New Recombinant Cold-Adapted and Organic Solvent Tolerant Lipase from Psychrophilic Pseudomonas sp. LSK25, Isolated from Signy Island Antarctica

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
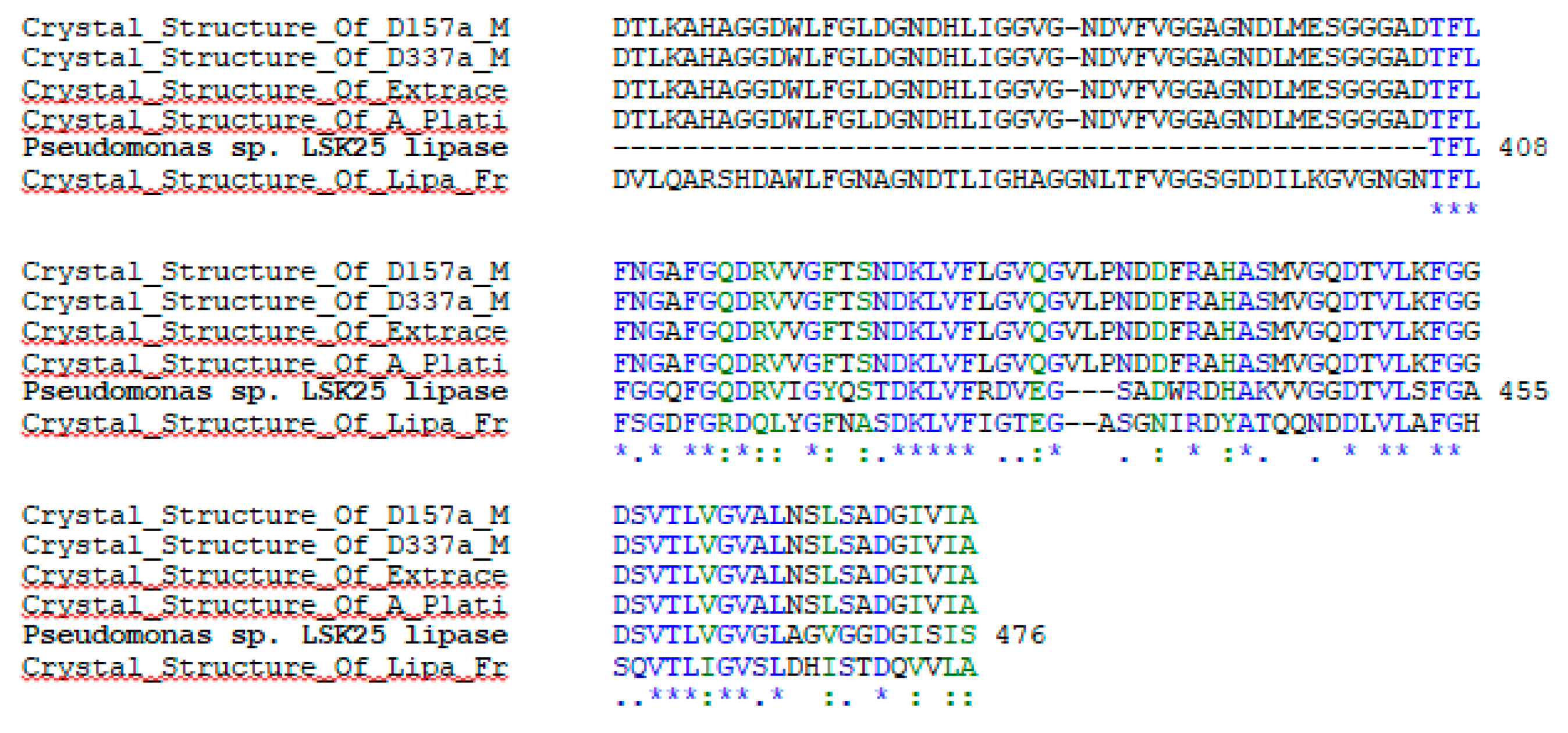
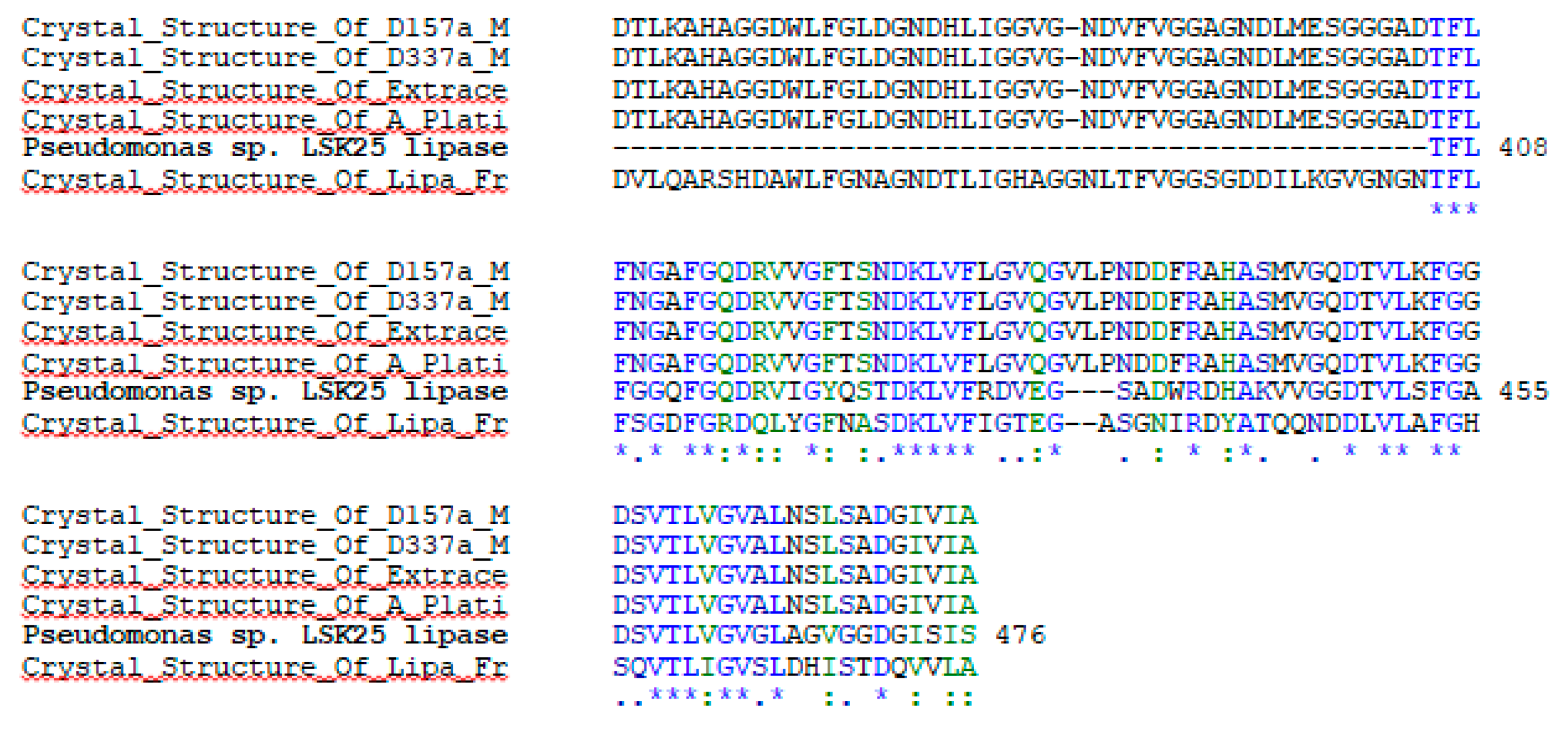
2.1. Sequence Analysis and