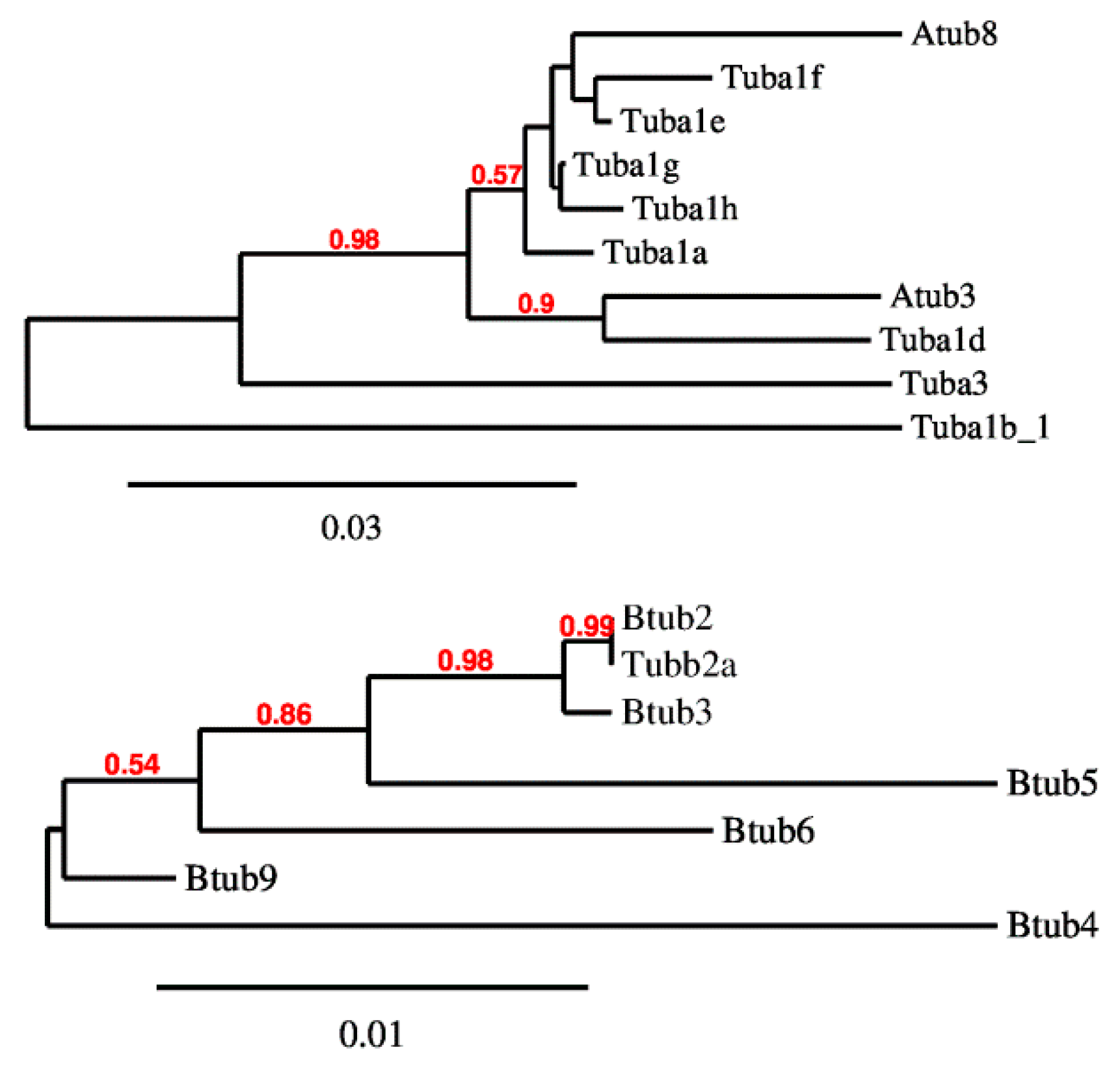
2.1. P. lividus α- and β-Tubulin Identification and Their Predicted PTMs
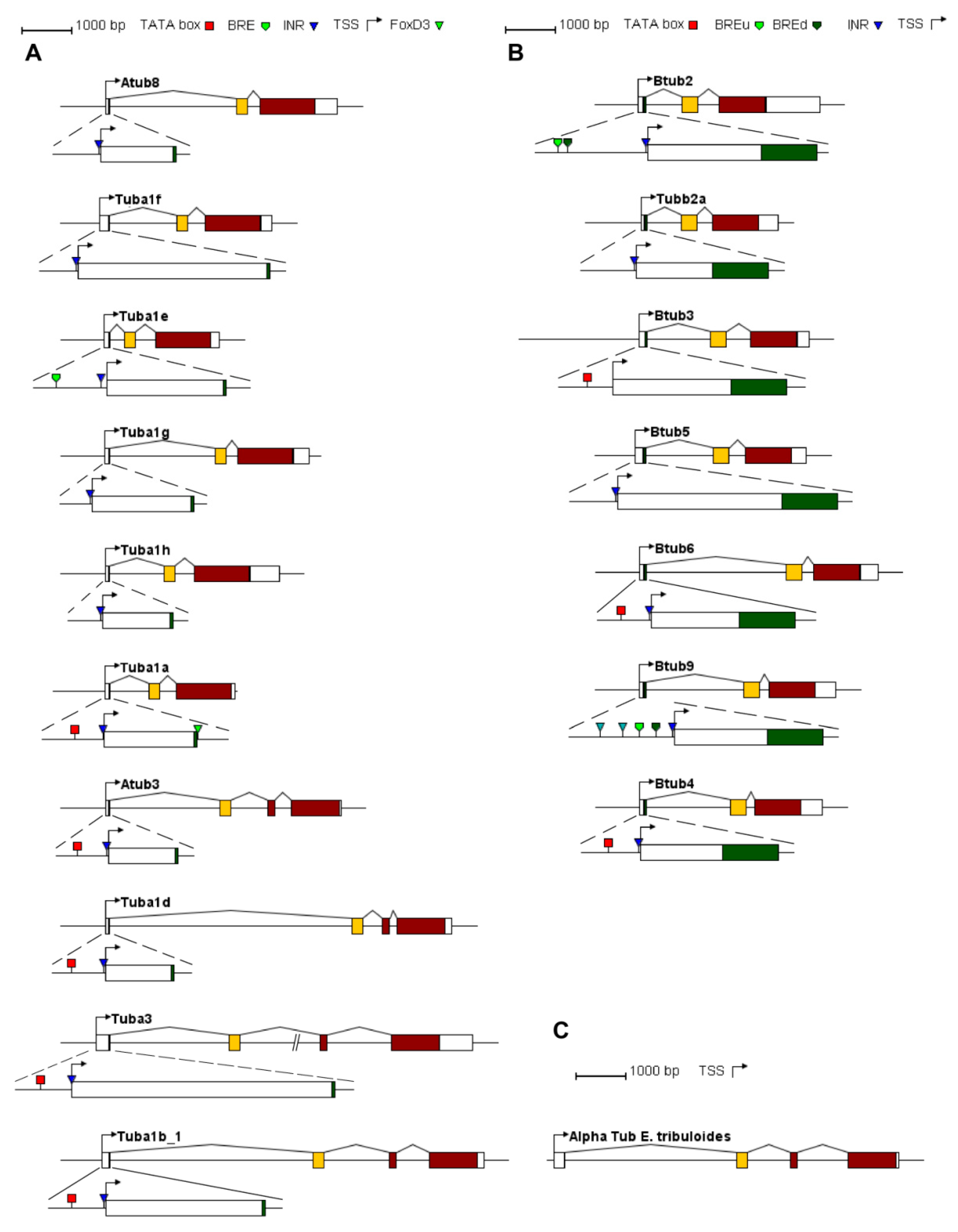
2.2. P. lividus α- and β-Tubulin Gene Organization
2.3. P. lividus α- and β-Tubulin Gene Expression
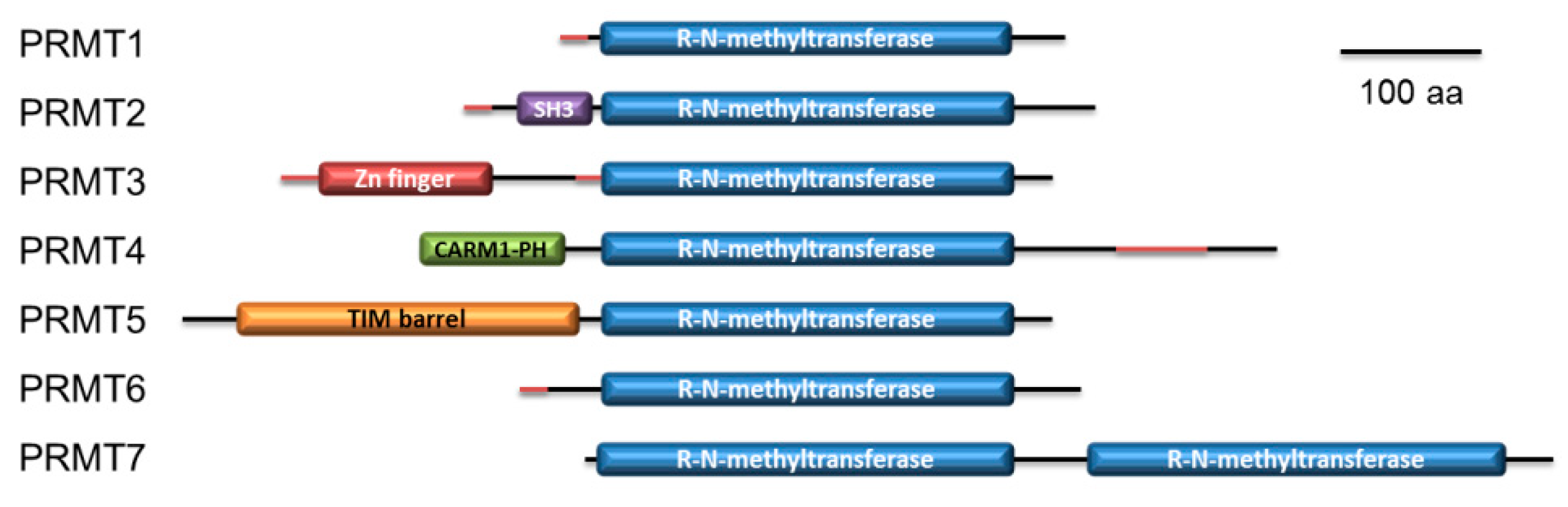
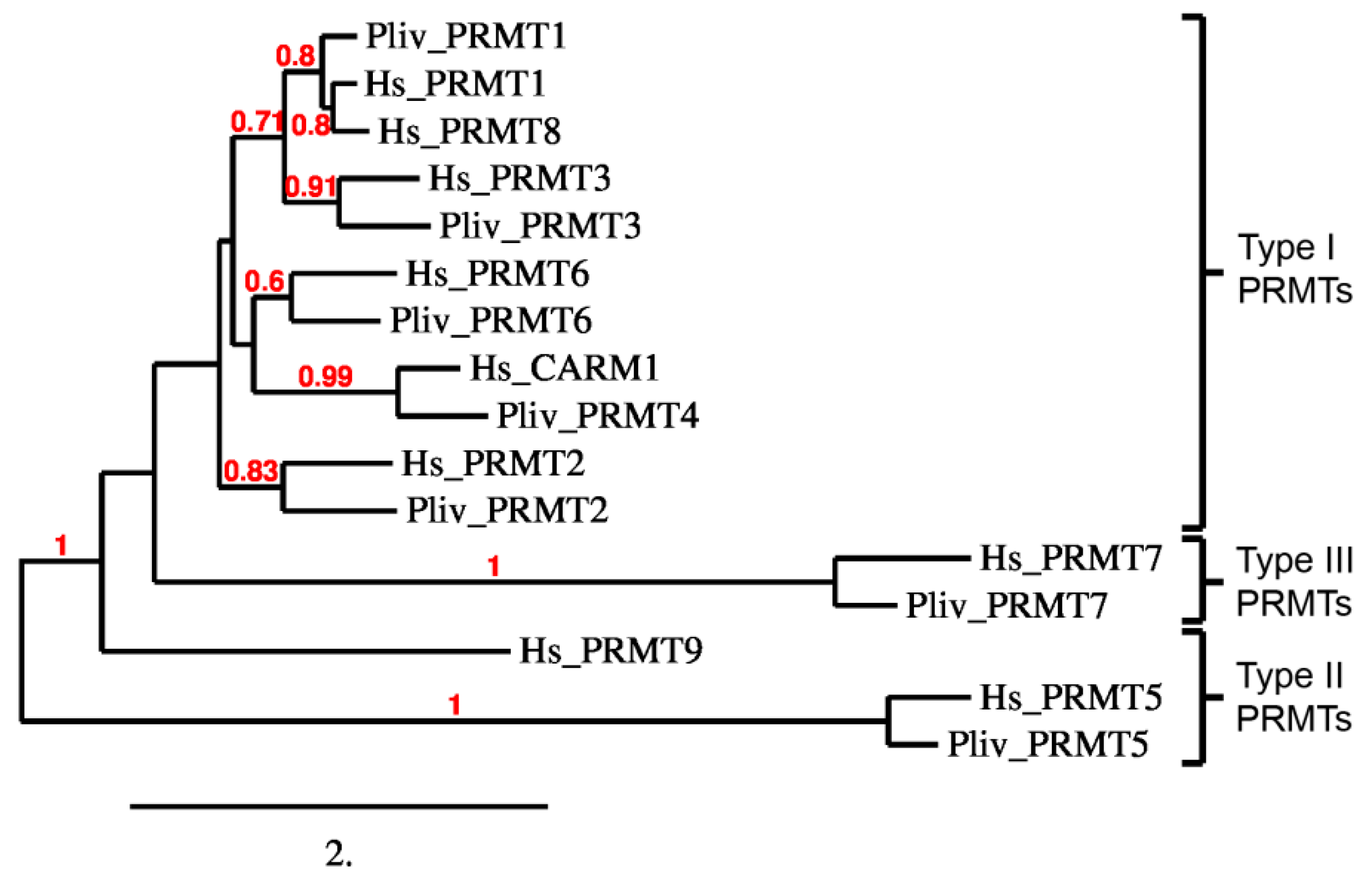
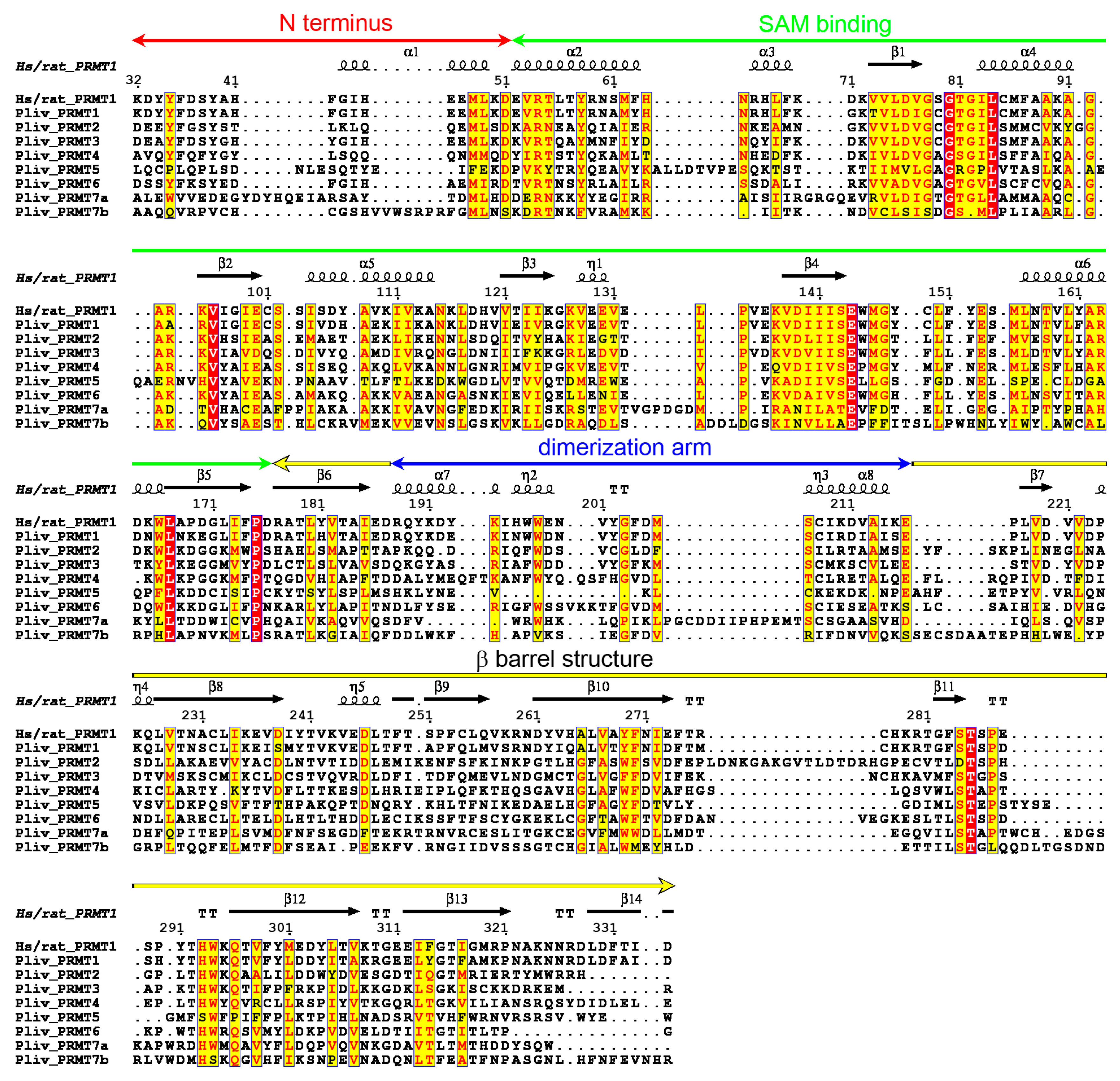
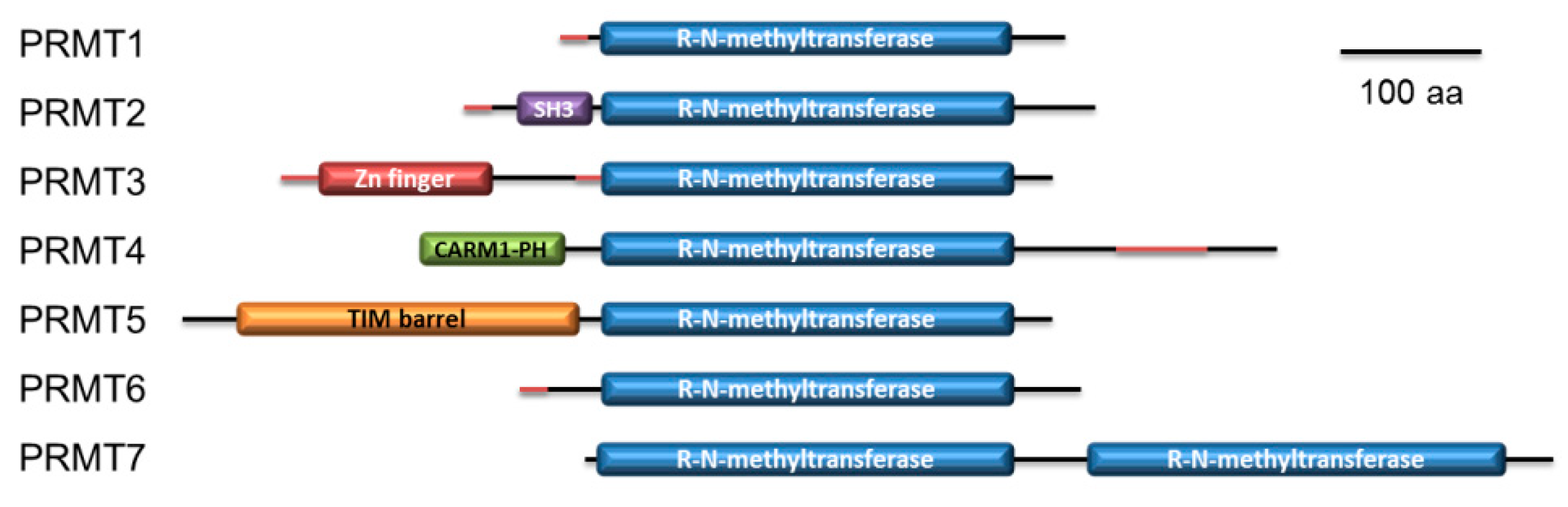
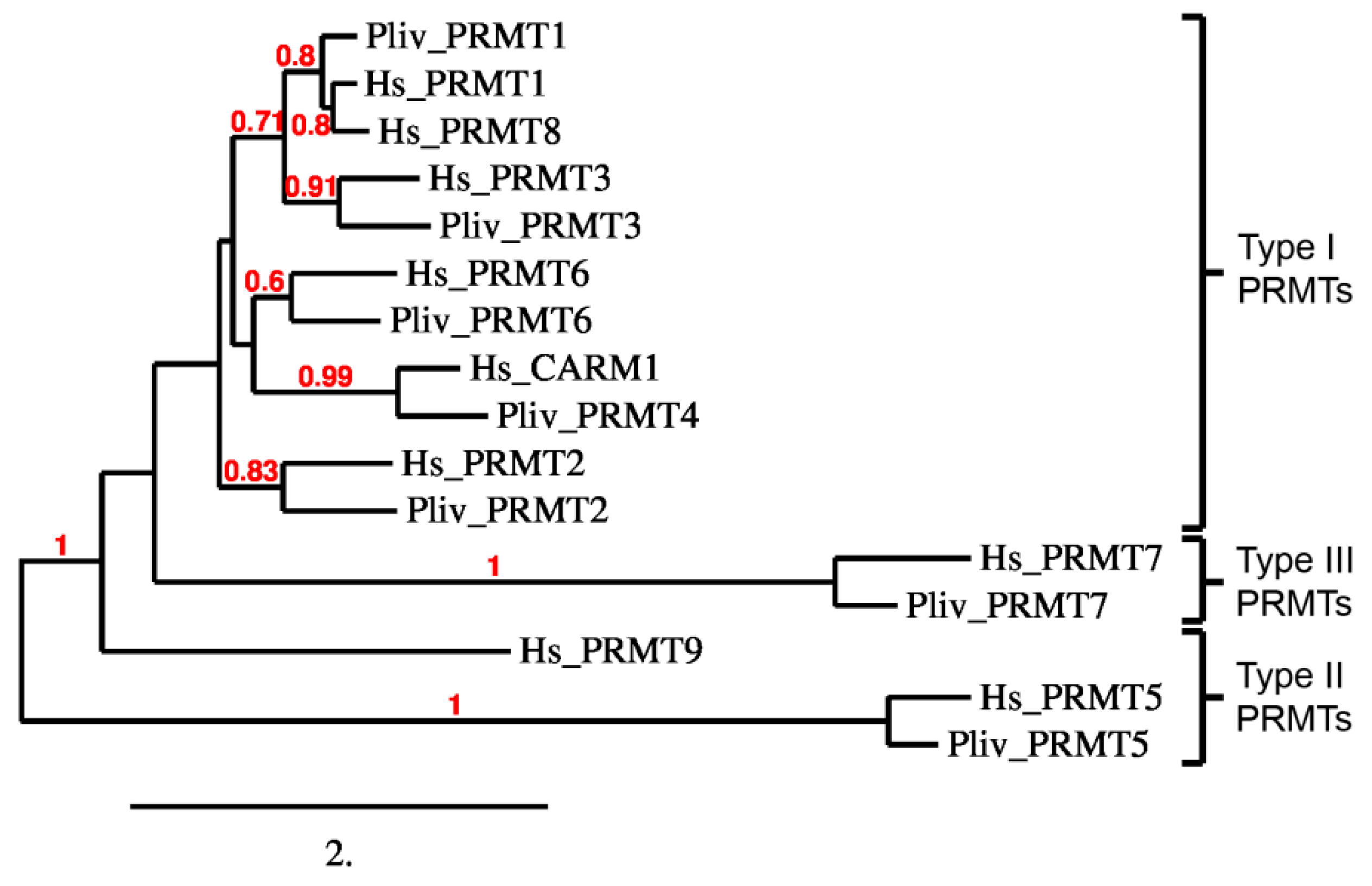
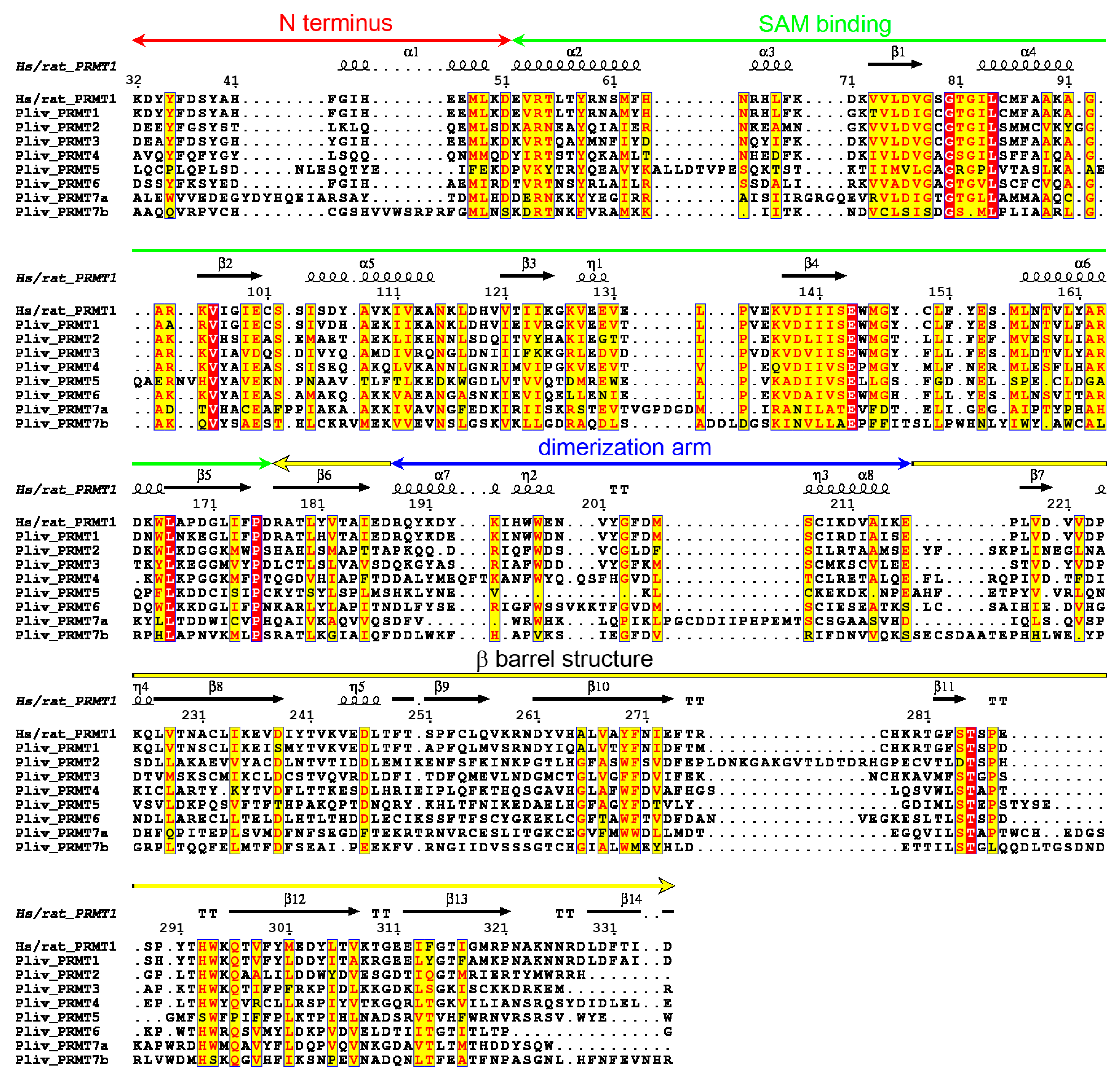
2.4. P. lividus PRMT Orthologues
2.5. P. lividus PRMT Expression
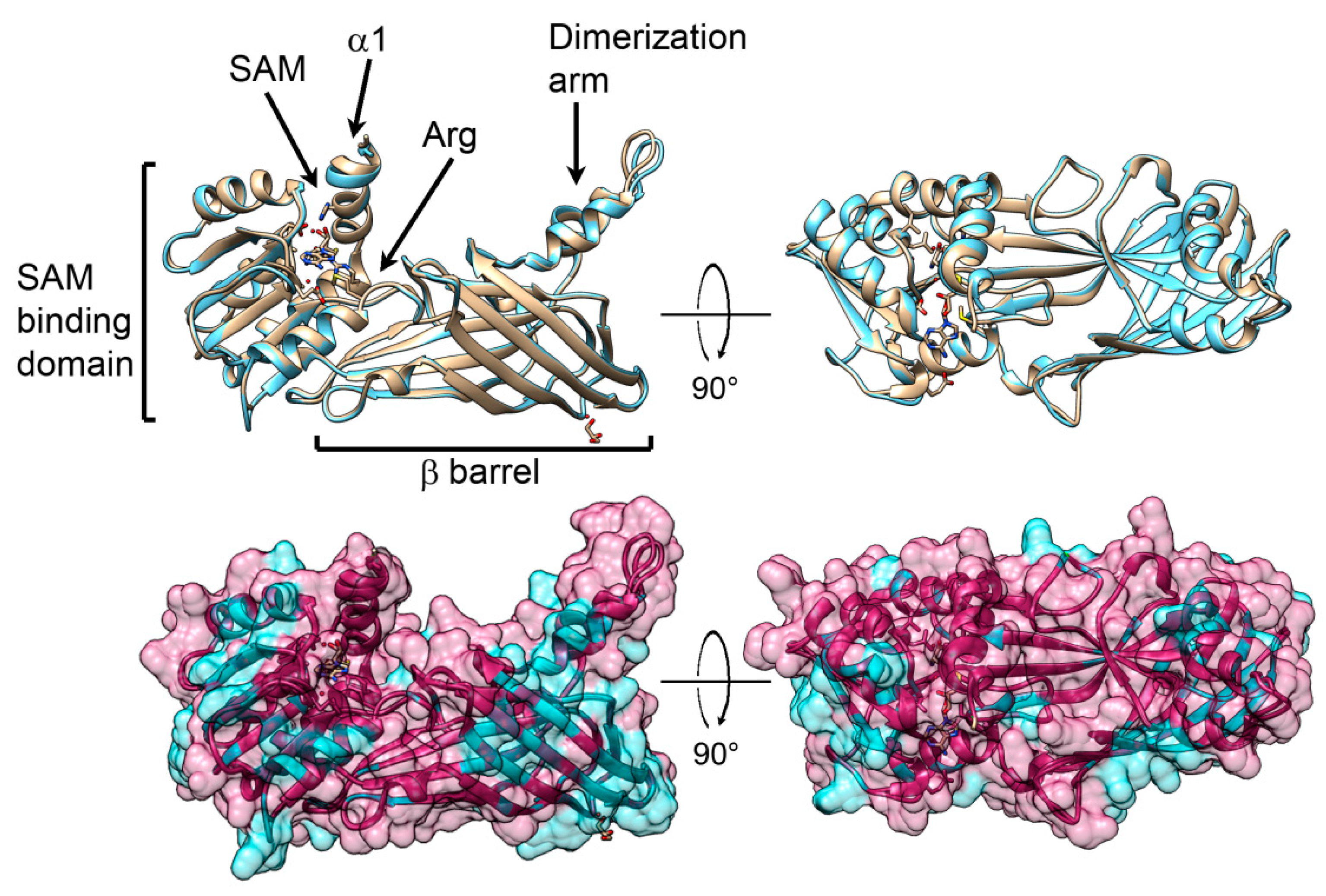
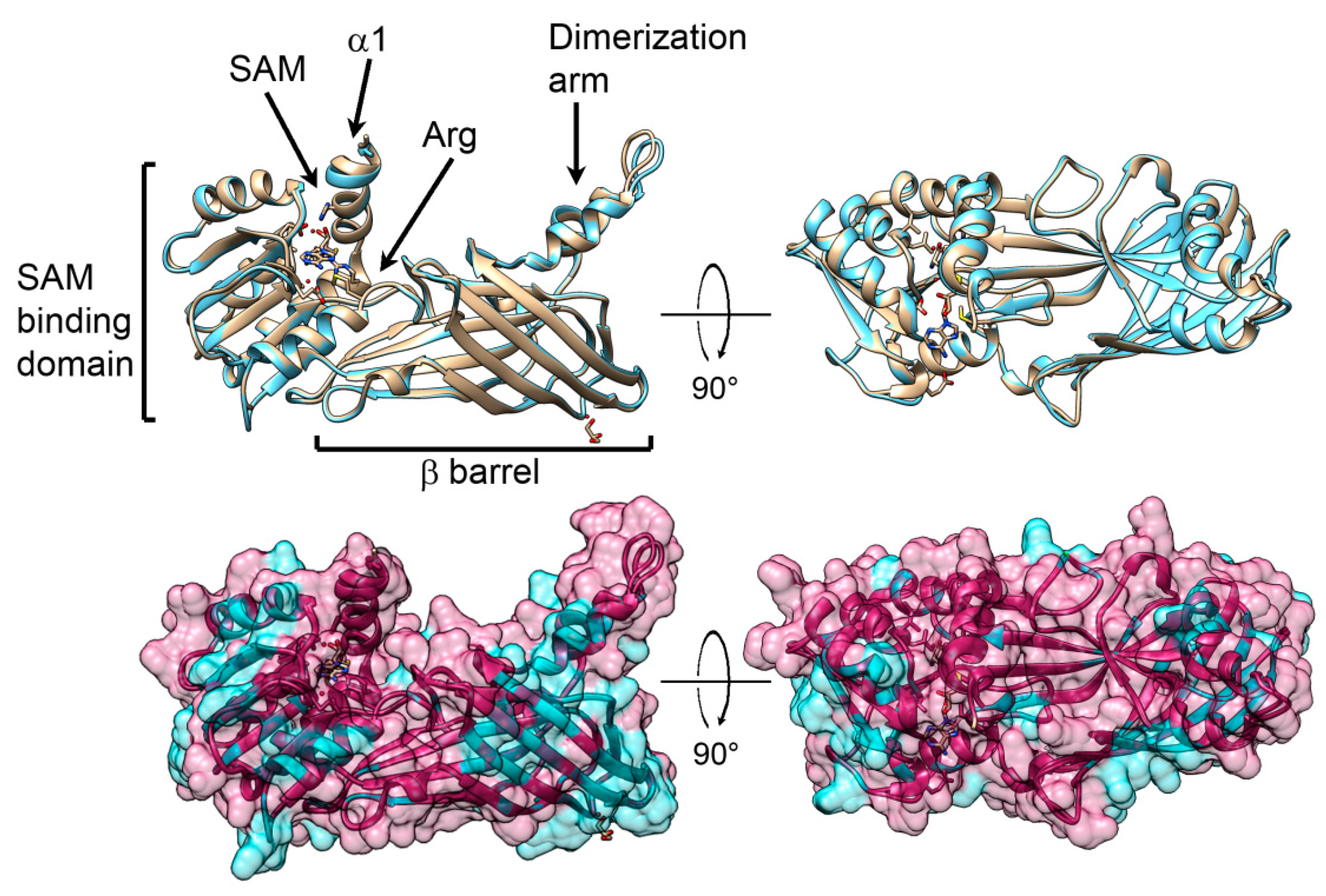
2.6. Predicted 3D Structural Model of P. lividus PRMT1
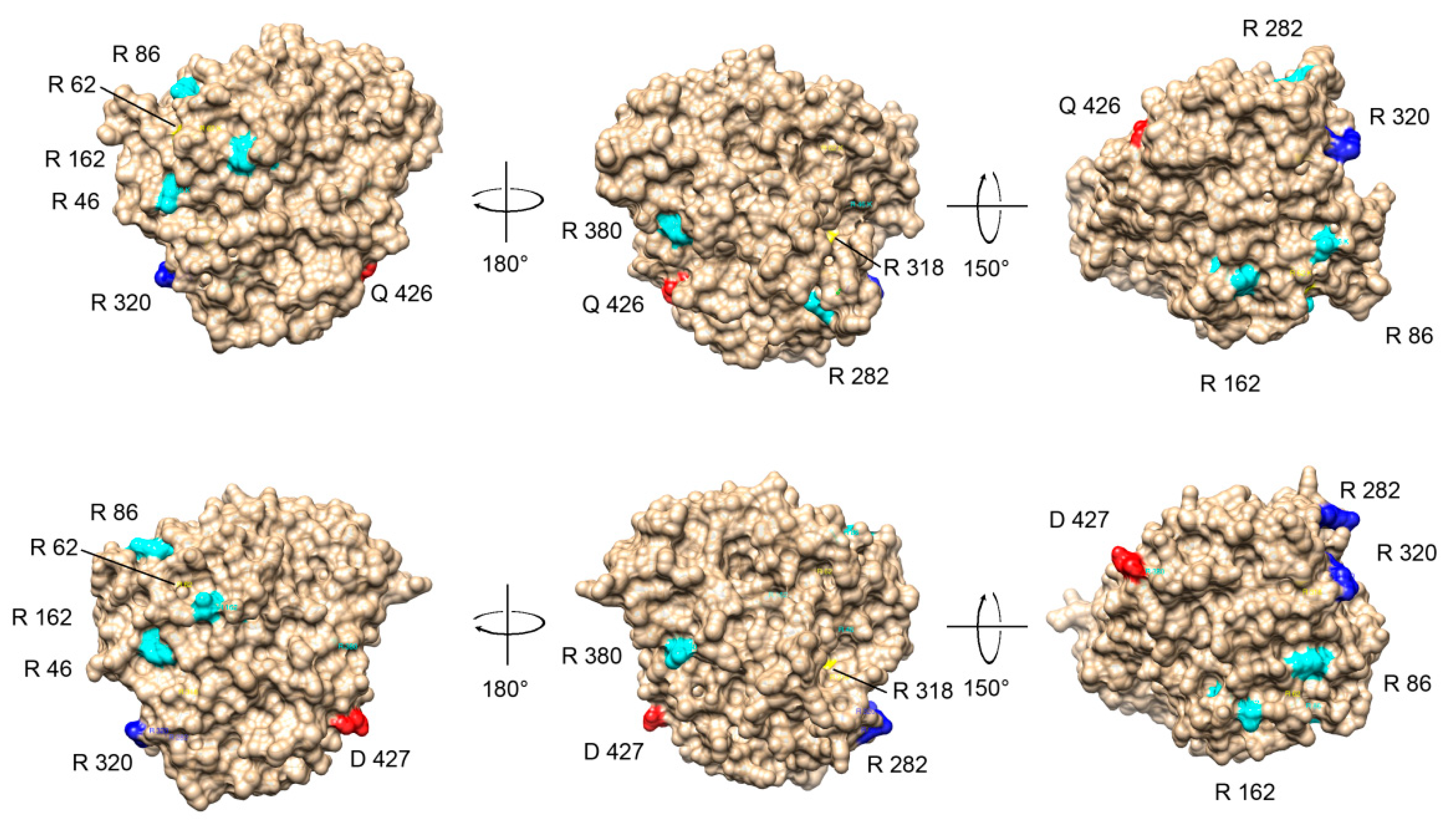
2.7. Tubulin Arginine Methylation
3. Materials and Methods
3.1. Data Mining
3.2. Embryo Cultures
3.3. RNA Purification, Reverse Transcription and DNA Extraction
3.4. cDNA Identification
3.5. RS-PCR and α- and β-Tubulin Gene Organization
3.6. RT-qPCR
3.7. Sequence and Structural Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADMA | asymmetric dimethylarginine |
| MAP | microtubule-associated protein |
| MMA | monomethylarginine |
| MSA | multiple sequence alignment |
| ORF | open reading frame |
| PRMT | protein arginine methyl transferase |
| PTM | post-translational modification |
| SDMA | symmetric dimethylarginine |
| TSS | transcription start site |
| UTR | untranslated region |
References
- Nogales, E. Structural insights into microtubule function. Annu. Rev. Biochem. 2000, 69, 277–302. [Google Scholar] [CrossRef] [PubMed]
- Downing, K.H.; Nogales, E. Tubulin structure: Insights into microtubule properties and functions. Curr. Opin. Struct. Biol. 1998, 8, 785–791. [Google Scholar] [CrossRef]
- Bowne-Anderson, H.; Hibbel, A.; Howard, J. Regulation of Microtubule Growth and Catastrophe: Unifying Theory and Experiment. Trends Cell Biol. 2015, 25, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Etienne-Manneville, S. From signaling pathways to microtubule dynamics: The key players. Curr. Opin. Cell Biol. 2010, 22, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Rosette, C.; Karin, M. Cytoskeletal control of gene expression: Depolymerization of microtubules activates NF-kappa B. J Cell Biol 1995, 128, 1111–1119. [Google Scholar] [CrossRef]
- Cho, S.G.; Sihn, C.-R.; Yoo, S.J.; Cho, K.K.; Lee, H.-G.; Choi, Y.-J.; Kim, S.H. Analysis of gene expression induced by microtubule-disrupting agents in HeLa cells using microarray. Cancer Lett. 2006, 241, 110–117. [Google Scholar] [CrossRef]
- Magidson, V.; O’Connell, C.B.; Lončarek, J.; Paul, R.; Mogilner, A.; Khodjakov, A. The spatial arrangement of chromosomes during prometaphase facilitates spindle assembly. Cell 2011, 146, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Etemad, B.; Kuijt, T.E.F.; Kops, G.J.P.L. Kinetochore-microtubule attachment is sufficient to satisfy the human spindle assembly checkpoint. Nat Commun. 2015, 6, 8987. [Google Scholar] [CrossRef] [PubMed]
- Bakhoum, S.F.; Thompson, S.L.; Manning, A.L.; Compton, D.A. Genome stability is ensured by temporal control of kinetochore-microtubule dynamics. Nat. Cell Biol. 2009, 11, 27–35. [Google Scholar] [CrossRef]
- Parker, A.L.; Teo, W.S.; McCarroll, J.A.; Kavallaris, M. An Emerging Role for Tubulin Isotypes in Modulating Cancer Biology and Chemotherapy Resistance. Int. J. Mol. Sci. 2017, 18, 1434. [Google Scholar] [CrossRef]
- Jordan, M.A. Mechanism of action of antitumor drugs that interact with microtubules and tubulin. Curr. Med. Chem. Anticancer Agents 2002, 2, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Rathinasamy, K.; Mohan, R.; Panda, D. Microtubule assembly dynamics: An attractive target for anticancer drugs. IUBMB Life 2008, 60, 368–375. [Google Scholar] [CrossRef]
- Gan, P.P.; McCarroll, J.A.; Po’uha, S.T.; Kamath, K.; Jordan, M.A.; Kavallaris, M. Microtubule dynamics, mitotic arrest, and apoptosis: Drug-induced differential effects of betaIII-tubulin. Mol. Cancer Ther. 2010, 9, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Kamath, K.; Wilson, L.; Cabral, F.; Jordan, M.A. BetaIII-tubulin induces paclitaxel resistance in association with reduced effects on microtubule dynamic instability. J. Biol. Chem. 2005, 280, 12902–12907. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.H.A.; Wu, S.-Y.; Lee, T.-R.; Chang, C.-Y.; Wu, J.-S.; Hsieh, H.-P.; Chang, J.-Y. Cancer cells acquire mitotic drug resistance properties through beta I-tubulin mutations and alterations in the expression of beta-tubulin isotypes. PLoS ONE 2010, 5, e12564. [Google Scholar] [CrossRef]
