Mitogen Activated Protein Kinases in Steatotic and Non-Steatotic Livers Submitted to Ischemia-Reperfusion

 and
and
Abstract
1. Introduction: Ischemia-Reperfusion, MAPKs and NAFLD patients
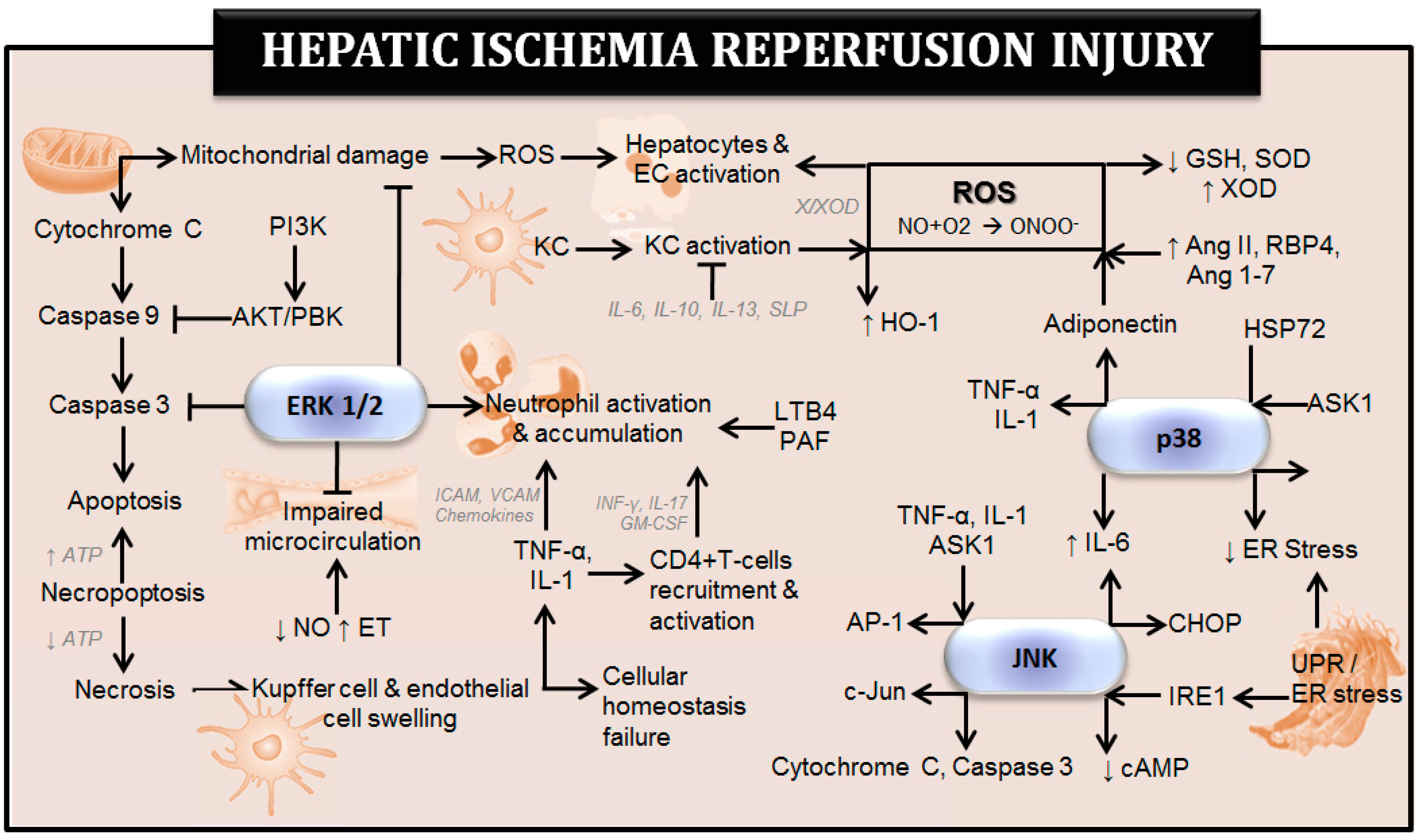
2. Role of MAPKs in Experimental Models of Hepatic I-R Injury Using Steatotic and Non-Steatotic Livers
2.1. p38 in Hepatic I-R and Their Therapeutic Implications
2.1.1. p38 in Hepatic I-R
2.1.2. Therapeutic Implications of p38 in Hepatic I-R
2.2. JNK in Hepatic I-R and Their Therapeutic Implications
2.2.1. JNK in Hepatic I-R
2.2.2. Therapeutic Implications of JNK in Hepatic I-R
2.3. ERK in Hepatic I-R and Their Therapeutic Implications
2.3.1. ERK in Hepatic I-R
2.3.2. Therapeutic Implications of ERK in Hepatic I-R
3. Role of MAPKs in the Clinical Practice of Hepatic I-R Injury Using Steatotic and Non-Steatotic Livers
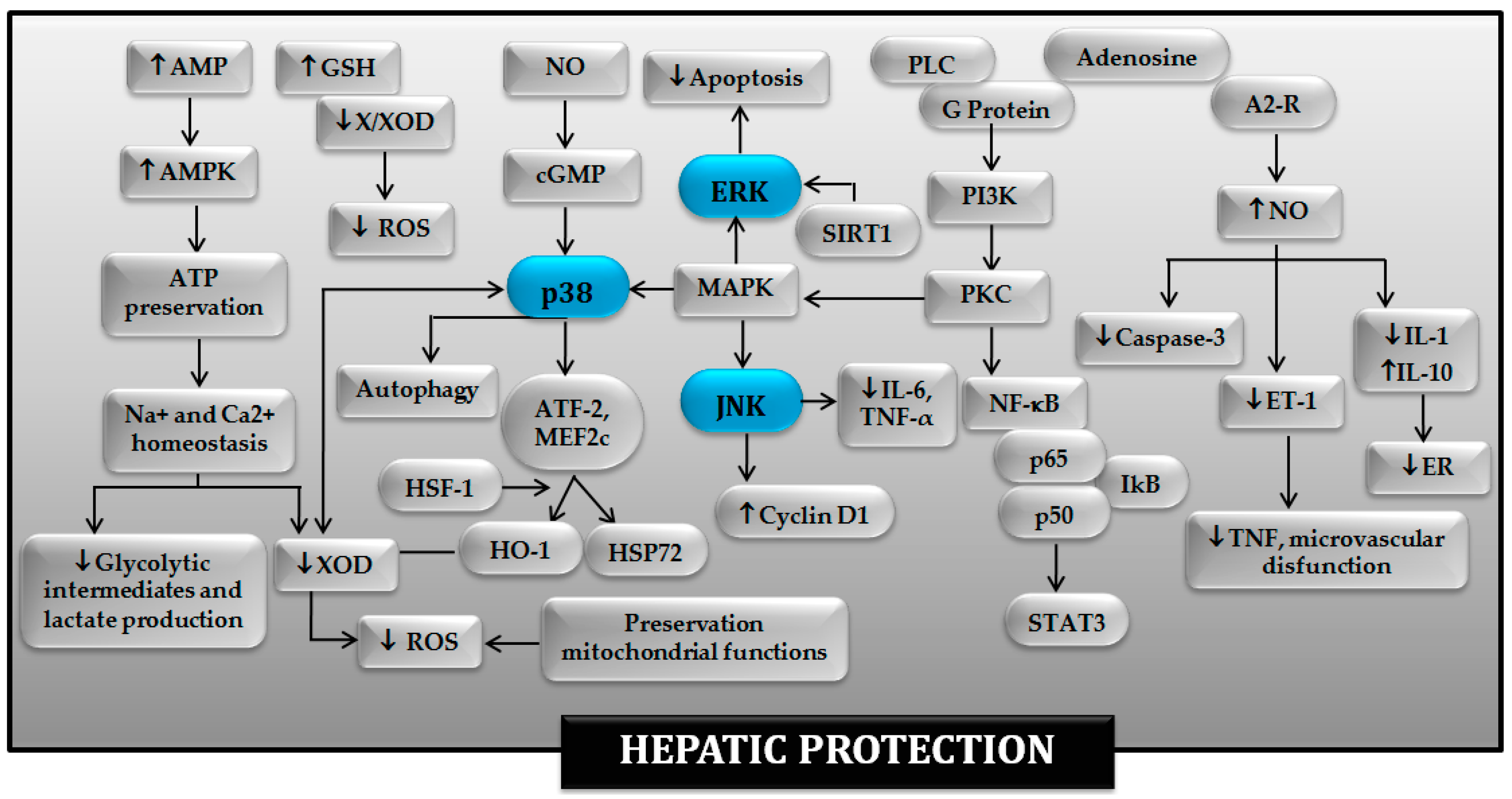
4. Involvement of MAPKs in the Benefits of PC in Hepatic I-R
5. Conclusions and Perspectives
Funding
Conflicts of Interest
Abbreviations
| A2-R | Adenosine 2 receptor |
| ACH | Acetylcholine |
| AGE | Advanced glycation endproduct |
| LD | Linear dichroism |
| Akt | Protein kinase B |
| AMP | AMP Adenosine monophosphate |
| Ang | Angiotensin |
| ANP | Atrial natriuretic peptide |
| AP-1 | Activating protein |
| ASK1 | Apoptosis signal-regulating kinase 1 |
| AST | Aspartate aminotransferase |
| ATF-2 | Activating transcription factor 2 |
| ATP | Adenosine triphosphate |
| Bcl-2 | B-cell lymphoma 2 |
| cAMP | Cyclic adenosine monophosphate |
| cGMP | Guanosine 3’,5’-cyclic monophosphate |
| CO | Carbon monoxide |
| CS-1 | Connecting segment-1 peptide |
| CXCL10 | C-X-C motif chemokine ligand 10 |
| DCD | Donation after circulatory death |
| EGF | Epidermal growth factor |
| eNOS | Endothelial nitric oxide synthase |
| ER | Endoplasmic reticulum |
| ERK 1/2 | Extracellular signal-regulated protein kinases |
| ET-1 | Endothelin-1 |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| GSH | Glutathione |
| HFD | High fat diet |
| HMP | Hypothermic machine perfusion |
| HO-1 | Heme oxygenase-1 |
| HSF-1 | Heat shock transcription factor-1 |
| HSP | Heat shock protein |
| I-R | Ischemia-reperfusion |
| ICAM | Intracellular cell adhesion molecule |
| IGF | Insulin-like growth factor |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| i.p | Intraperitoneal |
| i.v | Intravenous |
| JNK | c-Jun N-terminal kinase |
| KO | Knockout |
| LDH | Lactate dehydrogenase |
| LTB4 | Leucotriene B4 |
| MAPKs | Mitogen-activated protein kinases |
| MEF2c | Myocyte enhancer factor-2 |
| Min | Minutes |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NAFLD | Non-alcoholic fatty liver disease |
| NASH | Non-alcoholic steatohepatitis |
| NDQI-1 | NQDI-1–2,7-Dihydro-2,7-dioxo-3H-naphth-o[1,2,3dequin-oline-1-carboxylic acid ethyl ester] |
| NF-κB | Nuclear factor κ Beta |
| NO | Nitric oxide |
| PAF | Platelet activating factor |
| PC | Ischemic preconditioning |
| PDGF | Platelet-derived growth factor |
| PI3K | Phosphoinositide 3-kinase |
| PKC | Protein kinase C |
| PLC | Phospholipase C |
| RAGE | Receptor advanced glycation endproduct |
| RBP4 | Retinol binding protein 4 |
| ROS | Reactive oxygen species |
| SCS | Simple cold storage |
| SIRT1 | Sirtuin-1 |
| SLPI | Secretory leukocyte protease inhibitor |
| SOD | Superoxide dismutase |
| STAT3 | Signal transducer and activator of transcription-3 |
| Thr | Threonine |
| TNF-α | Tumor necrosis factor alpha |
| TUDCA | Tauroursodeoxycholic acid |
| Tyr | Tyrosine |
| X/XOD | Xanthine/xanthine oxidase |
| Z-Asp-cmk | Z-Asp-2,6-dichorobenzoyloxymethylketone |
References
- Casillas-Ramírez, A.; Mosbah, I.B.; Ramalho, F.; Roselló-Catafau, J.; Peralta, C. Past and future approaches to ischemia-reperfusion lesion associated with liver transplantation. Life Sci. 2006, 79, 1881–1894. [Google Scholar] [CrossRef]
- Serafin, A.; Rosello-Catafau, J.; Prats, N.; Xaus, C.; Gelpi, E.; Peralta, C. Ischemic preconditioning increases the tolerance of fatty liver to hepatic ischemia-reperfusion injury in the rat. Am. J. Pathol. 2002, 161, 587–601. [Google Scholar] [CrossRef]
- Clavien, P.; Harvey, P.; Strasberg, S. Preservation and reperfusion injuries in liver allografts. An overview and synthesis of current studies. Transplantation 1992, 53, 957–978. [Google Scholar] [CrossRef] [PubMed]
- Huguet, C.; Gavelli, A.; Chieco, P.; Bona, S.; Harb, J.; Joseph, J.M.; Jobard, J.; Gramaglia, M.; Lasserre, M. Liver ischemia for hepatic resection: Where is the limit? Surgery 1992, 111, 251–259. [Google Scholar] [PubMed]
- Busuttil, R.W.; Tanaka, K. The utility of marginal donors in liver transplantation. Liver Transpl. 2003, 9, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. Nonalcoholic fatty liver disease as a nexus of metabolic and hepatic diseases. Cell Metab. 2018, 27, 22–41. [Google Scholar] [CrossRef]
- Pratap, A.; Panakanti, R.; Yang, N.; Lakshmi, R.; Modanlou, K.A.; Eason, J.D.; Mahato, R.I. Cyclopamine attenuates acute warm ischemia reperfusion injury in cholestatic rat liver: Hope for marginal livers. Mol. Pharm. 2011, 8, 958–968. [Google Scholar] [CrossRef]
- Nadig, S.N.; Periyasamy, B.; Shafizadeh, S.F.; Polito, C.; Fiorini, R.N.; Rodwell, D.; Evans, Z.; Cheng, G.; Dunkelberger, D.; Schmidt, M.; et al. Hepatocellular ultrastructure after ischemia/reperfusion injury in human orthotopic liver transplantation. J. Gastrointest. Surg. 2004, 8, 695–700. [Google Scholar] [CrossRef]
- Verran, D.; Kusyk, T.; Painter, D.; Fisher, J.; Koorey, D.; Strasser, S.; Stewart, G.; McCaughan, G. Clinical experience gained from the use of 120 steatotic donor livers for orthotopic liver transplantation. Liver Transpl. 2003, 9, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Schaubel, D.E.; Sima, C.S.; Goodrich, N.P.; Feng, S.; Merion, R.M. The survival benefit of deceased donor liver transplantation as a function of candidate disease severity and donor quality. Am. J. Transplant. 2008, 8, 419–425. [Google Scholar]
- Ploeg, R.J.; D’ Alessandro, A.M.; Knechtle, S.J.; Stegall, M.D.; Pirsch, J.D.; Hoffmann, R.M.; Sasaki, T.; Sollinger, H.W.; Belzer, F.O.; Kalayoglu, M. Risk factors for primary dysfunction after liver transplantation-a multivariate analysis. Transplantation 1993, 55, 807–813. [Google Scholar] [CrossRef]
- Behrns, K.E.; Tsiotos, G.G.; DeSouza, N.F.; Krishna, M.K.; Ludwig, J.; Nagorney, D.M. Hepatic steatosis as a potential risk factor for major hepatic resection. J. Gastrointest. Surg. 1998, 2, 292–298. [Google Scholar] [CrossRef]
- Lopez-Neblina, F.; Toledo, A.H.; Toledo-Peryra, L.H. Molecular biology of apoptosis in ischemia and reperfusion. J. Investig. Surg. 2005, 18, 335–350. [Google Scholar] [CrossRef]
- King, L.A.; Toledo, A.H.; Rivera-Chavez, F.A.; Toledo-Pereyra, L.H. Role of p38 and JNK in liver ischemia and reperfusion. J. Hepatobiliary Pancreat. Surg. 2009, 16, 763–770. [Google Scholar] [CrossRef]
- Kobayashi, M.; Takeyoshi, I.; Yoshinari, D.; Matsumoto, K.; Morishita, Y. The role of mitogen-activated protein kinases and the participation of intestinal congestion in total hepatic ischemia–reperfusion injury. Hepatogastroenterology 2006, 53, 243–248. [Google Scholar]
- Kumar, S.; Boehm, J.; Lee, J.C. p38 MAP Kinases: Key signalling molecules as therapeutic targets for inflammatory diseases. Nat. Rev. Drug Discov. 2003, 2, 717–725. [Google Scholar] [CrossRef]
- Schnabl, B.; Bradham, C.A.; Bennett, B.L.; Manning, A.M.; Stefanovic, B.; Brenner, D.A. TAK1/JNK and p38 have opposite effects on rat hepatic stellate cells. Hepatology 2001, 34, 953–963. [Google Scholar] [CrossRef]
- Toledo-Pereyra, L.H.; Toledo, A.H.; Walsh, J.; Lopez-Neblina, F. Molecular signaling pathways in ischemia/reperfusion. Exp. Clin. Transplant. 2004, 2, 174–177. [Google Scholar]
- Bradham, C.A.; Stachlewitz, R.F.; Gao, W.; Qian, T.; Jayadev, S.; Jenkins, G.; Hannun, Y.; Lemasters, J.J.; Thurman, R.G.; Brenner, D.A. Reperfusion after liver transplantation in rats differentially activates the mitogen-activated protein kinases. Hepatology 1997, 25, 1128–1135. [Google Scholar] [CrossRef]
- Ding, C.; Wilding, J.P.; Bing, C. 1,25-dihydroxyvitamin D3 protects against macrophage-induced activation of NFκB and MAPK signaling and chemokine release in human adipocytes. PLoS ONE 2013, 8, e61707. [Google Scholar]
