Changes in Human Foetal Osteoblasts Exposed to the Random Positioning Machine and Bone Construct Tissue Engineering


 ,
,  ,
,
Abstract
1. Introduction
2. Results
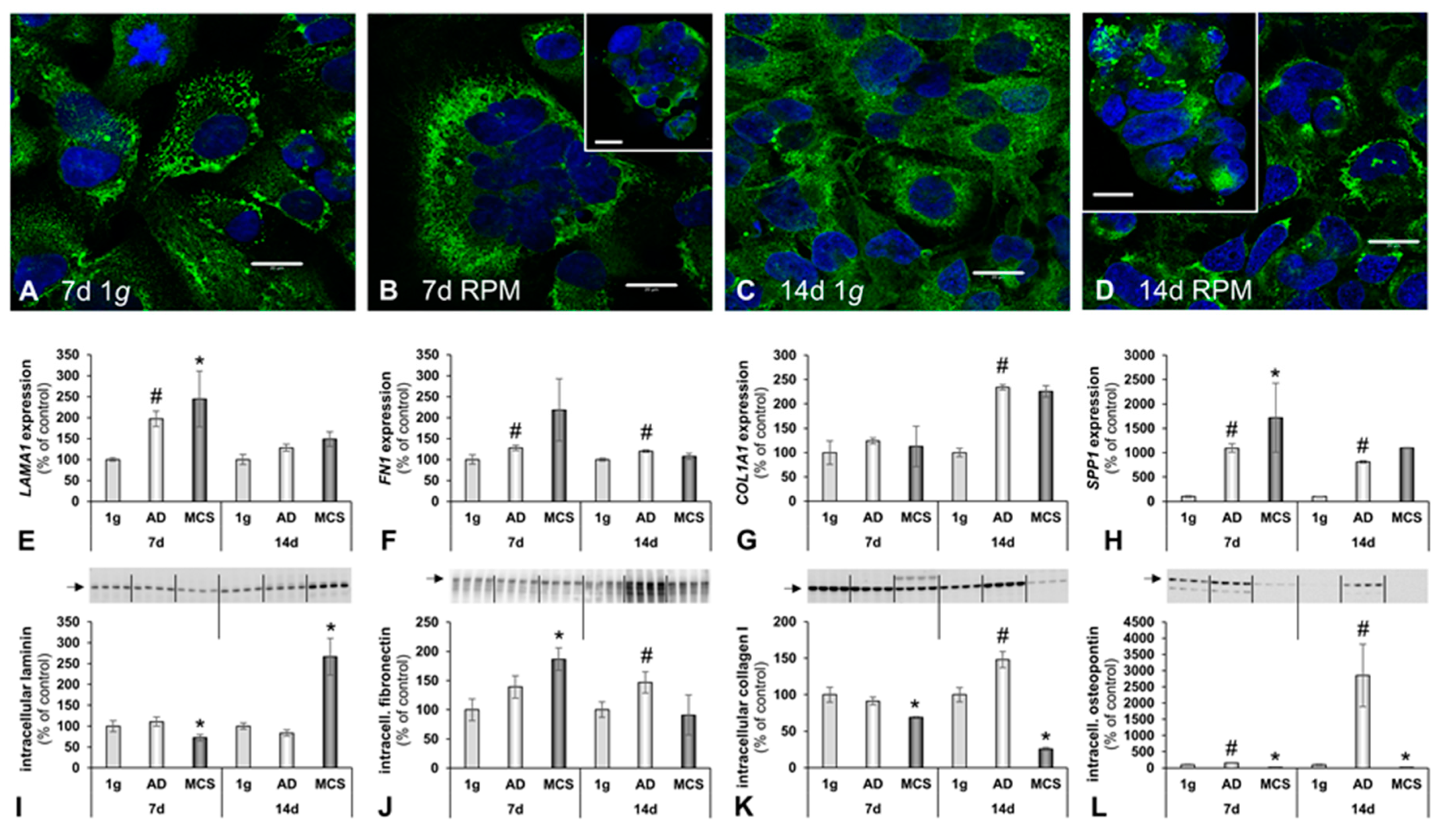
2.1. Morphological Examination of RPM-Exposed hFOB 1.19 Cells Reveal Signs of Organoid Formation
2.2. Alterations of the Cytoskeleton and the Extracellular Matrix
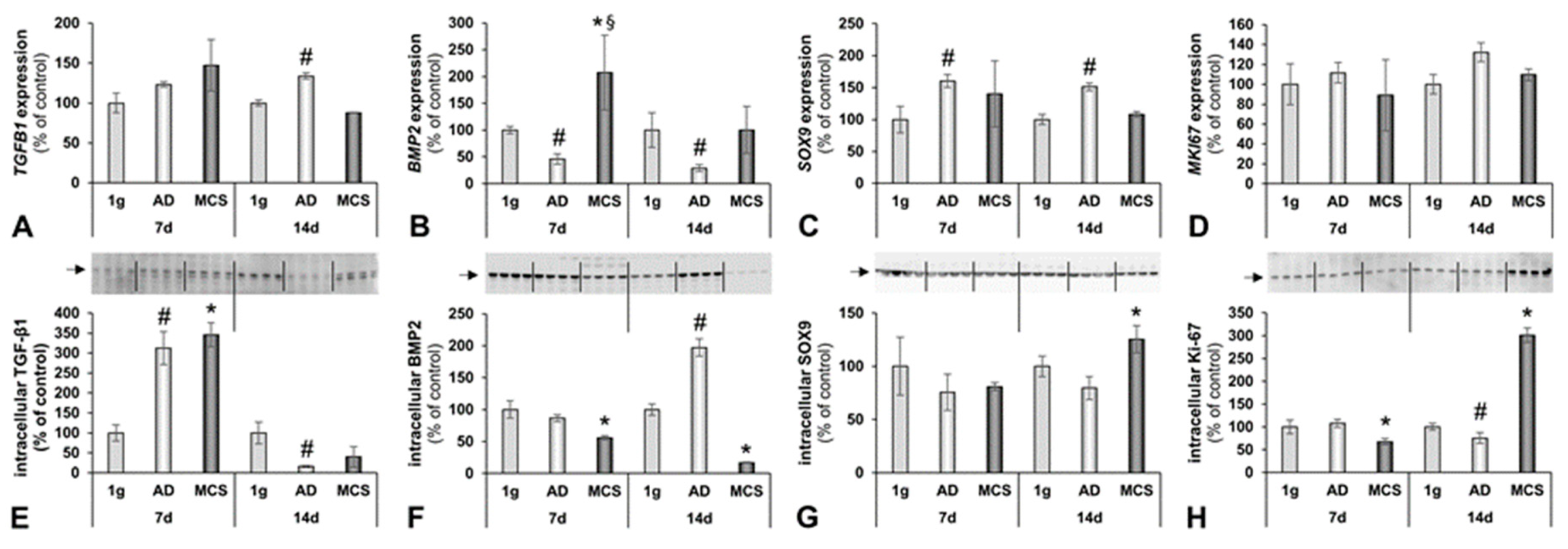
2.3. Changes in BMP2/TGF-β Signalling Pathways
2.4. Secretion of Soluble Factors in the Supernatant
2.5. Interactions of Proteins Involved in Bone Formation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Random Positioning Machine
4.3. F-Actin, Tubulin and Laminin Staining
4.4. Microscopy
4.5. RNA Isolation
4.6. Quantitative Real Time PCR
4.7. Determination of Intracellular Protein Levels
4.8. Measurement of Secreted Bone Biomarkers
4.9. Histological Staining
4.10. Determination of Calcification of the MCS
4.11. Pathway Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| µg | Microgravity |
| 2D | Two- dimensional |
| 3D | Three- dimensional |
| ACTB | Actin beta |
| ACTH | Adrenocorticotrophic hormone |
| AD | Adherent monolayers |
| ALC | Alcian blue |
| BMP2 | Bone morphogenic protein 2 |
| CLSM | Confocal laser scanning microscopy |
| CN | Clinostat |
| COL1A1 | Collagen type 1 alpha 1 |
| Dkk1 | Dickkopf- related protein 1 |
| ECM | Extra cellular matrix |
| FGF-23 | Fibroblast growth factor-23 |
| FN1 | Fibronectin 1 |
| HE | Haematoxylin–Eosin |
| hFOB | Human foetal osteoblast |
| IL-1β | Interleukin 1 beta |
| ISS | International space station |
| LAMA1 | Laminin subunit alpha 1 |
| MCS | Multicellular spheroid |
| MKI67 | Marker of proliferation Ki-67 |
| OC | Osteocalcin |
| OPG | Osteoprotegerin |
| OPN | Osteopontin |
| PTH | Parathyroid hormone |
| qRT-PCR | Quantitative real-time PCR |
| r- | Real |
| RANK | Receptor activator of nuclear factor-κB |
| RANKL | Receptor activator of nuclear factor-κB ligand |
| RCC | Rotating Cell Culture |
| RPM | Random Positioning Machine |
| RWV | Rotating wall vessel |
| s- | Simulated |
| SOST | Sclerostin |
| SOST | Sclerostin |
| SOX9 | SRY-Box 9 |
| SPP1 | Secreted Phosphoprotein 1 |
| TGF-β1 | Transforming growth factor beta 1 |
| TNF-1α | Tumour necrosis factor 1 alpha |
| TUBB | Beta tubulin |
| VIM | Vimentin |
References
- Grimm, D.; Egli, M.; Krüger, M.; Riwaldt, S.; Corydon, T.J.; Kopp, S.; Wehland, M.; Wise, P.; Infanger, M.; Mann, V.; et al. Tissue engineering under microgravity conditions-use of stem cells and specialized cells. Stem Cells Dev. 2018, 27, 787–804. [Google Scholar] [CrossRef]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Qian, A.; Yin, D.; Yang, P.; Jia, B.; Zhang, W.; Shang, P. Development of a ground-based simulated experimental platform for gravitational biology. IEEE Trans. Appl. Supercond. 2009, 19, 42–46. [Google Scholar] [CrossRef]
- Borst, A.G.; van Loon, J.J.W.A. Technology and developments for the random positioning machine, rpm. Microgravity Sci. Technol. 2008, 21, 287. [Google Scholar] [CrossRef]
- Mesland, D. Novel ground-based facilities for research in the effects of weight. ESA Microgravity News 1996, 9, 5–10. [Google Scholar]
- van Loon, J.J.W.A. Some history and use of the random positioning machine, rpm, in gravity related research. Adv. Space Res. 2007, 39, 1161–1165. [Google Scholar] [CrossRef]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated microgravity: Critical review on the use of random positioning machines for mammalian cell culture. Biomed. Res. Int. 2015, 2015, 971474. [Google Scholar] [CrossRef]
- Strauch, S.M.; Grimm, D.; Corydon, T.J.; Krüger, M.; Bauer, J.; Lebert, M.; Wise, P.; Infanger, M.; Richter, P. Current knowledge about the impact of microgravity on the proteome. Expert Review of Proteomics; Taylor & Francis: Thames, UK, 2018; pp. 1–12. [Google Scholar]
