Cytotoxicity of Human Hepatic Intrasinusoidal CD56bright Natural Killer Cells against Hepatocellular Carcinoma Cells

Abstract
1. Introduction
2. Results
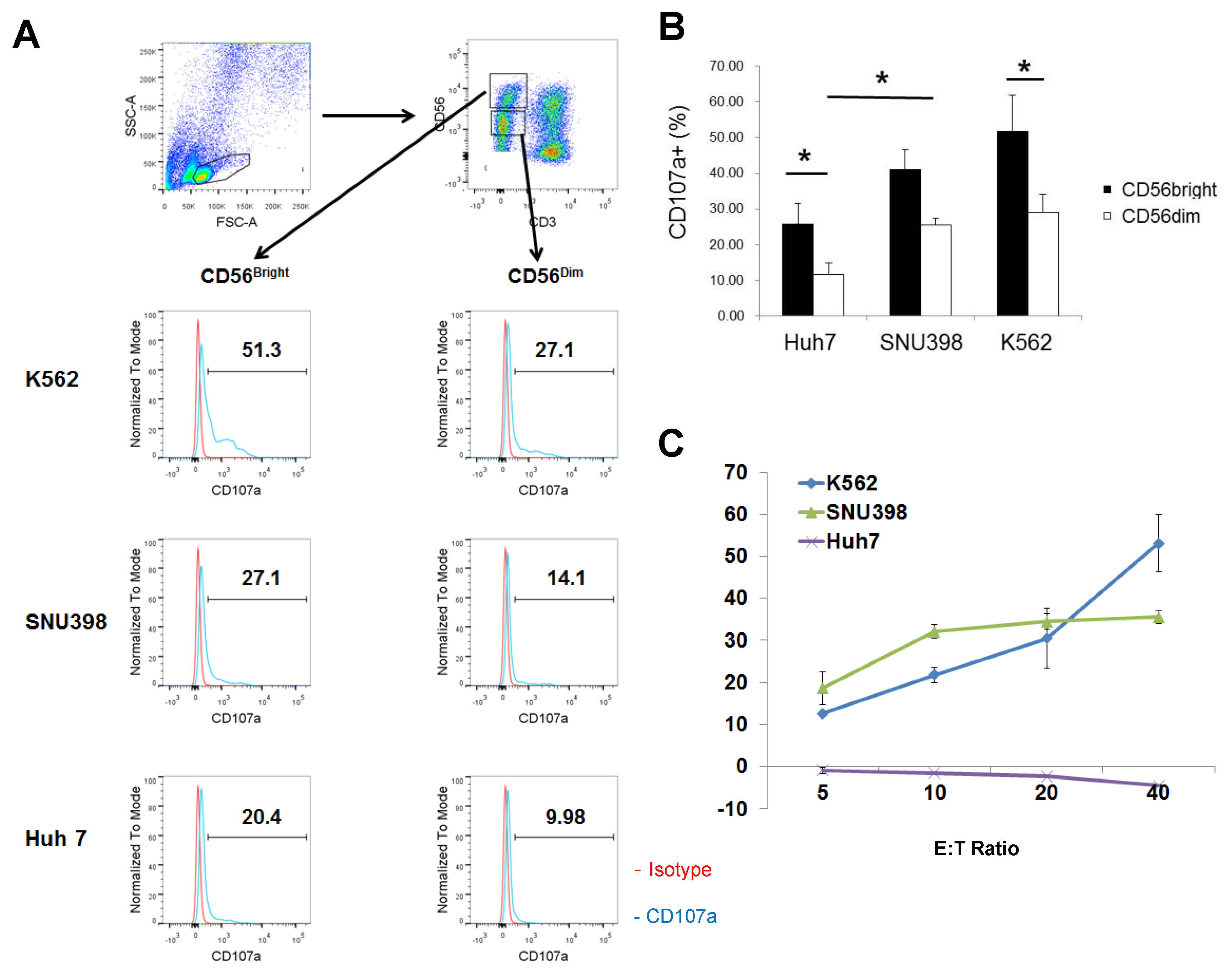
2.1. Strong Cytotoxicity of CD56bright HI NK Cells
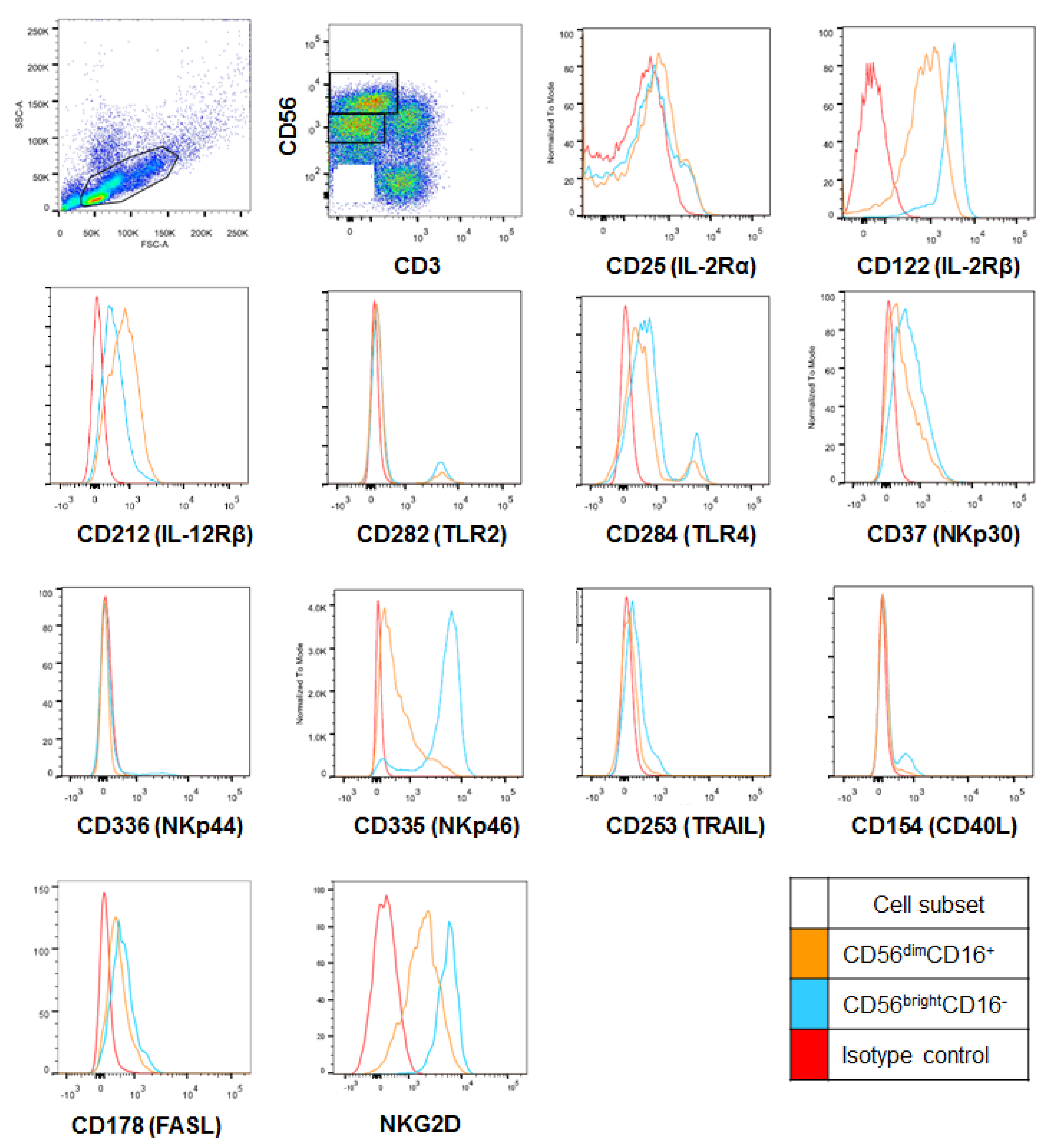
2.2. CD56bright HI NK Cells Express Cytotoxicity Receptors at Higher Levels
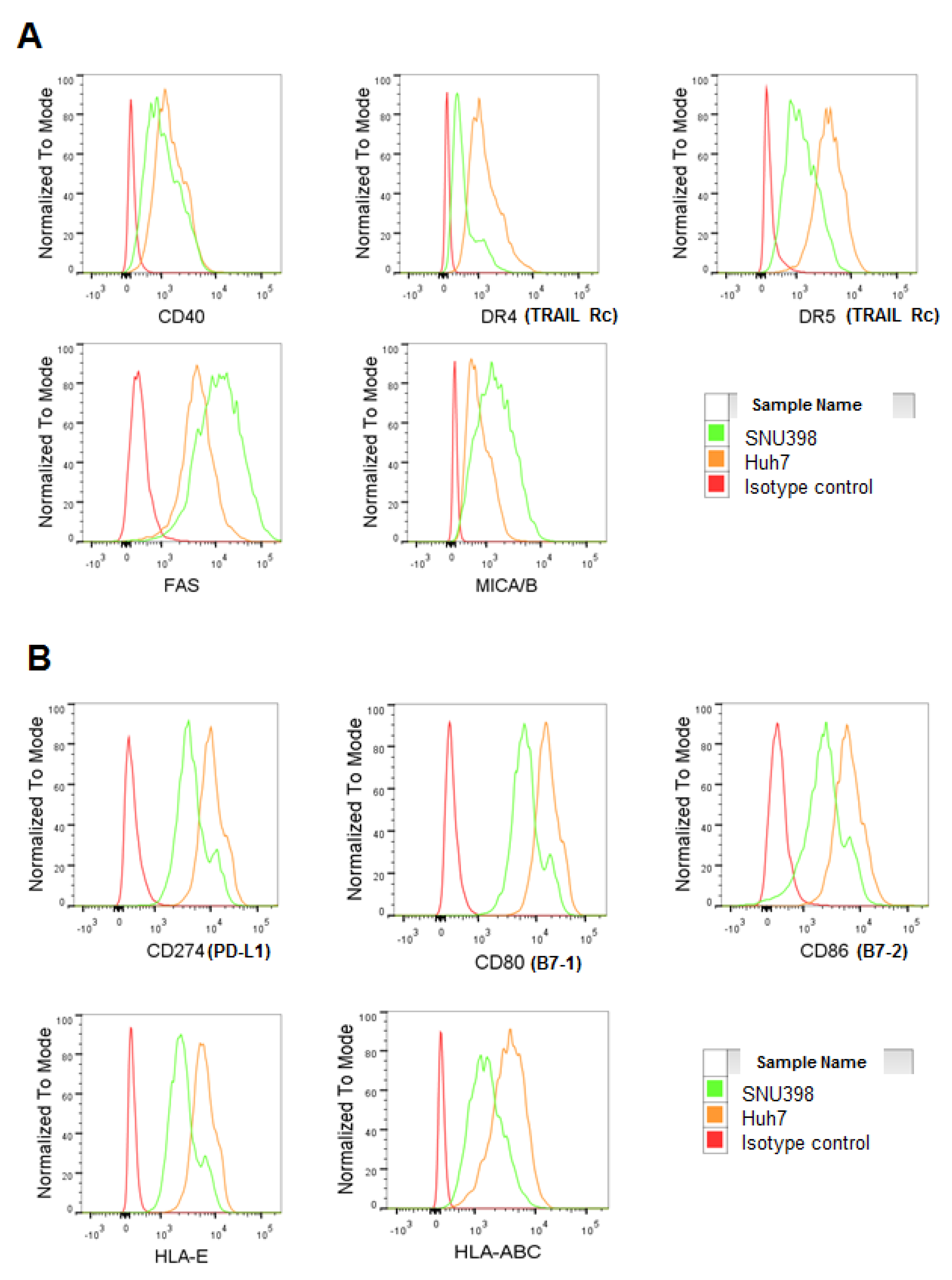
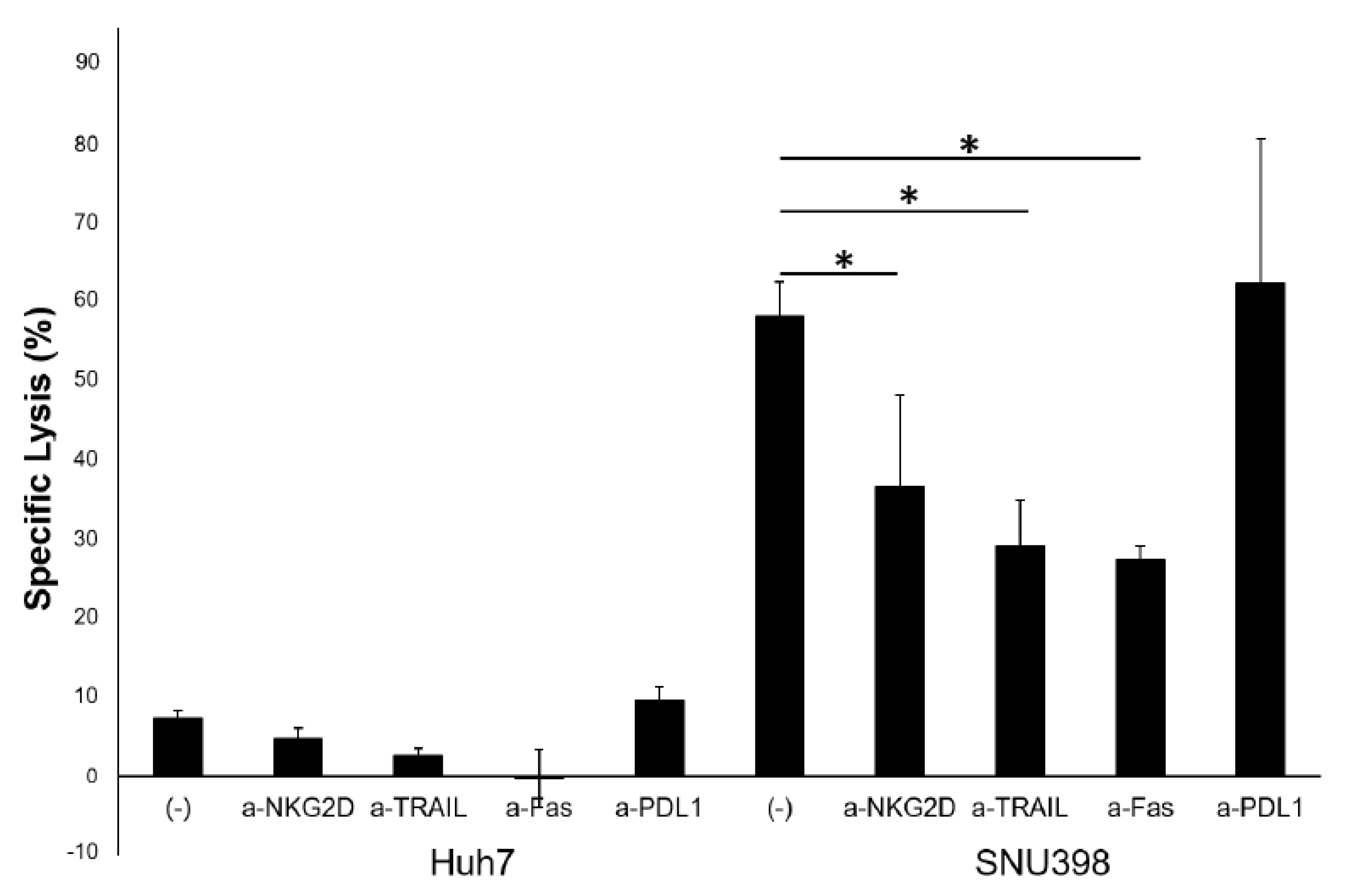
2.3. HI NK Cells Kill HCC Cells through NKG2D Ligands and FAS on HCC Cells
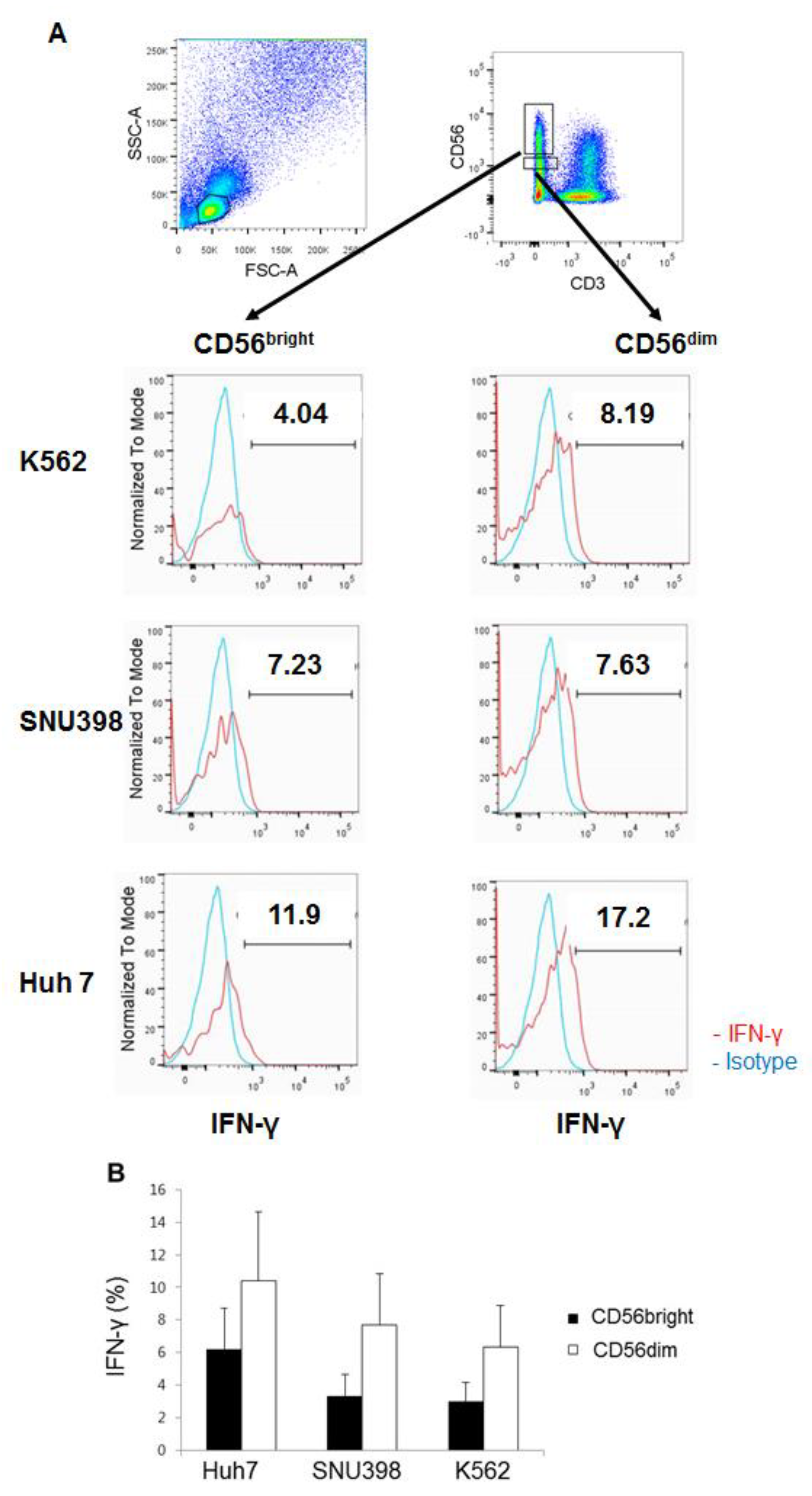
2.4. HI NK Cells Produce IFN-γ Well in Response to Huh7 Cells
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Flow Cytometry
4.3. Lactate Dehydrogenase (LDH) Assay
4.4. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BTLA | B- and T-lymphocyte attenuator |
| CTLA-4 | cytotoxic T-lymphocyte-associated antigen 4 |
| FASL | FAS ligand |
| HCC | hepatocellular carcinoma |
| HI | Hepatic intrasinusoidal |
| ILT2 | immunoglobulin-like transcript 2 |
| IFN-γ | interferon-γ |
| KIRs | killer-cell immunoglobulin-like receptors |
| LDH | lactate dehydrogenase |
| NK | natural killer |
| PB | peripheral blood |
| PD-1 | programmed cell death protein 1 |
| TRAIL | TNF-related apoptosis-inducing ligand |
| TNF-α | tumor necrosis factor-α |
References
- Kagi, D.; Ledermann, B.; Burki, K.; Seiler, P.; Odermatt, B.; Olsen, K.J.; Podack, E.R.; Zinkernagel, R.M.; Hengartner, H. Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice. Nature 1994, 369, 31–37. [Google Scholar] [CrossRef]
- Heusel, J.W.; Wesselschmidt, R.L.; Shresta, S.; Russell, J.H.; Ley, T.J. Cytotoxic lymphocytes require granzyme B for the rapid induction of DNA fragmentation and apoptosis in allogeneic target cells. Cell 1994, 76, 977–987. [Google Scholar] [CrossRef]
- Zamai, L.; Ahmad, M.; Bennett, I.M.; Azzoni, L.; Alnemri, E.S.; Perussia, B. Natural killer (NK) cell-mediated cytotoxicity: Differential use of TRAIL and Fas ligand by immature and mature primary human NK cells. J. Exp. Med. 1998, 188, 2375–2380. [Google Scholar] [CrossRef] [PubMed]