- Mutt, S.J.; Karhu, T.; Lehtonen, S.; Carlberg, C.; Saarnio, J.; Sebert, S.; Hypponen, E.; Jarvelin, M.R.; Herzing, K.H. Inhibition of cytokine secretion from adipocytes by 1,25-dihydroxyvitamin D via the NF-κB pathway. FASEB J. 2012, 26, 4400–4407. [Google Scholar] [CrossRef]
- Meeker, S.; Seamons, A.; Paik, J.; Treuting, P.M.; Brabb, T.; Grady, W.M.; Maggio-prince, L. Increased dietary vitamin D suppresses MAPK signaling, colitis, and colon cancer. Cancer Res. 2014, 74, 4398–4408. [Google Scholar] [CrossRef]
- Nelson, J.E.; Roth, C.L.; Wilson, L.A.; Yates, K.P.; Aouizerat, B.; Morgan-Stevenson, V.; Whalen, E.; Hoofnagle, A.; Mason, M.; Gersuk, V.; et al. Vitamin D deficiency is associated with increased risk of non-alcoholic steatohepatitis in adults with non-alcoholic fatty liver disease: Possible role for MAPK and NF-κB? Am. J. Gastroenterol. 2016, 111, 852–863. [Google Scholar] [CrossRef]
- Vega, M.I.; Huerta-Yepaz, S.; Garban, H.; Jazirehi, A.; Emmanouilides, C.; Bonavida, B. Rituximab inhibits p38 MAPK activity in 2F7 B NHL and decreases IL-10 transcription: Pivotal role of p38 MAPK in drug resistance. Oncogene 2004, 23, 3530–3540. [Google Scholar] [CrossRef]
- Hu, X.; Chen, J.; Wang, L.; Ivashkiv, L.B. Crosstalk among Jak-STAT, Toll-like receptor, and ITAM-dependent pathways in macrophage activation. J. Leukocyte Biol. 2007, 82, 237–243. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Karrar, A.; Pierobon, M.; Birerdinc, A.; Stepanova, M.; Abdelatif, D.; Younoszai, Z.; Jeffers, T.; Felix, S.; Jeiran, K.; et al. An exploratory study examining how nano-liquid chromatography-mass spectrometry and phosphoproteomics can differentiate patients with advanced fibrosis and higher percentage collagen in non-alcoholic fatty liver disease. BMC Med. 2018, 16, 170. [Google Scholar] [CrossRef]
- Wu, Q.; Li, S.; Li, X.; Sui, Y.; Yang, Y.; Dong, L.; Xie, B.; Sun, Z. Inhibition of advanced glycation end product formation by lotus seedpod oligomeric procyanidins through RAGE-MAPK signaling and NF-κB activation in high-fat-diet rats. J. Agric. Food Chem. 2015, 63, 6989–6998. [Google Scholar] [CrossRef]
- Banga, N.R.; Homer-Vanniasinkam, S.; Graham, A.; Al-Mukhtar, A.; White, S.A.; Prasad, K.R. Ischaemic preconditioning in transplantation and major resection of the liver. Br. J. Surg. 2005, 92, 528–538. [Google Scholar] [CrossRef]
- Fan, C.; Zwacka, R.M.; Engelhardt, J.F. Therapeutic approaches for ischemia/reperfusion injury in the liver. J. Mol. Med. 1999, 77, 577–592. [Google Scholar] [CrossRef]
- Jaeschke, H. Molecular mechanisms of hepatic ischemia-reperfusion injury and preconditioning. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G15–G26. [Google Scholar] [CrossRef]
- Lentsch, A.B.; Kato, A.; Yoshidome, H.; McMasters, K.M.; Edwards, M.J. Inflammatory mechanisms and therapeutic strategies for warm hepatic ischemia/reperfusion injury. Hepatology 2000, 32, 169–173. [Google Scholar] [CrossRef]
- Massip-Salcedo, M.; Roselló-Catafau, J.; Prieto, J.; Avíla, M.A.; Peralta, C. The response of the hepatocyte to ischemia. Liver Int. 2007, 27, 6–16. [Google Scholar] [CrossRef]
- Kobayashi, M.; Takeyoshi, I.; Yoshinari, D.; Matsumoto, K.; Morishita, Y. P38 mitogen-activated protein kinase inhibition attenuates ischemia–reperfusion injury of the rat liver. Surgery 2002, 131, 344–349. [Google Scholar] [CrossRef]
- Liang, R.; Nickkholgh, A.; Hoffmann, K.; Kern, M.; Schneider, H.; Sobirey, M.; Zorn, M.; Buchler, M.W.; Schemmer, P. Melatonin protects from hepatic reperfusion injury through inhibition of IKK and JNK pathways and modification of cell proliferation. J. Pineal Res. 2009, 46, 8–14. [Google Scholar] [CrossRef]
- Toledo-Pereyra, L.H.; Lopez-Neblina, F.; Toledo, A.H. Protein kinases in organ ischemia and reperfusion. J. Investig. Surg. 2008, 21, 215–226. [Google Scholar] [CrossRef]
- Tsung, A.; Hoffman, R.A.; Izuishi, K.; Critchlow, N.D.; Nakao, A.; Chan, M.H.; Lotze, M.T.; Geller, D.A.; Billiar, T.R. Hepatic ischemia/reperfusion injury involves functional TLR4 signaling in nonparenchymal cells. J. Immunol. 2005, 175, 7661–7668. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Brenner, D.A. Mechanisms of liver injury. I. TNF-α induced liver injury: Role of IKK, JNK, and ROS pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G583–G589. [Google Scholar] [CrossRef]
- Onishi, I.; Shimizu, K.; Tani, T.; Hashimoto, T.; Miwa, K. JNK activation and apoptosis during ischemia–reperfusion. Transplant. Proc. 1999, 31, 1077–1079. [Google Scholar] [CrossRef]
- Yoshinari, D.; Takeyoshi, I.; Kobayashi, M.; Koyama, T.; Iijima, K.; Ohwada, S.; Matsumoto, K.; Morishita, Y. Effects of a p38 mitogen-activated protein kinase inhibitor as an additive to university of wisconsin solution on reperfusion injury in liver transplantation. Transplantation 2001, 72, 22–27. [Google Scholar] [CrossRef]
- Massip-Salcedo, M.; Casillas-Ramirez, A.; Franco-Gou, R.; Bartrons, R.; Ben Mosbah, I.; Serafin, A.; Rosello-Catafau, J.; Peralta, C. Heat shock proteins and mitogen-activated protein kinases in steatotic livers undergoing ischemia–reperfusion: Some answers. Am. J. Pathol. 2006, 168, 1474–1485. [Google Scholar] [CrossRef]
- Rosseland, C.; Wierød, L.; Oksvold, M.P.; Werner, H.; Østvold, A.C.; Thoresen, G.H.; Paulsen, R.E.; Huitfeldt, H.S.; Skarpen, E. Cytoplasmic retention of peroxide-activated ERK provides survival in primary cultures of rat hepatocytes. Hepatology 2005, 42, 200–207. [Google Scholar] [CrossRef]
- Widmann, C.; Gibson, S.; Jarpe, M.B.; Johnson, G.L. Mitogen-activated protein kinase: Conservation of a three-kinase module from yeast to human. Physiol. Rev. 1999, 79, 143–180. [Google Scholar] [CrossRef]
- Kaminska, B. MAPK signaling pathways as molecular targets for anti-inflammatory therapy-from molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Casillas-Ramírez, A.; Zaouali, A.; Padrissa-Altés, S.; Ben Mosbah, I.; Pertosa, A.; Alfany-Fernández, I.; Bintanel-Morcillo, M.; Xaus, C.; Rimola, A.; Rodes, J.; Rosello-Catafau, J.; Peralta, C. Insulin-like growth factor and epidermal growth factor treatment: New approaches to protecting steatotic livers against ischemia-reperfusion injury. Endocrinology 2009, 150, 3153–3161. [Google Scholar] [CrossRef]
- Ramalho, F.S.; Alfany-Fernandez, I.; Casillas-Ramirez, A.; Massip-Salcedo, M.; Serafín, A.; Rimola, A.; Arroyo, V.; Rodes, J.; Rosello-Catafau, J.; Peralta, C. Are angiotensin II receptor antagonists useful strategies in steatotic and nonsteatotic livers in conditions of partial hepatectomy under ischemia-reperfusion? J. Pharmacol. Exp. Ther. 2009, 329, 130–140. [Google Scholar] [CrossRef]
- Ben Mosbah, I.; Alfany-Fernandez, I.; Martel, C.; Zaouali, M.A.; Bintanel-Morcillo, M.; Rimola, A.; Rodes, J.; Brenner, C.; Rosello-Catafau, J.; Peralta, C. Endoplasmic reticulum stress inhibition protects steatotic and non-steatotic livers in partial hepatectomy under ischemia-reperfusion. Cell Death Dis. 2010, 1, e52. [Google Scholar] [CrossRef]
- Cursio, R.; Filippa, N.; Miele, C.; Van Obberghen, E.; Gugenheim, J. Involvement of protein kinase B and mitogen-activated protein kinases in experimental normothermic liver ischemia-reperfusion injury. Br. J. Surg. 2006, 93, 752–761. [Google Scholar] [CrossRef]
- Massip-Salcedo, M.; Zaouali, M.A.; Padrissa-Altés, S.; Casillas-Ramirez, A.; Rodés, J.; Roselló-Catafau, J.; Peralta, C. Activation of peroxisome proliferator-activated receptor-alpha inhibits the injurious effects of adiponectin in rat steatotic liver undergoing ischemia-reperfusion. Hepatology 2008, 47, 461–472. [Google Scholar] [CrossRef]
- Fernández, L.; Carrasco-Chaumel, E.; Serafín, A.; Xaus, C.; Grande, L.; Rimola, A.; Roselló-Catafau, J.; Peralta, C. Is ischemic preconditioning a useful strategy in steatotic liver transplantation? Am. J. Transplant. 2004, 4, 888–889. [Google Scholar] [CrossRef]
- Alfany-Fernández, I.; Casillas-Ramírez, A.; Bintanel-Morcillo, M.; Brosnihan, K.B.; Ferrario, C.M.; Serafin, A.; Rimola, A.; Rodés, J.; Rosellò-Catafau, J.; Peralta, C. Therapeutic targets in liver transplantation: Angiotensin II in nonsteatotic grafts and angiotensin-(1-7) in steatotic grafts. Am. J. Transplant. 2009, 9, 439–451. [Google Scholar] [CrossRef]