- Hammond, T.G.; Lewis, F.C.; Goodwin, T.J.; Linnehan, R.M.; Wolf, D.A.; Hire, K.P.; Campbell, W.C.; Benes, E.; O’Reilly, K.C.; Globus, R.K.; et al. Gene expression in space. Nat. Med. 1999, 5, 359. [Google Scholar] [CrossRef] [PubMed]
- Hammer, B.E.; Kidder, L.S.; Williams, P.C.; Xu, W.W. Magnetic levitation of mc3t3 osteoblast cells as a ground-based simulation of microgravity. Microgravity Sci. Technol. 2009, 21, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Bauer, J.; Kossmehl, P.; Shakibaei, M.; Schoberger, J.; Pickenhahn, H.; Schulze-Tanzil, G.; Vetter, R.; Eilles, C.; Paul, M.; et al. Simulated microgravity alters differentiation and increases apoptosis in human follicular thyroid carcinoma cells. FASEB J. 2002, 16, 604–606. [Google Scholar] [CrossRef]
- Uva, B.M.; Masini, M.A.; Sturla, M.; Bruzzone, F.; Giuliani, M.; Tagliafierro, G.; Strollo, F. Microgravity-induced apoptosis in cultured glial cells. Eur. J. Histochem. 2002, 46, 209–214. [Google Scholar] [CrossRef]
- Masiello, M.G.; Cucina, A.; Proietti, S.; Palombo, A.; Coluccia, P.; D’Anselmi, F.; Dinicola, S.; Pasqualato, A.; Morini, V.; Bizzarri, M. Phenotypic switch induced by simulated microgravity on mda-mb-231 breast cancer cells. Biomed. Res. Int. 2014, 2014, 652434. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.L.; Reynolds, J.L.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight alters microtubules and increases apoptosis in human lymphocytes (jurkat). FASEB J. 1998, 12, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Li, C.F.; Sun, J.X.; Gao, Y.; Shi, F.; Pan, Y.K.; Wang, Y.C.; Sun, X.Q. Clinorotation-induced autophagy via hdm2-p53-mtor pathway enhances cell migration in vascular endothelial cells. Cell Death Dis. 2018, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.W.; Choi, S.H.; Namkoong, S.; Jang, I.S.; Seo, D.H.; Choi, I.; Kim, H.S.; Park, J. Simulated microgravity contributes to autophagy induction by regulating amp-activated protein kinase. DNA Cell Biol. 2014, 33, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Pietsch, J.; Wehland, M.; Schulz, H.; Saar, K.; Hubner, N.; Bauer, J.; Braun, M.; Schwarzwalder, A.; Segerer, J.; et al. Differential gene expression profile and altered cytokine secretion of thyroid cancer cells in space. FASEB J. 2014, 28, 813–835. [Google Scholar] [CrossRef]
- Corydon, T.J.; Kopp, S.; Wehland, M.; Braun, M.; Schutte, A.; Mayer, T.; Hulsing, T.; Oltmann, H.; Schmitz, B.; Hemmersbach, R.; et al. Alterations of the cytoskeleton in human cells in space proved by life-cell imaging. Sci. Rep. 2016, 6, 20043. [Google Scholar] [CrossRef]
- Moes, M.J.A.; Gielen, J.C.; Bleichrodt, R.-J.; van Loon, J.J.W.A.; Christianen, P.C.M.; Boonstra, J. Simulation of microgravity by magnetic levitation and random positioning: Effect on human a431 cell morphology. Microgravity Sci. Technol. 2011, 23, 249–261. [Google Scholar] [CrossRef]
- Unsworth, B.R.; Lelkes, P.I. Growing tissues in microgravity. Nat. Med. 1998, 4, 901–907. [Google Scholar] [CrossRef]
- Kopp, S.; Warnke, E.; Wehland, M.; Aleshcheva, G.; Magnusson, N.E.; Hemmersbach, R.; Corydon, T.J.; Bauer, J.; Infanger, M.; Grimm, D. Mechanisms of three-dimensional growth of thyroid cells during long-term simulated microgravity. Sci. Rep. 2015, 5, 16691. [Google Scholar] [CrossRef]
- Kopp, S.; Slumstrup, L.; Corydon, T.J.; Sahana, J.; Aleshcheva, G.; Islam, T.; Magnusson, N.E.; Wehland, M.; Bauer, J.; Infanger, M.; et al. Identifications of novel mechanisms in breast cancer cells involving duct-like multicellular spheroid formation after exposure to the random positioning machine. Sci. Rep. 2016, 6, 26887. [Google Scholar] [CrossRef]
- Kopp, S.; Sahana, J.; Islam, T.; Petersen, A.G.; Bauer, J.; Corydon, T.J.; Schulz, H.; Saar, K.; Huebner, N.; Slumstrup, L.; et al. The role of nfkappab in spheroid formation of human breast cancer cells cultured on the random positioning machine. Sci. Rep. 2018, 8, 921. [Google Scholar] [CrossRef]