- Long, E.O.; Kim, H.S.; Liu, D.; Peterson, M.E.; Rajagopalan, S. Controlling natural killer cell responses: Integration of signals for activation and inhibition. Annu. Rev. Immunol. 2013, 31, 227–258. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Kim, H.S. Targeting Checkpoint Receptors and Molecules for Therapeutic Modulation of Natural Killer Cells. Front. Immunol. 2018, 9, 2041. [Google Scholar] [CrossRef]
- Stojanovic, A.; Fiegler, N.; Brunner-Weinzierl, M.; Cerwenka, A. CTLA-4 is expressed by activated mouse NK cells and inhibits NK Cell IFN-gamma production in response to mature dendritic cells. J. Immunol. 2014, 192, 4184–4191. [Google Scholar] [CrossRef]
- Pesce, S.; Greppi, M.; Tabellini, G.; Rampinelli, F.; Parolini, S.; Olive, D.; Moretta, L.; Moretta, A.; Marcenaro, E. Identification of a subset of human natural killer cells expressing high levels of programmed death 1: A phenotypic and functional characterization. J. Allergy Clin. Immunol. 2017, 139, 335–346.e3. [Google Scholar] [CrossRef]
- Kwon, H.J.; Kim, N.; Kim, H.S. Molecular checkpoints controlling natural killer cell activation and their modulation for cancer immunotherapy. Exp. Mol. Med. 2017, 49, e311. [Google Scholar] [CrossRef]
- Takayama, T.; Sekine, T.; Makuuchi, M.; Yamasaki, S.; Kosuge, T.; Yamamoto, J.; Shimada, K.; Sakamoto, M.; Hirohashi, S.; Ohashi, Y.; et al. Adoptive immunotherapy to lower postsurgical recurrence rates of hepatocellular carcinoma: A randomised trial. Lancet 2000, 356, 802–807. [Google Scholar] [CrossRef]
- Weng, D.S.; Zhou, J.; Zhou, Q.M.; Zhao, M.; Wang, Q.J.; Huang, L.X.; Li, Y.Q.; Chen, S.P.; Wu, P.H.; Xia, J.C. Minimally invasive treatment combined with cytokine-induced killer cells therapy lower the short-term recurrence rates of hepatocellular carcinomas. J. Immunother. 2008, 31, 63–71. [Google Scholar] [CrossRef]
- Jinushi, M.; Takehara, T.; Tatsumi, T.; Kanto, T.; Groh, V.; Spies, T.; Kimura, R.; Miyagi, T.; Mochizuki, K.; Sasaki, Y.; et al. Expression and role of MICA and MICB in human hepatocellular carcinomas and their regulation by retinoic acid. Int. J. Cancer 2003, 104, 354–361. [Google Scholar] [CrossRef]