- Keller, M.; Gerbes, A.L.; Kulhanek-Heinze, S.; Gerwig, T.; Grutzner, U.; van Rooijen, N.; Vollmar, A.M.; Kiemer, A.K. Hepatocyte cytoskeleton during ischemia and reperfusion-influence of ANP-mediated p38 MAPK activation. World J. Gastroenterol. 2005, 11, 7418–7429. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Takahara, T.; Fujino, M.; Fukuhara, Y.; Sugiyama, T.; Li, X.K.; Takahara, S. Astaxanthin prevents ischemia-reperfusion injury of the steatotic liver in mice. PLoS ONE 2017, 12, e0187810. [Google Scholar] [CrossRef]
- Liang, T.; Xu, S.; Yu, J.; Shen, K.; Li, D.; Zheng, S. Activation pattern of mitogen-activated protein kinases in early phase of different size liver isografts in rats. Liver Transpl. 2005, 11, 1527–1532. [Google Scholar] [CrossRef] [PubMed]
- Ping, P.; Zhang, J.; Huang, S.; Cao, X.; Tang, X.L.; Li, R.C.; Zheng, Y.T.; Qiu, Y.; Clerk, A.; Sugden, P.; et al. PKC-dependent activation of p46/p54 JNKs during ischemic preconditioning in conscious rabbits. Am. J. Physiol. 1999, 277, H1771–H1785. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Gerl, R.E.; Schrader, J.W. Defining the involvement of p38α MAPK in the production of anti and proinflammatory cytokines using an SB203580-resistant form of the kinase. J. Biol. Chem. 2003, 278, 22237–22242. [Google Scholar] [CrossRef] [PubMed]
- Dumeus, S.; Shibu, M.A.; Lin, W.T.; Wang, M.F.; Lai, C.H.; Shen, C.Y.; Lin, Y.M.; Viswanadha, V.P.; Kuo, W.W.; Huang, C.Y. Bioactive peptide improves diet-induced hepatic fat deposition and hepatocyte proinflammatory response in SAMP8 ageing mice. Cell Physiol. Biochem. 2018, 48, 1942–1952. [Google Scholar] [CrossRef] [PubMed]
- Karin, M. Mitogen activated protein kinases as targets for development of novel anti-inflammatory drugs. Ann. Rheum. Dis. 2004, 63, ii62–ii64. [Google Scholar] [CrossRef]
- Kobayashi, K.; Oshima, K.; Muraoka, M.; Akao, T.; Totsuka, O.; Shimizu, H.; Sato, H.; Tanaka, K.; Konno, K.; Matsumoto, K.; et al. Effect of atrial natriuretic peptide on ischemia–reperfusion injury in a porcine total hepatic vascular exclusion model. World J. Gastroenterol. 2007, 13, 3487–3492. [Google Scholar] [CrossRef]
- Imarisio, C.; Alchera, E.; Revanna, C.B.; Valente, G.; Follenzi, A.; Trisolini, E.; Boldorini, R.; Carini, R. Oxidative and ER stress-dependent ASK1 activation in steatotic hepatocytes and Kupffer cells sensitizes mice fatty liver to ischemia/reperfusion injury. Free Radic. Biol. Med. 2017, 112, 141–148. [Google Scholar] [CrossRef]
- Deng, J.; Feng, J.; Liu, T.; Lu, X.; Wang, W.; Liu, N.; Lv, Y.; Liu, Q.; Guo, C.; Zhou, Y. Beraprost sodium preconditioning prevents inflammation, apoptosis, and autophagy during hepatic ischemia-reperfusion injury in mice via the P38 and JNK pathways. Drug Des. Dev. Ther. 2018, 12, 4067–4082. [Google Scholar] [CrossRef]
- Xu, S.; Niu, P.; Chen, K.; Xia, Y.; Yu, Q.; Liu, N.; Li, J.; Li, S.; Wu, L.; Feng, J.; et al. The liver protection of propylene glycol alginate sodium sulfate preconditioning against ischemia reperfusion injury: Focusing MAPK pathway activity. Sci. Rep. 2017, 7, 15175. [Google Scholar] [CrossRef]
- Yang, L.; Wang, W.; Wang, X.; Zhao, J.; Xiao, L.; Gui, W.; Fan, H.; Xia, J.; Li, Z.; Yan, J.; et al. Creg in hepatocytes ameliorates liver ischemia/reperfusion injury in a TAK1-dependent manner in mice. Hepatology 2019, 69, 294–313. [Google Scholar] [CrossRef]
- Ocuin, L.M.; Zeng, S.; Cavnar, M.J.; Sorenson, E.C.; Bamboat, Z.M.; Greer, J.B.; Kim, T.S.; Popow, R.; DeMatteo, R.P. Nilotinib protects the murine liver from ischemia/reperfusion injury. J. Hepatol. 2012, 57, 766–773. [Google Scholar] [CrossRef]
- Wang, X.; Mao, W.; Fang, C.; Tian, S.; Zhu, X.; Yang, L.; Huang, Z.; Li, H. Dusp14 protects against hepatic ischaemia-reperfusion injury via Tak1 suppression. J. Hepatol. 2018, 68, 118–129. [Google Scholar] [CrossRef]
- Kang, J.W.; Lee, S.M. Impaired expression of caveolin-1 contributes to hepatic ischemia and reperfusion injury. Biochem. Biophys. Res. Commun. 2014, 450, 1351–1357. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, S.; Wang, H.; Di, W.; Deng, C.; Jin, Z.; Yi, W.; Xiao, X.; Nie, Y.; Yang, Y. SIRT6 protects against hepatic ischemia/reperfusion injury by inhibiting apoptosis and autophagy related cell death. Free Radic. Biol. Med. 2018, 115, 18–30. [Google Scholar] [CrossRef]
- Li, J.; Wang, F.; Xia, Y.; Dai, W.; Chen, K.; Li, S.; Liu, T.; Zheng, Y.; Wang, J.; Lu, W.; et al. Astaxanthin pretreatment attenuates hepatic ischemia reperfusion-induced apoptosis and autophagy via the ROS/MAPK pathway in mice. Mar. Drugs 2015, 13, 3368–3387. [Google Scholar] [CrossRef]
- Guo, L.; Wu, X.; Zhang, Y.; Wang, F.; Li, J.; Zhu, J. Protective effects of gastrin-releasing peptide receptor antagonist RC-3095 in an animal model of hepatic ischemia/reperfusion injury. Hepatol. Res. 2019. [Google Scholar] [CrossRef]
- Zheng, N.; Liu, F.; Lu, H.; Zhan, Y.; Zhang, M.; Guo, W.; Ding, G. Schisantherin A protects against liver ischemia-reperfusion injury via inhibition of mitogen-activated protein kinase pathway. Int. Immunopharmacol. 2017, 47, 28–37. [Google Scholar] [CrossRef]
- Shin, J.K.; Kang, J.W.; Lee, S.M. Enhanced nitric oxide-mediated autophagy contributes to the hepatoprotective effects of ischemic preconditioning during ischemia and reperfusion. Nitric Oxide 2016, 58, 10–19. [Google Scholar] [CrossRef]
- Sun, L.; Wu, Q.; Nie, Y.; Cheng, N.; Wang, R.; Wang, G.; Zhang, D.; He, H.; Ye, R.D.; Qian, F. A role for MK2 in enhancing neutrophil-derived ROS production and aggravating liver ischemia/reperfusion injury. Front. Immunol. 2018, 9, 2610. [Google Scholar] [CrossRef]
- Teoh, N.; Dela Pena, A.; Farrell, G. Hepatic ischemic preconditioning in mice is associated with activation of NF-κB, p38 kinase, and cell cycle entry. Hepatology 2002, 36, 94–102. [Google Scholar] [CrossRef]
- Sakai, N.; Van Sweringen, H.L.; Quillin, R.C.; Schuster, R.; Blanchard, J.; Burns, J.M.; Tevar, A.D.; Edwards, M.J.; Lentsch, A.B. Interleukin-33 is hepatoprotective during liver ischemia/reperfusion in mice. Hepatology 2012, 56, 1468–1478. [Google Scholar] [CrossRef]
- Teoh, N.; Laclercq, I.; Pena, A.D.; Farrell, G. Low dose TNF-α protects against hepatic ischemia–reperfusion injury in mice: Implications for preconditioning. Hepatology 2003, 37, 118–128. [Google Scholar] [CrossRef]
- Teoh, N.; Field, J.; Sutton, J.; Farrell, G. Dual role of tumor necrosis factor-α in hepatic ischemia-reperfusion injury: Studies in tumor necrosis factor-α gene knockout mice. Hepatology 2004, 39, 412–421. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, D.; An, H.; Zhang, H.; Jiang, C.; Yang, B. Melatonin prevents lung injury induced by hepatic ischemia-reperfusion through anti-inflammatory and anti-apoptosis effects. Int. Immunopharmacol. 2015, 29, 462–467. [Google Scholar] [CrossRef]
- Gendy, A.M.; Abdallah, D.M.; El-Abhar, H.S. The potential curative effect of rebamipide in hepatic ischemia/reperfusion injury. Naunyn Schmiedebergs Arch. Pharmacol. 2017, 390, 691–700. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Kawahara, T.; Kumakura, S.; Hua, J.; Kugimiya, T.; Nagaoka, I.; Inada, E. Effect of oprinome, a phosphodiesterase III inhibitor, on hepatic ischemia-reperfusion injury in rats. Shock 2010, 33, 436–441. [Google Scholar] [CrossRef]
- Sun, P.P.; Yuan, F.; Xu, J.; Sai, K.; Chen, J.; Guan, S. Cryptotanshinone ameliorates hepatic normothermic ischemia and reperfusion injury in rats by anti-mitochondrial apoptosis. Biol. Pharm. Bull. 2014, 37, 1758–1765. [Google Scholar] [CrossRef]
- Iannelli, A.; de Sousa, G.; Zucchini, N.; Saint-Paul, M.C.; Gugenheim, J.; Rahmani, R. Anti-apoptotic pro-survival effect of clotrimazole in a normothermic ischemia reperfusion injury animal model. J. Surg. Res. 2001, 171, 101–107. [Google Scholar] [CrossRef]
- Rao, J.; Zhang, C.; Wang, P.; Lu, L.; Zhang, F. All-trans retinoic acid alleviates hepatic ischemia/reperfusion injury by enhancing manganese superoxide dismutase in rats. Biol. Pharm. Bull. 2010, 33, 869–875. [Google Scholar] [CrossRef]
- Schauer, R.J.; Gerbes, A.L.; Vonier, D.; op den Winkel, M.; Fraunberger, P.; Bilzer, M. Induction of cellular resistance against Kupffer cell-derived oxidant stress: A novel concept of hepatoprotection by ischemic preconditioning. Hepatology 2003, 37, 286–295. [Google Scholar] [CrossRef]
- Kapoor, S.; Berishvili, E.; Bandi, S.; Gupta, S. Ischemic preconditioning affects long-term cell fate through DNA damage-related molecular signaling and altered proliferation. Am. J. Pathol. 2014, 184, 2779–2790. [Google Scholar] [CrossRef]