- Svejgaard, B.; Wehland, M.; Ma, X.; Kopp, S.; Sahana, J.; Warnke, E.; Aleshcheva, G.; Hemmersbach, R.; Hauslage, J.; Grosse, J.; et al. Common effects on cancer cells exerted by a random positioning machine and a 2d clinostat. PLoS ONE 2015, 10, e0135157. [Google Scholar] [CrossRef]
- Grosse, J.; Wehland, M.; Pietsch, J.; Schulz, H.; Saar, K.; Hubner, N.; Eilles, C.; Bauer, J.; Abou-El-Ardat, K.; Baatout, S.; et al. Gravity-sensitive signaling drives 3-dimensional formation of multicellular thyroid cancer spheroids. FASEB J. 2012, 26, 5124–5140. [Google Scholar] [CrossRef]
- Dittrich, A.; Grimm, D.; Sahana, J.; Bauer, J.; Krüger, M.; Infanger, M.; Magnusson, N.E. Key proteins involved in spheroid formation and angiogenesis in endothelial cells after long-term exposure to simulated microgravity. Cell Physiol. Biochem. 2018, 45, 429–445. [Google Scholar] [CrossRef]
- Grimm, D.; Bauer, J.; Ulbrich, C.; Westphal, K.; Wehland, M.; Infanger, M.; Aleshcheva, G.; Pietsch, J.; Ghardi, M.; Beck, M.; et al. Different responsiveness of endothelial cells to vascular endothelial growth factor and basic fibroblast growth factor added to culture media under gravity and simulated microgravity. Tissue Eng. Part A 2010, 16, 1559–1573. [Google Scholar] [CrossRef]
- Ulbrich, C.; Westphal, K.; Pietsch, J.; Winkler, H.D.; Leder, A.; Bauer, J.; Kossmehl, P.; Grosse, J.; Schoenberger, J.; Infanger, M.; et al. Characterization of human chondrocytes exposed to simulated microgravity. Cell Physiol. Biochem. 2010, 25, 551–560. [Google Scholar] [CrossRef]
- Aleshcheva, G.; Bauer, J.; Hemmersbach, R.; Egli, M.; Wehland, M.; Grimm, D. Tissue engineering of cartilage on ground-based facilities. Microgravity Sci. Tech. 2016, 28, 237–245. [Google Scholar] [CrossRef]
- Boehrs, J.; Zaharias, R.S.; Laffoon, J.; Ko, Y.J.; Schneider, G.B. Three-dimensional culture environments enhance osteoblast differentiation. J. Prosthodont. 2008, 17, 517–521. [Google Scholar] [CrossRef]
- Schneider, G.B.; Boehrs, J.K.; Hoopes, J.V.; Seabold, D.A. Use of 3-dimensional environments to engineer osseous-like tissue. J. Dev. Biol. Tissue Eng. 2011, 3, 42–47. [Google Scholar]
- Amini, A.R.; Laurencin, C.T.; Nukavarapu, S.P. Bone tissue engineering: Recent advances and challenges. Crit. Rev. Biomed. Eng. 2012, 40, 363–408. [Google Scholar] [CrossRef]
- Su, G.N.; Hidaka, M.; Kimura, Y.; Yamamoto, G. In situ collagen gelation: A new method for constructing large tissue in rotary culture vessels. In Vitro Cell Dev. Biol. Anim. 2003, 39, 368–374. [Google Scholar] [CrossRef]
- Hidaka, M.; Su, G.N.; Chen, J.K.; Mukaisho, K.; Hattori, T.; Yamamoto, G. Transplantation of engineered bone tissue using a rotary three-dimensional culture system. In Vitro Cell Dev. Biol. Anim. 2007, 43, 49–58. [Google Scholar] [CrossRef]
- Ulbrich, C.; Wehland, M.; Pietsch, J.; Aleshcheva, G.; Wise, P.; van Loon, J.; Magnusson, N.; Infanger, M.; Grosse, J.; Eilles, C.; et al. The impact of simulated and real microgravity on bone cells and mesenchymal stem cells. Biomed. Res. Int. 2014, 2014, 928507. [Google Scholar] [CrossRef]
- Clarke, M.S.; Sundaresan, A.; Vanderburg, C.R.; Banigan, M.G.; Pellis, N.R. A three-dimensional tissue culture model of bone formation utilizing rotational co-culture of human adult osteoblasts and osteoclasts. Acta Biomater. 2013, 9, 7908–7916. [Google Scholar] [CrossRef]
- Harris, S.A.; Tau, K.R.; Enger, R.J.; Toft, D.O.; Riggs, B.L.; Spelsberg, T.C. Estrogen response in the hfob 1.19 human fetal osteoblastic cell line stably transfected with the human estrogen receptor gene. J. Cell. Biochem. 1995, 59, 193–201. [Google Scholar] [CrossRef]
