Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol

 ,
,  ,
,  ,
, 

Abstract
1. Health Beneficial Effects of Resveratrol (RSV)
2. Pharmacokinetic Characteristics of RSV
3. Methodological Approaches to Improve RSV Oral Bioavailability
3.1. Lipid Nanocarriers and Liposomes
3.2. Nanoemulsions
3.3. Micelles
3.4. Polymeric Nanoparticles
3.5. Solid Dispersions
3.6. Nanocrystals
3.7. Limitations and Similarities of Different Methodological Approaches
4. RSV Derivatives
4.1. Methoxylated RSV Derivatives
4.2. Hydroxylated RSV Derivatives
4.3. Halogenated Derivatives
4.4. Other RSV Derivatives
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Guerrero, R.F.; Garcia-Parrilla, M.C.; Puertas, B.; Cantos-Villar, E. Wine, resveratrol and health: A review. Nat. Prod. Commun. 2009, 4, 635–658. [Google Scholar] [PubMed]
- Tokusoglu, O.; Unal, M.K.; Yemis, F. Determination of the phytoalexin resveratrol (3,5,4′-trihydroxystilbene) in peanuts and pistachios by high-performance liquid chromatographic diode array (hplc-dad) and gas chromatography-mass spectrometry (gc-ms). J. Agric. Food Chem. 2005, 53, 5003–5009. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.; Delorgeril, M. Wine, alcohol, platelets, and the french paradox for coronary heart-disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Valentovic, M.A. Evaluation of resveratrol in cancer patients and experimental models. Adv. Cancer Res. 2018, 137, 171–188. [Google Scholar]
- Koushki, M.; Amiri-Dashatan, N.; Ahmadi, N.; Abbaszadeh, H.A.; Rezaei-Tavirani, M. Resveratrol: A miraculous natural compound for diseases treatment. Food Sci. Nutr. 2018, 6, 2473–2490. [Google Scholar] [CrossRef]
- Rahman, I.; Biswas, S.K.; Kirkham, P.A. Regulation of inflammation and redox signaling by dietary polyphenols. Biochem. Pharmacol. 2006, 72, 1439–1452. [Google Scholar] [CrossRef]
- Frankel, E.N.; Waterhouse, A.L.; Kinsella, J.E. Inhibition of human ldl oxidation by resveratrol. Lancet 1993, 341, 1103–1104. [Google Scholar] [CrossRef]
- Belguendouz, L.; Fremont, L.; Linard, A. Resveratrol inhibits metal ion-dependent and independent peroxidation of porcine low-density lipoproteins. Biochem. Pharmacol. 1997, 53, 1347–1355. [Google Scholar] [CrossRef]
- Han, J.; Suga, K.; Hayashi, K.; Okamoto, Y.; Umakoshi, H. Multi-level characterization of the membrane properties of resveratrol-incorporated liposomes. J Phys Chem B. 2017, 121, 4091–4098. [Google Scholar] [CrossRef]
- Voloshyna, I.; Hussaini, S.M.; Reiss, A.B. Resveratrol in cholesterol metabolism and atherosclerosis. J. Med. Food 2012, 15, 763–773. [Google Scholar] [CrossRef]
- Kutil, Z.; Temml, V.; Maghradze, D.; Pribylova, M.; Dvorakova, M.; Schuster, D.; Vanek, T.; Landa, P. Impact of wines and wine constituents on cyclooxygenase-1, cyclooxygenase-2, and 5-lipoxygenase catalytic activity. Mediat. Inflamm. 2014, 2014, 178931. [Google Scholar] [CrossRef]
- Wicinski, M.; Socha, M.; Walczak, M.; Wodkiewicz, E.; Malinowski, B.; Rewerski, S.; Gorski, K.; Pawlak-Osinska, K. Beneficial effects of resveratrol administration-focus on potential biochemical mechanisms in cardiovascular conditions. Nutrients 2018, 10, 1813. [Google Scholar] [CrossRef]
- Toliopoulos, I.K.; Simos, Y.V.; Oikonomidis, S.; Karkabounas, S.C. Resveratrol diminishes platelet aggregation and increases susceptibility of k562 tumor cells to natural killer cells. Indian J. Biochem. Biophys. 2013, 50, 14–18. [Google Scholar]
- Zordoky, B.N.M.; Robertson, I.M.; Dyck, J.R.B. Preclinical and clinical evidence for the role of resveratrol in the treatment of cardiovascular diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 1155–1177. [Google Scholar] [CrossRef]
- Gordish, K.L.; Beierwaltes, W.H. Resveratrol induces acute endothelium-dependent renal vasodilation mediated through nitric oxide and reactive oxygen species scavenging. Am. J. Physiol. Ren. Physiol. 2014, 306, F542–F550. [Google Scholar] [CrossRef]
- Cao, W.Y.; Dou, Y.; Li, A.P. Resveratrol boosts cognitive function by targeting sirt1. Neurochem. Res. 2018, 43, 1705–1713. [Google Scholar] [CrossRef]
- Wang, Y.J.; He, J.; Liao, M.Y.; Hu, M.X.; Li, W.Z.; Ouyang, H.L.; Wang, X.; Ye, T.H.; Zhang, Y.W.; Ouyang, L. An overview of sirtuins as potential therapeutic target: Structure, function and modulators. Eur. J. Med. Chem. 2019, 161, 48–77. [Google Scholar] [CrossRef]
- Capiralla, H.; Vingtdeux, V.; Zhao, H.T.; Sankowski, R.; Al-Abed, Y.; Davies, P.; Marambaud, P. Resveratrol mitigates lipopolysaccharide- and a ss-mediated microglial inflammation by inhibiting the tlr4/nf-?B/stat signaling cascade. J. Neurochem. 2012, 120, 461–472. [Google Scholar] [CrossRef]
- Pallas, M.; Casadesus, G.; Smith, M.A.; Coto-Montes, A.; Pelegri, C.; Vilaplana, J.; Camins, A. Resveratrol and neurodegenerative diseases: Activation of sirt1 as the potential pathway towards neuroprotection. Curr. Neurovasc. Res. 2009, 6, 70–81. [Google Scholar] [CrossRef]
- Lin, H.Y.; Sun, M.; Tang, H.Y.; Simone, T.M.; Wu, Y.H.; Grandis, J.R.; Cao, H.J.; Davis, P.J.; Davis, F.B. Resveratrol causes cox-2-and p53-dependent apoptosis in head and neck squamous cell cancer cells. J. Cell Biochem. 2008, 104, 2131–2142. [Google Scholar] [CrossRef]
- Murias, M.; Handler, N.; Erker, T.; Pleban, K.; Ecker, G.; Saiko, P.; Szekeres, T.; Jager, W. Resveratrol analogues as selective cyclooxygenase-2 inhibitors: Synthesis and structure-activity relationship. Bioorg. Med. Chem. 2004, 12, 5571–5578. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H.; Jang, H.J.; Chae, H.S.; Oh, Y.C.; Choi, J.G.; Lee, Y.S.; Kim, J.H.; Kim, Y.C.; Sohn, D.H.; Park, H.; et al. Anti-inflammatory mechanisms of resveratrol in activated hmc-1 cells: Pivotal roles of nf-kappa b and mapk. Pharmacol. Res. 2009, 59, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Slater, S.J.; Seiz, J.L.; Cook, A.C.; Stagliano, B.A.; Buzas, C.J. Inhibition of protein kinase c by resveratrol. Biochim. Et Biophys. Acta-Mol. Basis Dis. 2003, 1637, 59–69. [Google Scholar] [CrossRef]
- Beedanagari, S.R.; Bebenek, I.; Bui, P.; Hankinson, O. Resveratrol inhibits dioxin-induced expression of human cyp1a1 and cyp1b1 by inhibiting recruitment of the aryl hydrocarbon receptor complex and rna polymerase ii to the regulatory regions of the corresponding genes. Toxicol. Sci. 2009, 110, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Fujii-Kuriyama, Y. Metabolic activation of polycyclic aromatic hydrocarbons to carcinogens by cytochromes p450 1a1 and 1b1. Cancer Sci. 2004, 95, 1–6. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an anti-cancer agent: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1428–1447. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, S.Q.; Ying, Y.; Zhou, R.Q.; Mao, P. Mir-196b/mir-1290 participate in the antitumor effect of resveratrol via regulation of igfbp3 expression in acute lymphoblastic leukemia. Oncol. Rep. 2017, 37, 1075–1083. [Google Scholar] [PubMed]
- Li, D.; Wang, G.C.; Jin, G.G.; Yao, K.; Zhao, Z.J.; Bie, L.Y.; Guo, Y.J.; Li, N.; Deng, W.Y.; Chen, X.B.; et al. Resveratrol suppresses colon cancer growth by targeting the akt/stat3 signaling pathway. Int. J. Mol. Med. 2019, 43, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Yan, B.; Chen, K.; Jiang, Z.D.; Zhou, C.C.; Cao, J.Y.; Qian, W.K.; Li, J.; Sun, L.K.; Ma, J.G.; et al. Resveratrol-induced downregulation of naf-1 enhances the sensitivity of pancreatic cancer cells to gemcitabine via the ros/nrf2 signaling pathways. Oxid. Med. Cell. Longev. 2018, 2018, 9482018. [Google Scholar]