- Katsetos, C.D.; Dráber, P. Tubulins as therapeutic targets in cancer: From bench to bedside. Curr. Pharm. Des. 2012, 18, 2778–2792. [Google Scholar] [CrossRef] [PubMed]
- Verhey, K.J.; Gaertig, J. The tubulin code. Cell Cycle 2007, 6, 2152–2160. [Google Scholar] [CrossRef] [PubMed]
- Janke, C. The tubulin code: Molecular components, readout mechanisms, and functions. J. Cell Biol. 2014, 206, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Magiera, M.M.; Janke, C. Post-translational modifications of tubulin. Curr. Biol. 2014, 24, R351–R354. [Google Scholar] [CrossRef] [Green Version]
- Gadadhar, S.; Bodakuntla, S.; Natarajan, K.; Janke, C. The tubulin code at a glance. J. Cell Sci. 2017, 130, 1347–1353. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Brady, S.T. Post-translational modifications of tubulin: Pathways to functional diversity of microtubules. Trends Cell Biol. 2015, 25, 125–136. [Google Scholar] [CrossRef]
- Piller, S.; Jwad, N.; Hejazi, L.; Gamsjaeger, R.; Sucher, N. Protein Arginine Methylation of Tubulin Beta Decreases Binding of Taxol in Neuro2a Cells. FASEB J. 2015, 29, 717.16. [Google Scholar]
- Raposo, A.E.; Piller, S.C. Protein arginine methylation: An emerging regulator of the cell cycle. Cell Div. 2018, 13. [Google Scholar] [CrossRef]
- Nielsen, M.G.; Gadagkar, S.R.; Gutzwiller, L. Tubulin evolution in insects: Gene duplication and subfunctionalization provide specialized isoforms in a functionally constrained gene family. BMC Evol. Biol. 2010, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- Ludueña, R.F. Are tubulin isotypes functionally significant. Mol. Biol. Cell 1993, 4, 445–457. [Google Scholar]
- Verdier-Pinard, P.; Pasquier, E.; Xiao, H.; Burd, B.; Villard, C.; Lafitte, D.; Miller, L.M.; Angeletti, R.H.; Horwitz, S.B.; Braguer, D. Tubulin proteomics: Towards breaking the code. Anal. Biochem. 2009, 384, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, R.L.; Hoffman, M.P.; Obar, R.A.; McCafferty, S.S.; Gibbons, I.R.; Leone, A.D.; Cool, J.; Allgood, E.L.; Musante, A.M.; Judkins, K.M.; et al. Analysis of cytoskeletal and motility proteins in the sea urchin genome assembly. Dev. Biol. 2006, 300, 219–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briggs, E.; Wessel, G.M. In the beginning… Animal fertilization and sea urchin development. Dev. Biol. 2006, 300, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pederson, T. The sea urchin’s siren. Dev. Biol. 2006, 300, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Cavalieri, V.; Spinelli, G. Early asymmetric cues triggering the dorsal/ventral gene regulatory network of the sea urchin embryo. Elife 2014, 3, e04664. [Google Scholar] [CrossRef]
- Molina, M.D.; Quirin, M.; Haillot, E.; Jimenez, F.; Chessel, A.; Lepage, T. p38 MAPK as an essential regulator of dorsal-ventral axis specification and skeletogenesis during sea urchin development: A re-evaluation. Development 2017, 144, 2270–2281. [Google Scholar] [CrossRef]
- Howard-Ashby, M.; Materna, S.C.; Brown, C.T.; Tu, Q.; Oliveri, P.; Cameron, R.A.; Davidson, E.H. High regulatory gene use in sea urchin embryogenesis: Implications for bilaterian development and evolution. Dev. Biol. 2006, 300, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Britten, R.J.; Davidson, E.H. Gene regulation for higher cells: A theory. Science 1969, 165, 349–357. [Google Scholar] [CrossRef]
- Davidson, E.H. The Sea Urchin Genome: Where Will It Lead Us? Science 2006, 314, 939–940. [Google Scholar] [CrossRef] [Green Version]
- Chiarelli, R.; Martino, C.; Agnello, M.; Bosco, L.; Roccheri, M.C. Autophagy as a defense strategy against stress: Focus on Paracentrotus lividus sea urchin embryos exposed to cadmium. Cell Stress Chaperones 2016, 21, 19–27. [Google Scholar] [CrossRef]
- Pinsino, A.; Matranga, V. Sea urchin immune cells as sentinels of environmental stress. Dev. Comp. Immunol. 2015, 49, 198–205. [Google Scholar] [CrossRef]
- Boveri, T. Zur frage der entstehung maligner tumoren (Jena: Gustav Fischer) (English translation); Boveri, M., Translator; Williams and Wilkins: Baltimore, MD, USA, 1929. [Google Scholar]
- Knudson, A.G. Of sea urchins and worms: Development and cancer. Cell Death Differ. 2004, 11, 11–12. [Google Scholar] [CrossRef]
- Hultin, T. The effect of puromycin on proteinmetabolism and cell division in fertilized sea ur-chin eggs. Experientia 1961, 17, 410–411. [Google Scholar] [CrossRef]
- Evans, T.; Rosenthal, E.T.; Youngblom, J.; Distel, D.; Hunt, T. Cyclin: A protein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage division. Cell 1983, 33, 389–396. [Google Scholar] [CrossRef]
