Mitochondria Are Important Determinants of the Aging of Seeds




Abstract
:1. Aging Seeds
2. Mitochondrial Activity in Aging Seeds
3. Oxidative Damage in Aging Seed Mitochondria
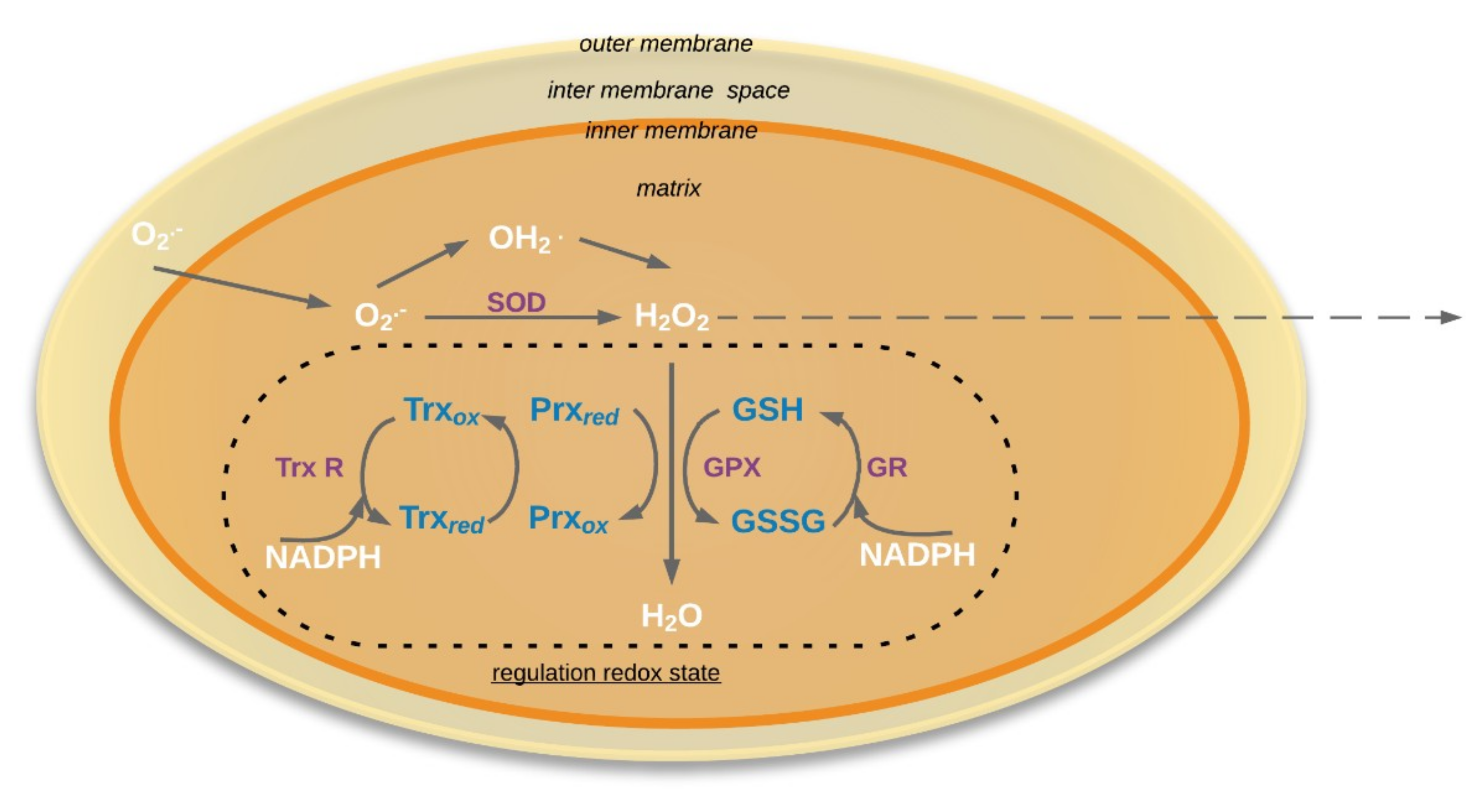
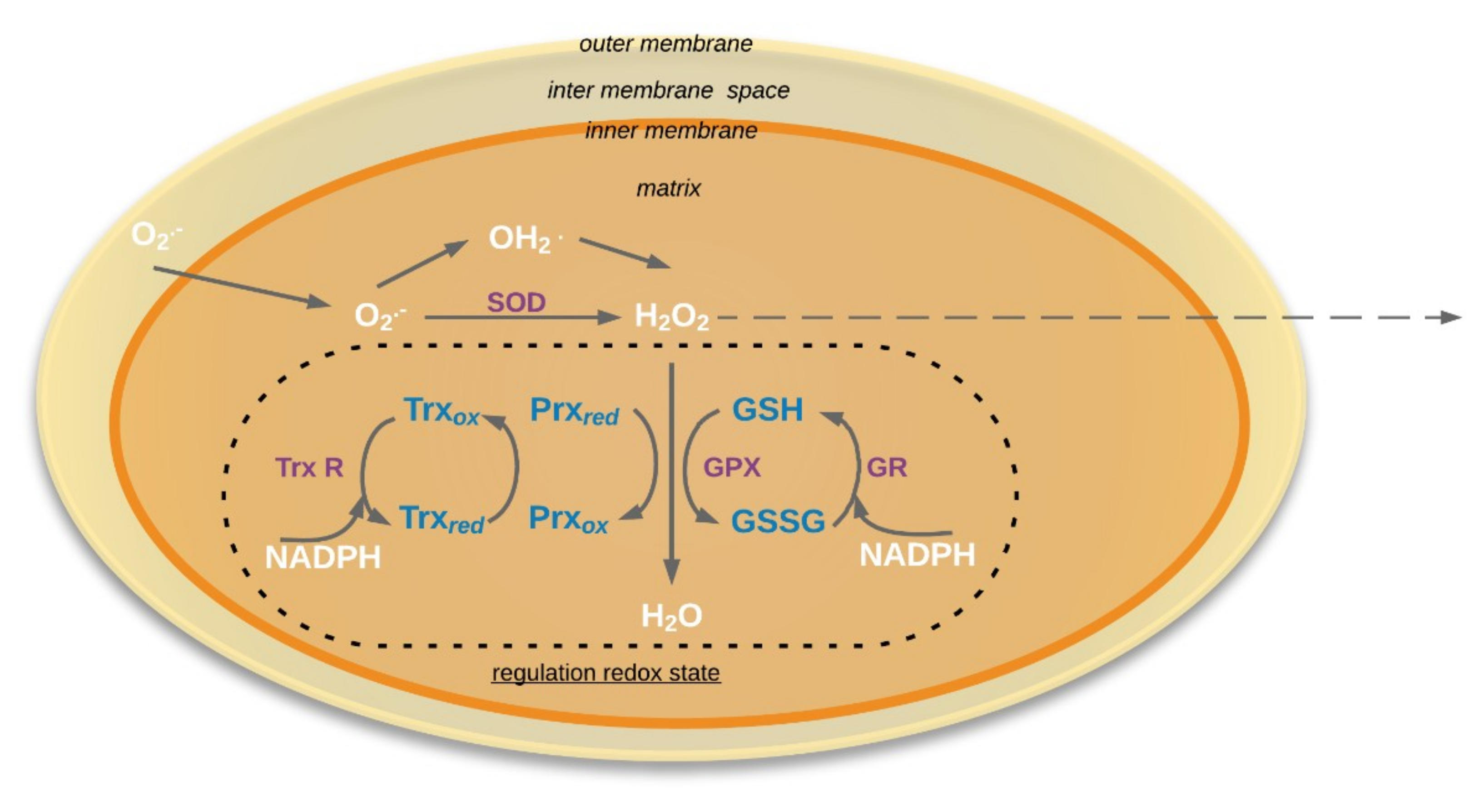
4. The Antioxidative System in Aging Seed Mitochondria
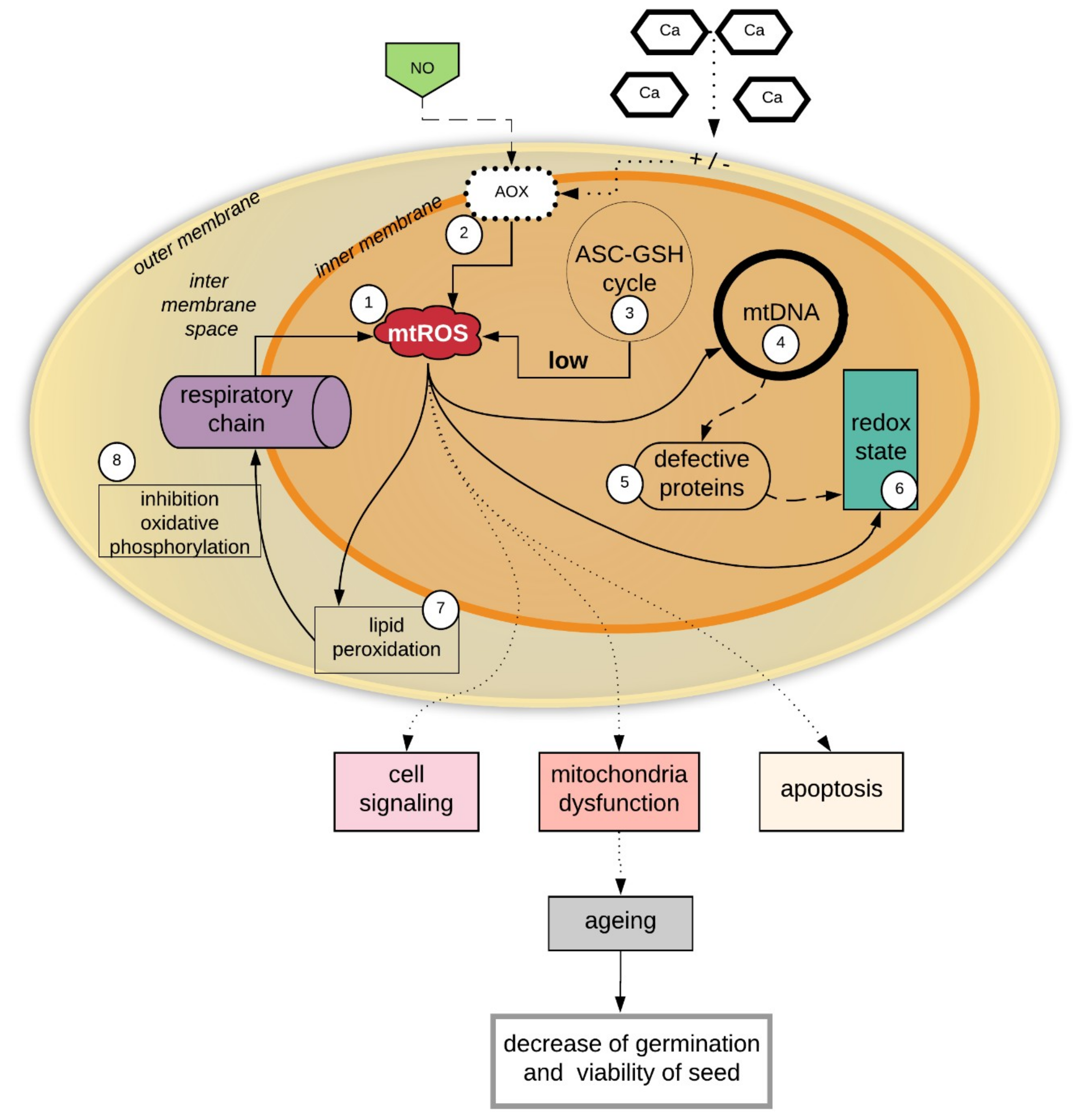
5. Is Mitochondrial Dysfunction the Cause of Seed Aging?
6. Mitochondrial Structure in the Process of Seed Aging
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.M.W.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy; Springer: New York, NY, USA, 2013. [Google Scholar]
- Groot, S.P.C.; Surki, A.A.; de Vos, R.C.H.; Kodde, J. Seed storage at elevated partial pressure of oxygen, a fast method for analysing seed aging under dry conditions. Ann. Bot. 2012, 110, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Walters, C.; Ballesteros, D.; Vertucci, V.A. Structural mechanics of seed deterioration: Standing the test of time. Plant Sci. 2010, 6, 565–573. [Google Scholar] [CrossRef]
- Pukacka, S.; Ratajczak, E. Age-related biochemical changes during storage of beech (Fagus sylvatica L.) seeds. Seed Sci. Res. 2007, 17, 45–53. [Google Scholar] [CrossRef]
- Ballesteros, D.; Walters, C. Detailed characterization of mechanical properties and molecular mobility within dry seed glasses: Relevance to the physiology of dry biological systems. Plant J. 2011, 68, 607–619. [Google Scholar] [CrossRef]
- Ratajczak, E.; Małecka, A.; Bagniewska-Zadworna, A.; Kalemba, E.M. The production, localization and spreading of reactive oxygen species contributes to the low vitality of long-term stored common beech (Fagus sylvatica L.) seeds. J. Plant Physiol. 2015, 174, 147–156. [Google Scholar] [CrossRef]
- Yin, G.K.; Whelan, J.; Wu, S.H.; Zhou, J.; Chen, B.; Chen, X.L.; Zhang, J.M.; He, X.; Xin, J.J.; Lu, X.X. Comprehensive mitochondrial metabolic shift during the critical node of seed aging in rice. PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: the dual role of reactive oxygen species in seed physiology. C RBiol. 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Rajjou, L.; Lovigny, Y.; Grott, S.P.C.; Belghazi, M.; Job, C.; Job, D. Proteome-wide characterization of seed aging in Arabidopsis. A comparison between artificial and natural aging protocols. Plant Physiol. 2008, 148, 20–41. [Google Scholar] [CrossRef]
- Bailly, C.; Kranner, I. Analyses of reactive oxygen species and antioxidants in relation to seed longevity and germination. Methods Mol. Biol. 2011, 773, 343–367. [Google Scholar] [PubMed]
- Bellani, M.; Salvini, L.; Dell’Aquila, A.; Scialabba, A. Reactive oxygen species release, vitamin E, fatty acid and phytosterol content of artificially aged radish (Raphanus sativus L.) seeds during germination. Acta. Physiol. Plant. 2012, 34, 1789–1799. [Google Scholar] [CrossRef]