- Subramaniam, M.; Jalal, S.M.; Rickard, D.J.; Harris, S.A.; Bolander, M.E.; Spelsberg, T.C. Further characterization of human fetal osteoblastic hfob 1.19 and hfob/er alpha cells: Bone formation in vivo and karyotype analysis using multicolor fluorescent in situ hybridization. J. Cell. Biochem. 2002, 87, 9–15. [Google Scholar] [CrossRef]
- Maniatopoulos, C.; Sodek, J.; Melcher, A.H. Bone formation in vitro by stromal cells obtained from bone marrow of young adult rats. Cell Tissue Res. 1988, 254, 317–330. [Google Scholar] [CrossRef]
- Xu, H.; Wu, F.; Zhang, H.; Yang, C.; Li, K.; Wang, H.; Yang, H.; Liu, Y.; Ding, B.; Tan, Y.; et al. Actin cytoskeleton mediates bmp2-smad signaling via calponin 1 in preosteoblast under simulated microgravity. Biochimie 2017, 138, 184–193. [Google Scholar] [CrossRef]
- Aisha, M.D.; Nor-Ashikin, M.N.; Sharaniza, A.B.; Nawawi, H.; Froemming, G.R. Orbital fluid shear stress promotes osteoblast metabolism, proliferation and alkaline phosphates activity in vitro. Exp. Cell Res. 2015, 337, 87–93. [Google Scholar] [CrossRef]
- Viale-Bouroncle, S.; Gosau, M.; Morsczeck, C. Laminin regulates the osteogenic differentiation of dental follicle cells via integrin-alpha2/-beta1 and the activation of the fak/erk signaling pathway. Cell Tissue Res. 2014, 357, 345–354. [Google Scholar] [CrossRef]
- Bentmann, A.; Kawelke, N.; Moss, D.; Zentgraf, H.; Bala, Y.; Berger, I.; Gasser, J.A.; Nakchbandi, I.A. Circulating fibronectin affects bone matrix, whereas osteoblast fibronectin modulates osteoblast function. J. Bone Miner. Res. 2010, 25, 706–715. [Google Scholar]
- Wu, M.; Chen, G.; Li, Y.P. Tgf-beta and bmp signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Yen, M.L.; Chien, C.C.; Chiu, I.M.; Huang, H.I.; Chen, Y.C.; Hu, H.I.; Yen, B.L. Multilineage differentiation and characterization of the human fetal osteoblastic 1.19 cell line: A possible in vitro model of human mesenchymal progenitors. Stem Cells 2007, 25, 125–131. [Google Scholar] [CrossRef]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; de Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [CrossRef]
- Kamiya, N.; Ye, L.; Kobayashi, T.; Mochida, Y.; Yamauchi, M.; Kronenberg, H.M.; Feng, J.Q.; Mishina, Y. Bmp signaling negatively regulates bone mass through sclerostin by inhibiting the canonical wnt pathway. Development 2008, 135, 3801–3811. [Google Scholar] [CrossRef]
- Gilbert, L.; He, X.; Farmer, P.; Boden, S.; Kozlowski, M.; Rubin, J.; Nanes, M.S. Inhibition of osteoblast differentiation by tumor necrosis factor-alpha. Endocrinology 2000, 141, 3956–3964. [Google Scholar] [CrossRef]
- Abbas, S.; Zhang, Y.H.; Clohisy, J.C.; Abu-Amer, Y. Tumor necrosis factor-alpha inhibits pre-osteoblast differentiation through its type-1 receptor. Cytokine 2003, 22, 33–41. [Google Scholar] [CrossRef]
- Huang, R.L.; Yuan, Y.; Tu, J.; Zou, G.M.; Li, Q. Opposing tnf-alpha/il-1beta- and bmp-2-activated mapk signaling pathways converge on runx2 to regulate bmp-2-induced osteoblastic differentiation. Cell Death Dis. 2014, 5, e1187. [Google Scholar] [CrossRef]
- Neve, A.; Corrado, A.; Cantatore, F.P. Osteocalcin: Skeletal and extra-skeletal effects. J. Cell. Physiol. 2013, 228, 1149–1153. [Google Scholar] [CrossRef]
- Glass, D.A., 2nd; Bialek, P.; Ahn, J.D.; Starbuck, M.; Patel, M.S.; Clevers, H.; Taketo, M.M.; Long, F.; McMahon, A.P.; Lang, R.A.; et al. Canonical wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev. Cell 2005, 8, 751–764. [Google Scholar] [CrossRef]
- Luan, X.; Lu, Q.; Jiang, Y.; Zhang, S.; Wang, Q.; Yuan, H.; Zhao, W.; Wang, J.; Wang, X. Crystal structure of human rankl complexed with its decoy receptor osteoprotegerin. J. Immunol. 2012, 189, 245–252. [Google Scholar] [CrossRef]
- Wu, X.; Guo, S.; Shen, G.; Ma, X.; Tang, C.; Xie, K.; Liu, J.; Guo, W.; Yan, Y.; Luo, E. Screening of osteoprotegerin-related feature genes in osteoporosis and functional analysis with DNA microarray. Eur. J. Med. Res. 2013, 18, 15. [Google Scholar] [CrossRef]