- Lim, E.J.; Do, G.M.; Shin, J.H.; Kwon, O. Protective effects of Acanthopanax divaricatus vat. albeofructus and its active compound on ischemia-reperfusion injury of rat liver. Biochem. Biophys. Res. Commun. 2013, 432, 599–605. [Google Scholar] [CrossRef]
- Ruess, D.A.; Probst, M.; Marjanovic, G.; Wittel, U.A.; Hopt, U.T.; Keck, T.; Bausch, D. HDACi valproic acid (VPA) and suberoylanilide hydroxamic acid (SAHA) delay but fail to protect against warm hepatic ischemia-reperfusion injury. PLoS ONE 2016, 11, e0161233. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, Q.; Mo, W.; Wu, L.; Li, S.; Li, J.; Liu, T.; Xu, S.; Fan, X.; Guo, C. Salidroside pretreatment attenuates apoptosis and autophagy during hepatic ischemia-reperfusion injury by inhibiting the mitogen-activated protein kinase pathway in mice. Drug Des. Dev. Ther. 2017, 11, 1989–2006. [Google Scholar] [CrossRef]
- Kawai, K.; Yokoyama, Y.; Kokuryo, T.; Watanabe, K.; Kitagawa, T.; Nagino, M. Inchinkoto, an herbal medicine, exerts beneficial effects in the rat liver under stress with hepatic ischemia-reperfusion and subsequent hepatectomy. Ann. Surg. 2010, 251, 692–700. [Google Scholar] [CrossRef]
- Ben Mosbah, I.; Duval, H.; Mbatchi, S.F.; Ribault, C.; Grandadam, S.; Pajaud, J.; Morel, F.; Boudjema, K.; Compagnon, P.; Corlu, A. Intermittent selective clamping improves rat liver regeneration by attenuating oxidative and endoplasmic reticulum stress. Cell Death Dis. 2014, 5, e1107. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, W.; Zhang, Y.; Wang, W.; Yao, F.; Yan, J.; Wan, C. 5′-Methylthioadenosine attenuates ischemia reperfusion injury after liver transplantation in rats. Inflammation 2014, 37, 1366–1373. [Google Scholar] [CrossRef]
- Pantazi, E.; Bejaoui, M.; Zaouali, M.A.; Folch-Puy, E.; Pinto-Rolo, A.; Panisello, A.; Palmeira, C.M.; Roselló-Catafau, J. Losartan activates sirtuin 1 in rat reduced-size orthotopic liver transplantation. World J. Gastroenterol. 2015, 21, 8021–8031. [Google Scholar] [CrossRef][Green Version]
- Kaizu, T.; Ikeda, A.; Nakao, A.; Tsung, A.; Toyokawa, H.; Ueki, S.; Geller, D.A.; Murase, N. Protection of transplant-induced hepatic ischemia/reperfusion injury with carbon monoxide via MEK/ERK1/2 pathway downregulation. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G236–G244. [Google Scholar] [CrossRef]
- Duarte, S.; Shen, X.D.; Fondevila, C.; Busuttil, R.W.; Coito, A.J. Fibronectin-α4β1 interactions in hepatic cold ischemia and reperfusion injury: Regulation of MMP-9 and MT1-MMP via the p38 MAPK pathway. Am. J. Transplant. 2012, 12, 2689–2699. [Google Scholar] [CrossRef]
- Kiemer, A.K.; Kulhanek-Heinze, S.; Gerwig, T.; Gerbes, A.L.; Vollmar, A.M. Stimulation of p38 MAPK by hormonal preconditioning with atrial natriuretic peptide. World J. Gastroenterol. 2002, 8, 707–711. [Google Scholar] [CrossRef]
- Amersi, F.; Shen, X.D.; Anselmo, D.; Melinek, J.; Iyer, S.; Southard, D.J.; Katori, M.; Volk, H.D.; Busuttil, R.W.; Buelow, R.; et al. Ex vivo exposure to carbon monoxide prevents hepatic ischemia/reperfusion injury through p38 MAP kinase pathway. Hepatology 2002, 35, 815–823. [Google Scholar] [CrossRef]
- Bendinelli, P.; Piccoletti, R.; Maroni, P.; Bernelli-Zazzera, A. The MAP kinase cascades are activated during post-ischemic liver reperfusion. FEBS Lett. 1996, 398, 193–197. [Google Scholar] [CrossRef]
- Lee, K.H.; Kim, S.E.; Lee, Y.S. SP600125, a selective JNK inhibitor, aggravates hepatic ischemia-reperfusion injury. Exp. Mol. Med. 2006, 38, 408–416. [Google Scholar] [CrossRef][Green Version]
- Zhai, Y.; Shen, X.D.; Gao, F.; Zhao, A.; Freitas, M.C.; Lassman, C.; Luster, A.D.; Busuttil, R.W.; Kupiec-Weglinski, J.W. CXCL10 regulates liver innate immune response against ischemia and reperfusion injury. Hepatology 2008, 47, 207–214. [Google Scholar] [CrossRef]
- Kang, J.W.; Koh, E.J.; Lee, S.M. Melatonin protects liver against ischemia and reperfusion injury through inhibition of toll-like receptor signaling pathway. J. Pineal Res. 2011, 50, 403–411. [Google Scholar] [CrossRef]
- Pratap, A.; Panakanti, R.; Yang, N.; Eason, J.D.; Mahato, R.I. Inhibition of endogenous hedgehog signaling protects against acute liver injury after ischemia reperfusion. Pharm. Res. 2010, 27, 2492–2504. [Google Scholar] [CrossRef]
- Zeng, S.; Dun, H.; Ippagunta, N.; Rosario, R.; Zhang, Q.Y.; Lefkowitch, J.; Yan, S.F.; Schmidt, A.M.; Emond, J.C. Receptor for advanced glycation end product (RAGE)-dependent modulation of early growth response-1 in hepatic ischemia/reperfusion injury. J. Hepatol. 2009, 50, 929–936. [Google Scholar] [CrossRef]
- Vilatoba, M.; Eckstein, C.; Bilbao, G.; Frennete, L.; Eckhoff, D.E.; Contreras, J.L. 17β-estradiol differentially activates mitogen-activated protein-kinases and improves survival following reperfusion injury of reduced-size liver in mice. Transplant. Proc. 2005, 37, 399–403. [Google Scholar] [CrossRef]
- Uehara, T.; Peng, X.X.; Bennett, B.; Satoh, Y.; Friedman, G.; Currin, R.; Brenner, D.A.; Lemasters, J. c-Jun N-terminal kinase mediates hepatic injury after rat liver transplantation. Transplantation 2004, 78, 324–332. [Google Scholar] [CrossRef]
- Uehara, T.; Bennett, B.; Sakata, S.T.; Satoh, Y.; Bilter, G.K.; Westwick, J.K.; Brenner, D.A. JNK mediates hepatic ischemia reperfusion injury. J. Hepatol. 2005, 42, 850–859. [Google Scholar] [CrossRef]
- Serafín, A.; Roselló-Catafau, J.; Prats, N.; Gelpí, E.; Rodés, J.; Peralta, C. Ischemic preconditioning affects interleukin release in fatty livers of rats undergoing ischemia/reperfusion. Hepatology 2004, 39, 688–698. [Google Scholar] [CrossRef]
- Westwick, J.K.; Weitzel, C.; Leffert, H.L.; Brenner, D.A. Activation of Jun kinase is an early event in hepatic regeneration. J. Clin. Investig. 1995, 95, 803–810. [Google Scholar] [CrossRef]
- Rai, R.M.; Yang, S.Q.; McClain, C.; Karp, C.L.; Klein, A.S.; Diehl, A.M. Kupffer cell depletion by gadolinium chloride enhances liver regeneration after partial hepatectomy in rats. Am. J. Physiol. 1996, 270, G909–G918. [Google Scholar] [CrossRef]
- Suzuki, S.; Nakamura, S.; Sakaguchi, T.; Ochiai, H.; Konno, H.; Baba, S.; Baba, S. Alteration of reticuloendothelial phagocytic function and tumor necrosis factor-alpha production after total hepatic ischemia. Transplantation 1997, 64, 821–827. [Google Scholar] [CrossRef]
- Watanabe, M.; Chijiiwa, K.; Kameoka, N.; Yamaguchi, K.; Kuroki, S.; Tanaka, M. Gadolinium pretreatment decreases survival and impairs liver regeneration after partial hepatectomy under ischemia/reperfusion in rats. Surgery 2000, 127, 456–463. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, C.; Cui, C.; Jiao, Y.; Li, P.; Zhu, L.; Yu, W.; Xia, Q.; Wen, D.; Yang, L. Indispensable role of β-arrestin2 in the protection of remifentanil preconditioning against hepatic ischemic reperfusion injury. Sci. Rep. 2019, 9, 2087. [Google Scholar] [CrossRef]
- Cobb, M.H. MAP kinase pathways. Prog. Biophys. Mol. Biol. 1999, 71, 479–500. [Google Scholar] [CrossRef]
- Park, S.W.; Kim, M.; Chen, S.W.; Brown, K.M.; D’Agati, V.D.; Lee, H.T. Sphinganine-1-phosphate protects kidney and liver after hepatic ischemia and reperfusion in mice through S1P1 receptor activation. Lab. Investig. 2010, 90, 1209–1224. [Google Scholar] [CrossRef]
- Song, J.; Zhang, Y.W.; Yao, A.H.; Yu, Y.; Hua, Z.Y.; Pu, L.Y.; Li, G.Q.; Li, X.C.; Zhang, F.; Sheng, G.Q.; et al. Adenoviral cardiotrophin-1 transfer improves survival and early graft function after ischemia and reperfusion in rat small-for-size liver transplantation model. Transpl. Int. 2008, 21, 372–383. [Google Scholar] [CrossRef]
- Kamo, N.; Shen, X.D.; Ke, B.; Busuttil, R.W.; Kupiec-Weglinski, J.W. Sotrastaurin, a protein kinase C inhibitor, ameliorates ischemia and reperfusion injury in rat orthotopic liver transplantation. Am. J. Transplant. 2011, 11, 2499–2507. [Google Scholar] [CrossRef]
- Eipel, C.; Hübschmann, U.; Abshagen, K.; Wagner, K.F.; Menger, M.D.; Vollmar, B. Erythropoietin as additive of HTK preservation solution in cold ischemia/reperfusion injury of steatotic livers. J. Surg. Res. 2010, 173, 171–179. [Google Scholar] [CrossRef]
- Moore, C.; Shen, X.D.; Fondevila, C.; Coito, A.J. Fibronectin-alpha4beta1 integrin interactions modulate p42/44 MAPK phosphorylation in steatotic liver cold ischemia-reperfusion injury. Transplant. Proc. 2005, 37, 432–434. [Google Scholar] [CrossRef]