- Hu, D.; Ma, G.; Wang, Q.; Yao, J.; Wang, Y.; Pritchard, H.W.; Wang, X. Spatial and temporal nature of reactive oxygen species production and programmed cell death in elm (Ulmus pumila L.) seeds during controlled deterioration. Plant Cell Environ. 2012, 35, 2045–2059. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Liu, L.; Gao, F.; Rampitsch, C.; Reinecke, D.M.; Ozga, J.A.; Ayele, B.T. Developmental and seed aging mediated regulation of antioxidative genes and differential expression of proteins during pre- and post-germinative phases in pea. J. Plant Physiol. 2012, 169, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.H.; Hong, D.; Roberts, E.H. An Intermediate Category of Seed Storage Behaviour?: I. Coffee. J. Exp. Bot. 1990, 41, 1167–1174. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed recalcitrance in perspective. Ann. Bot. Lond. 2008, 101, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M. Plant mitochondria and oxidative stress. Electron transport, NADPH turnover and metabolism of reactive oxygen species. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 561–569. [Google Scholar] [CrossRef]
- Howell, K.A.; Millar, A.H.; Whelan, J. Ordered assembly of mitochondria during rice germination begins with pro-mitochondrial structures rich in components of the protein import apparatus. Plant Mol. Biol. 2006, 60, 201–223. [Google Scholar] [CrossRef] [PubMed]
- Navrot, N.; Rouhier, N.; Gelhaye, E.; Jacquot, J.-P. ROS generation and antioxidant systems in plant mitochondria. Physiol. Plant. 2007, 129, 185–195. [Google Scholar] [CrossRef]
- Dunn, J.D.; Alvarez, L.A.J.; Zhang, X.; Soldati, T. Reactive oxygen species and mitochondria: A nexus of cellular homeostasis. Redox Biol. 2015, 6, 472–485. [Google Scholar] [CrossRef]
- Liberatore, K.L.; Dukowic-Schulze, S.; Miller, M.E.; Chen, C.; Kianian, S.F. The role of mitochondria in plant development and stress tolerance. Free Rad. Biol. Med. 2016, 100, 238–256. [Google Scholar] [CrossRef] [PubMed]
- Kowaltowski, A.J.; de Souza-Pinto, N.C.; Castilho, R.F.; Vercesi, A.E. Mitochondria and reactive oxygen species. Free Rad. Biol. Med. 2009, 47, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Lismont, C.; Nordgren, M.; VanVeldhoven, P.P.; Franse, M. Redox interplay between mitochondria and peroxisomes. Front Cell Dev. Biol. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Mrad, M.B.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Schjoerring, J.K.; Jahn, T.P. Membrane transport of hydrogen peroxide. Biochim. Biophys. Acta. 2006, 1758, 994–1003. [Google Scholar] [CrossRef]
- Barranco-Medina, S.; Krell, T.; Finkemeier, I.; Sevilla, F.; Lazaro, J.J.; Dietz, K.J. Biochemical and molecular characterization of the mitochondrial peroxiredoxin PsPrxII F from Pisum sativum. Plant Physiol. Biochem. 2007, 45, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Zhu, Y.; Cheng, H.; Yan, H.; Zhao, L.; Tang, J.; Ma, X.; Mao, P. Nitric oxide regulates seedling growth and mitochondrial responses in aged oat seeds. Int. J. Mol. Sci. 2018, 19, 1052. [Google Scholar] [CrossRef]