- Heo, J.R.; Kim, S.M.; Hwang, K.A.; Kang, J.H.; Choi, K.C. Resveratrol induced reactive oxygen species and endoplasmic reticulum stress-mediated apoptosis, and cell cycle arrest in the a375sm malignant melanoma cell line. Int. J. Mol. Med. 2018, 42, 1427–1435. [Google Scholar] [CrossRef]
- Wu, X.X.; Xu, Y.D.; Zhu, B.R.; Liu, Q.; Yao, Q.F.; Zhao, G. Resveratrol induces apoptosis in sgc-7901 gastric cancer cells. Oncol. Lett. 2018, 16, 2949–2956. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qiu, R.L.; Lin, Y.; Cai, Y.; Bian, Y.; Fan, Y.; Gao, X.J. Resveratrol suppresses human cervical carcinoma cell proliferation and elevates apoptosis via the mitochondrial and p53 signaling pathways. Oncol. Lett. 2018, 15, 9845–9851. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tong, L.; Luo, Y.; Li, X.; Chen, G.W.; Wang, Y.F. Resveratrol inhibits the proliferation and induces the apoptosis in ovarian cancer cells via inhibiting glycolysis and targeting ampk/mtor signaling pathway. J. Cell Biochem. 2018, 119, 6162–6172. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Oda, K.; Wada-Hiraike, O.; Sone, K.; Inaba, K.; Ikeda, Y.; Makii, C.; Miyasaka, A.; Kashiyama, T.; Tanikawa, M.; et al. Autophagy inhibition augments resveratrol-induced apoptosis in ishikawa endometrial cancer cells. Oncol. Lett. 2016, 12, 2560–2566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.C.; Yin, X.N.; Sui, S.G. Resveratrol inhibited the progression of human hepatocellular carcinoma by inducing autophagy via regulating p53 and the phosphoinositide 3-kinase/protein kinase b pathway. Oncol. Rep. 2018, 40, 2758–2765. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Martinez, D.; Soto, A.; Gil-Araujo, B.; Gallego, B.; Chiloeches, A.; Lasa, M. Resveratrol promotes apoptosis through the induction of dual specificity phosphatase 1 and sensitizes prostate cancer cells to cisplatin. Food Chem. Toxicol. 2018, 124, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Ferraz da Costa, D.C.; Campos, N.P.C.; Santos, R.A.; Guedes-da-Silva, F.H.; Martins-Dinis, M.; Zanphorlin, L.; Ramos, C.; Rangel, L.P.; Silva, J.L. Resveratrol prevents p53 aggregation in vitro and in breast cancer cells. Oncotarget 2018, 9, 29112–29122. [Google Scholar] [PubMed]
- Ko, J.H.; Sethi, G.; Um, J.Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The role of resveratrol in cancer therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef] [PubMed]
- Dybkowska, E.; Sadowska, A.; Swiderski, F.; Rakowska, R.; Wysocka, K. The occurrence of resveratrol in foodstuffs and its potential for supporting cancer prevention and treatment. A review. Rocz Panstw Zakl Hig 2018, 69, 5–14. [Google Scholar] [PubMed]
- Singh, C.K.; Ndiaye, M.A.; Ahmad, N. Resveratrol and cancer: Challenges for clinical translation. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 1178–1185. [Google Scholar] [CrossRef]
- Cai, H.; Scott, E.; Kholghi, A.; Andreadi, C.; Rufini, A.; Karmokar, A.; Britton, R.G.; Horner-Glister, E.; Greaves, P.; Jawad, D.; et al. Cancer chemoprevention: Evidence of a nonlinear dose response for the protective effects of resveratrol in humans and mice. Sci. Transl. Med. 2015, 7, 298ra117. [Google Scholar] [CrossRef] [PubMed]
- Howells, L.M.; Berry, D.P.; Elliott, P.J.; Jacobson, E.W.; Hoffmann, E.; Hegarty, B.; Brown, K.; Steward, W.P.; Gescher, A.J. Phase i randomized, double-blind pilot study of micronized resveratrol (srt501) in patients with hepatic metastases-safety, pharmacokinetics, and pharmacodynamics. Cancer Prev. Res. 2011, 4, 1419–1425. [Google Scholar] [CrossRef]
- Wenzel, E.; Somoza, V. Metabolism and bioavailability of trans-resveratrol. Mol. Nutr. Food Res. 2005, 49, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Gambini, J.; Ingles, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomez-Cabrera, M.C.; Vina, J.; et al. Properties of resveratrol: In vitro and in vivo studies about metabolism, bioavailability, and biological effects in animal models and humans. Oxid. Med. Cell. Longev. 2015, 2015, 837042. [Google Scholar] [CrossRef] [PubMed]
- Delmas, D.; Aires, V.; Limagne, E.; Dutartre, P.; Mazue, F.; Ghiringhelli, F.; Latruffe, N. Transport, stability, and biological activity of resveratrol. Resveratrol Health 2011, 1215, 48–59. [Google Scholar] [CrossRef]
- Boocock, D.J.; Patel, K.R.; Faust, G.E.S.; Normolle, D.P.; Marczylo, T.H.; Crowell, J.A.; Brenner, D.E.; Booth, T.D.; Gescher, A.; Steward, W.P. Quantitation of trans-resveratrol and detection of its metabolites in human plasma and urine by high performance liquid chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 848, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Ruotolo, R.; Calani, L.; Fietta, E.; Brighenti, F.; Crozier, A.; Meda, C.; Maggi, A.; Ottonello, S.; Del Rio, D. Anti-estrogenic activity of a human resveratrol metabolite. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, E.; Soldo, T.; Erbersdobler, H.; Somoza, V. Bioactivity and metabolism of trans-resveratrol orally administered to wistar rats. Mol. Nutr. Food Res. 2005, 49, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Marier, J.F.; Vachon, P.; Gritsas, A.; Zhang, J.; Moreau, J.P.; Ducharme, M.P. Metabolism and disposition of resveratrol in rats: Extent of absorption, glucuronidation, and enterohepatic recirculation evidenced by a linked-rat model. J. Pharmacol. Exp. Ther. 2002, 302, 369–373. [Google Scholar] [CrossRef]
- Patel, K.R.; Scott, E.; Brown, V.A.; Gescher, A.J.; Steward, W.P.; Brown, K. Clinical trials of resveratrol. Resveratrol. Health 2011, 1215, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.; Vaz-da-Silva, M.; Falcao, A.; Soares, E.; Costa, R.; Loureiro, A.I.; Fernandes-Lopes, C.; Rocha, J.F.; Nunes, T.; Wright, L.; et al. Pharmacokinetic and safety profile of trans-resveratrol in a rising multiple-dose study in healthy volunteers. Mol. Nutr. Food Res. 2009, 53, S7–S15. [Google Scholar] [CrossRef] [PubMed]
- Boocock, D.J.; Faust, G.E.S.; Patel, K.R.; Schinas, A.M.; Brown, V.A.; Ducharme, M.P.; Booth, T.D.; Crowell, J.A.; Perloff, M.; Gescher, A.J.; et al. Phase i dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: A review of clinical trials. Npj Precis. Oncol. 2017, 1, 35. [Google Scholar] [CrossRef] [PubMed]
- Amidon, G.L.; Lennernas, H.; Shah, V.P.; Crison, J.R. A theoretical basis for a biopharmaceutic drug classification—The correlation of in-vitro drug product dissolution and in-vivo bioavailability. Pharm. Res. 1995, 12, 413–420. [Google Scholar] [CrossRef]
- Lobenberg, R.; Amidon, G.L. Modern bioavailability, bioequivalence and biopharmaceutics classification system. New scientific approaches to international regulatory standards. Eur. J. Pharm. Biopharm. 2000, 50, 3–12. [Google Scholar] [CrossRef]
- Amri, A.; Chaumeil, J.C.; Sfar, S.; Charrueau, C. Administration of resveratrol: What formulation solutions to bioavailability limitations? J. Control. Release 2012, 158, 182–193. [Google Scholar] [CrossRef]
- Chang, C.W.; Wong, C.Y.; Wu, Y.T.; Hsu, M.C. Development of a solid dispersion system for improving the oral bioavailability of resveratrol in rats. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 239–249. [Google Scholar] [CrossRef]