- Saidenberg-Kermanac’h, N.; Cohen-Solal, M.; Bessis, N.; De Vernejoul, M.C.; Boissier, M.C. Role for osteoprotegerin in rheumatoid inflammation. Jt. Bone Spine 2004, 71, 9–13. [Google Scholar] [CrossRef]
- Bae, D.; Moon, S.H.; Park, B.G.; Park, S.J.; Jung, T.; Kim, J.S.; Lee, K.B.; Chung, H.M. Nanotopographical control for maintaining undifferentiated human embryonic stem cell colonies in feeder free conditions. Biomaterials 2014, 35, 916–928. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Kumagai, K.; Imai, S.; Miyatake, K.; Saito, T. Sclerostin is upregulated in the early stage of chondrogenic differentiation, but not required in endochondral ossification in vitro. PLoS ONE 2018, 13, e0201839. [Google Scholar] [CrossRef]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef]
- Dong, Q.; Zhu, X.; Dai, C.; Zhang, X.; Gao, X.; Wei, J.; Sheng, Y.; Zheng, Y.; Yu, J.; Xie, L.; et al. Osteopontin promotes epithelial-mesenchymal transition of hepatocellular carcinoma through regulating vimentin. Oncotarget 2016, 7, 12997–13012. [Google Scholar] [CrossRef]
- Mukherjee, B.B.; Nemir, M.; Beninati, S.; Cordella-Miele, E.; Singh, K.; Chackalaparampil, I.; Shanmugam, V.; DeVouge, M.W.; Mukherjee, A.B. Interaction of osteopontin with fibronectin and other extracellular matrix molecules. Ann. N. Y. Acad. Sci. 1995, 760, 201–212. [Google Scholar] [CrossRef]
- Tocchio, A.; Durmus, N.G.; Sridhar, K.; Mani, V.; Coskun, B.; El Assal, R.; Demirci, U. Magnetically guided self-assembly and coding of 3d living architectures. Adv. Mater. 2018, 30, 1705034. [Google Scholar] [CrossRef]
- Parfenov, V.A.; Koudan, E.V.; Bulanova, E.A.; Karalkin, P.A.; Pereira, F.D.; Norkin, N.E.; Norkin, N.E.; Knyazeva, A.D.; Gryadunova, A.A.; Petrov, O.F.; et al. Scaffold-free, label-free and nozzle-free biofabrication technology using magnetic levitational assembly. Biofabrication 2018, 10, 034104. [Google Scholar] [CrossRef]
- Anil-Inevi, M.; Yaman, S.; Yildiz, A.A.; Mese, G.; Yalcin-Ozuysal, O.; Tekin, H.C.; Ozcivici, E. Biofabrication of in situ self assembled 3d cell cultures in a weightlessness environment generated using magnetic levitation. Sci. Rep. 2018, 8, 7239. [Google Scholar] [CrossRef]
- Sahana, J.; Nassef, M.Z.; Wehland, M.; Kopp, S.; Krüger, M.; Corydon, T.J.; Infanger, M.; Bauer, J.; Grimm, D. Decreased e-cadherin in mcf7 human breast cancer cells forming multicellular spheroids exposed to simulated microgravity. Proteomics 2018, 18, e1800015. [Google Scholar] [CrossRef]
- Ko, Y.J.; Zaharias, R.S.; Seabold, D.A.; Lafoon, J.; Schneider, G.B. Osteoblast differentiation is enhanced in rotary cell culture simulated microgravity environments. J. Prosthodont. 2007, 16, 431–438. [Google Scholar] [CrossRef]
- Vorselen, D.; Roos, W.H.; MacKintosh, F.C.; Wuite, G.J.; van Loon, J.J. The role of the cytoskeleton in sensing changes in gravity by nonspecialized cells. FASEB J. 2014, 28, 536–547. [Google Scholar] [CrossRef]
- Hader, D.P.; Braun, M.; Grimm, D.; Hemmersbach, R. Gravireceptors in eukaryotes-a comparison of case studies on the cellular level. NPJ Microgravity 2017, 3, 13. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef]
- Egashira, K.; Sumita, Y.; Zhong, W.; Takashi, I.; Ohba, S.; Nagai, K.; Asahina, I. Bone marrow concentrate promotes bone regeneration with a suboptimal-dose of rhbmp-2. PLoS ONE 2018, 13, e0191099. [Google Scholar] [CrossRef]
- Qian, X.; Zhang, C.; Chen, G.; Tang, Z.; Liu, Q.; Chen, J.; Tong, X.; Wang, J. Effects of bmp-2 and fgf2 on the osteogenesis of bone marrow-derived mesenchymal stem cells in hindlimb-unloaded rats. Cell Biochem. Biophys. 2014, 70, 1127–1136. [Google Scholar] [CrossRef]
- Sefat, F.; Khaghani, S.A.; Nejatian, T.; Genedy, M.; Abdeldayem, A.; Moghaddam, Z.S.; Denyer, M.C.; Youseffi, M. Transforming growth factor beta (tgf-beta) isomers influence cell detachment of mg-63 bone cells. Tissue Cell 2015, 47, 567–574. [Google Scholar] [CrossRef]