- Drahota, Z.; Chowdhury, S.K.R.; Floryk, D.; Mracek, T.; Wilhelm, J.; Rauchova, H. Glycerophosphate-dependent hydrogen peroxide production by brown adiposetissue mitochondria and its activation by ferricyanide. J. Bioenerg. Biomembr. 2002, 34, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Starkov, A.A.; Fiskum, G.; Chinopoulos, C.; Lorenzo, B.J.; Browne, S.E.; Patel, M.S. Mitochondrial alpha-ketoglutarate dehydrogenase complex generates reactive oxygen species. J. Neurosci. 2004, 24, 7779–7788. [Google Scholar]
- Jardim-Messeder, D.; Caverzan, A.; Rauber, R.; de Souza Ferreira, E.; Margis-Pinheiro, M.; Galina, A. Galina Succinate dehydrogenase (mitochondrial complex II) is a source of reactive oxygen species in plants and regulates development and stress responses. New Phytol. 2015, 208, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Holzerová, E.; Prokisch, H. Mitochondria: Much ado about nothing? How dangerous is reactive oxygen species production? Int. J. Biochem. Cell Biol. 2015, 63, 16–20. [Google Scholar] [CrossRef]
- Gupta, K.J. Protein S-nitrosylation in plants: photorespiratory metabolism and NO signaling. Sci. Signal. 2011, 154, jc1. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Igamberdiev, A.U. Reactive nitrogen species in mitochondria and their implications in plant energy status and hypoxic stress tolerance. Front Plant Sci. 2016, 7, 369. [Google Scholar] [CrossRef]
- Krasuska, U.; Ciacka, K.; Andryka, P.; Bogatek, R.; Gniazdowska, A. „Nitrosative door” in seed dormancy alleviation and germination. In Reactive oxygen and nitrogen species signaling and communication in plants; Gupta, K.J., Igamberdiev, A.U., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 215–237. [Google Scholar]
- Krasuska, U.; Ciacka, K.; Orzechowski, S.; Fettke, J.; Bogatek, R.; Gniazdowska, A. Modification of the endogenous NO level influences apple embryos dormancy by alterations of nitrated and biotinylated protein patterns. Planta 2016, 244, 877–891. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.; Granett, A.L. Deterioration of stored pea seed: Changes in germination, membrane permeability and ultrastructure resulting from infection by Aspergillus ruber and from aging. Physiol. Plant Path. 1972, 3, 271–276. [Google Scholar] [CrossRef]
- Hallam, N.D.; Roberts, B.E.; Osborne, D.J. Embryogenesis and germination in rye (Secale cereale L.)—III. Fine structure and biochemistry of the non-viable embryo. Planta 1973, 110, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Parrish, D.J.; Leopold, A.C. On the mechanism of aging in soybean seeds. Plant Physiol. 1978, 61, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Sun, H.; Xin, X.; Qin, G.; Liang, Z.; Jing, X. Mitochondrial damage in the soybean seed axis during imbibition at chilling temperatures. Plant Cell Physiol. 