Expression of LSK25 Lipase
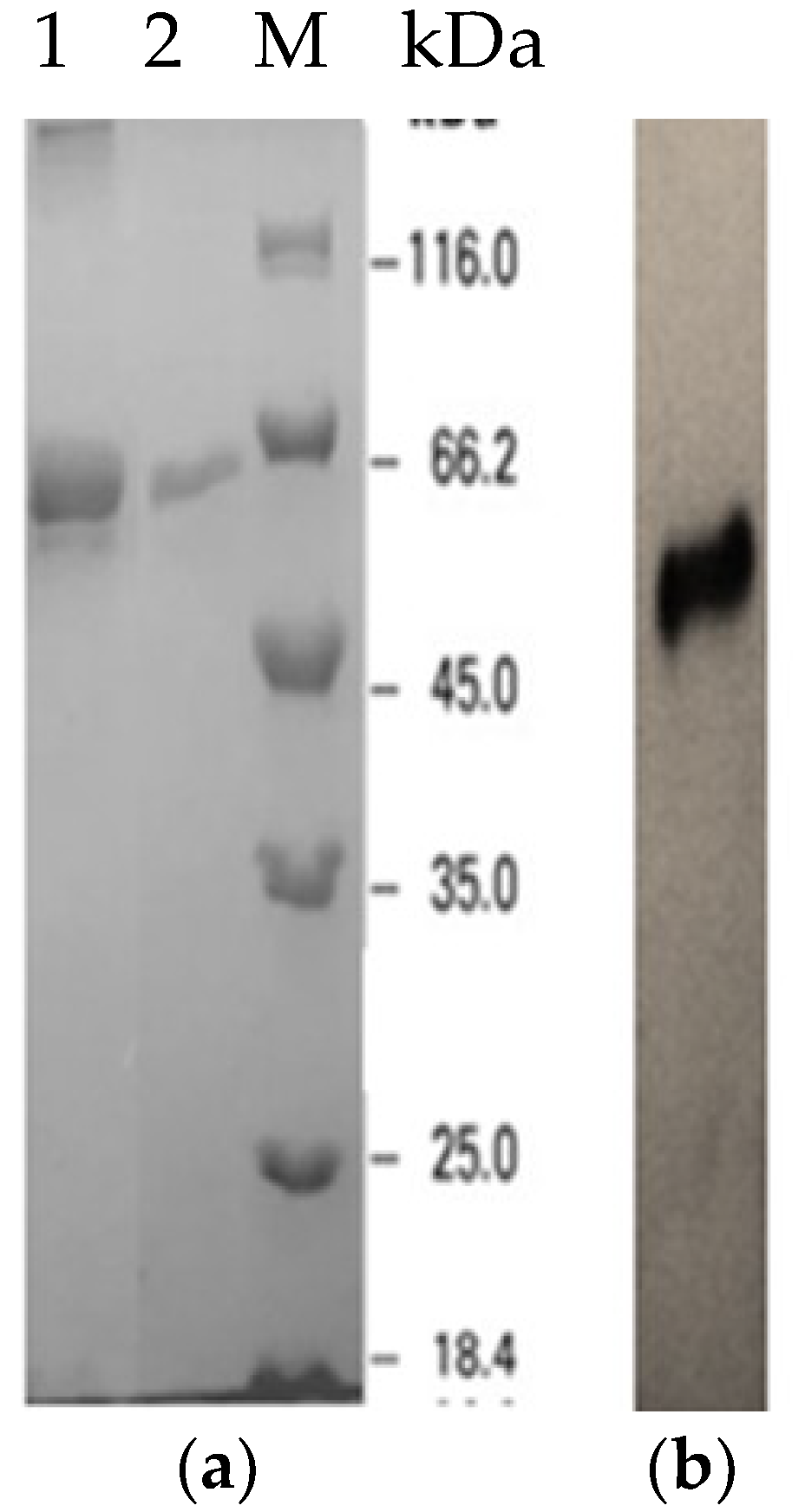
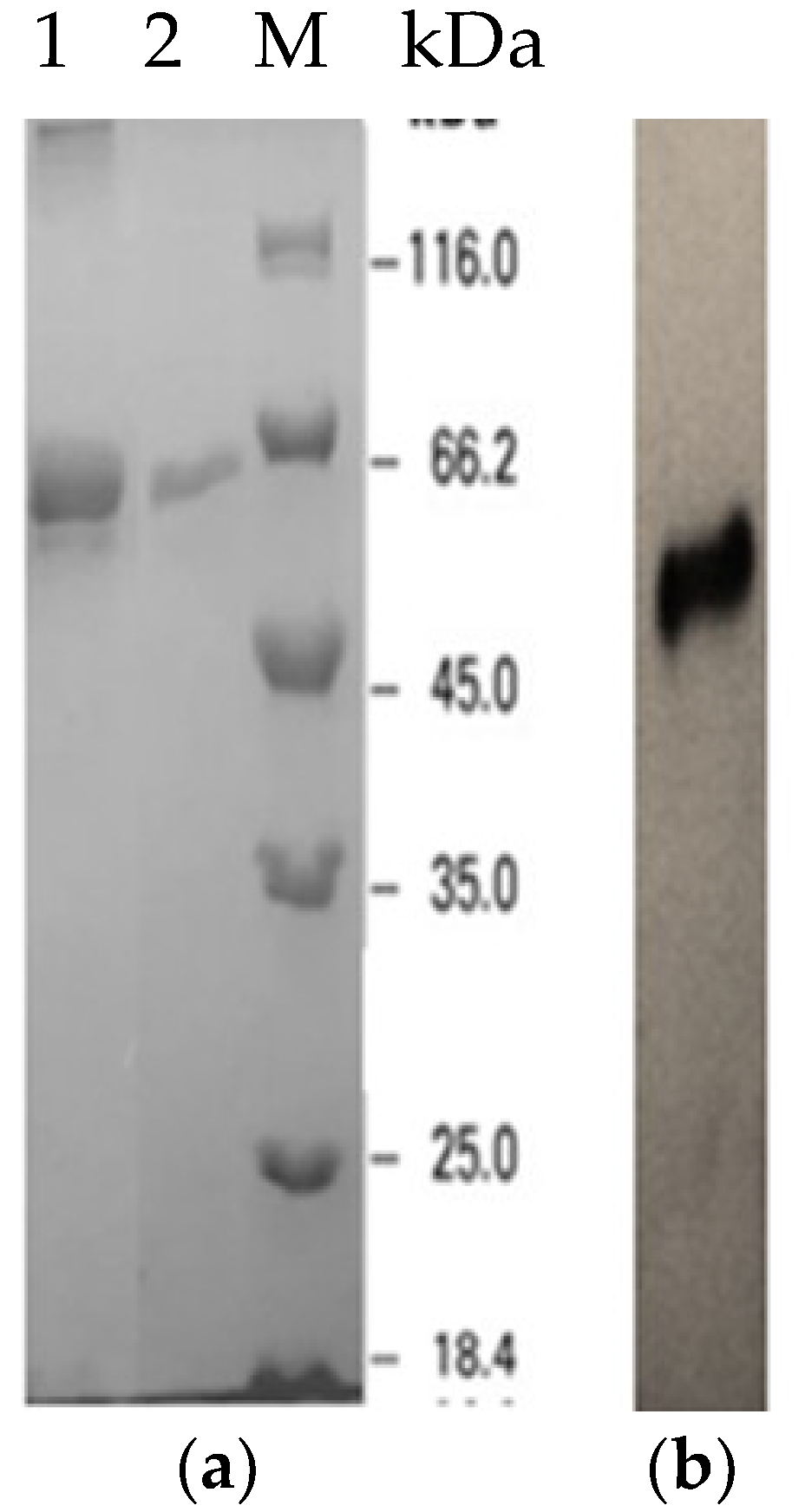