- Semenova, M.N.; Kiselyov, A.; Semenov, V.V. Sea urchin embryo as a model organism for the rapid functional screening of tubulin modulators. BioTechniques 2006, 40, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Hamaguchi, Y. Displacement of cleavage plane in the sea urchin egg by locally applied taxol. Cell Motil. Cytoskelet. 1998, 40, 211–219. [Google Scholar] [CrossRef]
- Kiselyov, A.S.; Semenova, M.N.; Chernyshova, N.B.; Leitao, A.; Samet, A.V.; Kislyi, K.A.; Raihstat, M.M.; Oprea, T.; Lemcke, H.; Lantow, M.; et al. Novel derivatives of 1,3,4-oxadiazoles are potent mitostatic agents featuring strong microtubule depolymerizing activity in the sea urchin embryo and cell culture assays. Eur. J. Med. Chem. 2010, 45, 1683–1697. [Google Scholar] [CrossRef]
- Demchuk, D.V.; Samet, A.V.; Chernysheva, N.B.; Ushkarov, V.I.; Stashina, G.A.; Konyushkin, L.D.; Raihstat, M.M.; Firgang, S.I.; Philchenkov, A.A.; Zavelevich, M.P.; et al. Synthesis and antiproliferative activity of conformationally restricted 1,2,3-triazole analogues of combretastatins in the sea urchin embryo model and against human cancer cell lines. Bioorg. Med. Chem. 2014, 22, 738–755. [Google Scholar] [CrossRef]
- Strobykina, I.Y.; Belenok, M.G.; Semenova, M.N.; Semenov, V.V.; Babaev, V.M.; Rizvanov, I.K.; Mironov, V.F.; Kataev, V.E. Triphenylphosphonium Cations of the Diterpenoid Isosteviol: Synthesis and Antimitotic Activity in a Sea Urchin Embryo Model. J. Nat. Prod. 2015, 78, 1300–1308. [Google Scholar] [CrossRef]
- Di Bernardo, M.G.; Gianguzza, F.; Ciaccio, M.; Palla, F.; Colombo, P.; Di Blasi, F.; Fais, M.; Spinelli, G. Nucleotide sequence of a full length cDNA clone encoding for beta-tubulin of the sea urchin Paracentrotus lividus. Nucleic Acids Res. 1989, 17, 5851. [Google Scholar] [CrossRef]
- Gianguzza, F.; Di Bernardo, M.G.; Sollazzo, M.; Palla, F.; Ciaccio, M.; Carra, E.; Spinelli, G. DNA sequence and pattern of expression of the sea urchin (Paracentrotus lividus) alpha-tubulin genes. Mol. Reprod. Dev. 1989, 1, 170–181. [Google Scholar] [CrossRef]
- Gianguzza, F.; Di Bernardo, M.G.; Fais, M.; Palla, F.; Casano, C.; Russo, R.; Spinelli, G. Sequence and expression of Paracentrotus lividus alpha tubulin gene. Nucleic Acids Res. 1990, 18, 4915. [Google Scholar] [CrossRef]
- Gianguzza, F.; Casano, C.; Ragusa, M. Alpha-tubulin marker gene of neural territory of sea urchin embryos detected by whole-mount in situ hybridization. Int. J. Dev. Biol. 1995, 39, 477–483. [Google Scholar]
- Casano, C.; Ragusa, M.; Cutrera, M.; Costa, S.; Gianguzza, F. Spatial expression of alpha and beta tubulin genes in the late embryogenesis of the sea urchin Paracentrotus lividus. Int. J. Dev. Biol. 1996, 40, 1033–1041. [Google Scholar]
- Costa, S.; Ragusa, M.A.; Drago, G.; Casano, C.; Alaimo, G.; Guida, N.; Gianguzza, F. Sea urchin neural alpha2 tubulin gene: Isolation and promoter analysis. Biochem. Biophys. Res. Commun. 2004, 316, 446–453. [Google Scholar] [CrossRef]
- Emanuele, M.; Costa, S.; Ragusa, M.A.; Gianguzza, F. Chromatin dynamics of the developmentally regulated P. lividus neural alpha tubulin gene. Int. J. Dev. Biol. 2011, 55, 591–596. [Google Scholar] [CrossRef]
- Ragusa, M.A.; Longo, V.; Emanuele, M.; Costa, S.; Gianguzza, F. In silico characterization of the neural alpha tubulin gene promoter of the sea urchin embryo Paracentrotus lividus by phylogenetic footprinting. Mol. Biol. Rep. 2012, 39, 2633–2644. [Google Scholar] [CrossRef]
- Costa, S.; Nicosia, A.; Cuttitta, A.; Gianguzza, F.; Ragusa, M.A. An Intronic cis-Regulatory Element Is Crucial for the Alpha Tubulin Pl-Tuba1a Gene Activation in the Ciliary Band and Animal Pole Neurogenic Domains during Sea Urchin Development. PLoS ONE 2017, 12, e0170969. [Google Scholar] [CrossRef]
- Gildor, T.; Malik, A.; Sher, N.; Avraham, L.; Ben-Tabou de-Leon, S. Quantitative developmental transcriptomes of the Mediterranean sea urchin Paracentrotus lividus. Mar. Genom. 2016, 25, 89–94. [Google Scholar] [CrossRef]
- Di Natale, M.; Bennici, C.; Biondo, G.; Masullo, T.; Monastero, C.; Tagliavia, M.; Torri, M.; Costa, S.; Ragusa, M.A.; Cuttitta, A.; et al. Aberrant gene expression profiles in Mediterranean sea urchin reproductive tissues after metal exposures. Chemosphere 2019, 216, 48–58. [Google Scholar] [CrossRef]
- Ragusa, M.A.; Costa, S.; Gianguzza, M.; Roccheri, M.C.; Gianguzza, F. Effects of cadmium exposure on sea urchin development assessed by SSH and RT-qPCR: Metallothionein genes and their differential induction. Mol. Biol. Rep. 2013, 40, 2157–2167. [Google Scholar] [CrossRef]