2009, 50, 1305–1318. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-Q.; Cheng, H.-Y.; Møller, I.M.; Song, S.-Q. The role of recovery of mitochondrial structure and function in desiccation tolerance of pea seeds. Physiol. Plant. 2012, 144, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Tian, Q.; Yin, G.; Chen, X.; Zhang, J.; Ng, S.; Lu, X. Reduced mitochondrial and ascorbate-glutathione activity after artificial aging in soybean seed. J. Plant Physiol. 2014, 171, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Xue, H.; Pritchard, H.W.; Wang, X. Reactive oxygen species—provoked mitochondria—dependent cell death during aging of elm (Ulmus pumila L.) seeds. Plant J. 2015, 81, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.S.; Wang, M.Y.; Li, M.L.; Mao, P.S. Mitochondrial structural and antioxidant system responses to aging in oat (Avena sativa L.) seeds with different moisture contents. Plant Physiol. Biochem. 2015, 94, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Fu, H.; Zhou, X.; Chen, Z.; Luo, Y.; Cui, B.; Chen, G.; Liu, J. Comparative proteomic analysis of seed embryo proteins associated with seed storability in rice (Oryza sativa L.) during natural aging. Plant Physiol. Biochem. 2016, 103, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Xue, H.; Pritchard, H.W.; Wang, X. Changes in the mitochondrial protein profile due to ROS eruption during aging of elm (Ulmus pumila L.) seeds. Plant Physiol Biochem. 2017, 114, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Osuna, D.; Colville, L.; Lorenzo, O.; Graeber, K.; Dennis, E.S.; Peacock, W.J. Transcriptome-wide mapping of pea seed aging reveals a pivotal role for genes related to oxidative stress and programmed cell death. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Rhoads, D.M.; Umbach, A.L.; Subbaiah, C.C.; Siedow, J.N. Mitochondrial reactive oxygen species. Contribution to oxidative stress and interorganellar signaling. Plant Physiol. 2006, 141, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Prabhakaran, S.A. Mechanism, measurement and prevention of oxidative stress in male reproductive physiology. Indian J. Exp. Biol. 2005, 43, 963–974. [Google Scholar] [PubMed]
- Bykova, N.V.; Egsgaard, H.; Møller, I.M. Identification of 14 new phosphoproteins involved in important plant mitochondrial processes. FEBS Lett. 2003, 540, 141–146. [Google Scholar] [CrossRef]
- Ito, J.; Heazlewood, J.L.; Millar, A.H. The plant mitochondrial proteome and the challenge of defining the post-translational modifications responsible for signalling and stress effects on respiratory functions. Physiol. Plant. 2007, 129, 207–224. [Google Scholar] [CrossRef]