- Cao, W.; Xi, X.; Hao, Z.; Li, W.; Kong, Y.; Cui, L.; Ma, C.; Ba, D.; He, W. RAET1E2, a soluble isoform of the UL16-binding protein RAET1E produced by tumor cells, inhibits NKG2D-mediated NK cytotoxicity. J. Biol. Chem. 2007, 282, 18922–18928. [Google Scholar] [CrossRef]
- Kamimura, H.; Yamagiwa, S.; Tsuchiya, A.; Takamura, M.; Matsuda, Y.; Ohkoshi, S.; Inoue, M.; Wakai, T.; Shirai, Y.; Nomoto, M.; et al. Reduced NKG2D ligand expression in hepatocellular carcinoma correlates with early recurrence. J. Hepatol. 2012, 56, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Sha, W.H.; Zeng, X.H.; Min, L. The correlation between NK cell and liver function in patients with primary hepatocellular carcinoma. Gut Liver 2014, 8, 298–305. [Google Scholar] [CrossRef]
- Zou, Y.; Chen, T.; Han, M.; Wang, H.; Yan, W.; Song, G.; Wu, Z.; Wang, X.; Zhu, C.; Luo, X.; et al. Increased killing of liver NK cells by Fas/Fas ligand and NKG2D/NKG2D ligand contributes to hepatocyte necrosis in virus-induced liver failure. J. Immunol. 2010, 184, 466–475. [Google Scholar] [CrossRef]
- Sheppard, S.; Guedes, J.; Mroz, A.; Zavitsanou, A.M.; Kudo, H.; Rothery, S.M.; Angelopoulos, P.; Goldin, R.; Guerra, N. The immunoreceptor NKG2D promotes tumour growth in a model of hepatocellular carcinoma. Nat. Commun. 2017, 8, 13930. [Google Scholar] [CrossRef]
- Ishiyama, K.; Ohdan, H.; Ohira, M.; Mitsuta, H.; Arihiro, K.; Asahara, T. Difference in cytotoxicity against hepatocellular carcinoma between liver and periphery natural killer cells in humans. Hepatology 2006, 43, 362–372. [Google Scholar] [CrossRef]
- Kim, H.R.; Park, H.J.; Park, J.H.; Kim, S.J.; Kim, K.; Kim, J. Characteristics of the killing mechanism of human natural killer cells against hepatocellular carcinoma cell lines HepG2 and Hep3B. Cancer Immunol. Immunother. 2004, 53, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Wisse, E.; Tian, Z. Liver natural killer cells: Subsets and roles in liver immunity. Cell. Mol. Immunol. 