- Spogli, R.; Bastianini, M.; Ragonese, F.; Iannitti, R.G.; Monarca, L.; Bastioli, F.; Nakashidze, I.; Brecchia, G.; Menchetti, L.; Codini, M.; et al. Solid dispersion of resveratrol supported on magnesium dihydroxide (resv@mdh) microparticles improves oral bioavailability. Nutrients 2018, 10, 1925. [Google Scholar] [CrossRef]
- Leuner, C.; Dressman, J. Improving drug solubility for oral delivery using solid dispersions. Eur. J. Pharm. Biopharm. 2000, 50, 47–60. [Google Scholar] [CrossRef]
- Singh, S.K.; Makadia, V.; Sharma, S.; Rashid, M.; Shahi, S.; Mishra, P.R.; Wahajuddin, M.; Gayen, J.R. Preparation and in-vitro/in-vivo characterization of trans-resveratrol nanocrystals for oral administration. Drug Deliv. Transl. Res. 2017, 7, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Lucio, M.; Martins, S.; Lima, J.L.C.; Reis, S. Novel resveratrol nanodelivery systems based on lipid nanoparticles to enhance its oral bioavailability. Int. J. Nanomed. 2013, 8, 177–187. [Google Scholar]
- Pandita, D.; Kumar, S.; Poonia, N.; Lather, V. Solid lipid nanoparticles enhance oral bioavailability of resveratrol, a natural polyphenol. Food Res. Int. 2014, 62, 1165–1174. [Google Scholar] [CrossRef]
- Ting, Y.W.; Jiang, Y.; Ho, C.T.; Huang, Q.R. Common delivery systems for enhancing in vivo bioavailability and biological efficacy of nutraceuticals. J. Funct. Foods 2014, 7, 112–128. [Google Scholar] [CrossRef]
- Basavaraj, S.; Betageri, G.V. Improved oral delivery of resveratrol using proliposomal formulation: Investigation of various factors contributing to prolonged absorption of unmetabolized resveratrol. Expert Opin. Drug Deliv. 2014, 11, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Zu, Y.J.; Overby, H.; Ren, G.F.; Fan, Z.Y.; Zhao, L.; Wang, S. Resveratrol liposomes and lipid nanocarriers: Comparison of characteristics and inducing browning of white adipocytes. Colloids Surf. B Biointerfaces 2018, 164, 414–423. [Google Scholar] [CrossRef]
- Sessa, M.; Tsao, R.; Liu, R.H.; Ferrari, G.; Donsi, F. Evaluation of the stability and antioxidant activity of nanoencapsulated resveratrol during in vitro digestion. J. Agric. Food Chem. 2011, 59, 12352–12360. [Google Scholar] [CrossRef]
- Sessa, M.; Balestrieri, M.L.; Ferrari, G.; Servillo, L.; Castaldo, D.; D’Onofrio, N.; Donsi, F.; Tsao, R. Bioavailability of encapsulated resveratrol into nanoemulsion-based delivery systems. Food Chem. 2014, 147, 42–50. [Google Scholar] [CrossRef]
- Yen, C.C.; Chang, C.W.; Hsu, M.C.; Wu, Y.T. Self-nanoemulsifying drug delivery system for resveratrol: Enhanced oral bioavailability and reduced physical fatigue in rats. Int. J. Mol. Sci. 2017, 18, 1853. [Google Scholar] [CrossRef]
- Yang, F.F.; Zhou, J.; Hu, X.; Cong, Z.Q.; Liu, C.Y.; Pan, R.L.; Chang, Q.; Liu, X.M.; Liao, Y.H. Improving oral bioavailability of resveratrol by a udp-glucuronosyltransferase inhibitory excipient-based self-microemulsion. Eur. J. Pharm. Sci. 2018, 114, 303–309. [Google Scholar] [CrossRef]
- Wiedmann, T.S.; Liang, W.; Kamel, L. Solubilization of drugs by physiological mixtures of bile salts. Pharm. Res. 2002, 19, 1203–1208. [Google Scholar] [CrossRef]
- Atanackovic, M.; Posa, M.; Heinle, H.; Gojkovic-Bukarica, L.; Cvejic, J. Solubilization of resveratrol in micellar solutions of different bile acids. Colloids Surf. B Biointerfaces 2009, 72, 148–154. [Google Scholar] [CrossRef]
- Calvo-Castro, L.A.; Schiborr, C.; David, F.; Ehrt, H.; Voggel, J.; Sus, N.; Behnam, D.; Bosy-Westphal, A.; Frank, J. The oral bioavailability of trans-resveratrol from a grapevine-shoot extract in healthy humans is significantly increased by micellar solubilization. Mol. Nutr. Food Res. 2018, 62, 1701057. [Google Scholar] [CrossRef]
- Penalva, R.; Morales, J.; Gonzalez-Navarro, C.J.; Larraneta, E.; Quincoces, G.; Penuelas, I.; Irache, J.M. Increased oral bioavailability of resveratrol by its encapsulation in casein nanoparticles. Int. J. Mol. Sci. 2018, 19, 2816. [Google Scholar] [CrossRef]
- Bachar, M.; Mandelbaum, A.; Portnaya, I.; Perlstein, H.; Even-Chen, S.; Barenholz, Y.; Danino, D. Development and characterization of a novel drug nanocarrier for oral delivery, based on self-assembled beta-casein micelles. J. Control. Release 2012, 160, 164–171. [Google Scholar] [CrossRef]
- Zhang, J.; Mi, Q.; Shen, M. Resveratrol binding to collagen and its biological implication. Food Chem. 2012, 131, 879–884. [Google Scholar] [CrossRef]
- Shao, J.F.; Li, X.L.; Lu, X.W.; Jiang, C.; Hu, Y.; Li, Q.P.; You, Y.P.; Fu, Z. Enhanced growth inhibition effect of resveratrol incorporated into biodegradable nanoparticles against glioma cells is mediated by the induction of intracellular reactive oxygen species levels. Colloids Surf. B Biointerfaces 2009, 72, 40–47. [Google Scholar] [CrossRef]
- Adams, M.L.; Lavasanifar, A.; Kwon, G.S. Amphiphilic block copolymers for drug delivery. J. Pharm. Sci. 2003, 92, 1343–1355. [Google Scholar] [CrossRef]
- Wan, S.Q.; Zhang, L.; Quan, Y.Y.; Wei, K. Resveratrol-loaded plga nanoparticles: Enhanced stability, solubility and bioactivity of resveratrol for non-alcoholic fatty liver disease therapy. R. Soc. Open Sci. 2018, 5, 181457. [Google Scholar] [CrossRef]
- Nassir, A.M.; Shahzad, N.; Ibrahim, I.A.A.; Ahmad, I.; Md, S.; Ain, M.R. Resveratrol-loaded plga nanoparticles mediated programmed cell death in prostate cancer cells. Saudi Pharm. J. 2018, 26, 876–885. [Google Scholar] [CrossRef]
- Siu, F.Y.K.; Ye, S.T.; Lin, H.; Li, S.J. Galactosylated plga nanoparticles for the oral delivery of resveratrol: Enhanced bioavailability and in vitro anti-inflammatory activity. Int. J. Nanomed. 2018, 13, 4133–4144. [Google Scholar] [CrossRef] [PubMed]
- Maier-Salamon, A.; Hagenauer, B.; Wirth, M.; Gabor, F.; Szekeres, T.; Jager, W. Increased transport of resveratrol across monolayers of the human intestinal Caco-2 cells is mediated by inhibition and saturation of metabolites. Pharm. Res. 2006, 23, 2107–2115. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Pai, R.S. Optimized plga nanoparticle platform for orally dosed trans-resveratrol with enhanced bioavailability potential. Expert Opin. Drug Deliv. 2014, 11, 647–659. [Google Scholar] [CrossRef]
- Barenholz, Y. Liposome application: Problems and prospects. Curr. Opin. Colloid Interface Sci. 2001, 6, 66–77. [Google Scholar] [CrossRef]
- Watkins, R.; Wu, L.; Zhang, C.M.; Davis, R.M.; Xu, B. Natural product-based nanomedicine: Recent advances and issues. Int. J. Nanomed. 2015, 10, 6055–6074. [Google Scholar]
- Vasconcelos, T.; Sarmento, B.; Costa, P. Solid dispersions as strategy to improve oral bioavailability of poor water soluble drugs. Drug. Discov. Today 2007, 12, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yao, G.L.; Liu, X.T.; Yin, H.P. Preparation nanocrystals of poorly soluble plant compounds using an ultra-small-scale approach. Aaps Pharmscitech 2017, 18, 2610–2617. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Aldawsari, F.S.; Aguiar, R.P.; Wiirzler, L.A.M.; Aguayo-Ortiz, R.; Aljuhani, N.; Cuman, R.K.N.; Medina-Franco, J.L.; Siraki, A.G.; Velazquez-Martinez, C.A. Anti-inflammatory and antioxidant properties of a novel resveratrol-salicylate hybrid analog. Bioorg. Med. Chem. Lett. 2016, 26, 1411–1415. [Google Scholar] [CrossRef]
- Antus, C.; Radnai, B.; Dombovari, P.; Fonai, F.; Avar, P.; Matyus, P.; Racz, B.; Sumegi, B.; Veres, B. Anti-inflammatory effects of a triple-bond resveratrol analog: Structure and function relationship. Eur. J. Pharm. 2015, 748, 61–67. [Google Scholar] [CrossRef]