- Xu, D.; Zhu, J.; Jeong, S.; Li, D.; Hua, X.; Huang, L.; Zhang, J.; Luo, Y.; Xia, Q. Rictor deficiency aggravates hepatic ischemia/reperfusion injury in mice by suppressing autophagy and regulating MAPK signaling. Cell Physiol. Biochem. 2018, 45, 2199–2212. [Google Scholar] [CrossRef]
- Toledo-Pereyra, L.H.; Lopez-Neblina, F.; Reuben, J.S.; Toledo, A.H.; Ward, P.A. Selectin inhibition modulates Akt/MAPK signaling and chemokine expression after liver ischemia-reperfusion. J. Investig. Surg. 2004, 17, 303–313. [Google Scholar] [CrossRef]
- Man, K.; Ng, K.T.; Lee, T.K.; Lo, C.M.; Sun, C.K.; Li, X.L.; Zhao, Y.; Ho, J.W.; Fan, S.T. FTY720 attenuates hepatic ischemia-reperfusion injury in normal and cirrhotic livers. Am. J. Transplant. 2005, 5, 40–49. [Google Scholar] [CrossRef]
- Vekemans, K.; van Pelt, J.; Komuta, M.; Wylin, T.; Heedfeld, V.; Detry, O.; Monbaliu, D.; Pirenne, J. Attempt to rescue discarded human liver grafts by end ischemic hypothermic oxygenated machine perfusion. Transplant. Proc. 2011, 43, 3455–3459. [Google Scholar] [CrossRef]
- Murry, C.E.; Jennings, R.B.; Reimer, K.A. Preconditioning with ischemia: A delay of lethal cell injury in ischemic myocardium. Circulation 1986, 74, 1124–1136. [Google Scholar] [CrossRef]
- Przyklenk, K.; Bauer, B.; Ovize, M.; Kloner, R.A.; Whittaker, P. Regional ischemic “preconditioning” protects remote virgin myocardium from subsequent sustained coronary occlusion. Circulation 1993, 87, 893–899. [Google Scholar] [CrossRef]
- Bahde, R.; Spiegel, H.U. Hepatic ischaemia-reperfusion injury from bench to bedside. Br. J. Surg. 2010, 97, 1461–1475. [Google Scholar] [CrossRef]
- Desai, K.K.; Dikdan, G.S.; Shareef, A.; Koneru, B. Ischemic preconditioning of the liver: A few perspectives from the bench to bedside translation. Liver Transpl. 2008, 14, 1569–1577. [Google Scholar] [CrossRef]
- Clavien, P.A.; Selzner, M.; Rudiger, H.A.; Graft, R.; Kadry, Z.; Rousson, V.; Jochum, W. A prospective randomized study in 100 consecutive patients undergoing major liver resection with versus without ischemic preconditioning. Ann. Surg. 2003, 238, 843–850. [Google Scholar] [CrossRef]
- Koneru, B.; Shareef, A.; Dikdan, G.; Desai, K.; Klein, K.M.; Peng, B.; Wachsberg, R.H.; de la Torre, A.N.; Debroy, M.; Fisher, A.; Wilson, D.J.; Samanta, A.K. The ischemic preconditioning paradox in deceased donor liver transplantation-evidence from a prospective randomized single blind clinical trial. Am. J. Transplant. 2007, 7, 2788–2796. [Google Scholar] [CrossRef]
- Robertson, F.P.; Magill, L.J.; Wright, G.P.; Fuller, B.; Davidson, B.R. A systematic review and meta-analysis of donor ischaemic preconditioning in liver transplantation. Transpl. Int. 2016, 29, 1147–1154. [Google Scholar] [CrossRef]
- Jiménez-Castro, M.B.; Meroño, N.; Mendes-Braz, M.; Gracia-Sancho, J.; Martínez-Carreres, L.; Cornide-Petronio, M.E.; Casillas-Ramirez, A.; Rodés, J.; Peralta, C. The effect of brain death in rat steatotic and non-steatotic liver transplantation with previous ischemic preconditioning. J. Hepatol. 2015, 62, 83–91. [Google Scholar] [CrossRef]
- Cornide-Petronio, M.E.; Negrete-Sánchez, E.; Mendes-Braz, M.; Casillas-Ramírez, A.; Bujaldon, E.; Meroño, N.; Martínez-Carreres, L.; Gracia-Sancho, J.; Rodés, J.; Jiménez-Castro, M.B.; et al. The effect of high-mobility group box 1 in rat steatotic and nonsteatotic liver transplantation from donors after brain death. Am. J. Transplant. 2016, 16, 1148–1159. [Google Scholar] [CrossRef]
- Franco-Gou, R.; Peralta, C.; Massip-Salcedo, M.; Xaus, C.; Serafín, A.; Roselló-Catafau, J. Protection of reduced-size liver for transplantation. Am. J. Transplant. 2004, 4, 1408–1420. [Google Scholar] [CrossRef]
- Barrier, A.; Olaya, N.; Chiappini, F.; Roser, F.; Scatton, O.; Artus, C.; Franc, B.; Dudoit, S.; Flahault, A.; Debuire, B.; et al. Ischemic preconditioning modulates the expression of several genes, leading to the overproduction of IL-1Ra, iNOS, and Bcl-2 in a human model of liver ischemia-reperfusion. FASEB J. 2005, 19, 1617–1626. [Google Scholar] [CrossRef]
- Cutrin, J.C.; Perrelli, M.G.; Cavalieri, B.; Peralta, C.; Rosello-Catafau, J.; Poli, G. Microvascular dysfunction induced by reperfusion injury and protective effect of ischemic preconditioning. Free Radic. Biol. Med. 2002, 33, 1200–1208. [Google Scholar] [CrossRef]
- Serafin, A.; Fernandez-Zabalegui, L.; Prats, N.; Wu, Z.Y.; Rosello-Catafau, J.; Peralta, C. Ischemic preconditioning: Tolerance to hepatic ischemia-reperfusion injury. Histol. Histopathol. 2004, 19, 281–289. [Google Scholar]
- Carini, R.; Albano, E. Recent insights on the mechanisms of liver preconditioning. Gastroenterology 2003, 125, 1480–1491. [Google Scholar] [CrossRef]
- Selzner, N.; Rudiger, H.; Graf, R.; Clavien, P. Protective strategies against ischemic injury of the liver. Gastroenterology 2003, 125, 917–936. [Google Scholar] [CrossRef]
- Alchera, E.; Dal Ponte, C.; Imarisio, C.; Albano, E.; Carini, R. Molecular mechanisms of liver preconditioning. World J. Gastroenterol. 2010, 16, 6058–6067. [Google Scholar] [CrossRef]
- Richards, J.A.; Wigmore, S.J.; Devey, L.R. Heme oxygenase system in hepatic ischemia-reperfusion injury. World J. Gastroenterol. 2010, 16, 6068–6078. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, J.; Xiong, X.; Xu, Y.; Zhang, H.; Huang, C.; Tian, Y.; Jiao, C.; Wang, X.; Li, X. Remote ischemic preconditioning protects against liver ischemia-reperfusion injury via heme oxygenase-1- induced autophagy. PLoS ONE 2014, 9, e98834. [Google Scholar] [CrossRef]
- Carini, R.; Alchera, E.; De Cesaris, M.G.; Splendore, R.; Piranda, D.; Baldanzi, G.; Albano, E. Purinergic P2Y2 receptors promote hepatocyte resistance to hipoxia. J. Hepatol. 2006, 45, 236–245. [Google Scholar] [CrossRef]
- Jin, L.M.; Jin, S.F.; Liu, Y.X.; Zhou, L.; Xie, H.Y.; Yan, S.; Xu, X.; Zheng, S.S. Ischemic preconditioning enhances hepatocyte proliferation in the early phase after ischemia under hemi-hepatectomy in rats. Hepatobiliary Pancreat. Dis. Int. 2012, 11, 521–526. [Google Scholar] [CrossRef]
- Pantazi, E.; Zaouali, M.A.; Bejaoui, M.; Serafin, A.; Folch-Puy, E.; Petegnief, V.; De Vera, N.; Ben Abdennebi, H.; Rimola, A.; Roselló-Catafau, J. Silent information regulator 1 protects the liver against ischemia-reperfusion injury: Implications in steatotic liver ischemic preconditioning. Transpl. Int. 2014, 27, 493–503. [Google Scholar] [CrossRef]
- Terada, K.; Kaziro, Y.; Satoh, T. Analysis of Ras-dependent signals that prevent caspase-3 activation and apoptosis induced by cytokine deprivation in hematopoietic cells. Biochem. Biophys. Res. Commun. 2000, 267, 449–455. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Experimental Model | Changes in p38 Induced by I-R | Treatment with p38 Modulators | Dose and Administration | Effect of p38 Modulators | |
|---|---|---|---|---|---|
| Species | Surgical Procedure | ||||
| Mice | Partial Warm I-R I: 45 min-R: 120 min [59] | ↑ p38 | NQDI-1 | 0.25 mg/kg, i.p., twice a week before surgery | ↓ p38 phosphorylation, damage |
| Rats | Partial Warm I-R I: 60 min-R: 30 min [40] | ↑ p38 | SB203580 | 1 mg/kg, i.p., 24 h before I-R | ↓ p38 phosphorylation, damage ↑ HSP72 |
| Partial Warm I-R I: 60 min-R: 30 min [40] | ↑ p38 | Anisomycin PC | 0.1 mg/kg, i.p., 24 h before PC | ↑ p38 phosphorylation, damage | |
| Mice | Total Warm I-R I: 15 min-R: 3 h [52] | ↑ p38 | Astaxanthin | 25 mg/kg, oral, 48 h, 24 h and 40 min before I-R | ↓ p38 phosphorylation, damage apoptosis |
| Rats | I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [44] | ↑ p38 | Anisomycin | 0.1 mg/kg, na, 24 h before surgery | No activity changes in p38 |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [44] | ↑ p38 | SB203580 | 1 mg/kg, na, 24 h before surgery | ↓ p38 phosphorylation, damage | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [44] | ↑ p38 | EGF | 100 µg/kg, na, before surgery every 8 h for 3 doses | ↓ p38 phosphorylation | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [45] | ↑ p38 | Losartan PD123319 | Losartan: 5 mg/kg, oral, 24 and 1.5 h before surgery PD123319: 30 mg/kg, i.v., 5 min before surgery | ↓ Damage ↑ p38 phosphorylation, regeneration | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [46] | ↑ p38 | TUDCA | 50 mg/kg, na, before surgery | ↓ mitochondrial damage, ER stress ↑ p38 phosphorylation, regeneration | |