- Zhou, N.; Li, Q.; Lin, X.; Hu, N.; Liao, J.Y.; Lin, L.B.; Zhao, C.; Hu, Z.M.; Liang, X.; Xu, W.; et al. Bmp2 induces chondrogenic differentiation, osteogenic differentiation and endochondral ossification in stem cells. Cell Tissue Res. 2016, 366, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Aleshcheva, G.; Wehland, M.; Sahana, J.; Bauer, J.; Corydon, T.J.; Hemmersbach, R.; Frett, T.; Egli, M.; Infanger, M.; Grosse, J.; et al. Moderate alterations of the cytoskeleton in human chondrocytes after short-term microgravity produced by parabolic flight maneuvers could be prevented by up-regulation of bmp-2 and sox-9. FASEB J. 2015, 29, 2303–2314. [Google Scholar] [CrossRef]
- Gao, L.; Sheu, T.J.; Dong, Y.; Hoak, D.M.; Zuscik, M.J.; Schwarz, E.M.; Hilton, M.J.; O’Keefe, R.J.; Jonason, J.H. Tak1 regulates sox9 expression in chondrocytes and is essential for postnatal development of the growth plate and articular cartilages. J. Cell Sci. 2013, 126, 5704–5713. [Google Scholar] [CrossRef]
- Weber, M.; Sotoca, A.M.; Kupfer, P.; Guthke, R.; van Zoelen, E.J. Dynamic modelling of microrna regulation during mesenchymal stem cell differentiation. BMC Syst. Biol. 2013, 7, 124. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Joiner, D.M.; Oyserman, S.M.; Sharma, P.; Goldstein, S.A.; He, X.; Hauschka, P.V. Bone mass is inversely proportional to dkk1 levels in mice. Bone 2007, 41, 331–339. [Google Scholar] [CrossRef]
- Kenkre, J.S.; Bassett, J. The bone remodelling cycle. Ann. Clin. Biochem. 2018, 55, 308–327. [Google Scholar] [CrossRef]
- Cassuto, J.; Folestad, A.; Gothlin, J.; Malchau, H.; Karrholm, J. The key role of proinflammatory cytokines, matrix proteins, rankl/opg and wnt/beta-catenin in bone healing of hip arthroplasty patients. Bone 2018, 107, 66–77. [Google Scholar] [CrossRef]
- Wei, J.; Karsenty, G. An overview of the metabolic functions of osteocalcin. Rev. Endocr. Metab. Disord. 2015, 16, 93–98. [Google Scholar] [CrossRef]
- Infanger, M.; Kossmehl, P.; Shakibaei, M.; Baatout, S.; Witzing, A.; Grosse, J.; Bauer, J.; Cogoli, A.; Faramarzi, S.; Derradji, H.; et al. Induction of three-dimensional assembly and increase in apoptosis of human endothelial cells by simulated microgravity: Impact of vascular endothelial growth factor. Apoptosis 2006, 11, 749–764. [Google Scholar] [CrossRef]
- Tamplen, M.; Fowler, T.; Markey, J.; Knott, P.D.; Suva, L.J.; Alliston, T. Treatment with anti-sclerostin antibody to stimulate mandibular bone formation. Head Neck 2018, 40, 1453–1460. [Google Scholar] [CrossRef]
- Veeriah, V.; Zanniti, A.; Paone, R.; Chatterjee, S.; Rucci, N.; Teti, A.; Capulli, M. Interleukin-1beta, lipocalin 2 and nitric oxide synthase 2 are mechano-responsive mediators of mouse and human endothelial cell-osteoblast crosstalk. Sci. Rep. 2016, 6, 29880. [Google Scholar] [CrossRef]
- Samad, T.A.; Moore, K.A.; Sapirstein, A.; Billet, S.; Allchorne, A.; Poole, S.; Bonventre, J.V.; Woolf, C.J. Interleukin-1beta-mediated induction of cox-2 in the cns contributes to inflammatory pain hypersensitivity. Nature 2001, 410, 471–475. [Google Scholar] [CrossRef]
- Azuma, Y.; Kaji, K.; Katogi, R.; Takeshita, S.; Kudo, A. Tumor necrosis factor-alpha induces differentiation of and bone resorption by osteoclasts. J. Biol. Chem. 2000, 275, 4858–4864. [Google Scholar] [CrossRef]
- Corydon, T.J.; Mann, V.; Slumstrup, L.; Kopp, S.; Sahana, J.; Askou, A.L.; Magnusson, N.E.; Echegoyen, D.; Bek, T.; Sundaresan, A.; et al. Reduced expression of cytoskeletal and extracellular matrix genes in human adult retinal pigment epithelium cells exposed to simulated microgravity. Cell Physiol. Biochem. 2016, 40, 1–17. [Google Scholar] [CrossRef]
- Pietsch, J.; Bauer, J.; Weber, G.; Nissum, M.; Westphal, K.; Egli, M.; Grosse, J.; Schönberger, J.; Eilles, C.; Infanger, M.; et al. Proteome analysis of thyroid cancer cells after long-term exposure to a random positioning machine. Microgravity Sci. Technol. 2011, 23, 381–390. [Google Scholar] [CrossRef]