- Ragusa, M.A.; Nicosia, A.; Costa, S.; Cuttitta, A.; Gianguzza, F. Metallothionein Gene Family in the Sea Urchin Paracentrotus lividus: Gene Structure, Differential Expression and Phylogenetic Analysis. Int. J. Mol. Sci. 2017, 18, 812. [Google Scholar] [CrossRef]
- Fry, A.M.; O’Regan, L.; Montgomery, J.; Adib, R.; Bayliss, R. EML proteins in microtubule regulation and human disease. Biochem. Soc. Trans. 2016, 44, 1281–1288. [Google Scholar] [CrossRef] [Green Version]
- Fourest-Lieuvin, A.; Peris, L.; Gache, V.; Garcia-Saez, I.; Juillan-Binard, C.; Lantez, V.; Job, D. Microtubule Regulation in Mitosis: Tubulin Phosphorylation by the Cyclin-dependent Kinase Cdk1. Mol. Biol. Cell 2006, 17, 1041–1050. [Google Scholar] [CrossRef] [Green Version]
- Gambardella, C.; Morgana, S.; Bari, G.D.; Ramoino, P.; Bramini, M.; Diaspro, A.; Falugi, C.; Faimali, M. Multidisciplinary screening of toxicity induced by silica nanoparticles during sea urchin development. Chemosphere 2015, 139, 486–495. [Google Scholar] [CrossRef]
- Stephens, R.E. Tubulin in sea urchin embryonic cilia: Post-translational modifications during regeneration. J. Cell Sci. 1992, 101 Pt 4, 837–845. [Google Scholar]
- Portran, D.; Schaedel, L.; Xu, Z.; Théry, M.; Nachury, M.V. Tubulin acetylation protects long-lived microtubules against mechanical ageing. Nat. Cell Biol. 2017, 19, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Schaedel, L.; Portran, D.; Aguilar, A.; Gaillard, J.; Marinkovich, M.P.; Théry, M.; Nachury, M.V. Microtubules acquire resistance from mechanical breakage through intralumenal acetylation. Science 2017, 356, 328–332. [Google Scholar] [CrossRef]
- Barisic, M.; Silva e Sousa, R.; Tripathy, S.K.; Magiera, M.M.; Zaytsev, A.V.; Pereira, A.L.; Janke, C.; Grishchuk, E.L.; Maiato, H. Microtubule detyrosination guides chromosomes during mitosis. Science 2015, 348, 799–803. [Google Scholar] [CrossRef] [Green Version]
- Sirajuddin, M.; Rice, L.M.; Vale, R.D. Regulation of microtubule motors by tubulin isotypes and post-translational modifications. Nat. Cell Biol. 2014, 16, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, D.W. Autoregulated control of tubilin synthesis in animal cells. Curr. Opin. Cell Biol. 1989, 1, 10–14. [Google Scholar] [CrossRef]
- Eddé, B.; Rossier, J.; Le Caer, J.P.; Desbruyères, E.; Gros, F.; Denoulet, P. Posttranslational glutamylation of alpha-tubulin. Science 1990, 247, 83–85. [Google Scholar] [CrossRef]
- Redeker, V.; Levilliers, N.; Schmitter, J.M.; Le Caer, J.P.; Rossier, J.; Adoutte, A.; Bré, M.H. Polyglycylation of tubulin: A posttranslational modification in axonemal microtubules. Science 1994, 266, 1688–1691. [Google Scholar] [CrossRef]
- Mary, J.; Redeker, V.; Le Caer, J.P.; Rossier, J.; Schmitter, J.M. Posttranslational modifications in the C-terminal tail of axonemal tubulin from sea urchin sperm. J. Biol. Chem. 1996, 271, 9928–9933. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, C.; White, D.; Cosson, J.; Huitorel, P.; Eddé, B.; Desbruyères, E.; Paturle-Lafanechère, L.; Multigner, L.; Job, D.; Cibert, C. The polyglutamylated lateral chain of alpha-tubulin plays a key role in flagellar motility. J. Cell Sci. 1996, 109, 1545–1553. [Google Scholar] [PubMed]
- Huitorel, P.; White, D.; Fouquet, J.-P.; Kann, M.-L.; Cosson, J.; Gagnon, C. Differential distribution of glutamylated tubulin isoforms along the sea urchin sperm axoneme. Mol. Reprod. Dev. 2002, 62, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Kann, M.-L.; Soues, S.; Levilliers, N.; Fouquet, J.-P. Glutamylated tubulin: Diversity of expression and distribution of isoforms. Cell Motil. Cytoskelet. 2003, 55, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Löwe, J.; Li, H.; Downing, K.H.; Nogales, E. Refined structure of alpha beta-tubulin at 3.5 A resolution. J. Mol. Biol. 2001, 313, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Park, I.Y.; Chowdhury, P.; Tripathi, D.N.; Powell, R.T.; Dere, R.; Terzo, E.A.; Rathmell, W.K.; Walker, C.L. Methylated α-tubulin antibodies recognize a new microtubule modification on mitotic microtubules. MAbs 2016, 8, 1590–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, C.N.I.; Gasteiger, E.; Wilkins, M.R. Identification of arginine- and lysine-methylation in the proteome of Saccharomyces cerevisiae and its functional implications. BMC Genom. 2010, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Gu, H.; Zhou, J.; Mulhern, D.; Wang, Y.; Lee, K.A.; Yang, V.; Aguiar, M.; Kornhauser, J.; Jia, X.; et al. Immunoaffinity Enrichment and Mass Spectrometry Analysis of Protein Methylation. Mol. Cell. Proteom. 2014, 13, 372–387. [Google Scholar] [CrossRef] [PubMed]