2016, 13, 328. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, P.; Wang, G.; Liu, X.; Sun, X.; Zhang, X.; Li, H.; Qi, J.; Ding, L.; Li, T.; et al. Description of organ-specific phenotype, and functional characteristics of tissue resident lymphocytes from liver transplantation donor and research on immune tolerance mechanism of liver. Oncotarget 2018, 9, 15552–15565. [Google Scholar] [CrossRef]
- Jonsson, J.R.; Hogan, P.G.; Balderson, G.A.; Ooi, L.L.; Lynch, S.V.; Strong, R.W.; Powell, E.E. Human liver transplant perfusate: An abundant source of donor liver-associated leukocytes. Hepatology 1997, 26, 1111–1114. [Google Scholar] [PubMed]
- Moroso, V.; Metselaar, H.J.; Mancham, S.; Tilanus, H.W.; Eissens, D.; van der Meer, A.; van der Laan, L.J.; Kuipers, E.J.; Joosten, I.; Kwekkeboom, J. Liver grafts contain a unique subset of natural killer cells that are transferred into the recipient after liver transplantation. Liver Transpl. 2010, 16, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Ohira, M.; Ishiyama, K.; Tanaka, Y.; Doskali, M.; Igarashi, Y.; Tashiro, H.; Hiraga, N.; Imamura, M.; Sakamoto, N.; Asahara, T.; et al. Adoptive immunotherapy with liver allograft-derived lymphocytes induces anti-HCV activity after liver transplantation in humans and humanized mice. J. Clin. Investig. 2009, 119, 3226–3235. [Google Scholar] [CrossRef]
- Tashiro, H.; Ishiyama, K.; Ohira, M.; Igarashi, Y.; Tahara, H.; Ide, K.; Onoe, T.; Tanaka, Y.; Ohdan, H. Impact of adjuvant immunotherapy using liver allograft-derived lymphocytes on bacteremia in living-donor liver transplantation. Transplantation 2011, 92, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Levi, D.M.; Tzakis, A.G. Liver natural killer cell inoculum for liver transplantation with hepatocellular carcinoma. Curr. Opin. Organ Transplant. 2013, 18, 690–694. [Google Scholar] [CrossRef]
- Kim, N.; Park, P.J.; Jung, M.K.; Song, G.W.; Jung, D.H.; Lee, S.G.; Ahn, C.S.; Hwang, S. Differential function of natural killer cells in the liver graft perfusate of Korean population. Transplant. Proc. 2013, 45, 2886–2891. [Google Scholar] [CrossRef]