- Chimento, A.; Sirianni, R.; Saturnino, C.; Caruso, A.; Sinicropi, M.S.; Pezzi, V. Resveratrol and its analogs as antitumoral agents for breast cancer treatment. Mini-Rev. Med. Chem. 2016, 16, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Sale, S.; Verschoyle, R.D.; Boocock, D.; Jones, D.J.L.; Wilsher, N.; Ruparelia, K.C.; Potter, G.A.; Farmer, P.B.; Steward, W.P.; Gescher, A.J. Pharmacokinetics in mice and growth-inhibitory properties of the putative cancer chemopreventive agent resveratrol and the synthetic analogue trans 3,4,5,4′-tetramethoxystilbene. Br. J. Cancer 2004, 90, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Zhang, W.; Go, M.L.; Choo, Q.Y.; Ho, P.C. Determination of z-3,5,4′-trimethoxystilbene in rat plasma by a simple hplc method: Application in a pre-clinical pharmacokinetic study. J. Pharm. Biomed. Anal. 2010, 53, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yeo, S.C.M.; Elhennawy, M.G.A.A.; Xiang, X.Q.; Lin, H.S. Determination of naturally occurring resveratrol analog trans-4,4′-dihydroxystilbene in rat plasma by liquid chromatography-tandem mass spectrometry: Application to a pharmacokinetic study. Anal. Bioanal. Chem. 2015, 407, 5793–5801. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yeo, S.C.M.; Elhennawy, M.G.A.A.; Lin, H.S. Oxyresveratrol: A bioavailable dietary polyphenol. J. Funct. Foods 2016, 22, 122–131. [Google Scholar] [CrossRef]
- Moran, B.W.; Anderson, F.P.; Devery, A.; Cloonan, S.; Butler, W.E.; Varughese, S.; Draper, S.M.; Kenny, P.T.M. Synthesis, structural characterisation and biological evaluation of fluorinated analogues of resveratrol. Bioorg. Med. Chem. 2009, 17, 4510–4522. [Google Scholar] [CrossRef]
- Li, X.Z.; Wei, X.; Zhang, C.J.; Jin, X.L.; Tang, J.J.; Fan, G.J.; Zhou, B. Hypohalous acid-mediated halogenation of resveratrol and its role in antioxidant and antimicrobial activities. Food Chem. 2012, 135, 1239–1244. [Google Scholar] [CrossRef]
- Estrela, J.M.; Ortega, A.; Mena, S.; Rodriguez, M.L.; Asensi, M. Pterostilbene: Biomedical applications. Crit. Rev. Clin. Lab. Sci. 2013, 50, 65–78. [Google Scholar] [CrossRef]
- Fulda, S. Resveratrol and derivatives for the prevention and treatment of cancer. Drug. Discov. Today 2010, 15, 757–765. [Google Scholar] [CrossRef]
- Lin, H.S.; Yue, B.D.; Ho, P.C. Determination of pterostilbene in rat plasma by a simple hplc-uv method and its application in pre-clinical pharmacokinetic study. Biomed. Chromatogr. 2009, 23, 1308–1315. [Google Scholar] [CrossRef]
- Kapetanovic, I.M.; Muzzio, M.; Huang, Z.H.; Thompson, T.N.; McCormick, D.L. Pharmacokinetics, oral bioavailability, and metabolic profile of resveratrol and its dimethylether analog, pterostilbene, in rats. Cancer Chemother Pharm. 2011, 68, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Perecko, T.; Jancinova, V.; Drabikova, K.; Nosal, R.; Harmatha, J. Structure-efficiency relationship in derivatives of stilbene. Comparison of resveratrol, pinosylvin and pterostilbene. Neuroendocrinol. Lett. 2008, 29, 802–805. [Google Scholar] [PubMed]
- McCormack, D.; McFadden, D. A review of pterostilbene antioxidant activity and disease modification. Oxidative Med. Cell. Longev. 2013, 2013, 575482. [Google Scholar] [CrossRef] [PubMed]
- Satheesh, M.A.; Pari, L. The antioxidant role of pterostilbene in streptozotocin-nicotinamide-induced type 2 diabetes mellitus in wistar rats. J. Pharm. Pharmacol. 2006, 58, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Pari, L.; Satheesh, M.A. Effect of pterostilbene on hepatic key enzymes of glucose metabolism in streptozotocin- and nicotinamide-induced diabetic rats. Life Sci. 2006, 79, 641–645. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Liu, K.X.; Fu, S.P.; Li, Z.; Yu, X. Pterostilbene attenuates the inflammatory reaction induced by ischemia/reperfusion in rat heart. Mol. Med. Rep. 2015, 11, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, L.; Li, F.; Hu, C.P.; Zhang, Z. Restoration of sirt1 function by pterostilbene attenuates hypoxia-reoxygenation injury in cardiomyocytes. Eur. J. Pharm. 2016, 776, 26–33. [Google Scholar] [CrossRef]
- Kosuru, R.; Cai, Y.; Kandula, V.; Yan, D.; Wang, C.Y.; Zheng, H.; Li, Y.L.; Irwin, M.G.; Singh, S.; Xia, Z.Y. Ampk contributes to cardioprotective effects of pterostilbene against myocardial ischemia-reperfusion injury in diabetic rats by suppressing cardiac oxidative stress and apoptosis. Cell. Physiol. Biochem. 2018, 46, 1381–1397. [Google Scholar] [CrossRef]
- Kosuru, R.; Kandula, V.; Rai, U.; Prakash, S.; Xia, Z.Y.; Singh, S. Pterostilbene decreases cardiac oxidative stress and inflammation via activation of ampk/nrf2/ho-1 pathway in fructose-fed diabetic rats. Cardiovasc. Drugs Ther. 2018, 32, 147–163. [Google Scholar] [CrossRef]
- Li, Y.R.; Li, S.M.; Lin, C.C. Effect of resveratrol and pterostilbene on aging and longevity. Biofactors 2018, 44, 69–82. [Google Scholar] [CrossRef]
- Li, Q.S.; Chen, L.; Liu, X.W.; Li, X.D.; Cao, Y.; Bai, Y.; Qi, F.J. Pterostilbene inhibits amyloid—Induced neuroinflammation in a microglia cell line by inactivating the nlrp3/caspase-1 inflammasome pathway. J. Cell Biochem. 2018, 119, 7053–7062. [Google Scholar] [CrossRef] [PubMed]
- McCormack, D.E.; Mannal, P.; McDonald, D.; Tighe, S.; Hanson, J.; McFadden, D. Genomic analysis of pterostilbene predicts its antiproliferative effects against pancreatic cancer in vitro and in vivo. J. Gastrointest. Surg. 2012, 16, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.Q.; Yang, Y.; Di, S.Y.; Feng, X.; Liu, D.; Jiang, S.; Hu, W.; Qin, Z.G.; Li, Y.; Lv, J.J.; et al. Pterostilbene exerts anticancer activity on non-small-cell lung cancer via activating endoplasmic reticulum stress. Sci. Rep. 2017, 7, 8091. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.Y.; Liu, Z.S.; Zhang, Z.D.; Levenson, A.S.; Li, K. Pterostilbene increases pten expression through the targeted downregulation of microrna-19a in hepatocellular carcinoma. Mol. Med. Rep. 2018, 17, 5193–5201. [Google Scholar] [CrossRef] [PubMed]
- Tolba, M.F.; Abdel-Rahman, S.Z. Pterostilbine, an active component of blueberries, sensitizes colon cancer cells to 5-fluorouracil cytotoxicity. Sci. Rep. 2015, 5, 15239. [Google Scholar] [CrossRef]
- Pan, M.H.; Chang, Y.H.; Badmaev, V.; Nagabhushanam, K.; Ho, C.T. Pterostilbene induces apoptosis and cell cycle arrest in human gastric carcinoma cells. J. Agric. Food Chem. 2007, 55, 7777–7785. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.G.; Alosi, J.A.; McDonald, D.E.; McFadden, D.W. Effects of pterostilbene on melanoma alone and in synergy with inositol hexaphosphate. Am. J. Surg. 2009, 198, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Butt, N.A.; Kumar, A.; Dhar, S.; Rimando, A.M.; Akhtar, I.; Hancock, J.C.; Lage, J.M.; Pound, C.R.; Lewin, J.R.; Gomez, C.R.; et al. Targeting mta1/hif-1 signaling by pterostilbene in combination with histone deacetylase inhibitor attenuates prostate cancer progression. Cancer Med. 2017, 6, 2673–2685. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Du, J.; Mi, Y.; Li, T.Y.; Gong, Y.; Ouyang, H.S.; Hou, Y. Long non-coding rnas contribute to the inhibition of proliferation and emt by pterostilbene in human breast cancer. Front. Oncol. 2018, 8, 629. [Google Scholar] [CrossRef]