| Experimental Model | Changes in p38 Induced by I-R | Treatment with p38 Modulators | Dose and Administration | Effect of p38 Modulators | |
|---|---|---|---|---|---|
| Species | Surgical Procedure | ||||
| Mice | Partial Warm I-R I: 45 min-R: 2, 8, 24 h [60] | ↑ p38 | Beraprost sodium | 50, 100 μg/kg/d, oral, 7 days before I-R | ↓ p38 phosphorylation, damage, apoptosis, autophagy |
| Partial Warm I-R I: 45 min-R: 24 h [61] | ↑ p38 | Propylene glycol alginate sodium sulfate | 25, 50 mg/kg, i.p., 1 h before surgery | ↓ p38 phosphorylation, damage, apoptosis, autophagy, inflammation | |
| Partial Warm I-R I: 60 min-R: 1, 3, 6, 12, 24 h [62] | ↑ p38 | 5Z-7-oxozeaenol | 5 mg/kg, i.p., 30 min before I-R | ↓ p38 phosphorylation, NF-κB, damage, inflammation | |
| Partial Warm I-R I: 60 min-R: 1, 3, 6, 12 h [63] | ↑ p38 | Nilotinib | 30 mg/kg, oral, 12 and 2 h before I-R | ↓ p38 phosphorylation, damage, inflammatory cytokines | |
| Partial Warm I-R I: 60 min-R: 1, 6, 24 h [64] | ↑ p38 | 5Z-7-oxozeaenol | Adenovirus vectors, i.v., 4 weeks before I-R | ↓ p38 phosphorylation, damage, inflammation | |
| Partial Warm I-R I: 60 min-R: 1, 4, 24 h [65] | ↑ p38 | Methyl-beta-cyclodextrin | 10, 25, 50 mg/kg, i.p., 48 and 24 h before I-R | ↑ p38 phosphorylation, damage, apoptosis | |
| Partial Warm I-R I: 60 min-R: 2, 4, 8, 12, 24 h [66] | ↑ p38 | SIRT6 -/- KO | na | ↓ p38 phosphorylation, damage, NF-κB | |
| Partial Warm I-R I: 60 min-R: 2, 4, 24 h [67] | ↑ p38 | Astaxanthin | 30, 60 mg/kg, oral, 14 consecutive days before I-R | ↓ p38 phosphorylation, damage, apoptosis, ROS, autophagy | |
| Partial Warm I-R I: 60 min-R: 3, 6, 12, 24 h [68] | ↑ p38 | RC-3095 | 3 mg/kg, i.p., at the time of reperfusion | ↓ p38 phosphorylation, damage, inflammation | |
| Partial Warm I-R I: 60 min-R: 6 h [69] | ↑ p38 | Schisantherin A | 200 mg/kg/d, oral, 5 days | ↓ p38 phosphorylation, damage | |
| Partial Warm I-R I: 60 min-R: 6 h [70] | ↑ p38 | PC | 10 min ischemia, 10 min reperfusion | ↓ p38 phosphorylation, damage | |
| Partial Warm I-R I: 60 min-R: 6, 18 h [71] | ↑ p38 | MK2 -/- KO | na | ↓ p38 phosphorylation, damage, ROS production | |
| Partial Warm I-R I: 90 min-R: 90 min [72] | No activity changes in p38 | PC | 10 min ischemia, 10 min reperfusion | ↓ Damage, NF-κB ↑ p38 phosphorylation | |
| Partial Warm I-R I: 90 min-R: 1, 4, 8 h [73] | ↑ p38 | IL-33 recombinant | 5, 10 µg/mice, i.p., 16 and 1 h before I-R | ↓ Damage ↑ p38 phosphorylation, NF-κB, cyclin D1, Bcl-2 | |
| Partial Warm I-R I: 90 min-R: 2, 24, 44 h [74] | ↑ p38 | TNF-α + PC (I-R: 10 min) | 1, 5 µg/kg, na, 30 min before I-R | ↓ Damage ↑ p38 phosphorylation, NF-κB, STAT3, cyclin D1, cdk4 | |
| Partial Warm I-R I: 90 min-R: 2, 24, 44 h [75] | ↑ p38 | TNF -/- KO | na | ↓ Damage ↑ p38 phosphorylation | |
| Rats | Partial Warm I-R I: 30 min-R: 3 h [76] | ↑ p38 | Melatonin | 10 mg/kg, i.v., 15 min before I & 10 min before R | ↓ p38 phosphorylation, damage |
| Partial Warm I-R I: 30 min-R: 72 h [77] | ↑ p38 | Rebamipide | 60, 100 mg/kg, i.p., after reperfusion and for 2 days | ↓ p38 phosphorylation, damage, HMGB1, caspase-3 ↑ ATP | |
| Partial Warm I-R I: 60 min-R: 30 min [40] | ↑ p38 | SB203580 | 1 mg/kg, i.p., 24 h before I-R | ↓ p38 phosphorylation, damage ↑ HSP72 | |
| Partial Warm I-R I: 60 min-R: 30 min [40] | ↑ p38 | Anisomycin PC | 0.1 mg/kg, i.p., 24 h before PC | ↑ p38 phosphorylation, damage | |
| Partial Warm I-R I: 60 min-R: 3 h [78] | ↑ p38 | Olprinone | 20 µg/kg, i.v., 5 min before reperfusion | ↓ p38 phosphorylation, damage, NF-κB, IL-6, TNFα, ICAM-1 ↑ cAMP | |
| Partial Warm I-R I: 60 min-R: 4 h [79] | ↑ p38 | Cryptotan- shinone | 50 mg/kg/d, i.p., 7 days before I-R | ↓ p38 phosphorylation, damage, apoptosis | |
| Partial Warm I-R I: 60 min-R: 6 h [80] | ↑ p38 | Clotrimazole | 50 mg/kg, i.p., 3 days before I-R | No activity changes in p38 ↓ Damage, apoptosis | |
| Partial Warm I-R I: 60 min-R: 24 h [81] | ↑ p38 | All-tras retinoic acid | 15 mg/kg/d, i.p., 14 days before I-R | ↓ p38 phosphorylation, damage | |
| Partial Warm I-R I: 60 min-R: 120 min [82] | ↑ p38 | Anisomycin | 0.5 µg/ml, i.v., 60-70 min in 100 total min of I-R | ↑ p38 phosphorylation, damage, GSH/GSSG redox system | |
| Partial Warm I-R I: 60 min-R: 5 days [83] | No activity changes in p38 | PC | 10 min ischemia, 15 min reperfusion | ↑ p38 phosphorylation | |
| Partial Warm I-R I: 60 min-R: 7 days [84] | No activity changes in p38 | Acanthopanax divaricatus vat. albeofructus | 600 mg/kg, oral, 2 weeks before I-R | ↓ Damage ↑ p38 phosphorylation, IL-10 | |
| Partial Warm I-R I: 90 min-R: 6, 11, 24, 60 h [85] | ↑ p38 | HDACi valproic & suberoylanilide hydroxamic acids | Valproic acid: 300 mg/kg, i.p., 24, 12 and 0 h before I-R, Suberoylanilide: 60 mg/kg, i.p., 24 and 0 h before I-R | ↓ p38 phosphorylation ↑ Damage | |
| Partial Warm I-R I: 120 min-R: 1, 3, 6 h [47] | ↑ p38 | Z-Asp-cmk | 50 mg/ml, i.v., before I-R | ↓ Damage, apoptosis ↑ p38 phosphorylation | |
| Mice | Total Warm I-R I: 45 min-R: 2, 8, 24 h [86] | ↑ p38 | Salidroside | 20 mg/kg, i.p., 2 h before I-R | ↓ p38 phosphorylation, damage, apoptosis, autophagy |
| Rats | Total Warm I-R I: 30 min-R: 30, 60, 90, 120 min [33] | ↑ p38 | FR167653 | 0.1 mg/kg/h, i.v., 30 minutes before I to 2 h after R | ↓ p38 phosphorylation, damage, IL-1β, TNF-α |
| Pig | Total Warm I-R I: 120 min-R: 2 h [58] | ↑ p38 | ANP | 0.1 μg/kg/min, i.v., 30 min before I to the end of the R | ↓ Damage, TNF-α, NF-κB ↑ p38 phosphorylation |
| Rats | I-R with Partial (70%) Hepatectomy I: 15 min-R: 1 h [87] | ↑ p38 | Inchinkoto | 1, 2 g/kg, na, 3 days before I-R | ↓ p38 phosphorylation, damage ↑ Survival |
| I-R with Partial (70%) Hepatectomy I: 30 min-R: 3, 9, 18, 24, 30, 48 h [88] | No activity changes in p38 | PC | 10 min ischemia, 5 min reperfusion | ↓ Damage, apoptosis ↑ p38 phosphorylation, mitochondrial integrity | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [44] | ↑ p38 | Anisomycin | 0.1 mg/kg, na, 24 h before I-R | No activity changes in p38 | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [44] | ↑ p38 | SB203580 | 1 mg/kg, na, 24 h before I-R | ↓ p38 phosphorylation, damage | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [44] | ↑ p38 | EGF | 100 µg/kg, na, before surgery every 8 h for 3 doses | ↓ p38 phosphorylation, damage | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [45] | ↑ p38 | Losartan PD123319 | Losartan: 5 mg/kg, oral, 24 and 1.5 h before surgery. PD123319: 30 mg/kg, i.v., 5 min before surgery | ↓ Damage ↑ p38 phosphorylation, regeneration | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [46] | ↑ p38 | TUDCA | 100 mg/kg, na, before surgery | ↓ Mitochondrial damage, ER stress ↑ p38 phosphorylation, regeneration | |
| Rats | Liver transplantation Cold I: 30 min-R: 3, 6, 24 h [89] | ↑ p38 | 5′-Methylthio- adenosine | 96 μmol/kg, i.p., 30 min before harvest | ↓ p38 phosphorylation, damage, NF-κB, inflammation |
| Liver transplantation Cold I: 1 h-R: 24 h [90] | ↑ p38 | Losartan | 5 mg/kg, oral, 24 and 1 h before surgery | ↓ p38 phosphorylation, damage ↑ SIRT1 | |
| Liver transplantation Cold I: 18 h-R: 1, 3, 6, 24, 48 h [91] | ↑ p38 | Carbon monoxide | 100 ppm in recipient, 1 h before surgery and 1, 3, and 6 h after surgery | No activity changes in p38 ↓ Damage | |
| Liver transplantation Cold I: 24 h-R: 2 h [51] | ↑ p38 | ANP | 200 nmol/L, na, 10 min after perfusion | ↓ Damage, alterations of the hepatocyte cytoskeleton ↑ p38 phosphorylation | |
| Liver transplantation Cold I: 24 h-R: 6, 24 h [92] | ↑ p38 | CS-1 peptide | 500 µg/rat, i.v, before cold storage and reperfusion. 1 mg/rat, i.p., 1 h after transplant in recipients | ↓ p38 phosphorylation, damage, TNF-α, INF-γ, iNOS, leukocyte recruitment ↑ Survival | |
| Liver transplantation Cold I: 30 h-R: 6, 24 h [39] | ↑ p38 | FR167653 as additive | 100 ml/kg, in perfused, before removal the liver | ↓ p38 phosphorylation, damage ↑ Survival | |
| Rats | Isolated Liver Perfused Cold I: 24 h-R: 45 min [51] | ↑ p38 | SB203580 + ANP | SB203580: 2 µmol/L, na, after perfusion. ANP: 200 nmol/L, na, 10 min after perfusion. | ↓ p38 phosphorylation, Hsp27 ↑ Damage |
| Isolated Liver Perfused Cold I: 24 h-R: 2 h [93] | ↓ p38 | ANP | 200 nM added to the perfusate for 20 min | ↓ Damage ↑ p38 activity | |