- Grimm, D.; Jabusch, H.C.; Kossmehl, P.; Huber, M.; Fredersdorf, S.; Griese, D.P.; Kramer, B.K.; Kromer, E.P. Experimental diabetes and left ventricular hypertrophy: Effects of beta-receptor blockade. Cardiovasc. Pathol. 2002, 11, 229–237. [Google Scholar] [CrossRef]
- Meloan, S.N.; Puchtler, H. Chemical mechanisms of staining methods: Von kossa’s technique: What von kossa really wrote and a modified reaction for selective demonstration of inorganic phosphates. J. Histotechnol. 1985, 8, 11–13. [Google Scholar] [CrossRef]
- Aleshcheva, G.; Sahana, J.; Ma, X.; Hauslage, J.; Hemmersbach, R.; Egli, M.; Infanger, M.; Bauer, J.; Grimm, D. Changes in morphology, gene expression and protein content in chondrocytes cultured on a random positioning machine. PLoS ONE 2013, 8, e79057. [Google Scholar] [CrossRef]
- Dhurjati, R.; Liu, X.; Gay, C.V.; Mastro, A.M.; Vogler, E.A. Extended-term culture of bone cells in a compartmentalized bioreactor. Tissue Eng. 2006, 12, 3045–3054. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA Levels [% of 1 g Control] | |||||||||||||||||||
| Gene | |||||||||||||||||||
| Sample | ACTB | BMP2 | COL1A1 | FN1 | SPP1 | LAMA1 | MKI67 | SOX9 | TGFB1 | TUBB | VIM | ||||||||
| 7 days | 1 g | 100.0 ± 11.0 | 100.0 ± 6.7 | 100.0 ± 24.3 | 100.0 ± 10.8 | 100.0 ± 14.8 | 100.0 ± 3.6 | 100.0 ± 20.6 | 100.0 ± 20.8 | 100.0 ± 12.1 | 100.0 ± 15.1 | 100.0 ± 7.2 | |||||||
| AD | ▲181.2 ± 2.9 | ▼45.6 ± 9.7 | 124.0 ± 6.0 | ▲127.8 ± 6.2 | ▲1094.0 ± 89.0 | ▲197.3 ± 18.4 | 111.4 ± 10.3 | ▲160.1 ± 10.1 | 123.2 ± 3.5 | ▲297.4 ± 27.8 | ▲162.3 ± 8.5 | ||||||||
| MCS | ▲154.7 ± 19.7 | ▲207.2 ± 69.9 | 112.6 ± 41.7 | 218.5 ± 73.9 | ▲1719.2 ± 709.5 | ▲244.4 ± 67.0 | 89.0 ± 35.9 | 140.1 ± 51.6 | 147.1 ± 32.3 | ▲236.2 ± 74.5 | ▲166.7 ± 38.5 | ||||||||
| 14 days | 1 g | 100.0 ± 8.6 | 100.0 ± 32.1 | 100.0 ± 8.6 | 100.0 ± 3.6 | 100.0 ± 3.4 | 100.0 ± 11.5 | 100.0 ± 9.6 | 100.0 ± 8.1 | 100.0 ± 3.8 | 100.0 ± 11.3 | 100.0 ± 11.1 | |||||||
| AD | 113.1 ± 25.8 | ▼28.8 ± 6.5 | ▲234.0 ± 5.3 | ▲119.8 ± 2.7 | ▲813.4 ± 22.2 | 127.9 ± 8.1 | 132.1 ± 9.5 | ▲151.2 ± 5.9 | ▲133.8 ± 4.0 | ▲125.7 ± 4.8 | ▼76.1 ± 3.5 | ||||||||
| MCS | 225.2 ± 61.7 | 100.1 ± 43.8 | 225.7 ± 11.4 | 107.7 ± 7.0 | 1098.2 ± 11.3 | 148.9 ± 17.6 | 109.7 ± 5.8 | 108.0 ± 4.3 | 87.7 ± 0.7 | 67.3 ± 2.7 | 72.1 ± 1.7 | ||||||||
| Intracellular Protein Levels [% of 1 g Control] | |||||||||||||||||||
| Protein | |||||||||||||||||||
| Sample | β-actin | BMP2 | Col 1 | FN | OPN | laminin | Ki-67 | SOX9 | TGF-β1 | β-tubulin | vimentin | ||||||||
| 7 days | 1 g | 100.0 ± 7.5 | 100.0 ± 13.6 | 100.0 ± 10.4 | 100.0 ± 18.8 | 100.0 ± 16.6 | 100.0 ± 13.2 | 100.0 ± 14.8 | 100.0 ± 27.3 | 100.0 ± 20.1 | 100.0 ± 8.2 | 100.0 ± 6.9 | |||||||
| AD | 105.8 ± 12.9 | 86.5 ± 5.4 | 91.4 ± 5.2 | 139.2 ± 19.2 | ▲154.4 ± 13.2 | 110.7 ± 11.1 | 107.8 ± 9.1 | 75.3 ± 17.0 | ▲312.1 ± 41.4 | ▼59.7 ± 23.9 | 107.4 ± 29.3 | ||||||||
| MCS | 83.2 ± 10.4 | ▼55.7 ± 3.2 | ▼68.8 ± 1.1 | ▲186.9 ± 19.1 | ▼19.4 ± 7.5 | ▼72.1 ± 7.7 | ▼67.5 ± 6.5 | 80.9 ± 3.7 | ▲345.9 ± 30.4 | ▼44.3 ± 5.6 | ▼21.5 ± 6.3 | ||||||||
| 14 days | 1 g | 100.0 ± 13.1 | 100.0 ± 8.8 | 100.0 ± 9.8 | 100.0 ± 13.2 | 100.0 ± 24.6 | 100.0 ± 7.6 | 100.0 ± 7.5 | 100.0 ± 9.6 | 100.0 ± 27.3 | 100.0 ± 11.3 | 100.0 ± 19.2 | |||||||
| AD | 84.8 ± 8.0 | ▲196.9 ± 13.6 | ▲148.1 ± 11.1 | ▲146.9 ± 18.0 | ▲2852.2 ± 964.4 | 83.5 ± 7.6 | ▼75.3 ± 12.0 | 79.6 ± 10.8 | ▼15.5 ± 2.4 | ▲188.8 ± 18.0 | 69.3 ± 4.3 | ||||||||
| MCS | 103.9 ± 11.7 | ▼16.9 ± 1.0 | ▼25.5 ± 2.1 | 91.0 ± 34.3 | ▼14.8 ± 17.3 | ▲266.4 ± 44.4 | ▲300.8 ± 15.9 | ▲125.4 ± 12.8 | 39.8 ± 26.0 | ▼35.7 ± 3.8 | ▼1.6 ± 0.8 | ||||||||
| Protein Secretion [% of 1 g Control] | |||||||||||||||||||
| Protein | |||||||||||||||||||
| Sample | DKK1 | IL-1β | OC | OPG | OPN | SOST | TNF-α | ||||||||||||