- Liu, B.; Luo, X.J.; Yang, Z.B.; Zhang, J.J.; Li, T.B.; Zhang, X.J.; Ma, Q.L.; Zhang, G.G.; Hu, C.P.; Peng, J. Inhibition of nox/vpo1 pathway and inflammatory reaction by trimethoxystilbene in prevention of cardiovascular remodeling in hypoxia-induced pulmonary hypertensive rats. J. Cardiovasc. Pharm. 2014, 63, 567–576. [Google Scholar] [CrossRef]
- Nguyen, C.B.; Kotturi, H.; Waris, G.; Mohammed, A.; Chandrakesan, P.; May, R.; Sureban, S.; Weygant, N.; Qu, D.F.; Rao, C.V.; et al. (z)-3,5,4′-trimethoxystilbene limits hepatitis c and cancer pathophysiology by blocking microtubule dynamics and cell-cycle progression. Cancer Res. 2016, 76, 4887–4896. [Google Scholar] [CrossRef] [PubMed]
- Aldawsari, F.S.; Velazquez-Martinez, C.A. 3,4′,5-trans-trimethoxystilbene; a natural analogue of resveratrol with enhanced anticancer potency. Investig. New Drugs 2015, 33, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Traversi, G.; Fiore, M.; Leone, S.; Basso, E.; Di Muzio, E.; Polticelli, F.; Degrassi, F.; Cozzi, R. Resveratrol and its methoxy-derivatives as modulators of DNA damage induced by ionising radiation. Mutagenesis 2016, 31, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Traversi, G.; Fiore, M.; Percario, Z.; Degrassi, F.; Cozzi, R. The resveratrol analogue trimethoxystilbene inhibits cancer cell growth by inducing multipolar cell mitosis. Mol. Carcinog. 2017, 56, 1117–1126. [Google Scholar] [CrossRef]
- Piotrowska, H.; Myszkowski, K.; Amarowicz, R.; Murias, M.; Kulcenty, K.; Wierzchowski, M.; Jodynis-Liebert, J. Different susceptibility of colon cancer dld-1 and lovo cell lines to apoptosis induced by dmu-212, a synthetic resveratrol analogue. Toxicol. In Vitro 2013, 27, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Horvath, Z.; Marihart-Fazekas, S.; Saiko, P.; Grusch, M.; Ozsuy, M.; Harik, M.; Handler, N.; Erker, T.; Jaeger, W.; Fritzer-Szekeres, M.; et al. Novel resveratrol derivatives induce apoptosis and cause cell cycle arrest in prostate cancer cell lines. Anticancer Res. 2007, 27, 3459–3464. [Google Scholar] [PubMed]
- Piotrowska-Kempisty, H.; Klupczynska, A.; Trzybulska, D.; Kulcenty, K.; Sulej-Suchomska, A.M.; Kucinska, M.; Mikstacka, R.; Wierzchowski, M.; Murias, M.; Baer-Dubowska, W.; et al. Role of cyp1a1 in the biological activity of methylated resveratrol analogue, 3,4,5,4′-tetramethoxystilbene (dmu-212) in ovarian cancer a-2780 and non-cancerous hose cells. Toxicol. Lett. 2017, 267, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.X.; Yao, X.J.; Xu, S.W.; Wong, V.K.W.; He, J.X.; Ding, J.; Xue, W.W.; Mujtaba, T.; Michelangeli, F.; Huang, M.; et al. (z)3,4,5,4′-trans-tetramethoxystilbene, a new analogue of resveratrol, inhibits gefitinb-resistant non-small cell lung cancer via selectively elevating intracellular calcium level. Sci. Rep. 2015, 5, 16348. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.S.; Molavi, O.; Haddadi, A.; Lai, R.; Gossage, R.A.; Lavasanifar, A. Resveratrol analog trans 3,4,5,4′-tetramethoxystilbene (dmu-212) mediates anti-tumor effects via mechanism different from that of resveratrol. Cancer Chemother Pharm. 2008, 63, 27–35. [Google Scholar] [CrossRef]
- Piotrowska, H.; Kujawska, M.; Nowicki, M.; Petzke, E.; Ignatowicz, E.; Krajka-Kuzniak, V.; Zawierucha, P.; Wierzchowski, M.; Murias, M.; Jodynis-Liebert, J. Effect of resveratrol analogue, dmu-212, on antioxidant status and apoptosis-related genes in rat model of hepatocarcinogenesis. Hum. Exp. Toxicol. 2017, 36, 160–175. [Google Scholar] [CrossRef]
- Chun, Y.J.; Oh, Y.K.; Kim, B.J.; Kim, D.; Kim, S.S.; Choi, H.K.; Kim, M.Y. Potent inhibition of human cytochrome p450 1b1 by tetramethoxystilbene. Toxicol. Lett. 2009, 189, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P.; Chun, Y.J.; Kim, D.; Gillam, E.M.J.; Shimada, T. Cytochrome p4501b1: A target for inhibition in anticarcinogenesis strategies. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2003, 523, 173–182. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, Y.; Kim, M.Y.; Chun, Y.J.; Kim, S. Potent inhibition of recombinant human cytochrome p-450 1a1 by pentamethoxystilbene. J. Toxicol. Environ. Health Part A Curr. Issues 2004, 67, 1987–2000. [Google Scholar] [CrossRef]
- Pan, M.H.; Lin, C.L.; Tsai, J.H.; Ho, C.T.; Chen, W.J. 3,5,3′,4′,5′-pentamethoxystilbene (mr-5), a synthetically methoxylated analogue of resveratrol, inhibits growth and induces g1 cell cycle arrest of human breast carcinoma mcf-7 cells. J. Agric. Food Chem. 2010, 58, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Li, H.T.; Wu, W.K.K.; Zheng, Z.P.; Che, C.T.; Yu, L.; Li, Z.J.; Wu, Y.C.; Cheng, K.W.; Yu, J.; Cho, C.H.; et al. 2,3′,4,4′,5′-pentamethoxy-trans-stilbene, a resveratrol derivative, is a potent inducer of apoptosis in colon cancer cells via targeting microtubules. Biochem. Pharm. 2009, 78, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Spatafora, C.; Tringali, C.; Ho, P.C. Determination of trans-2,4,3′,4′,5′-pentamethoxystilbene in rat plasma and its application to a pharmacokinetic study. J. Pharm. Biomed. Anal. 2012, 57, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Coppa, T.; Lazze, M.C.; Cazzalini, O.; Perucca, P.; Pizzala, R.; Bianchi, L.; Stivala, L.A.; Forti, L.; Maccario, C.; Vannini, V.; et al. Structure-activity relationship of resveratrol and its analogue, 4,4′-dihydroxy-trans-stilbene, toward the endothelin axis in human endothelial cells. J. Med. Food 2011, 14, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Fang, J.G.; Chen, W.F.; Zhou, B.; Yang, L.; Liu, Z.L. Structure-activity relationship studies of resveratrol and its analogues by the reaction kinetics of low density lipoprotein peroxidation. Bioorg. Chem. 2006, 34, 142–157. [Google Scholar] [CrossRef]
- Saha, B.; Subramanian, M.; Gupta, P.; Patro, B.S.; Ray, J.; Bandyopadhyay, S.K.; Chattopadhyay, S. Trans-4,4′-dihydroxystilbene (dhs) protects pc12 cells from oxidative damage but induces reactive oxygen species-mediated apoptosis in shsy-5y cell line. Indian J. Exp. Biol. 2016, 54, 719–728. [Google Scholar]
- Chen, C.W.; Li, Y.; Hu, S.; Zhou, W.; Meng, Y.; Li, Z.; Zhang, Y.; Sun, J.; Bo, Z.; DePamphilis, M.L.; et al. Dhs (trans-4,4′-dihydroxystilbene) suppresses DNA replication and tumor growth by inhibiting rrm2 (ribonucleotide reductase regulatory subunit m2). Oncogene 2018, 1. [Google Scholar] [CrossRef]
- Saha, B.; Pai, G.B.; Subramanian, M.; Gupta, P.; Tyagi, M.; Patro, B.S.; Chattopadhyay, S. Resveratrol analogue, trans-4,4′-dihydroxystilbene (dhs), inhibits melanoma tumor growth and suppresses its metastatic colonization in lungs. Biomed. Pharmacother. 2018, 107, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Patro, B.S.; Koli, M.; Pai, G.; Ray, J.; Bandyopadhyay, S.K.; Chattopadhyay, S. Trans-4,4′-dihydroxystilbene (dhs) inhibits human neuroblastoma tumor growth and induces mitochondrial and lysosomal damages in neuroblastoma cell lines. Oncotarget 2017, 8, 73905–73924. [Google Scholar] [CrossRef]
- Maccario, C.; Savio, M.; Ferraro, D.; Bianchi, L.; Pizzala, R.; Pretali, L.; Forti, L.; Stivala, L.A. The resveratrol analog 4,4′-dihydroxy-trans-stilbene suppresses transformation in normal mouse fibroblasts and inhibits proliferation and invasion of human breast cancer cells. Carcinogenesis 2012, 33, 2172–2180. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, H.; Kucinska, M.; Murias, M. Biological activity of piceatannol: Leaving the shadow of resveratrol. Mutat. Res. Rev. Mutat. Res. 2012, 750, 60–82. [Google Scholar] [CrossRef] [PubMed]