- Harmon, C.; Robinson, M.W.; Fahey, R.; Whelan, S.; Houlihan, D.D.; Geoghegan, J.; O’Farrelly, C. Tissue-resident Eomes(hi) T-bet(lo) CD56(bright) NK cells with reduced proinflammatory potential are enriched in the adult human liver. Eur. J. Immunol. 2016, 46, 2111–2120. [Google Scholar] [CrossRef]
- Stegmann, K.A.; Robertson, F.; Hansi, N.; Gill, U.; Pallant, C.; Christophides, T.; Pallett, L.J.; Peppa, D.; Dunn, C.; Fusai, G.; et al. CXCR6 marks a novel subset of T-bet(lo)Eomes(hi) natural killer cells residing in human liver. Sci. Rep. 2016, 6, 26157. [Google Scholar]
- Marquardt, N.; Beziat, V.; Nystrom, S.; Hengst, J.; Ivarsson, M.A.; Kekalainen, E.; Johansson, H.; Mjosberg, J.; Westgren, M.; Lankisch, T.O.; et al. Cutting edge: Identification and characterization of human intrahepatic CD49a+ NK cells. J. Immunol. 2015, 194, 2467–2471. [Google Scholar] [CrossRef]
- Takeda, K.; Cretney, E.; Hayakawa, Y.; Ota, T.; Akiba, H.; Ogasawara, K.; Yagita, H.; Kinoshita, K.; Okumura, K.; Smyth, M.J. TRAIL identifies immature natural killer cells in newborn mice and adult mouse liver. Blood 2005, 105, 2082–2089. [Google Scholar] [CrossRef]
- Gordon, S.M.; Chaix, J.; Rupp, L.J.; Wu, J.; Madera, S.; Sun, J.C.; Lindsten, T.; Reiner, S.L. The transcription factors T-bet and Eomes control key checkpoints of natural killer cell maturation. Immunity 2012, 36, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.A.; Rosario, M.; Romee, R.; Berrien-Elliott, M.M.; Schneider, S.E.; Leong, J.W.; Sullivan, R.P.; Jewell, B.A.; Becker-Hapak, M.; Schappe, T.; et al. CD56bright NK cells exhibit potent antitumor responses following IL-15 priming. J. Clin. Investig. 2017, 127, 4042–4058. [Google Scholar] [CrossRef] [PubMed]
- Kenna, T.; Golden-Mason, L.; Norris, S.; Hegarty, J.E.; O’Farrelly, C.; Doherty, D.G. Distinct subpopulations of gamma delta T cells are present in normal and tumor-bearing human liver. Clin. Immunol. 2004, 113, 56–63. [Google Scholar] [CrossRef]