- Winger, A.M.; Taylor, N.L.; Heazlewood, J.H.; Day, D.A.; Millar, A.H. The cytotoxic lipid peroxidation product 4-hydroxy-2-nomenal covalently modifies a selective range of proteins linked to respiratory function in plant mitochondria. J. Biol. Chem. 2007, 282, 37436–37447. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, B.K.; Askerlund, P.; Bykova, N.V.; Egsgaard, H.; Møller, I.M. Identification of oxidised proteins in the matrix of rice leaf mitochondria by immunoprecipitation and two-dimensional liquid chromatography-tandem mass spectrometry. Phytochemistry 2004, 65, 1839–1851. [Google Scholar] [CrossRef] [PubMed]
- Moller, I.M.; Kristensen, B.K. Protein oxidation in plant mitochondria as a stress indicator. Photochem. Photobiol. Sci. 2004, 3, 730–735. [Google Scholar] [CrossRef]
- Shokolenko, I.; Venediktova, N.; Bochkareva, A.; Wilson, G.L.; Alexeyev, M.F. Oxidative stress induces degradation of mitochondrial DNA. Nucleic Acids Res. 2009, 37, 2539–2548. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, RP.; Li, L.; Huang, S.; Pong Lee, C.; Millar, A.H.; Taylor, N.L. Mitochondrial composition, function and stress response in plants. J. Integr. Plant Biol. 2012, 11, 887–906. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Li, W.; He, H.Y.; Li, C.Z.; He, L.F. Mitochondrial alterations during Al-induced PCD in peanut root tips. Plant Physiol. Biochem. 2014, 7, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Free radical theory of aging: an update: increasing the functional lifespan. Ann. N. Y. Acad. Sci. 2006, 1067, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Oelze, M.L.; Kandlbinder, A.; Dietz, K.J. Redox regulation and overreduction control in the photosynthesizing cell: Complexity in redox regulatory networks. Bioch. Bioph. Acta. 2008, 1780, 1261–1272. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, E.; Dietz, K.J. Regulatory redox state in tree seeds. Acta Soc. Bot. Poloniae 2017, 86, 35–67. [Google Scholar] [CrossRef]
- Gama, F.; Keech, O.; Eymery, F.; Finkemeier, I.; Gelhaye, E.; Gardeströ>m, P.; Dietz, K.J.; Rey, P.; Jacquot, J.-P.; Rouhie, N. The mitochondrial type II peroxiredoxin from poplar. Physiol. Plant 2007, 129, 196–206. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Kibinza, S.; Bazin, J.; Bailly, C.; Farrant, J.M.; Corbineau, F.; El-Maarouf-Bouteau, H. Catalase is a key enzyme in seed recovery from aging during priming. Plant Sci. 2011, 181, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 9, 405–410. [Google Scholar] [CrossRef]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot. 2002, 372, 1237–1247. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox regulation and photosynthetic organisms: Signaling, acclimation and practical implications. Antiox. Redox. Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Talla, S.; Riazunnisa, K.; Padmavathi, L.; Sunil, B.; Rajsheel, P.; Raghavendra, A.S. Ascorbic acid is a key participant during the interactions between chloroplasts and mitochondria to optimize photosynthesis and protect against photoinhibition. J. Biosci. 2011, 36, 163–173. [Google Scholar] [CrossRef]