- Setoguchi, Y.; Oritani, Y.; Ito, R.; Inagaki, H.; Maruki-Uchida, H.; Ichiyanagi, T.; Ito, T. Absorption and metabolism of piceatannol in rats. J. Agric. Food Chem. 2014, 62, 2541–2548. [Google Scholar] [CrossRef] [PubMed]
- Brents, L.K.; Medina-Bolivar, F.; Seely, K.A.; Nair, V.; Bratton, S.M.; Nopo-Olazabal, L.; Patel, R.Y.; Liu, H.N.; Doerksen, R.J.; Prather, P.L.; et al. Natural prenylated resveratrol analogs arachidin-1 and-3 demonstrate improved glucuronidation profiles and have affinity for cannabinoid receptors. Xenobiotica 2012, 42, 139–156. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, H.; Ito, R.; Setoguchi, Y.; Oritani, Y.; Ito, T. Administration of piceatannol complexed with alpha-cyclodextrin improves its absorption in rats. J. Agric. Food Chem. 2016, 64, 3557–3563. [Google Scholar] [CrossRef]
- Dhanapal, J.; Balaraman Ravindrran, M. Chitosan/poly (lactic acid)-coated piceatannol nanoparticles exert an in vitro apoptosis activity on liver, lung and breast cancer cell lines. Artif. Cells Nanomed. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Messiad, H.; Amira-Guebailia, H.; Houache, O. Reversed phase high performance liquid chromatography used for the physicochemical and thermodynamic characterization of piceatannol/beta-cyclodextrin complex. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 926, 21–27. [Google Scholar] [CrossRef]
- Wang, D.P.; Zhang, Y.F.; Zhang, C.Q.; Gao, L.; Li, J. Piceatannol pretreatment alleviates acute cardiac injury via regulating pi3k-akt-enos signaling in h9c2 cells. Biomed. Pharmacother. 2019, 109, 886–891. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.H.; Chen, X.; Zhang, N.; Li, G. Piceatannol attenuates behavioral disorder and neurological deficits in aging mice via activating the nrf2 pathway. Food Funct. 2018, 9, 371–378. [Google Scholar] [CrossRef]
- Seyed, M.A.; Jantan, I.; Bukhari, S.N.A.; Vijayaraghavan, K. A comprehensive review on the chemotherapeutic potential of piceatannol for cancer treatment, with mechanistic insights. J. Agric. Food Chem. 2016, 64, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, J.H.; Wang, L.; Han, M.; Xiao, F.; Lan, X.Q.; Li, Y.Q.; Xu, G.; Yao, Y. Glucocorticoid receptor agonist dexamethasone attenuates renal ischemia/reperfusion injury by up-regulating enos/inos. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Franceschelli, S.; Pesce, M.; Ferrone, A.; Gatta, D.M.P.; Patruno, A.; De Lutiis, M.A.; Quiles, J.L.; Grilli, A.; Felaco, M.; Speranza, L. Biological effect of licochalcone c on the regulation of pi3k/akt/enos and nf-kappa b/inos/no signaling pathways in h9c2 cells in response to lps stimulation. Int. J. Mol. Sci. 2017, 18, 690. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, K.W.; Lee, H.J. Protective effects of piceatannol against beta-amyloid-induced neuronal cell death. Signal Transduct. Pathw. Part C 2007, 1095, 473–482. [Google Scholar] [CrossRef]
- Jin, C.Y.; Molagoda, I.M.N.; Park, C.; Kwon, T.K.; Yun, S.J.; Kim, W.J.; Kim, G.Y.; Choi, Y.H. Piceatannol-induced apoptosis is reversed by n-acetyl-l-cysteine through restoration of xiap expression. Biol. Pharm. Bull 2018, 41, 1372–1378. [Google Scholar] [CrossRef]
- Ko, H.S.; Lee, H.J.; Kim, S.H.; Lee, E.O. Piceatannol suppresses breast cancer cell invasion through the inhibition of mmp-9: Involvement of pi3k/akt and nf-kappa b pathways. J. Agric. Food Chem. 2012, 60, 4083–4089. [Google Scholar] [CrossRef]
- Kuo, P.L.; Hsu, Y.L. The grape and wine constituent piceatannol inhibits proliferation of human bladder cancer cells via blocking cell cycle progression and inducing fas/membrane bound fas ligand-mediated apoptotic pathway. Mol. Nutr. Food Res. 2008, 52, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Jayasooriya, R.G.P.T.; Lee, Y.G.; Kang, C.H.; Lee, K.T.; Choi, Y.H.; Park, S.Y.; Hwang, J.K.; Kim, G.Y. Piceatannol inhibits mmp-9-dependent invasion of tumor necrosis factor-alpha-stimulated du145 cells by suppressing the akt-mediated nuclear factor-kappa b pathway. Oncol. Lett. 2013, 5, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.C.; Lin, C.Y.; Lin, H.Y.; Wu, J.M. Akt/mtor as novel targets of polyphenol piceatannol possibly contributing to inhibition of proliferation of cultured prostate cancer cells. ISRN Urol. 2012, 2012, 272697. [Google Scholar] [CrossRef]
- Du, M.T.; Zhang, Z.; Gao, T. Piceatannol induced apoptosis through up-regulation of microrna-181a in melanoma cells. Biol. Res. 2017, 50, 36. [Google Scholar] [CrossRef] [PubMed]
- Kita, Y.; Miura, Y.; Yagasaki, K. Antiproliferative and anti-invasive effect of piceatannol, a polyphenol present in grapes and wine, against hepatoma ah109a cells. J. Biomed. Biotechnol. 2012, 2012, 672416. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.; Hsieh, T.C.; Halicka, H.D.; Darzynkiewicz, Z.; Wu, J.M. Upregulation of pd-l1 expression by resveratrol and piceatannol in breast and colorectal cancer cells occurs via hdac3/p300-mediated nf-kappa b signaling. Int. J. Oncol. 2018, 53, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Storniolo, C.E.; Moreno, J.J. Resveratrol analogs with antioxidant activity inhibit intestinal epithelial cancer Caco-2 cell growth by modulating arachidonic acid cascade. J. Agric. Food Chem. 2018, 67, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Park, H.; Park, S.Y.; Jun, J.G.; Park, J.H.Y. The grape component piceatannol induces apoptosis in du145 human prostate cancer cells via the activation of extrinsic and intrinsic pathways. J. Med. Food 2009, 12, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Li, H.T.; Wu, W.K.K.; Zheng, Z.P.; Che, C.T.; Li, Z.J.; Xu, D.D.; Wong, C.C.M.; Ye, C.G.; Sung, J.J.Y.; Cho, C.H.; et al. 3,3′,4,5,5′-pentahydroxy-trans-stilbene, a resveratrol derivative, induces apoptosis in colorectal carcinoma cells via oxidative stress. Eur. J. Pharm. 2010, 637, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kang, N.J.; Rogozin, E.A.; Oh, S.M.; Heo, Y.S.; Pugliese, A.; Bode, A.M.; Lee, H.J.; Dong, Z.G. The resveratrol analogue 3,5,3′,4′,5′-pentahydroxy-trans-stilbene inhibits cell transformation via mek. Int. J. Cancer 2008, 123, 2487–2496. [Google Scholar] [CrossRef] [PubMed]
- Saiko, P.; Horvath, Z.; Murias, M.; Handler, N.; Jaeger, W.; Erker, T.; Fritzer-Szekeres, M.; Szekeres, T. Antitumor effects of 3,3′, 4,4′, 5,5′-hexahydroxystilbene in hl-60 human promyelocytic leukemia cells. Nucleosides Nucleotides Nucleic Acids 2006, 25, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Ovesna, Z.; Kozics, K.; Bader, Y.; Saiko, P.; Handler, N.; Erker, T.; Szekere, T. Antioxidant activity of resveratrol, piceatannol and 3,3′,4,4′,5,5′-hexahydroxy-trans-stilbene in three leukemia cell lines. Oncol. Rep. 2006, 16, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Murias, M.; Luczak, M.W.; Niepsuj, A.; Krajka-Kuzniak, V.; Zielinska-Przyjemska, M.; Jagodzinski, P.P.; Jager, W.; Szekeres, T.; Jodynis-Liebert, J. Cytotoxic activity of 3,3′,4,4′,5,5′-hexahydroxystilbene against breast cancer cells is mediated by induction of p53 and downregulation of mitochondrial superoxide dismutase. Toxicol. In Vitro 2008, 22, 1361–1370. [Google Scholar] [CrossRef]
- Han, Y.S.; Quashie, P.K.; Mesplede, T.; Xu, H.T.; Quan, Y.D.; Jaeger, W.; Szekeres, T.; Wainberg, M.A. A resveratrol analog termed 3,3,4,4,5,5-hexahydroxy-trans-stilbene is a potent hiv-1 inhibitor. J. Med. Virol. 2015, 87, 2054–2060. [Google Scholar] [CrossRef]