| Isolated Liver Perfused Cold I: 24 h-R: 2 h [94] | ↑ p38 | SB203580 | 25 mg/kg, oral, 1 h before harvest and ex vivo perfusion with 20 µmol/L plus CO | ↓ Damage, apoptosis | |
| Experimental Model | Changes in JNK Induced by I-R | Treatment with JNK Modulators | Dose and Administration | Effect of JNK Modulators | |
|---|---|---|---|---|---|
| Species | Surgical Procedure | ||||
| Mice | Partial Warm I-R I: 45 min-R: 120 min [59] | ↑ JNK | NQDI-1 | 0.25 mg/kg, i.p., twice a week before surgery | ↓ JNK phosphorylation, damage |
| Rats | Partial Warm I-R I: 60 min-R: 30 min [40] | ↑ JNK | SP600125 | 6 mg/kg, s.c., 1 h before I-R | ↓ JNK phosphorylation, damage |
| Mice | Total Warm I-R I: 15 min-R: 3 h [52] | ↑ JNK | Astaxanthin | 25 mg/kg, oral, 48, 24 h and 40 min before I-R | ↓ JNK phosphorylation, damage, apoptosis |
| Rats | I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [46] | ↑ JNK | TUDCA | 100 mg/kg, na, before surgery | ↓ JNK phosphorylation, damage, caspase 12 |
| Experimental Model | Changes in JNK Induced by I-R | Treatment with JNK Modulators | Dose and Administration | Effect of JNK MODULATORS | |
|---|---|---|---|---|---|
| Species | Surgical Procedure | ||||
| Mice | Partial Warm I-R I: 45 min-R: 2 h [109] | ↑ JNK | Remifentanil | 30 μg/kg, i.p., 10 min before I-R | ↓ JNK phosphorylation, damage |
| Partial Warm I-R I: 45 min-R: 2, 8, 24 h [60] | ↑ JNK | Beraprost sodium | 50, 100 μg/kg/d, oral, 7 days before I-R | ↓ JNK phosphorylation, damage, apoptosis, autophagy | |
| Partial Warm I-R I: 45 min-R: 24 h [61] | ↑ JNK | Propylene glycol alginate sodium sulfate | 25, 50 mg/kg, i.p., 1 h before surgery | ↓ JNK phosphorylation, damage, apoptosis, autophagy, inflammation | |
| Partial Warm I-R I: 60 min-R: 15 min, 1 h [96] | ↑ JNK | SP600125 | 10 mg/kg, i.p., at the onset of ischemia and 10 mg/kg, s.c., after reperfusion | ↓ JNK phosphorylation ↑ Damage, leukocyte infiltration | |
| Partial Warm I-R I: 60 min-R: 1, 3, 6, 12, 24 h [62] | ↑ JNK | 5Z-7-oxozeaenol | 5 mg/kg, i.p., 30 min before I-R | ↓ JNK phosphorylation, damage, inflammation, NF-κB | |
| Partial Warm I-R I: 60 min-R: 1, 6, 24 h [64] | ↑ JNK | 5Z-7-oxozeaenol | Adenovirus vectors, i.v., 4 weeks before I-R | ↓ JNK phosphorylation, damage, inflammation | |
| Partial Warm I-R I: 60 min-R: 1, 4, 24 h [65] | ↑ JNK | Methyl-beta-cyclodextrin | 10, 25, 50 mg/kg, i.p., 48 and 24 h before I-R | ↑ JNK phosphorylation, damage, apoptosis | |
| Partial Warm I-R I: 60 min-R: 2, 4, 8, 12, 24 h [66] | ↑ JNK | SIRT6 -/- KO | na | ↓ JNK phosphorylation, damage, NF-κB | |
| Partial Warm I-R I: 60 min-R: 2, 4, 24 h [67] | ↑ JNK | Astaxanthin | 30, 60 mg/kg, oral, 14 consecutive days before I-R | ↓ JNK phosphorylation, damage, apoptosis, ROS, autophagy | |
| Partial Warm I-R I: 60 min-R: 6 h [69] | ↑ JNK | Schisantherin A | 200 mg/kg/d, oral, 5 days | ↓ JNK phosphorylation, damage | |
| Partial Warm I-R I: 60 min-R: 6 h [70] | ↑ JNK | PC | 10 min ischemia, 10 min reperfusion | No activity changes in JNK ↓ Damage | |
| Partial Warm I-R I: 90 min-R: 1, 2, 4, 6 h [97] | ↑ JNK | CXCL10 -/- KO | na | No activity changes in JNK ↓ Damage, neutrophil infiltration | |
| Partial Warm I-R I: 90 min-R: 90 min [72] | No activity changes in JNK | PC | 10 min ischemia, 10 min reperfusion | ↓ Damage, NF-κB ↑ JNK phosphorylation | |
| Partial Warm I-R I: 90 min-R: 2, 4 h [108] | ↑ JNK | SB203580 | shRNA plasmids: 10 μg in 2 ml, i.v., 72 h before I-R | ↓ JNK phosphorylation, damage | |
| Partial Warm I-R I: 90 min-R: 2, 24, 44 h [74] | ↑ JNK | TNF-α + PC (I-R: 10 min) | 1, 5 µg/kg, na, 30 min before I-R | ↓ Damage ↑ JNK phosphorylation | |
| Partial Warm I-R I: 90 min-R: 2, 24, 44 h [75] | ↑ JNK | TNF -/- KO | na | ↓ JNK phosphorylation, damage | |
| Rats | Partial Warm I-R I: 30 min-R: 3 h [76] | ↑ JNK | Melatonin | 10 mg/kg, i.v., 15 min before I & 10 min before R | ↓ JNK phosphorylation, damage |
| Partial Warm I-R I: 60 min-R: 30 min [95] | ↑ JNK | IL-1 RA | 100–400 µg/100 g, i.p., 30 min before I and immediately after R | ↓ JNK phosphorylation, damage, IL-6, TNF-α | |
| Partial Warm I-R I: 60 min-R: 30 min [40] | ↑ JNK | SP600125 | 6 mg/kg, s.c., 1 h before I-R | ↓ JNK phosphorylation, damage | |
| Partial Warm I-R I: 60 min-R: 1, 5 h [98] | ↑ JNK | Melatonin | 10 mg/kg, i.v., 15 min before I & 10 min before R | ↓ JNK phosphorylation, damage, inflammatory signaling | |
| Partial Warm I-R I: 60 min-R: 3 h [78] | ↑ JNK | Olprinone | 20 µg/kg, i.v., 5 min before reperfusion | ↓ JNK phosphorylation, damage, IL-6, TNF-α, NF-κB, ICAM-1 ↑ cAMP | |
| Partial Warm I-R I: 60 min-R: 4 h [79] | ↑ JNK | Cryptotan- shinone | 50 mg/kg/d, i.p., 7 days before I-R | ↓ JNK phosphorylation, damage, apoptosis | |
| Partial Warm I-R I: 60 min-R: 6 h [80] | ↑ JNK | Clotrimazole | 50 mg/kg, i.p., 3 days before I-R | ↓ JNK phosphorylation, damage, apoptosis | |
| Partial Warm I-R I: 60 min-R: 7 days [84] | ↑ JNK | Acanthopanax divaricatus vat. albeofructus | 600 mg/kg, oral, 2 weeks before I-R | ↓ JNK phosphorylation, damage ↑ IL-10 | |
| Partial Warm I-R I: 90 min-R: 1 h [99] | ↑ JNK | Cyclopamine | 10 mg/kg, i.p., 3 days and 1 h before surgery | ↓ JNK phosphorylation, damage, inflammation | |
| Partial Warm I-R I: 90 min-R: 6, 11, 24, 60 h [85] | ↑ JNK | HDACi valproic & suberoylanilide hydroxamic acids | Valproic acid: 300 mg/kg, i.p., 24, 12 and 0 h before I-R, Suberoylanilide: 60 mg/kg, i.p., 24 and 0 h before I-R | ↑ JNK phosphorylation, damage | |
| Partial Warm I-R I: 120 min-R: 1, 3, 6 h [47] | ↑ JNK | Z-Asp-cmk | 50 mg/ml, i.v., before I-R | ↓ Damage, apoptosis ↑ JNK phosphorylation | |
| Mice | Total Warm I-R I: 45 min-R: 2, 8, 24 h [86] | ↑ JNK | Salidroside | 20 mg/kg, i.p., 2 h before I-R | ↓ JNK phosphorylation, damage, apoptosis, autophagy |
| Total Warm I-R I: 10, 15 min-R: 10, 20, 30 min [38] | ↑ JNK | d-α-tocopherol | 40 mg/kg, i.p., 2 weeks before surgery | ↓ JNK phosphorylation, damage, apoptosis | |
| Mice | I-R with Partial (30%) Hepatectomy I: 75 min-R: 1 h [100] | ↑ JNK | RAGE -/- KO | na | ↓ JNK phosphorylation, damage, neutrophil infiltration, TNF-α |
| I-R with Partial (70%) Hepatectomy I: 45 min-R: 1, 6, 24 h [101] | ↑ JNK | 17β-Estradiol | 1 µg in 100 ml, s.c., 24 h before I-R | ↓ JNK phosphorylation ↑ Regeneration, survival | |
| Rats | I-R with Partial (30%) Hepatectomy I: 60 min-R: 60 min [103] | ↑ JNK | CC0223105 CC0209766 CC-401 | 3-20 mg/kg, i.v., 15 min before ischemia | ↓ JNK phosphorylation, damage, cytochrome c release, apoptosis, ROS ↑ Survival |
| I-R with Partial (68%) Hepatectomy I: 30 min-R: 3, 8, 48 h [34] | ↑ JNK | Melatonin | 50 mg/kg, oral, 2 h before ischemia | ↓ JNK phosphorylation, damage, iNOS, leukocyte infiltration ↑ Survival | |
| I-R with Partial (70%) Hepatectomy I: 30 min-R: 3, 9, 18, 21, 24, 30, 48 h [88] | No activity changes in JNK | PC (I-R: 10-5 min) | 10 min ischemia, 5 min reperfusion | ↓ JNK phosphorylation, damage, apoptosis ↑ Mitochondrial integrity | |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [46] | ↑ JNK | TUDCA | 100 mg/kg, na, before surgery | ↓ JNK phosphorylation, damage, caspase 12 | |
| Rats | Liver transplantation Cold I: 18 h-R: 1, 3, 6, 24, 48 h [91] | ↑ JNK | Carbon monoxide | 100 ppm in recipient, 1 h before surgery and 1, 3, and 6 h after surgery | No activity changes in JNK ↓ Damage |
| Liver transplantation Cold I: 30 h-R: 6, 24 h [39] | ↑ JNK | FR167653 as additive | 100 ml/kg, in perfused, before removal the liver | No activity changes in JNK ↓ Damage ↑ Survival | |
| Liver transplantation Cold I: 44 h-R: 15 min, 1, 8 h [102] | ↑ JNK | CC-401 | 10 mg/kg, i.v., 10 min before removal the liver and 20 µM in the solution for 44 h | ↓ JNK phosphorylation, damage, TNF-α, caspase 3, apoptosis | |
| Rats | Isolated Liver Perfused I: 24 h-R: 2 h [93] | ↑ JNK | ANP | 200 nM added to the perfusate for 20 min | No activity changes in JNK ↓ Damage |
| Experimental Model | Changes in ERK Induced by I-R | Treatment with ERK Modulators | Dose and Administration | Effect of ERK Modulators | |
|---|---|---|---|---|---|
| Species | Surgical Procedure | ||||
| Mice | Total Warm I-R I: 15 min-R: 3 h [52] | ↓ ERK | Astaxanthin | 25 mg/kg, oral, 48, 24 h and 40 min before I-R | ↓ ERK phosphorylation, damage, apoptosis |