- Prajapati, K.; Perez, C.; Rojas, L.B.P.; Burke, B.; Guevara-Patino, J.A. Functions of NKG2D in CD8+ T cells: An opportunity for immunotherapy. Cell. Mol. Immunol. 2018, 15, 470–479. [Google Scholar] [CrossRef]
- Sedy, J.R.; Bjordahl, R.L.; Bekiaris, V.; Macauley, M.G.; Ware, B.C.; Norris, P.S.; Lurain, N.S.; Benedict, C.A.; Ware, C.F. CD160 activation by herpesvirus entry mediator augments inflammatory cytokine production and cytolytic function by NK cells. J. Immunol. 2013, 191, 828–836. [Google Scholar] [CrossRef]
- Park, S.C.; Jeen, Y.M.; Jeen, Y.T. Approach to cytomegalovirus infections in patients with ulcerative colitis. Korean J. Intern. Med. 2017, 32, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Ku, J.L.; Park, J.G. Biology of SNU cell lines. Cancer Res. Treat. 2005, 37, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Li, W.; Hou, Z.; Han, Q.; Lan, P.; Zhang, J.; Tian, Z.; Zhang, C. HBV suppresses expression of MICA/B on hepatoma cells through up-regulation of transcription factors GATA2 and GATA3 to escape from NK cell surveillance. Oncotarget 2016, 7, 56107–56119. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lin, Y.T.; Yan, X.L.; Ding, Y.L.; Wu, Y.L.; Chen, W.N.; Lin, X. Hepatitis B virus core protein inhibits Fas-mediated apoptosis of hepatoma cells via regulation of mFas/FasL and sFas expression. FASEB J. 2015, 29, 1113–1123. [Google Scholar] [CrossRef]
- Park, S.H. Trends in the seroprevalence of hepatitis B surface antigen in the South Korean population. Int J. Infect. Dis. 2012, 16, e669–e672. [Google Scholar] [CrossRef] [PubMed]
- Rolle, A.; Mousavi-Jazi, M.; Eriksson, M.; Odeberg, J.; Soderberg-Naucler, C.; Cosman, D.; Karre, K.; Cerboni, C. Effects of human cytomegalovirus infection on ligands for the activating NKG2D receptor of NK cells: Up-regulation of UL16-binding protein (ULBP)1 and ULBP2 is counteracted by the viral UL16 protein. J. Immunol. 2003, 171, 902–908. [Google Scholar] [CrossRef]
- Fielding, C.A.; Aicheler, R.; Stanton, R.J.; Wang, E.C.; Han, S.; Seirafian, S.; Davies, J.; McSharry, B.P.; Weekes, M.P.; Antrobus, P.R.; et al. Two novel human cytomegalovirus NK cell evasion functions target MICA for lysosomal degradation. PLoS Pathog. 2014, 10, e1004058. [Google Scholar] [CrossRef] [PubMed]