- Paulitschke, V.; Schicher, N.; Szekeres, T.; Jager, W.; Elbling, L.; Riemer, A.B.; Scheiner, O.; Trimurtulu, G.; Venkateswarlu, S.; Mikula, M.; et al. 3,3′,4,4′,5,5′-hexahydroxystilbene impairs melanoma progression in a metastatic mouse model. J. Investig. Dermatol. 2010, 130, 1668–1679. [Google Scholar] [CrossRef]
- Mayhoub, A.S.; Marler, L.; Kondratyuk, T.P.; Park, E.J.; Pezzuto, J.M.; Cushman, M. Optimization of the aromatase inhibitory activities of pyridylthiazole analogues of resveratrol. Bioorg. Med. Chem. 2012, 20, 2427–2434. [Google Scholar] [CrossRef]
- Chimento, A.; Sala, M.; Gomez-Monterrey, I.M.; Musella, S.; Bertamino, A.; Caruso, A.; Sinicropi, M.S.; Sirianni, R.; Puoci, F.; Parisi, O.I.; et al. Biological activity of 3-chloro-azetidin-2-one derivatives having interesting antiproliferative activity on human breast cancer cell lines. Bioorg. Med. Chem. Lett. 2013, 23, 6401–6405. [Google Scholar] [CrossRef]
- Lee, E.J.; Min, H.Y.; Park, H.J.; Chung, H.J.; Kim, S.; Han, Y.N.; Lee, S.K. G2/m cell cycle arrest and induction of apoptosis by a stilbenoid, 3,4,5-trimethoxy-4′-bromo-cis-stilbene, in human lung cancer cells. Life Sci. 2004, 75, 2829–2839. [Google Scholar] [CrossRef]
- Sala, M.; Chimento, A.; Saturnino, C.; Gomez-Monterrey, I.M.; Musella, S.; Bertamino, A.; Milite, C.; Sinicropi, M.S.; Caruso, A.; Sirianni, R.; et al. Synthesis and cytotoxic activity evaluation of 2,3-thiazolidin-4-one derivatives on human breast cancer cell lines. Bioorg. Med. Chem. Lett. 2013, 23, 4990–4995. [Google Scholar] [CrossRef]
- Kim, H.J.; Yang, K.M.; Park, Y.S.; Choi, Y.J.; Yun, J.H.; Son, C.H.; Suh, H.S.; Jeong, M.H.; Jo, W.S. The novel resvertrol analogue hs-1793 induces apoptosis via the mitochondrial pathway in murine breast cancer cells. Int. J. Oncol. 2012, 41, 1628–1634. [Google Scholar] [CrossRef]
- Kim, J.A.; Kim, D.H.; Hossain, M.A.; Kim, M.Y.; Sung, B.; Yoon, J.H.; Suh, H.; Jeong, T.C.; Chung, H.Y.; Kim, N.D. Hs-1793, a resveratrol analogue, induces cell cycle arrest and apoptotic cell death in human breast cancer cells. Int. J. Oncol. 2014, 44, 473–480. [Google Scholar] [CrossRef]
- Tang, Y.W.; Shi, C.J.; Yang, H.L.; Cai, P.; Liu, Q.H.; Yang, X.L.; Kong, L.Y.; Wang, X.B. Synthesis and evaluation of isoprenylation-resveratrol dimer derivatives against alzheimer’s disease. Eur. J. Med. Chem. 2019, 163, 307–319. [Google Scholar] [CrossRef]
- Gajjar, K.; Martin-Hirsch, P.L.; Martin, F.L. Cyp1b1 and hormone-induced cancer. Cancer Lett. 2012, 324, 13–30. [Google Scholar] [CrossRef]
- Shimada, T.; Hayes, C.L.; Yamazaki, H.; Amin, S.; Hecht, S.S.; Guengerich, F.P.; Sutter, T.R. Activation of chemically diverse procarcinogens by human cytochrome p-450 1b1. Cancer Res. 1996, 56, 2979–2984. [Google Scholar] [PubMed]
- Hayes, C.L.; Spink, D.C.; Spink, B.C.; Cao, J.Q.; Walker, N.J.; Sutter, T.R. 17 beta-estradiol hydroxylation catalyzed by human cytochrome p450 1b1. Proc. Natl. Acad. Sci. USA 1996, 93, 9776–9781. [Google Scholar] [CrossRef]
- Hosoya, T.; Harada, N.; Mimura, J.; Motohashi, H.; Takahashi, S.; Nakajima, O.; Morita, M.; Kawauchi, S.; Yamamoto, M.; Fujii-Kuriyama, Y. Inducibility of cytochrome p450 1a1 and chemical carcinogenesis by benzo[a]pyrene in ahr repressor-deficient mice. Biochem. Biophys. Res. Commun. 2008, 365, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Ovesna, Z.; Horvathova-Kozics, K. Structure-activity relationship of trans-resveratrol and its analogues. Neoplasma 2005, 52, 450–455. [Google Scholar] [PubMed]
- Lappano, R.; De Marco, P.; De Francesco, E.M.; Chimento, A.; Pezzi, V.; Maggiolini, M. Cross-talk between gper and growth factor signaling. J. Steroid Biochem. Mol. Biol. 2013, 137, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Chimento, A.; Casaburi, I.; Rosano, C.; Avena, P.; De Luca, A.; Campana, C.; Martire, E.; Santolla, M.F.; Maggiolini, M.; Pezzi, V.; et al. Oleuropein and hydroxytyrosol activate gper/gpr30-dependent pathways leading to apoptosis of er-negative skbr3 breast cancer cells. Mol. Nutr. Food Res. 2014, 58, 478–489. [Google Scholar] [CrossRef]




{kind=link}
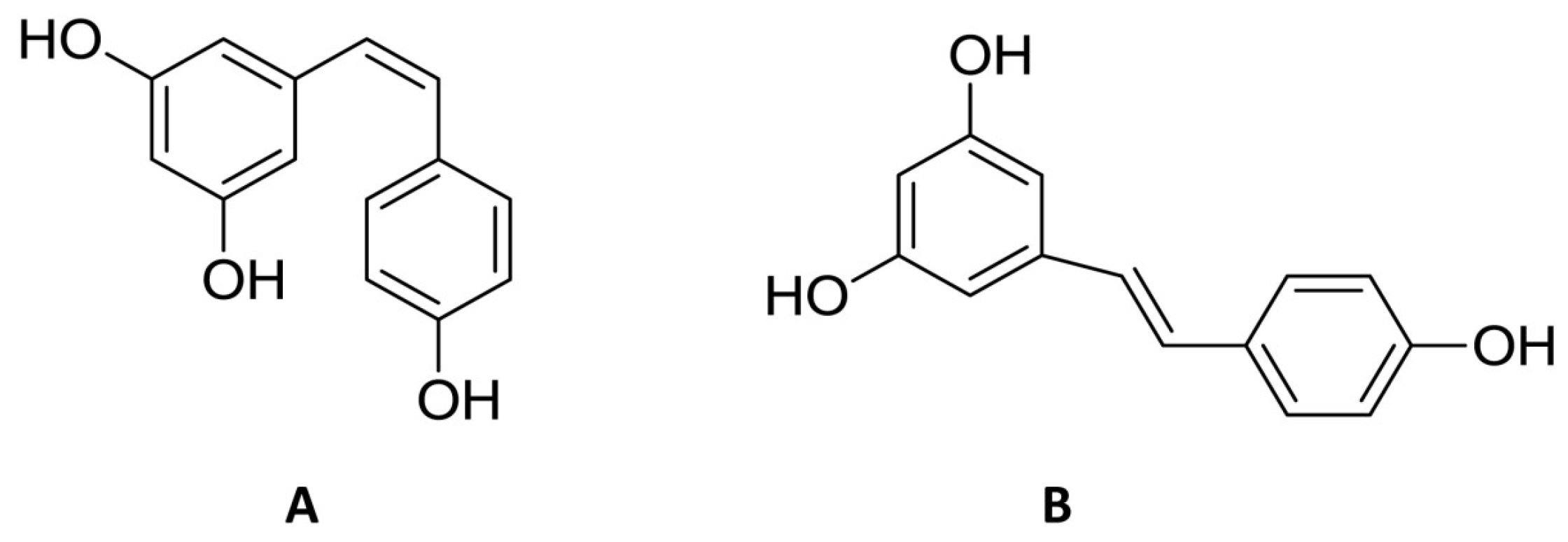{kind=link}
{kind=link}
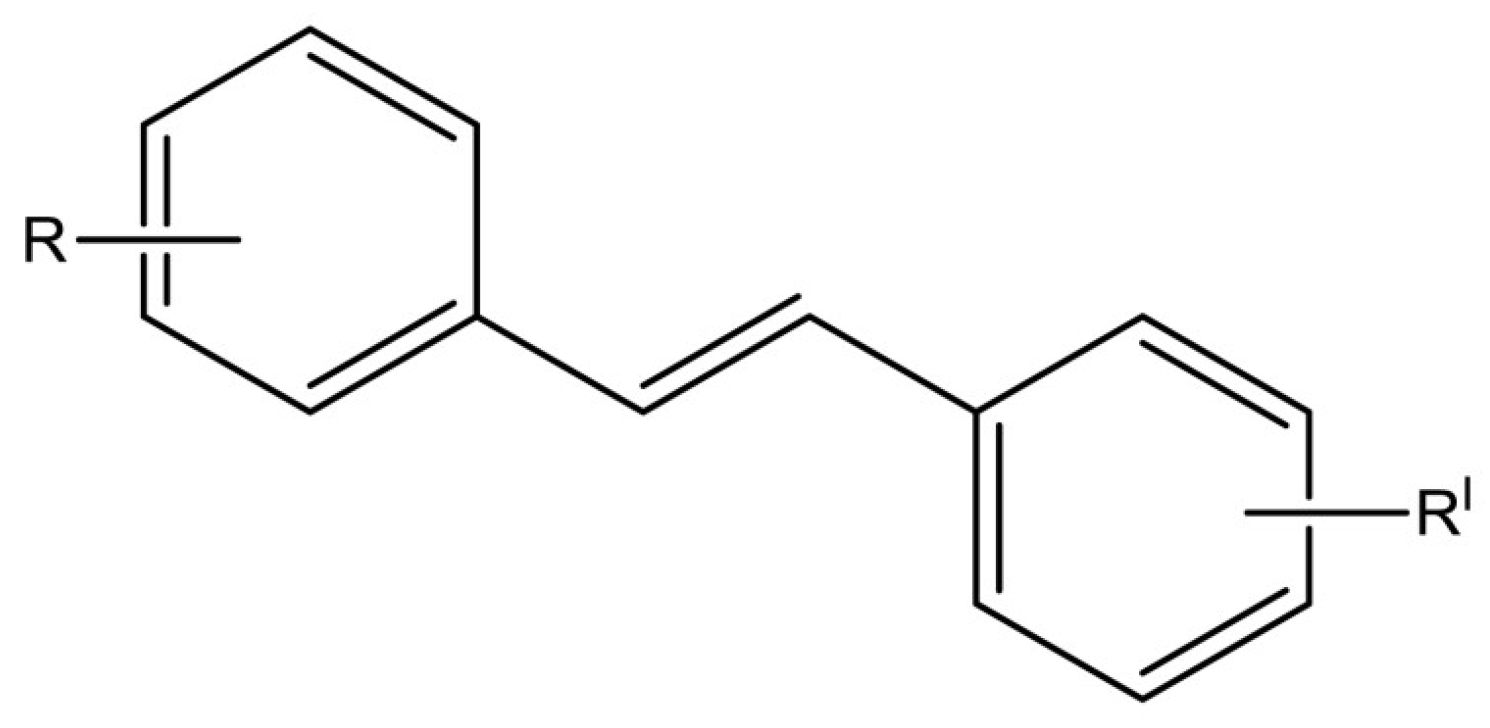{kind=link}
| Biological Effects | Mechanisms | References |
|---|---|---|
| Antioxidant | Decrease of ROS and free radicals; increase of endogenous antioxidant biosynthesis | [6] |
| Reduction of copper-catalyzed oxidation | [7] | |
| Inhibition of LDL peroxidation | [8] | |
| Inhibition of membrane lipids peroxidation | [9] | |
| Decrease of intracellular concentration of ApoB, cholesterol esters and triglycerides secretion rate | [10] | |
| Anti-Inflammatory | Inhibition of COX-1, COX-2, and 5-lipoxygenase catalytic activity | [11] |
| Inhibition of PAF, TNF-α, and histamine | [12] | |
| Cardioprotective | Inhibition of chemotactic factors formation and platelet aggregation | [13,14] |
| Increase of eNOS expression and NO synthesis | [15] | |
| Neuroprotective | Increase of SIRT1 activity | [16] |
| Reduction of cytokines production in activated microglia | [18] | |
| Prevention of free radical-mediated damage through SIRT1 pathway activation | [19] | |
| Antitumor | Inhibition of | |
| Cyclooxygenases | [20,21] | |
| NF-κB | [22] | |
| Kinases such as protein kinase C | [23] | |
| CYPA1 and CYPB1 | [24] | |
| Apoptosis induction and proliferation inhibition in several tumors: | [26] | |
| Lymphoblastic leukemia | [27] | |
| Colon | [28] | |
| Pancreatic | [29] | |
| Melanoma | [30] | |
| Gastric | [31] | |
| Cervical | [32] | |
| Ovarian | [33] | |
| Endometrial | [34] | |
| Liver | [35] | |
| Prostate | [36] | |
| Breast | [37] |
| Delivery System | Models/Methods Used | Results of Studies | References |
|---|---|---|---|
| Lipid Nanocarriers or Liposomes | |||
| Solid lipid nanoparticles (SLNs) and nanostructured lipid carriers (NLCs) | In vitro release simulation studies in liquid dosage forms by cellulose dialysis bag method and in gastrointestinal transit using gastric and intestinal fluid; stability studies by measurements of particle size, polydispersity index, and zeta potential. | Prolonged release over several hours for both nanosystems. ↑ Stability. | [62] |
| Lipid nanocarriers (RSV-nano) or liposomes (RSV-lipo) encapsulating RSV | In vitro release study by dialysis bag method; stability studies by measurements of particle size, polydispersity index, and zeta potential; studies on 3T3-L1 mouse adipocytes cell line | ↑ Solubility and stability. RSV-nano release more prolonged than RSV-lipo. Biological activity: RSV-lipo > RSV-nano. ↑ Cellular content by both RSV-nano and RSV-lipo. No cellular toxicity. | [66] |
| Nanoemulsions | |||
| Nanoemulsions encapsulating RSV | In vitro release study by dialysis bag method; stability studies by measurements of droplet size and polydispersity index; studies on Caco-2 human intestinal cell line | ↑ Solubility and stability. Sustained release. ↑ Membrane passive transport and cellular uptake. No cellular toxicity. | [68] |
| Self-nanoemulsifying drug delivery systems (SNEDDS) | In vitro solubility studies in different solvents; stability studies by measurements of droplet size, polydispersity index, and dispersibility test. | ↑ Solubility and stability. | [69] |
| In vivo studies on Sprague-Dawley rats. | Improved in rate and extent of absorption. ↑ AUC (3.3-fold) and Cmax (2.2-fold) from SNEDDS. The t1/2 and Tmax: RSV-SNEDDS = RSV-solution groups. ↑ Oral bioavailability (3.2-fold). Antifatigue pharmacological effect in rats. | ||