- Kerchev, P.I.; Pellny, T.K.; Vivancos, P.D.; Kiddle, G.; Hedden, P.; Driscoll, S.; Vanacker, H.; Verrier, P.; Hancock, R.D.; Foyer, C.H. The transcription factor ABI4 Is required for the ascorbic acid-dependent regulation of growth and regulation of jasmonate-dependent defense signaling pathways in Arabidopsis. Plant Cell. 2011, 23, 3319–3334. [Google Scholar] [CrossRef] [PubMed]
- Zechmann, B. Subcellular distribution of ascorbate in plants. Plant Sign. Behav. 2011, 6, 360–363. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic Acid-A potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front Plant Sci. 2017, 26, 613. [Google Scholar] [CrossRef] [PubMed]
- Schwarzländer, M.; Finkemeier, I. Mitochondrial energy and redox signaling in plants. Antioxid. Redox. Signal. 2013, 18, 2122–2144. [Google Scholar] [CrossRef] [PubMed]
- Chew, O.; Whelan, J.; Millar, A.H. Molecular definition of the ascorbate-glutathione cycle in Arabidopsis mitochondria reveals dual targeting of antioxidant defenses in plants. J. Biol. Chem. 2003, 278, 46869–46877. [Google Scholar] [CrossRef]
- Mailloux, R.J.; Jin, X.; Willmore, W.G. Redox regulation of mitochondrial function with emphasis on cysteine oxidation reactions. Redox Biol. 2013, 19, 123–139. [Google Scholar] [CrossRef]
- Barranco-Medina, S.; Kakorin, S.; Lázaro, J.J.; Dietz, K.J. Thermodynamics of the dimer–decamer transition of reduced human and plant 2-Cys peroxiredoxin. Biochemistry 2008, 47, 7196–7204. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J. Peroxiredoxins in plants and cyanobacteria. Antiox. Redox Sign. 2011, 15, 1129–1159. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, E.; Dietz, K.J.; Kalemba, E.M. The occurrence of peroxiredoxins and changes in redox state in Acer platanoides and Acer pseudoplatanus during seed development. J. Plant Grow. Regul. 2018. [Google Scholar] [CrossRef]
- Finkemeier, I.; Goodman, M.; Lamkemeyer, P.; Kandlbinder, A.; Sweetlove, L.J.; Dietz, K.J. The mitochondrial type II peroxiredoxin F is essential for redox homeostasis and root growth of Arabidopsis thaliana under stress. J. Biol. Chem. 2005, 280, 12168–12180. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N.; Jacquot, J.P. The plant multigenic family of thiol peroxidases. Free Radic. Biol. Med. 2005, 38, 1413–1421. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, E.; Ströher, E.; Oelze, M.L.; Kalemba, E.M.; Pukacka, S.; Dietz, K.J. The involvement of the mitochondrial peroxiredoxin PrxIIF in defining physiological differences between orthodox and recalcitrant seeds of two Acer species. Funct. Plant Biol. 2013, 40, 1005–1017. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Balmer, Y. Redox regulation: A broadening horizon. Ann. Rev. Plant Biol. 2005, 56, 187–220. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Noguchi, K.; Motohash, K.; Hisabori, T. Systematic exploration of thioredoxin target proteins in plant mitochondria. Plant Cell Physiol. 2013, 54, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N.; Cerveau, D.; Couturier, J.; Reichheld, J.P.; Rey, P. Involvement of thiol-based mechanisms in plant development. Biochim. Biophys. Acta. 2015, 1850, 1479–1496. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, F.; Camejo, D.; Ortiz-Espín, A.; Calderón, A.; Lázaro, J.J.; Jiménez, A. The thioredoxin/peroxiredoxin/sulfiredoxin system: current overview on its redox function in plants and regulation by reactive oxygen and nitrogen species. J. Exp. Bot. 2015, 66, 2945–2955. [Google Scholar] [CrossRef] [PubMed]
- Shahpiri, A.; Svensson, B.; Finni, Ch. From Proteomics to Structural Studies of Cytosolic/Mitochondrial-Type Thioredoxin Systems in Barley Seeds. Mol. Plant 2009, 2, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Cai, N.; Tanaka, C.K.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin reduction alters the solubility of proteins of wheat starchy endosperm: An early event in cereal germination. Plant Cell Physiol. 2004, 45, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Gelhaye, E.; Rouhier, N.; Jacquot, J.P. The thioredoxin h system of higher plants. Plant Physiol. Biochem. 2004, 42, 265–271. [Google Scholar] [CrossRef]
- Balmer, Y.; Vensel, W.H.; Tanaka, C.K.; Hurkman, W.J.; Schurmann, P.; Droux, M.; et al. Thioredoxin links redox to the regulation of fundamental processes of plant mitochondria. PNAS 2004, 101, 2642–2647. [Google Scholar] [CrossRef]
- Sanz-Barrio, R.; Fernández-San Millán, A.; Carballeda, J.; Corral-Martínez, P.; Seguí-Simarro, J.M.; Farran, I. Chaperone-like properties of tobacco plastid thioredoxins f and m. J. Exp. Bot. 2012, 63, 365–379. [Google Scholar] [CrossRef]
- Hill, S.; Van Remmen, H. Mitochondrial stress signaling in longevity: A new role for mitochondrial function in aging. Redox Biol. 2014, 27, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Youle R., J.; Finkel, T. The mitochondrial basis of aging. Mol. Cell 2016, 61, 654–666. [Google Scholar] [CrossRef]
- Gredilla, R.; Bohr, V.A.; Stevnsner, T. Mitochondrial DNA repair and association with aging—An update. Exp. Gerontol. 2010, 45, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, W.M.; Bray, C.M.; West, C.E. The importance of safeguarding genome integrity in germination and seed longevity. J. Exp. Bot. 2015, 66, 3549–3558. [Google Scholar] [CrossRef] [PubMed]
- Paszkiewicz, G.; Gualberto, J.M.; Benamar, A.; Macherel, D.; Logana, D.C. Arabidopsis seed mitochondria are bioenergetically active immediately upon imbibition and specialize via biogenesis in preparation for autotrophic growth. Plant Cell. 2017, 29, 109–128. [Google Scholar] [CrossRef]
- Fu, B.F.; Ahmed, Z.; Diederichsen, A. Towards a better monitoring of seed aging under ex situ seed conservation. Conserv. Physiol. 2015, 3, cov026. [Google Scholar] [CrossRef] [PubMed]