- Nwangwu, C.A.; Weiher, H.; Schmidt-Wolf, I.G.H. Increase of CIK cell efficacy by upregulating cell surface MICA and inhibition of NKG2D ligand shedding in multiple myeloma. Hematol. Oncol. 2017, 35, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.J.; Zhao, X.Y.; Huang, X.J. Strategies for Enhancing and Preserving Anti-leukemia Effects Without Aggravating Graft-Versus-Host Disease. Front. Immunol. 2018, 9, 3041. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | |||
| % | CD3−CD56brightCD16− | CD3−CD56dimCD16+ | Fold (CD56dim/CD56bright) |
| CD25 | 1.87 ± 1.54 | 2.27 ± 3.12 | 1.25 |
| CD122 (IL2Rβ) | 85.99 ± 10.94 | 83.27 ± 18.86 | 0.96 |
| CD212 (IL12Rβ) *** | 19.85 ± 19.45 | 46.92 ± 31.47 | 2.36 |
| TLR2 | 4.56 ± 5.91 | 4.19 ± 6.21 | 0.91 |
| TLR4 | 37 ± 38.94 | 30.8 ± 37.53 | 0.83 |
| NKG2D ** | 97.86 ± 3.04 | 80.1 ± 13.54 | 0.81 |
| NKp30 | 50.35 ± 14.99 | 50.79 ± 19.12 | 1.00 |
| NKp44 ** | 5.86 ± 4.37 | 1.56 ± 2.13 | 0.26 |
| NKp46 ** | 77.1 ± 12.47 | 50.74 ± 27.11 | 0.65 |
| CD253 (TRAIL) *** | 12.41 ± 11.56 | 6.83 ± 8.93 | 0.55 |
| CD178 (FASL) * | 18.73 ± 18.63 | 14.06 ± 20.32 | 0.75 |
| CD154 (CD40L) | 1.81 ± 1.78 | 4.03 ± 4.42 | 2.22 |
| B | |||
| MFI | CD3−CD56brightCD16− | CD3−CD56dimCD16+ | Fold (CD56dim/CD56bright) |
| CD25 | 119.68 ± 107.31 | 174.24 ± 164.65 | 1.45 |
| CD122 (IL2Rβ) | 3326.73 ± 1455.05 | 1917.64 ± 589.08 | 0.57 |
| CD212 (IL12Rβ) *** | 350.58 ± 177.04 | 661.58 ± 239.14 | 1.88 |
| TLR2 | 239.10 ± 113.86 | 271.60 ± 71.91 | 1.13 |
| TLR4 | 723.45 ± 431.32 | 488.73 ± 215.92 | 0.67 |
| NKG2D *** | 4712.7 ± 874.1 | 2311.57 ± 603.8 | 0.49 |
| NKp30 | 986.27 ± 510.16 | 912.82 ± 414.95 | 0.92 |
| NKp44 ** | 159.75 ± 114.52 | 77.74 ± 46.5 | 0.48 |
| NKp46 ** | 4647.67 ± 1747.12 | 1357.25 ± 466.69 | 0.29 |
| CD253 (TRAIL) *** | 348.08 ± 180.2 | 207.17 ± 79.25 | 0.59 |
| CD178 (FASL) * | 368.82 ± 188.33 | 301.18 ± 152.1 | 0.81 |
| CD154 (CD40L) | 151.5 ± 22.84 | 240.5 ± 78.56 | 1.58 |