| UDP-glucuronosyltransferase (UGT) inhibitory excipient-based self-microemulsion (SME). | In vitro release study by dialysis bag method; stability studies by measurements of particle size, polydispersity index and zeta potential; studies on Caco-2 human intestinal cell line. | ↑ Solubility and stability. Sustained release in SME1 and SME2. ↑ Cellular uptake and transport. No cellular toxicity. | [70] |
| In vivo studies on male Wistar rats. | ↑ Intestinal permeability and lymphatic transport. ↑ AUC (11.52-fold) and Cmax. (19.14-fold) in rat plasma treated with SME1. ↑ AUC (1.95-fold) and Cmax. (5-fold) in rat plasma treated with SME2. The t1/2 and Tmax: SME1= SM2= RSV free. ↑ Oral bioavailability in rat plasma treated with SME1 (76.1%) and SME2 (12.9%). | ||
| Micelles | |||
| Bile acids micellar solutions. | In vitro solubility studies in buffer solutions of Na-salts of different bile acids; studies on red blood cells from rabbit | ↑ Solubility Micellar solution of 3,7,12-triketocholic acids have lowest membranolytical potential and biggest affinity for RSV solubilization. | [72] |
| Micellar solution of vineatrol30. | In vitro studies on Caco-2 human intestinal cell line. | ↑ Permeability through the cell membrane. | [73] |
| In vivo studies on twelve healthy volunteers (six women and six men). | ↓ Biotransformation during transit through the enterocytes. ↑ AUC (5-fold) and Cmax (10.6-fold) vs. vineatrol30, in all subjects. ↓ Tmax in all subjects. ↑ Urinary excretion (4.5-fold) vs. vineatrol30, in all subjects. ↑ Oral bioavailability (5- fold) vs. vineatrol30. | ||
| Casein nanoparticles encapsulating RSV | In vitro release study using simulated gastric (SGF) and intestinal (SIF) fluids; stability studies by measurements of particle size, polydispersity index and zeta potential. | Controlled release rate. ↑ Stability | [74] |
| In vivo studies on Male Wistar rats. | ↑ Capability to reach the intestinal ephitelium Promotion of RSV intestinal lymphatic transport. ↑ Absorption. ↑ AUC (9.8-fold) and Cmax (1.45-fold). ↑ t1/2 (9-fold) and Tmax (3- fold). ↑ Oral bioavailability (10- fold). | ||
| Polymeric Nanoparticles | |||
| RSV-poly(lactic-co-glycolic acid) (PLGA) nanoparticles (RSV-PLGA-NPs). | In vitro release study by dialysis bag method; stability studies by measurements of particle size, polydispersity index and zeta potential and encapsulation efficiency; studies on HepG2 human hepatoma cell line. | ↑ Solubility and stability. ↑ Encapsulation efficiency and drug loading for RSV. Sustained and slow RSV release from RSV-PLGANPs. ↑ Cellular uptake. ↑RSV-PLGA-NPs bioactivity (lipogenesis reduction, lipolysis promotion and hepatocellular proliferation reduction). | [79] |
| RSV-loaded galactosylated PLGA nanoparticles (RSV-GNPs) | In vitro release study by dialysis bag method; stability studies by measurements of particle size, polydispersity index, zeta potential and encapsulation efficiency; studies on Caco-2 human intestinal cell line; RAW 264.7 macrophage cell line. | Slower drug release in water. ↑ Stability and entrapment efficiency. ↑ Cellular uptake of RSV-GNPs. ↑ Bioactivity (anti-inflammatory efficacy). | [81] |
| In vivo studies on Sprague-Dawley rats. | ↑Permeability and intestinal absorption after oral administration. ↑ AUC (2-fold) and Cmax (1.8- fold). ↑ Oral bioavailability (2-fold). | ||
| Solid Dispersions | |||
| Solid dispersion of RSV on Magnesium DiHydroxide (RSV@MDH) | In vitro solubility study by dissolution test in simulated gastric environment. | ↑ Solubility and dissolution rate. | [59] |
| In vivo studies on New Zealand White hybrid rabbits. | ↑ In vivo absorption of RSV from RSV@MDH. ↑ AUC (3.3-fold) and Cmax (1.33-fold). ↑ Tmax (2-fold). ↑Oral bioavailability (3-fold). | ||
| GPEDP (grape peel extract-loaded dripping pill) into a solid dispersion | In vitro solubility profile by HPLC; dissolution study by the paddle method. | ↑ Solubility and dissolution rate. | [58] |
| In vivo studies on Sprague-Dawley rats. | ↑ Absorption (12-fold). ↑ AUC (1.92-fold) and Cmax (7-fold) vs. GPE. ↑ Oral bioavailability (12 fold higher) vs. GPE. | ||
| Nanocrystals | |||
| RSV nanocrystals (NCs) | In vitro dissolution study by dialysis bag diffusion method; stability studies by measurements of particle size, polydispersity index, zeta potential and drug content remained; studies on MDA-MB231 breast cancer cell line. | ↑ Solubility, stability and dissolution rate. NCs enhanced the RSV delivery in the cells. ↑ RSV bioactivity in NCs form (cell cytotoxicity increase, cell cycle arrest, and apoptosis induction). | [61] |
| In vivo studies on Sprague-Dawley rats. | ↑ Absorption and uptake across the intestinal barrier. ↑ AUC (3.5-fold) and Cmax (2.2-fold). |
| Resveratrol Derivatives | References | Resveratrol Derivatives | References |
|---|---|---|---|
 trans-3,5-dimethoxy-4′-hydroxystilbene (pterostilbene) (1) | [98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119] |  trans-3,4′,5-trimethoxystilbene (2) | [120,121,122,123,124] |
 trans-3,4,5,4′-tetramethoxystilbene (DMU 212 or TMS) (3) | [92,125,126,127,128,129,130,131,132] |  trans-2,4,3′,4′,5′-pentamethoxystilbene (PMS) (4) | [132,133,134,135,136] |
 trans-4,4′-dihydroxystilbene (DHS) (5) | [94,137,138,139,140,141,142,143] |  Trans-3′,4′,3,5-tetrahydroxy-stilbene (piceatannol) (6) | [144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165] |
 3,3′,4,5,5′-pentahydroxy-trans-stilbene (PHS) (7) | [166,167] |  3,3′,4,4′,5,5′-hexahydroxystilbene (8) | [168,169,170,171,172] |
 2,3-thiazolidin-4-one RSV derivatives (9) | [173,174] |  (E)-3,5-difluoro-4′-acetoxystilbene (10) | [96] |
 3,4,5-trimethoxy-4′-bromo-cis-stilbene (11) | [175] |  2,3-thiazolidin-4-one RSV derivatives (12) | [173,176] |
 4-(6-hydroxy-2-naphthalen-2-yl)-1,3-benzenediol (HS-1793) (13) | [177,178] |  [5-((E)-2-(3-(3,5-dihydroxy-4-(3-methylbut-2-en-1-yl)phenyl)-2-(4-hydroxyphenyl)-2,3-dihydrobenzofuran-5-yl)vinyl)-2-(3-methylbut-2-en-1-yl)benzene-1,3-diol] (14) | [179] |
 [5-((E)-2-(3-(5-hydroxy-2,2-dimethylchroman-7-yl)-2-(4-hydroxyphenyl)-2,3-dihydrobenzofuran-5-yl)vinyl)-2-(3-methylbut-2-en-1-yl)benzene-1,3-diol] (15) | [179] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chimento, A.; De Amicis, F.; Sirianni, R.; Sinicropi, M.S.; Puoci, F.; Casaburi, I.; Saturnino, C.; Pezzi, V. Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 1381. https://doi.org/10.3390/ijms20061381
Chimento A, De Amicis F, Sirianni R, Sinicropi MS, Puoci F, Casaburi I, Saturnino C, Pezzi V. Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol. International Journal of Molecular Sciences. 2019; 20(6):1381. https://doi.org/10.3390/ijms20061381
Chicago/Turabian StyleChimento, Adele, Francesca De Amicis, Rosa Sirianni, Maria Stefania Sinicropi, Francesco Puoci, Ivan Casaburi, Carmela Saturnino, and Vincenzo Pezzi. 2019. "Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol" International Journal of Molecular Sciences 20, no. 6: 1381. https://doi.org/10.3390/ijms20061381
APA StyleChimento, A., De Amicis, F., Sirianni, R., Sinicropi, M. S., Puoci, F., Casaburi, I., Saturnino, C., & Pezzi, V. (2019). Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol. International Journal of Molecular Sciences, 20(6), 1381. https://doi.org/10.3390/ijms20061381