| 7 days | 1 g | 100.0 ± 1.4 | 100.0 ± 13.0 | 100.0 ± 32.6 | 100.0 ± 15.3 | 100.0 ± 14.7 | 100.0 ± 7.8 | 100.0 ± 8.7 | |||||||||||
| RPM | 98.8 ± 2.5 | ▼34.8 ± 5.7 | 112.0 ± 47.8 | ▼42.6 ± 19.0 | ▲206.7 ± 38.1 | ▼49.4 ± 16.9 | ▼46.2 ± 12.2 | ||||||||||||
| 14 days | 1 g | 100.0 ± 6.6 | 100.0 ± 11.2 | 100.0 ± 24.9 | 100.0 ± 26.2 | 100.0 ± 14.8 | 100.0 ± 32.6 | 100.0 ± 7.5 | |||||||||||
| RPM | 102.1 ± 6.0 | 125.6 ± 13.5 | ▲148.9 ± 28.0 | ▲154.1 ± 22.5 | ▲579.2 ± 20.8 | 74.4 ± 21.4 | 96.0 ± 11.0 | ||||||||||||
| Gene | Primer Name | Sequence |
|---|---|---|
| 18S rRNA | 18S-F | GGAGCCTGCGGCTTAATTT |
| 18S-R | CAACTAAGAACGGCCATGCA | |
| ACTB | ACTB-F | TGCCGACAGGATGCAGAAG |
| ACTB-R | GCCGATCCACACGGAGTACT | |
| BMP2 | BMP2-F | GACCTGTATCGCAGGCACTCA |
| BMP2-R | TCGTTTCTGGTAGTTCTTCCAAAGA | |
| COL1A1 | COL1A1-F | ACGAAGACATCCCACCAATCAC |
| COL1A1-R | CGTTGTCGCAGACGCAGAT | |
| FN1 | FN1-F | TGAGGAGCATGGTTTTAGGAGAA |
| FN1-R | TCCTCATTTACATTCGGCGTATAC | |
| LAMA1 | LAMA1-F | TGCTCATGGTCAATGCTAATCTG |
| LAMA1-R | TCTATCAATCCTCTTCCTTGGACAA | |
| MKI67 | MKI67-F | TGGGGAAAGTAGGTGTGAAAGAAG |
| MKI67-R | CTCCTTAAACGTTCTGATGCTCTTG | |
| SOX9 | SOX9-F | TTGAGACCTTCGACGTCAATGAG |
| SOX9-R | TCTGGCCACGAGTGGCC | |
| SPP1 | SPP1-F | CGAGGTGATAGTGTGGTTTATGGA |
| SPP1-R | CGTCTGTAGCATCAGGGTACTG | |
| TBP | TATA-F | GTGACCCAGCATCACTGTTTC |
| TATA-R | GCAAACCAGAAACCCTTGCG | |
| TGFB1 | TGFB1-F | CACCCGCGTGCTAATGGT |
| TGFB1-R | AGAGCAACACGGGTTCAGGTA | |
| TUBB | TUBB-F | CTGGACCGCATCTCTGTGTACTAC |
| TUBB-R | GACCTGAGCGAACAGAGTCCAT | |
| VIM | VIM-F | TTCAGAGAGAGGAAGCCGAAAAC |
| VIM-R | AGATTCCACTTTGCGTTCAAGGT |
| Antibody Target | Company/No | Host | Size (kDa) | Used Dilution |
|---|---|---|---|---|
| β-actin | Sigma/A5316 | Ms | 42 | 1:2000 |
| BMP2 | Novus biological/ | Rb | 44 | 1:500 |
| Collagen I | Sigma/C2456 | Ms | ~130 | 1:500 |
| Cyclophilin | Abcam/AB178397 | Rb | 20 | 1:2000 |
| Fibronectin | Invitrogen/MA1198 | Ms | 250 | 1:1000 |
| Ki-67 | Santa cruz/c-15402 | Rb | 250 | 1:200 |
| Laminin | Invitrogen/PA1-1673 | Rb | ~200 | 1:1000 |
| Osteopontin | Sigma/07264 | Rb | 50 | 1:1000 |
| SOX9 | Abcam/ab26414 | Rb | 56 | 1:1000 |
| TGF-β1 | Abcam/ab92486 | Rb | 44 | 1:500 |
| TNF-α | Abcam/ab9739 | Rb | 26 | 1:500 |
| β-tubulin | Sigma/T5293 | Ms | 50 | 1:1000 |
| Vimentin | Sigma/WH0007431 | Ms | ~53 | 1:1000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mann, V.; Grimm, D.; Corydon, T.J.; Krüger, M.; Wehland, M.; Riwaldt, S.; Sahana, J.; Kopp, S.; Bauer, J.; Reseland, J.E.; et al. Changes in Human Foetal Osteoblasts Exposed to the Random Positioning Machine and Bone Construct Tissue Engineering. Int. J. Mol. Sci. 2019, 20, 1357. https://doi.org/10.3390/ijms20061357
Mann V, Grimm D, Corydon TJ, Krüger M, Wehland M, Riwaldt S, Sahana J, Kopp S, Bauer J, Reseland JE, et al. Changes in Human Foetal Osteoblasts Exposed to the Random Positioning Machine and Bone Construct Tissue Engineering. International Journal of Molecular Sciences. 2019; 20(6):1357. https://doi.org/10.3390/ijms20061357
Chicago/Turabian StyleMann, Vivek, Daniela Grimm, Thomas J Corydon, Marcus Krüger, Markus Wehland, Stefan Riwaldt, Jayashree Sahana, Sascha Kopp, Johann Bauer, Janne E. Reseland, and et al. 2019. "Changes in Human Foetal Osteoblasts Exposed to the Random Positioning Machine and Bone Construct Tissue Engineering" International Journal of Molecular Sciences 20, no. 6: 1357. https://doi.org/10.3390/ijms20061357
APA StyleMann, V., Grimm, D., Corydon, T. J., Krüger, M., Wehland, M., Riwaldt, S., Sahana, J., Kopp, S., Bauer, J., Reseland, J. E., Infanger, M., Mari Lian, A., Okoro, E., & Sundaresan, A. (2019). Changes in Human Foetal Osteoblasts Exposed to the Random Positioning Machine and Bone Construct Tissue Engineering. International Journal of Molecular Sciences, 20(6), 1357. https://doi.org/10.3390/ijms20061357