| A | |||
| % | CD56brightCD16− | CD56dimCD16+ | Fold (CD56dim/CD56bright) |
| CD279 (PD-1) | 9.59 ± 4.33 | 15 ± 5.16 | 1.56 |
| CD94 ** | 94.8 ± 3.37 | 68.45 ± 4.03 | 0.72 |
| CD85j * | 35.75 ± 3.71 | 50.1 ± 3.29 | 1.4 |
| CD272 (BTLA) * | 25 ± 3.29 | 29.75 ± 3.95 | 1.19 |
| CD152 (CTLA-4) | 10.03 ± 2.53 | 8.23 ± 1.24 | 0.82 |
| B | |||
| MFI | CD56brightCD16− | CD56dimCD16+ | Fold (CD56dim/CD56bright) |
| CD279 (PD-1) ** | 4149 ± 277.88 | 1998 ± 150.93 | 0.48 |
| CD94 | 280 ± 80.18 | 309.5 ± 67.73 | 1.1 |
| CD85j | 1444.5 ± 237.49 | 1682 ± 212.04 | 1.16 |
| CD272 (BTLA) * | 477.5 ± 93.62 | 639.5 ± 116.7 | 1.33 |
| CD152 (CTLA-4) | 257 ± 86.19 | 208.5 ± 42.66 | 0.811 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.; Han, J.; Baek, J.-S.; Tak, E.; Song, G.-W.; Lee, S.-G.; Jung, D.-H.; Park, G.-C.; Ahn, C.-S.; Kim, N. Cytotoxicity of Human Hepatic Intrasinusoidal CD56bright Natural Killer Cells against Hepatocellular Carcinoma Cells. Int. J. Mol. Sci. 2019, 20, 1564. https://doi.org/10.3390/ijms20071564
Hwang S, Han J, Baek J-S, Tak E, Song G-W, Lee S-G, Jung D-H, Park G-C, Ahn C-S, Kim N. Cytotoxicity of Human Hepatic Intrasinusoidal CD56bright Natural Killer Cells against Hepatocellular Carcinoma Cells. International Journal of Molecular Sciences. 2019; 20(7):1564. https://doi.org/10.3390/ijms20071564
Chicago/Turabian StyleHwang, Shin, Jaeseok Han, Ji-Seok Baek, Eunyoung Tak, Gi-Won Song, Sung-Gyu Lee, Dong-Hwan Jung, Gil-Chun Park, Chul-Soo Ahn, and Nayoung Kim. 2019. "Cytotoxicity of Human Hepatic Intrasinusoidal CD56bright Natural Killer Cells against Hepatocellular Carcinoma Cells" International Journal of Molecular Sciences 20, no. 7: 1564. https://doi.org/10.3390/ijms20071564
APA StyleHwang, S., Han, J., Baek, J.-S., Tak, E., Song, G.-W., Lee, S.-G., Jung, D.-H., Park, G.-C., Ahn, C.-S., & Kim, N. (2019). Cytotoxicity of Human Hepatic Intrasinusoidal CD56bright Natural Killer Cells against Hepatocellular Carcinoma Cells. International Journal of Molecular Sciences, 20(7), 1564. https://doi.org/10.3390/ijms20071564