| Rats | I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [46] | ↑ ERK | TUDCA | 50 mg/kg, na, before surgery | ↓ Damage, GSK3β activity ↑ ERK phosphorylation |
| Rats | Liver transplantation Cold I: 4 h-R: 1, 3, 7 days [115] | ↑ ERK | CS-1 peptide | 500 µg/rat, i.v., before and during surgery and 1 mg/rat/day, i.v., 3 days in recipients | ↓ ERK phosphorylation, damage, IL-2, IFN-γ, T-cell |
| Liver transplantation Cold I: 6 h-R: 4 h [50] | No activity changes in ERK | Angiotensin II receptor antagonists | AT1R: 5 mg/kg, oral, 24 and 1.5 h before surgery AT2R: 30 mg/kg, i.v., 5 min before surgery | ↓ Damage ↑ ERK phosphorylation, survival | |
| Mice | Isolated liver perfused Cold I: 24 h-R: 2 h [114] | ↓ ERK | Erythropoietin as additive | 10 IU/mL in solution | ↓ ERK phosphorylation, damage |
| Experimental Model | Changes in ERK Induced by I-R | Treatment with ERK Modulators | Dose and Administration | Effect of ERK Modulators | |
|---|---|---|---|---|---|
| Species | Surgical Procedure | ||||
| Mice | Partial Warm I-R I: 30 min-R: 4 h [7] | ↑ ERK | Cyclopamine | 10 mg/kg/day, na, 14 days before I-R | ↓ ERK phosphorylation, damage, Akt, apoptosis |
| Partial Warm I-R I: 45 min-R: 2 h [109] | ↑ ERK | Remifentanil | 30 μg/kg, i.p., 10 min before I-R | ↓ ERK phosphorylation, damage | |
| Partial Warm I-R I: 45 min-R: 24 h [61] | ↑ ERK | Propylene glycol alginate sodium sulfate | 25, 50 mg/kg, i.p., 1 h before surgery | ↓ ERK phosphorylation, damage, apoptosis, autophagy, inflammation | |
| Partial Warm I-R I: 60 min-R: 1, 6, 24 h [64] | ↑ ERK | 5Z-7-oxozeaenol | Adenovirus vectors, i.v., 4 weeks before I-R | No activity changes in ERK ↓ Damage, inflammation | |
| Partial Warm I-R I: 60 min-R: 1, 4, 24 h [65] | ↑ ERK | Methyl-beta-cyclodextrin | 10, 25, 50 mg/kg, i.p., 48 and 24 h before I-R | No activity changes in ERK ↑ Damage, apoptosis | |
| Partial Warm I-R I: 60 min-R: 2, 4, 8, 12, 24 h [66] | ↑ ERK | SIRT6 -/- KO | na | ↓ ERK phosphorylation, damage, NF-κB | |
| Partial Warm I-R I: 60 min-R: 2, 4, 24 h [67] | ↑ ERK | Astaxanthin | 30, 60 mg/kg, oral, 14 consecutive days before I-R | ↓ ERK phosphorylation, damage, apoptosis, ROS, autophagy | |
| Partial Warm I-R I: 60 min-R: 3, 6, 12, 24 h [68] | ↑ ERK | RC-3095 | 3 mg/kg, i.p., at the time of reperfusion | ↓ ERK phosphorylation, damage, inflammation | |
| Partial Warm I-R I: 60 min-R: 6 h [69] | ↑ ERK | Schisantherin A | 200 mg/kg/d, oral, 5 days | ↓ ERK phosphorylation, damage | |
| Partial Warm I-R I: 60 min-R: 6 h [70] | ↑ ERK | PC | 10 min ischemia, 10 min reperfusion | ↓ ERK phosphorylation, damage | |
| Partial Warm I-R I: 90 min-R: 2, 4 h [116] | ↑ ERK | SB203580 | shRNA plasmids: 10 μg in 2 ml, i.v., 72 h before I-R | ↓ ERK phosphorylation, damage | |
| Partial Warm I-R I: 90 min-R: 3 h [117] | ↑ ERK | TBC-1269 | 10, 20, 40 mg/kg, i.v., 1 h before reperfusion, and 15 min after reperfusion | ↓ ERK phosphorylation, damage, neutrophil infiltration, inflammation ↑ Akt, proliferation | |
| Partial Warm I-R I: 90 min-R: 1, 2, 4, 6 h [97] | ↑ ERK | CXCL10 -/- KO | na | ↓ ERK phosphorylation, damage, neutrophil infiltration | |
| Rats | Partial Warm I-R I: 60 min-R: 1, 5 h [98] | ↑ ERK | Melatonin | 10 mg/kg, i.v., 15 min before ischemia & before reperfusiom | ↓ ERK phosphorylation, damage, inflammatory signaling cascades |
| Partial Warm I-R I: 60 min-R: 30 min [95] | ↑ ERK | IL-1 RA | 100-400 µg/100 g, i.p., 30 min before I and immediately after R | ↓ ERK phosphorylation, damage, IL-6, TNF-α | |
| Partial Warm I-R I: 60 min-R: 6 h [80] | ↑ ERK | Clotrimazole | 50 mg/kg, i.p., 3 days before I-R | ↓ Damage, apoptosis ↑ ERK phosphorylation, PXR | |
| Partial Warm I-R I: 60 min-R: 20, 60, 90 min, 6, 24 h [118] | ↑ ERK | FTY720 | 1 mg/kg, i.v., 20 min before I and 10 min before R | ↓ ERK phosphorylation, damage, Egr-1, apoptosis ↑ Cell survival, Akt signaling | |
| Partial Warm I-R I: 60 min-R: 7 days [84] | ↑ ERK | Acanthopanax divaricatus vat albeofructus | 600 mg/kg, oral, 2 weeks before I-R | ↓ ERK phosphorylation, damage ↑ IL-10 | |
| Partial Warm I-R I: 90 min-R: 1 h [99] | ↑ ERK | Cyclopamine | 10 mg/kg, i.p., 3 days and 1 h before reperfusion | ↓ ERK phosphorylation, damage, inflammation | |
| Partial Warm I-R I: 60 min-R: 5 days [83] | No activity changes in ERK | PC | 15 min ischemia, 15 min reperfusion | ↑ ERK phosphorylation | |
| Mice | Total Warm I-R I: 45 min-R: 2, 8, 24 h [86] | ↑ ERK | Salidroside | 20 mg/kg, i.p., 2 h before I-R | ↓ ERK phosphorylation, damage, apoptosis, autophagy |
| Mice | I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [111] | Not evaluated | Sphinganine-1-phosphate | 0.1 mg/kg, i.v., before reperfusion and 0.2 mg/kg, s.c., 2 h after reperfusion | ↓ Damage, vascular dysfunction ↑ ERK phosphorylation, endothelial cell integrity |
| I-R with Partial (30%) Hepatectomy I: 75 min-R: 1 h [100] | ↑ ERK | RAGE -/- KO | na | ↓ ERK phosphorylation, damage, neutrophil infiltration | |
| Rats | I-R with Partial (70%) Hepatectomy I: 30 min-R: 3, 9, 18, 21, 24, 30, 48 h [88] | No activity changes in ERK | PC | 10 min ischemia, 5 min reperfusion | ↓ Damage, apoptosis ↑ ERK phosphorylation, mitochondrial integrity |
| I-R with Partial (70%) Hepatectomy I: 60 min-R: 24 h [46] | ↑ ERK | TUDCA | 100 mg/kg, na, before surgery | ↓ Damage, GSK3β activity ↑ ERK phosphorylation | |
| Rats | Liver transplantation Cold I: 1 h-R: 24 h [90] | ↑ ERK | Losartan | 5 mg/kg, oral, 24 and 1 h before surgery | No activity changes in ERK ↓ Damage ↑ SIRT1 |
| Liver transplantation Cold I: 80 min-R: 2, 6, 24 h [112] | ↑ ERK | Cardiotrophin-1 | pAdTrack-CMV vector, na | ↓ Damage, Bcl-2, caspase-3, apoptosis ↑ ERK phosphorylation, graft survival | |
| Liver transplantation Cold I: 6 h-R: 4 h [50] | No activity changes in ERK | Angiotensin II receptor antagonists | AT1R: 5 mg/kg, oral, 24 and 1.5 h before surgery AT2R: 30 mg/kg, i.v., 5 min before surgery | ↓ Damage ↑ ERK phosphorylation, survival | |
| Liver transplantation Cold I: 18 h-R: 1, 3, 6, 24, 48 h [91] | ↑ ERK | Carbon monoxide | 100 ppm in recipient, 1 h before surgery and 1, 3, and 6 h after surgery | ↓ ERK phosphorylation, damage | |
| Liver transplantation Cold I: 30 h-R: 6, 24 h [113] | ↑ ERK | Sotraustaurin | 30 mg/kg, oral, 1) 90 min before organ recovery and 3 days in recipients and 2) 3 days in recipients | ↓ ERK phosphorylation, damage, caspase-3, apoptosis | |
| Mice | Isolated liver perfused Cold I: 24 h-R: 2 h [114] | No activity changes in ERK | Erythropoietin as additive | 10 IU/mL in solution | ↓ ERK phosphorylation, damage |
| Experimental Model | Changes in MAPK Induced by I-R | Treatment with MAPK Modulators | Effect of MAPK Modulators | ||
|---|---|---|---|---|---|
| Species | Surgical Procedure | Type of Liver | |||
| Human | Isolated Liver Perfused Cold I: >10 h SCS + 4h HMP R: 2 h [119] | Marginal (steatotic, not usable DCD, donor malignancy, failed allocation, atheromatosis, poor perfusion, rising liver function test) | ↑ MAPK | Hypothermic machine perfusion | No benefits on histological score. ↓ MAPK, AST, LDH |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Castro, M.B.; Cornide-Petronio, M.E.; Gracia-Sancho, J.; Casillas-Ramírez, A.; Peralta, C. Mitogen Activated Protein Kinases in Steatotic and Non-Steatotic Livers Submitted to Ischemia-Reperfusion. Int. J. Mol. Sci. 2019, 20, 1785. https://doi.org/10.3390/ijms20071785
Jiménez-Castro MB, Cornide-Petronio ME, Gracia-Sancho J, Casillas-Ramírez A, Peralta C. Mitogen Activated Protein Kinases in Steatotic and Non-Steatotic Livers Submitted to Ischemia-Reperfusion. International Journal of Molecular Sciences. 2019; 20(7):1785. https://doi.org/10.3390/ijms20071785
Chicago/Turabian StyleJiménez-Castro, Mónica B., María Eugenia Cornide-Petronio, Jordi Gracia-Sancho, Araní Casillas-Ramírez, and Carmen Peralta. 2019. "Mitogen Activated Protein Kinases in Steatotic and Non-Steatotic Livers Submitted to Ischemia-Reperfusion" International Journal of Molecular Sciences 20, no. 7: 1785. https://doi.org/10.3390/ijms20071785
APA StyleJiménez-Castro, M. B., Cornide-Petronio, M. E., Gracia-Sancho, J., Casillas-Ramírez, A., & Peralta, C. (2019). Mitogen Activated Protein Kinases in Steatotic and Non-Steatotic Livers Submitted to Ischemia-Reperfusion. International Journal of Molecular Sciences, 20(7), 1785. https://doi.org/10.3390/ijms20071785