- Plett, K.L.; Raposo, A.E.; Bullivant, S.; Anderson, I.C.; Piller, S.C.; Plett, J.M. Root morphogenic pathways in Eucalyptus grandis are modified by the activity of protein arginine methyltransferases. BMC Plant Biol. 2017, 17, 62. [Google Scholar] [CrossRef]
- Alushin, G.M.; Lander, G.C.; Kellogg, E.H.; Zhang, R.; Baker, D.; Nogales, E. High-resolution microtubule structures reveal the structural transitions in αβ-tubulin upon GTP hydrolysis. Cell 2014, 157, 1117–1129. [Google Scholar] [CrossRef]
- Chaaban, S.; Brouhard, G.J. A microtubule bestiary: Structural diversity in tubulin polymers. Mol. Biol. Cell 2017, 28, 2924–2931. [Google Scholar] [CrossRef]
- Sitbon, E.; Pietrokovski, S. Occurrence of protein structure elements in conserved sequence regions. BMC Struct. Biol. 2007, 7, 3. [Google Scholar] [CrossRef]
- Nicosia, A.; Maggio, T.; Costa, S.; Salamone, M.; Tagliavia, M.; Mazzola, S.; Gianguzza, F.; Cuttitta, A. Maintenance of a Protein Structure in the Dynamic Evolution of TIMPs over 600 Million Years. Genome Biol. Evol. 2016, 8, 1056–1071. [Google Scholar] [CrossRef] [Green Version]
- Perumal, B.S.; Sakharkar, K.R.; Chow, V.T.K.; Pandjassarame, K.; Sakharkar, M.K. Intron position conservation across eukaryotic lineages in tubulin genes. Front. Biosci. 2005, 10, 2412–2419. [Google Scholar] [CrossRef]
- Morales, Y.; Cáceres, T.; May, K.; Hevel, J.M. Biochemistry and regulation of the protein arginine methyltransferases (PRMTs). Arch. Biochem. Biophys. 2016, 590, 138–152. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Cheng, X. Structure of the Predominant Protein Arginine Methyltransferase PRMT1 and Analysis of Its Binding to Substrate Peptides. Structure 2003, 11, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Frankel, A.; Cook, R.J.; Kim, S.; Paik, W.K.; Williams, K.R.; Clarke, S.; Herschman, H.R. PRMT1 is the predominant type I protein arginine methyltransferase in mammalian cells. J. Biol. Chem. 2000, 275, 7723–7730. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein-protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef]
- Sarkar, G.; Turner, R.T.; Bolander, M.E. Restriction-site PCR: A direct method of unknown sequence retrieval adjacent to a known locus by using universal primers. Genome Res. 1993, 2, 318–322. [Google Scholar] [CrossRef]
- Solovyev, V.; Kosarev, P.; Seledsov, I.; Vorobyev, D. Automatic annotation of eukaryotic genes, pseudogenes and promoters. Genome Biol. 2006, 7, S10. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- Di Tommaso, P.; Moretti, S.; Xenarios, I.; Orobitg, M.; Montanyola, A.; Chang, J.-M.; Taly, J.-F.; Notredame, C. T-Coffee: A web server for the multiple sequence alignment of protein and RNA sequences using structural information and homology extension. Nucleic Acids Res. 2011, 39, W13–W17. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Salamone, M.; Nicosia, A.; Bennici, C.; Quatrini, P.; Catania, V.; Mazzola, S.; Ghersi, G.; Cuttitta, A. Comprehensive analysis of a Vibrio parahaemolyticus strain extracellular serine protease VpSP37. PLoS ONE 2015, 10, e0126349. [Google Scholar] [CrossRef]
- Cuttitta, A.; Ragusa, M.A.; Costa, S.; Bennici, C.; Colombo, P.; Mazzola, S.; Gianguzza, F.; Nicosia, A. Evolutionary conserved mechanisms pervade structure and transcriptional modulation of allograft inflammatory factor-1 from sea anemone Anemonia viridis. Fish Shellfish. Immunol. 2017, 67, 86–94. [Google Scholar] [CrossRef]
- Nicosia, A.; Bennici, C.; Biondo, G.; Costa, S.; Di Natale, M.; Masullo, T.; Monastero, C.; Ragusa, M.A.; Tagliavia, M.; Cuttitta, A. Characterization of Translationally Controlled Tumour Protein from the Sea Anemone Anemonia viridis and Transcriptome Wide Identification of Cnidarian Homologues. Genes 2018, 9, 30. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Giannakakou, P.; Gussio, R.; Nogales, E.; Downing, K.H.; Zaharevitz, D.; Bollbuck, B.; Poy, G.; Sackett, D.; Nicolaou, K.C.; Fojo, T. A common pharmacophore for epothilone and taxanes: Molecular basis for drug resistance conferred by tubulin mutations in human cancer cells. Proc. Natl. Acad. Sci. USA 2000, 97, 2904–2909. [Google Scholar] [CrossRef] [Green Version]
- Romancino, D.P.; Anello, L.; Lavanco, A.; Buffa, V.; Di Bernardo, M.; Bongiovanni, A. A sea urchin in vivo model to evaluate Epithelial-Mesenchymal Transition. Dev. Growth Differ. 2017, 59, 141–151. [Google Scholar] [CrossRef]