- Madreiter-Sokolowski, C.T.; Waldeck-Weiermair, M.; Bourguignon, M.P.; Villeneuve, N.; Gottschalka, B.; Klec, Ch.; Stryeck, S.; Radulovic, S.; Parichatikan, W.; Madl, T.; et al. Enhanced inter-compartmental Ca2+ flux modulates mitochondrial metabolism and apoptotic threshold during aging. Redox. Biol. 2019, 20, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Berkowitz, O.; Selinski, J.; Xu, Y.; Hartmann, A.; Whelan, J. Stress responsive mitochondrial proteins in Arabidopsis thaliana. Free Radic. Biol. Med. 2018, 122, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, S.; Enriquez, J.A.; Scorrano, L. Mitochondrial Cristae: Where beauty meets functionality. Trends Biochem. Sci. 2018, 41, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Paepe, R.; Foyer, C.H. Mitochondrial redox biology and homeostasis in plants. Trends Plant Sci. 2007, 12, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Daum, B.; Walter, A.; Horst, A.; Osiewacz, H.D.; Kühlbrandt, W. Age-dependent dissociation of ATP synthase dimers and loss of inner-membrane cristae in mitochondria. PNAS 2013, 110, 15301–15306. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Organism (Species) | Processes | References |
|---|---|---|
| Pea (Pisum sativum L.) | Germination after Aspergillus ruber infection, aging | [35] |
| Rye (Secale cereal L.) | Embryogenesis, germination | [36] |
| Soybean [Glycine max (L.) Merr] | Aging | [37] |
| Pea (Pisum sativum) | Germination | [25] |
| Soybean [Glycine max (L.) Merr] | Germination, imbibition | [38] |
| Pea (Pisum sativum cv. Jizhuang) | Germination | [39] |
| Soybean [G.max (L.) Merr] | Aging | [40] |
| Elm (Ulmus pumila L.) | Aging | [41] |
| Oat (Avena sativa L.) | Aging | [42] |
| Rice (Oryza sativa L.) | Aging | [7] |
| Rice (Oryza sativa L.) | Aging | [43] |
| Elm (Ulmus pumila L.) | Aging | [44] |
| Oat (Avena sativa L.) | Aging | [26] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ratajczak, E.; Małecka, A.; Ciereszko, I.; Staszak, A.M. Mitochondria Are Important Determinants of the Aging of Seeds. Int. J. Mol. Sci. 2019, 20, 1568. https://doi.org/10.3390/ijms20071568
Ratajczak E, Małecka A, Ciereszko I, Staszak AM. Mitochondria Are Important Determinants of the Aging of Seeds. International Journal of Molecular Sciences. 2019; 20(7):1568. https://doi.org/10.3390/ijms20071568
Chicago/Turabian StyleRatajczak, Ewelina, Arleta Małecka, Iwona Ciereszko, and Aleksandra M. Staszak. 2019. "Mitochondria Are Important Determinants of the Aging of Seeds" International Journal of Molecular Sciences 20, no. 7: 1568. https://doi.org/10.3390/ijms20071568
APA StyleRatajczak, E., Małecka, A., Ciereszko, I., & Staszak, A. M. (2019). Mitochondria Are Important Determinants of the Aging of Seeds. International Journal of Molecular Sciences, 20(7), 1568. https://doi.org/10.3390/ijms20071568