- Nasuchon, N.; Hirasaka, K.; Yamaguchi, K.; Okada, J.; Ishimatsu, A. Effects of elevated carbon dioxide on contraction force and proteome composition of sea urchin tube feet. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 21, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Ragusa, M.A.; Costa, S.; Cuttitta, A.; Gianguzza, F.; Nicosia, A. Coexposure to sulfamethoxazole and cadmium impairs development and attenuates transcriptional response in sea urchin embryo. Chemosphere 2017, 180, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Pinsino, A.; Bergami, E.; Della Torre, C.; Vannuccini, M.L.; Addis, P.; Secci, M.; Dawson, K.A.; Matranga, V.; Corsi, I. Amino-modified polystyrene nanoparticles affect signalling pathways of the sea urchin (Paracentrotus lividus) embryos. Nanotoxicology 2017, 11, 201–209. [Google Scholar] [CrossRef]
- Martino, C.; Bonaventura, R.; Byrne, M.; Roccheri, M.; Matranga, V. Effects of exposure to gadolinium on the development of geographically and phylogenetically distant sea urchins species. Mar. Environ. Res. 2017, 128, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Bonaventura, R.; Russo, R.; Zito, F.; Matranga, V. Combined Effects of Cadmium and UVB Radiation on Sea Urchin Embryos: Skeleton Impairment Parallels p38 MAPK Activation and Stress Genes Overexpression. Chem. Res. Toxicol. 2015, 28, 1060–1069. [Google Scholar] [CrossRef]
- Bonaventura, R.; Matranga, V. Overview of the molecular defense systems used by sea urchin embryos to cope with UV radiation. Mar. Environ. Res. 2017, 128, 25–35. [Google Scholar] [CrossRef]
- Chiarelli, R.; Agnello, M.; Bosco, L.; Roccheri, M.C. Sea urchin embryos exposed to cadmium as an experimental model for studying the relationship between autophagy and apoptosis. Mar. Environ. Res. 2014, 93, 47–55. [Google Scholar] [CrossRef]
- Anello, L.; Cavalieri, V.; Di Bernardo, M. Developmental effects of the protein kinase inhibitor kenpaullone on the sea urchin embryo. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2018, 204, 36–44. [Google Scholar] [CrossRef]
- Semenova, M.N.; Demchuk, D.V.; Tsyganov, D.V.; Chernysheva, N.B.; Samet, A.V.; Silyanova, E.A.; Kislyi, V.P.; Maksimenko, A.S.; Varakutin, A.E.; Konyushkin, L.D.; et al. Sea Urchin Embryo Model As a Reliable in Vivo Phenotypic Screen to Characterize Selective Antimitotic Molecules. Comparative evaluation of Combretapyrazoles, -isoxazoles, -1,2,3-triazoles, and -pyrroles as Tubulin-Binding Agents. ACS Comb. Sci. 2018, 20, 700–721. [Google Scholar] [CrossRef]
- Chernysheva, N.B.; Maksimenko, A.S.; Andreyanov, F.A.; Kislyi, V.P.; Strelenko, Y.A.; Khrustalev, V.N.; Semenova, M.N.; Semenov, V.V. Regioselective synthesis of 3,4-diaryl-5-unsubstituted isoxazoles, analogues of natural cytostatic combretastatin A4. Eur. J. Med. Chem. 2018, 146, 511–518. [Google Scholar] [CrossRef]
- Eurtivong, C.; Semenov, V.; Semenova, M.; Konyushkin, L.; Atamanenko, O.; Reynisson, J.; Kiselyov, A. 3-Amino-thieno[2,3-b]pyridines as microtubule-destabilising agents: Molecular modelling and biological evaluation in the sea urchin embryo and human cancer cells. Bioorg. Med. Chem. 2017, 25, 658–664. [Google Scholar] [CrossRef]
- Oliveri, P.; Davidson, E.H.; McClay, D.R. Activation of pmar1 controls specification of micromeres in the sea urchin embryo. Dev. Biol. 2003, 258, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Stepicheva, N.A.; Song, J.L. High Throughput Microinjections of Sea Urchin Zygotes. J. Vis. Exp. 2014. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
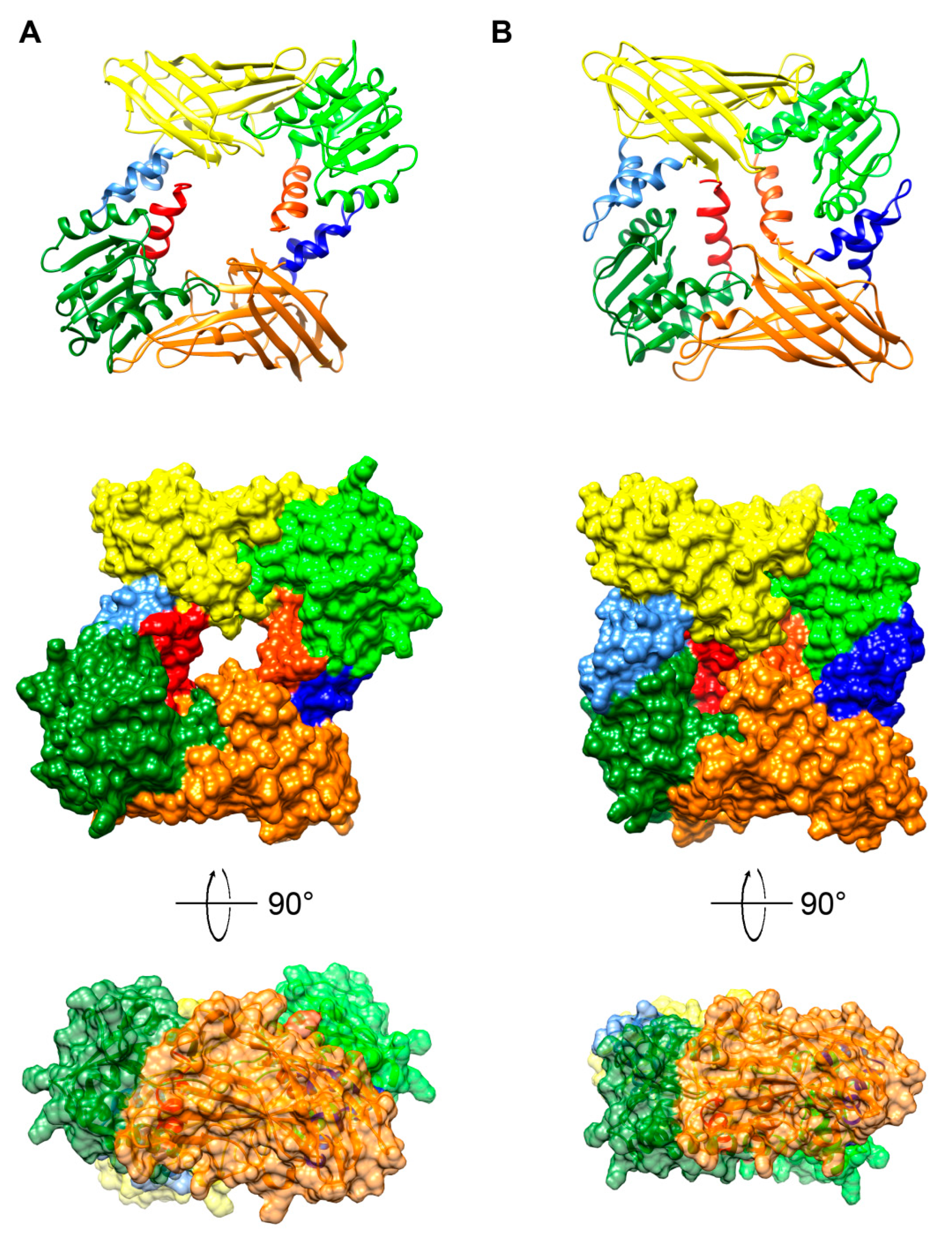{kind=link}
{kind=link}
| mRNA Length (Kb) | ORF Length (nt) | Protein Length (aa) | Molecular Weight a | pI b | |
|---|---|---|---|---|---|
| Atub8 | 1.9 | 1359 | 452 | 50,189.55 | 4.90 |
| Tuba1f | 1.7 | 1359 | 452 | 50,186.66 | 4.90 |
| Tuba1e | 1.7 | 1356 | 451 | 50,087.57 | 4.93 |
| Tuba1g | 1.8 | 1359 | 452 | 50,206.67 | 4.90 |
| Tuba1h | 2.0 | 1359 | 452 | 50,208.64 | 4.90 |
| Tuba1a | 1.5 | 1359 | 452 | 50,192.64 | 4.90 |
| Atub3 | 1.5 | 1356 | 451 | 50,212.59 | 4.91 |
| Tuba1d | 1.6 | 1359 | 452 | 50,243.73 | 4.88 |
| Tuba3 | 2.4 | 1362 | 453 | 50,407.98 | 4.83 |
| Tuba1b_1 | 1.7 | 1359 | 452 | 50,353.76 | 4.97 |
| Btub2 | 1.9 | 1344 | 447 | 50,051.16 | 4.73 |
| Tubb2a | 1.8 | 1344 | 447 | 50,051.16 | 4.73 |
| Btub3 | 1.7 | 1344 | 447 | 50,033.13 | 4.73 |
| Btub5 | 1.9 | 1341 | 446 | 49,990.17 | 4.79 |
| Btub6 | 1.8 | 1341 | 446 | 49,963.08 | 4.74 |
| Btub9 | 2.0 | 1341 | 446 | 50,000.08 | 4.72 |
| Btub4 | 1.9 | 1338 | 445 | 49,837.91 | 4.76 |
| Core Promoter Elements | Expression | ||||||
|---|---|---|---|---|---|---|---|
| Gene Name | BRE | TATA | INR | Egg | Early Blastula | Late Blastula | Prism |
| Atub8 | + | + | ++ | + | + | ||
| Tuba1f | + | - | - | + | - | ||
| Tuba1e | + | + | - | - | - | ++ | |
| Tuba1g | + | + | +++ | ++ | ++ | ||
| Tuba1h | + | + | ++ | + | + | ||
| Tuba1a | + | + | - | + | ++ | ++ | |
| Atub3 | + | + | - | - | - | + | |
| Tuba1d | + | + | - | - | - | + | |
| Tuba3 | + | + | - | - | ++ | + | |
| Tuba1b_1 | + | + | + | - | - | ++ | |
| Btub2 | + | + | - | ++ | - | +++ | |
| Tubb2a | + | - | +++ | ++ | ++ | ||
| Btub3 | + | - | ++ | - | +++ | ||
| Btub5 | + | - | + | ++ | ++ | ||
| Btub6 | + | + | - | - | - | ++ | |
| Btub9 | + | + | +++ | ++ | + | + | |
| Btub4 | + | + | - | - | - | + | |
| mRNA Length (Kb) | ORF Length (nt) | Protein Length (aa) | Molecular Weight | pI | |
|---|---|---|---|---|---|
| PRMT1 | 1.6 | 1077 | 358 | 41,123.81 | 5.33 |
| PRMT2 | 3.0 | 1353 | 450 | 50,762.22 | 5.06 |
| PRMT3 | 2.2 | 1647 | 548 | 62,149.50 | 4.60 |
| PRMT4 | 3.4 | 1185 | 394 | 68,586.26 | 6.77 |
| PRMT5 | 2.5 | 1881 | 626 | 71,161.21 | 6.16 |
| PRMT6 | 2.4 | 1854 | 617 | 44,301.14 | 5.04 |
| PRMT7 | 3.1 | 2061 | 686 | 77,028.46 | 5.55 |
| Expression | ||||
|---|---|---|---|---|
| Gene Name | Egg | Early Blastula | Late Blastula | Prism |
| PRMT1 | +++ | ++++ | ++++ | ++ |
| PRMT2 | + | + | + | - |
| PRMT3 | - | + | ++ | + |
| PRMT4 | + | + | + | - |
| PRMT5 | + | + | + | + |
| PRMT6 | - | + | + | - |
| PRMT7 | + | ++ | - | - |
| Experimentally Defined Methylated Residues | Predicted Methylable P. lividus Residues | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Site 1 | Type 2 | Tissue | Reference 3 | Protein Name | Accession # | −6/+6 Peptide 4 | P. lividus Proteins | −6/+6 Peptide 4 | P. lividus Protein | −6/+6 Peptide 4 |
| α339 | MMA | HCT116 cells | [77] | TUBA1B | NP_006073 | ATIKTKrSIQFVD | ALL 6 | ATIKTKrTIQFVD | Atub8 | ATIKTKrSIQFVD |
| α339 | MMA | HCT116 cells | [77] | TUBA4A | NP_005991 | AAIKTKrSIQFVD | Atub3 | ANIKTKrTIQFVD | ||
| α338 | MMA | 2,9 F 5 | [77] | TUBA3C | XP_486246 | ATIKTKrTIQFVD | ||||
| α339 | MMA | 2,9 F | [77] | TUBA1B | P05213 | ATIKTKrSIQFVD | ||||
| α339 | MMA | 2,9 F | [77] | TUBA4A | P68368 | AAIKTKrSIQFVD | ||||
| β46 | MMA | 2,0 F | [77] | TUBB2A | Q7TMM9 | SDLQLErINVYYN | ALL | SDLQLErINVYYN | ||
| β46 | MMA | 2,0 F | [77] | TUBB2B | Q9CWF2 | SDLQLErINVYYN | ||||
| β62 | Neuro2a cells | [22] | TUBB3 | Q13509 | SHKYVPrAILVDL | ALL 6 | GGKYVPrAVLVDL | Btub6 | GGKYVPrAALVDL | |
| β86 | MMA | 2,7 F | [77] | TUBB2A | Q7TMM9 | PFGQIFrPDNFVF | ALL 6 | PFGQIFrPDNFVF | Btub4 | PFGQIYrPDNFVF |
| β86 | MMA | 2,7 F | [77] | TUBB2B | Q9CWF2 | PFGQIFrPDNFVF | Btub6 | PFGQIFrPDNFIF | ||
| β86 | MMA | 2,7 F | [77] | TUBB2C | P68372 | PFGQIFrPDNFVF | ||||
| β86 | MMA | 2,7 F | [77] | TUBB4 | Q9D6F9 | PFGQIFrPDNFVF | ||||
| β86 | MMA | 2,7 F | [77] | TUBB | P99024 | PFGQIFrPDNFVF | ||||
| β162 | ADMA | mouse brain | [77] | TUBB | NP_035785 | REEYPDrIMNTFS | ALL 6 | REEYPDrIMNTFS | Btub4 | REEYPDrVMNTFS |
| β282 | Neuro2a cells | [76] | TUBB3 | Q13509 | RGSQQYrALTVPE | ALL | RGSQQYrALTVPE | |||
| β318 | MMA | HCT116 cells | [77] | TUBB2C | NP_006079 | TVAAVFrGRMSMK | ALL 6 | TVAAIFrGRMSMK | Btub5 | TVAAMFrGRMSMK |
| β318 | ADMA | mouse brain | [77] | TUBB | NP_035785 | TVAAVFrGRMSMK | ||||
| β318 | DIMETH | S. cerevisiae | [76] | YFL037W | P02557 | TVAAFFrGKVSVK | ||||
| β320 | ADMA | mouse brain | [77] | TUBB | NP_035785 | AAVFRGrMSMKEV | ALL 6 | AAIFRGrMSMKEV | Btub5 | AAMFRGrMSMKEV |
| β380 | MMA | HCT116 cells | [77] | TUBB8 | NP_817124 | IQELFKrVSEQFT | ALL | IQELFKrISEQFT | ||
| α79 | MMA | root tissue 7 | [78] | α tubulin | XP_010044940 | TV(I/V)DEVRSGTYRQ | ||||
| β162 | MMA | root tissue 7 | [78] | β tubulin 5 | NP_001289666 | REEYPDRMMLTFS | ||||
| Target | Forward Primer - 3′RACE | Reverse Primer- 5′RACE | Ta 1 |
|---|---|---|---|
| Atub8 | TATACCAACTTGAACCGTC | AGATGAGAAATCCTTGGAGA | 48 |
| Tuba1f | ATCTATGATATATGCCGTCG | GACATGGACTGAGATACATT | 47 |
| Tuba1e | ATTACGGAAAGAAGTCCAAG | ACTGAAGAAGGTGTTAAAGG | 48 |
| Atub3 | CTGTTGTCGAGCCATATAA | GCATAGTTATTAGCAGCATC | 47 |
| Tuba1d | CTGTAGTTGAGCCCTATAAC | ACTGAGATACATTCCCTCAT | 48 |
| Tuba3 | GGAGAAGGACTATGAAGAAG | AAGGATTGAGTTGTATGGTT | 47 |
| Tuba1b_1 | GAATCCATTTCCCTCTTGTA | TATCCATGACTCTGTCAATG | 47 |
| Btub6 | GATATCTGTTTCCGTACCTT | CTGTAGCCTCATTGTAGTAG | 47 |
| Btub9 | CCTGAATCATCTCGTATCAG | CACGCATAATGATACAGATG | 47 |
| Btub4 | AGCTCTGTACGATATTTGTT | CAGCTTGTAGATGAACAATC | 47 |
| PRMT2 | ATAGAAGGTACAACCCTACC | ATTGCTTCCATCTCCATTAC | 48 |
| PRMT3 | CAAGGATAAGGTGGTGTTAG | TGGAGAGGAGAAGATTTCAT | 48 |
| PRMT4 | TGAGCAGGTTGATATCATTG | CTTTGTGTTTAGTGTGTGTG | 48 |
| PRMT5 | CAAATGTGCTATCTTCTCCA | TTTCCTTCTACGAACTCTCT | 48 |
| PRMT6 | AGTCCTCATTTACATTCAGC | CCTAGGAGGCTTAGTAGAAG | 48 |
| PRMT7 | CTACATATCCACATGCTCAC | AGGGGATGTTCACCATATTA | 48 |
| Name | Forward Primer | Reverse Primer | Ta 1 |
|---|---|---|---|
| Atub8 | CGGCACATCGGACACTGTGA | AGCCGTGGGCGTTGATTGAT | 58 |
| Tuba1f | TGTAGCGAGCGACTCAAGCG | TGAGCAAAAGGAGGCTACGGC | 58 |
| Tuba1e | ACCCATCCATCACTTGGCACG | TCCCCACATTTTGGCGAATGA | 55 |
| Atub3 | CGACGTCTGCTAGCTCACCTTA | TGGTTTATTGATCAGCTCTCATGGC | 56 |
| Tuba1d | GACGCTTCGCAGCTCTGTCT | TGCATCGAAGGAAGGGGGAT | 56 |
| Tuba3_UP | GTCTCGCCGATTTCGCCACT | TGTGCTTGCCAGCTCCAGTC | 58 |
| Tuba3_DW | GCGAACCGGTACGTATCGTCA | TTCATCGTAGAATCTTGGAACGCC | 56 |
| Tuba1b_1 | CGCATCGAACACGGCTCTGA | GGTATCGTGGCCGTGCGAT | 58 |
| Btub6 | CCTTCTAAGCGAATTGCAAGGTG | TCTGCTTGTACATGCTGCCAGA | 55 |
| Btub9 | GACGGTTGCACAGCATGCAC | GTCCGGACGCAAGAGTGGTC | 58 |
| Btub4 | GGTCTGCTTGTGTGTCCCCG | TAACGAGTCCGACCAGGGGG | 58 |
| Target | Forward Primer - 3′RSO | Reverse Primer- 5′RSO | Ta 1 |
|---|---|---|---|
| Atub8 | TTATAAGGCAACCTCCAACC | GTTTCACAGATTGCTCACAG | 50 |
| Tuba1f | ACAACGAACTAAAGGCTCAT | GTATGATGTGCCGTAAGCTA | 50 |
| Tuba1e | GCCAAAATGTGGGGATTAAG | TAGACGGATTTGAGGCAAAA | 50 |
| Atub3 | TAGAATTCAGCCATGAGAGC | TTTGAGGAGCTTTGACAACT | 50 |
| Tuba1d | AATCGTCAACAGTTTGCTTC | TATTGAAGAGACAGAGCTGC | 50 |
| Tuba3 | TTCTGACTTCTTAGTGCCTT | AGTAGAAGTGGCGAAATCGG | 50 |
| Tuba1b_1 | AGAGGAAGATGACGATTGTG | ATAGGTTAGGCTTCAACAGC | 50 |
| Btub6 | CACTTGAAAGGAACATCTGC | AATAATGCACAGGAGAGGTG | 50 |
| Btub9 | TTCCAAAACATTTGCCTGTC | AATGGCGGAAAAATTGTGTT | 50 |
| Btub4 | CGACCTGTGGATACATCATT | AGATGTAAAGAAACGGGGAC | 50 |
| RSO-Bam | TAATACGACTCACTATAGGGAGANNNNNNNNNNGGATCC | ||
| RSO-Eco | TAATACGACTCACTATAGGGAGANNNNNNNNNNGAATTC | ||
| RSO-Nde | TAATACGACTCACTATAGGGAGANNNNNNNNNNCATATG | ||
| RSO-Xba | TAATACGACTCACTATAGGGAGANNNNNNNNNNTCTAGA | ||
| RSO-Sau | TAATACGACTCACTATAGGGAGANNNNNNNNNNGATC | ||
| RSO-Taq | TAATACGACTCACTATAGGGAGANNNNNNNNNNTCGA | ||
| RSO_T7 | TAATACGACTCACTATAGGGAGA | 50 | |
| Name | Forward Primer | Reverse Primer |
|---|---|---|
| Atub8 | TGAGCAATCTGTGAAACCTCCTC | TACAACTCCCAGCAGGCATTACC |
| Tuba1f | CTTACGGCACATCATACGTTGC | CGACATGGACTGAGATACATTCCC |
| Tuba1e | CTGAGCATTTTGCCTCAAATCC | TCCGACGTGGATAGAGATACATTC |
| Atub3 | CAGTGCAGTTGTCAAAGCTCCTC | GGCTGGATACCATGCTCAAGAC |
| Tuba1d | AAAGCTCACTTCAAGACGCTTCG | CGACATGGACTGAGATACATTCCC |
| Tuba3 | GCCGATTTCGCCACTTCTACTTAG | AGGCATTTCCCATCTGGACAC |
| Tuba1b_1 | TCTTCGTTGCTGTTGAAGCC | TCAAGGCAATAGAGTTCCCAGC |
| Btub6 | CCTAAGCAAATTGCAGGGTGTAAC | GCCTGTAAATGTACAATCTCACGC |
| Btub9 | CCACGAACACAATTTTTCCG | CCAGCTTGTAAGTGAACAATCTCAC |
| Btub4 | TTTCCCAAAGAGTCGTGGTG | CCAGCTTGTAGATGAACAATCTCAC |
| PRMT1 | GGGAGGACAGGGGGACGGAC | CAGCCCCAGCTTTGGCAGCA |
| PRMT2 | GCCTGGCGGAAATGGGGGAG | TTCCCTTGCGTGCCCACCAC |
| PRMT3 | CTGCTCACCATGGGCGCTGC | AGCCTTCCAATCGGTTTGCGTGT |
| PRMT4 | ACAGCAAGGCAGGGTGGTGC | GCATACATGCCCCCAGCCCC |
| PRMT5 | GTCCGCAGCCGGAGCGTATG | GGGCATCGAGGGCACCATCA |
| PRMT6 | GGCAGAGCCAGAGCCTGTTGG | GCCGCAATCTCTTCCGCACCT |
| PRMT7 | TCGCTGGCGCCTGGAGTACA | CCTGGGCCTTGAGAATGCAGGG |
| 18S rRNA | GAATGTCTGCCCTATCAACTTTCG | TTGGATGTGGTAGCCGTTTCTC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragusa, M.A.; Nicosia, A.; Costa, S.; Casano, C.; Gianguzza, F. A Survey on Tubulin and Arginine Methyltransferase Families Sheds Light on P. lividus Embryo as Model System for Antiproliferative Drug Development. Int. J. Mol. Sci. 2019, 20, 2136. https://doi.org/10.3390/ijms20092136
Ragusa MA, Nicosia A, Costa S, Casano C, Gianguzza F. A Survey on Tubulin and Arginine Methyltransferase Families Sheds Light on P. lividus Embryo as Model System for Antiproliferative Drug Development. International Journal of Molecular Sciences. 2019; 20(9):2136. https://doi.org/10.3390/ijms20092136
Chicago/Turabian StyleRagusa, Maria Antonietta, Aldo Nicosia, Salvatore Costa, Caterina Casano, and Fabrizio Gianguzza. 2019. "A Survey on Tubulin and Arginine Methyltransferase Families Sheds Light on P. lividus Embryo as Model System for Antiproliferative Drug Development" International Journal of Molecular Sciences 20, no. 9: 2136. https://doi.org/10.3390/ijms20092136
APA StyleRagusa, M. A., Nicosia, A., Costa, S., Casano, C., & Gianguzza, F. (2019). A Survey on Tubulin and Arginine Methyltransferase Families Sheds Light on P. lividus Embryo as Model System for Antiproliferative Drug Development. International Journal of Molecular Sciences, 20(9), 2136. https://doi.org/10.3390/ijms20092136