2.2. Expression and Purification of Recombinant LSK25 Lipase
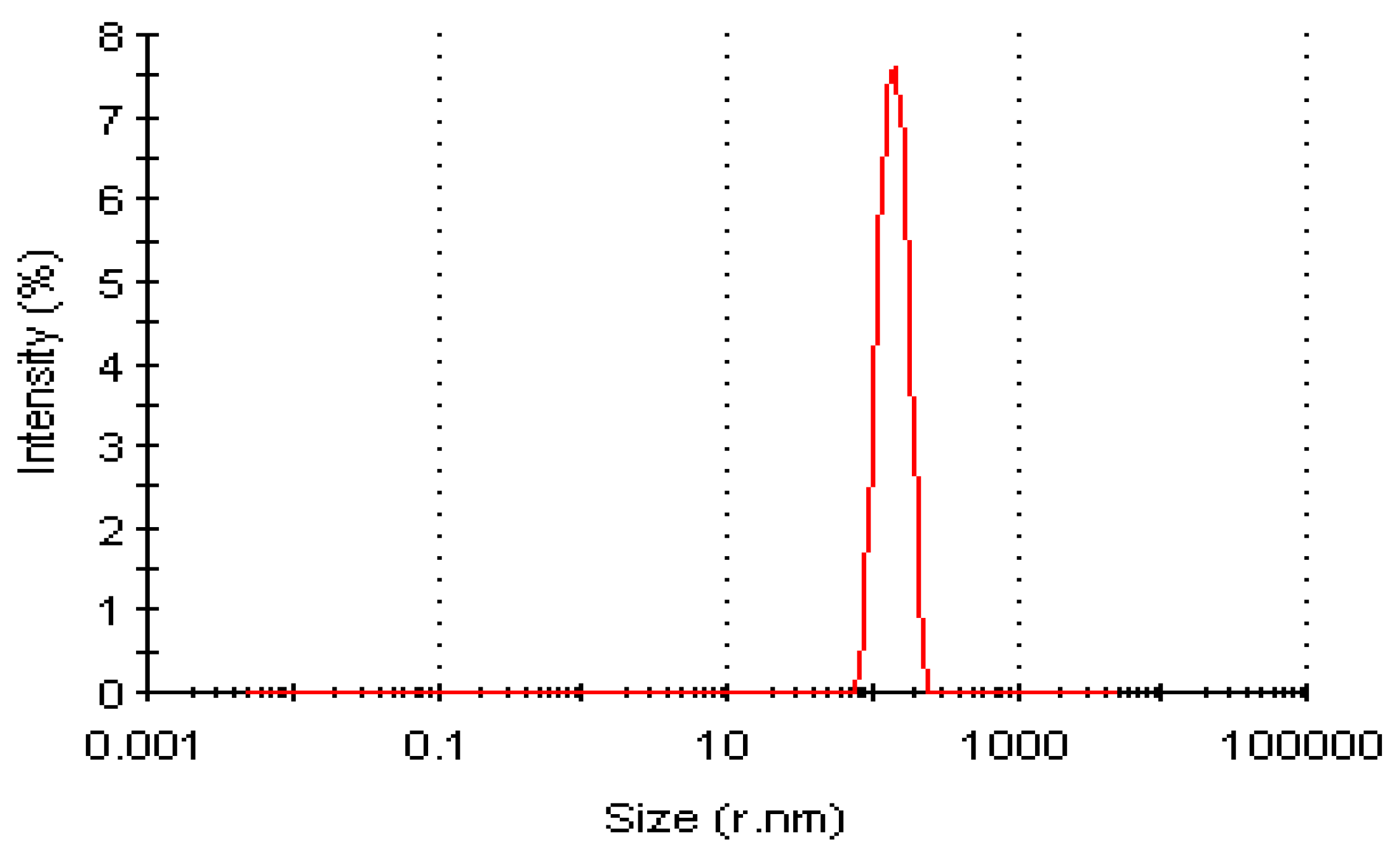
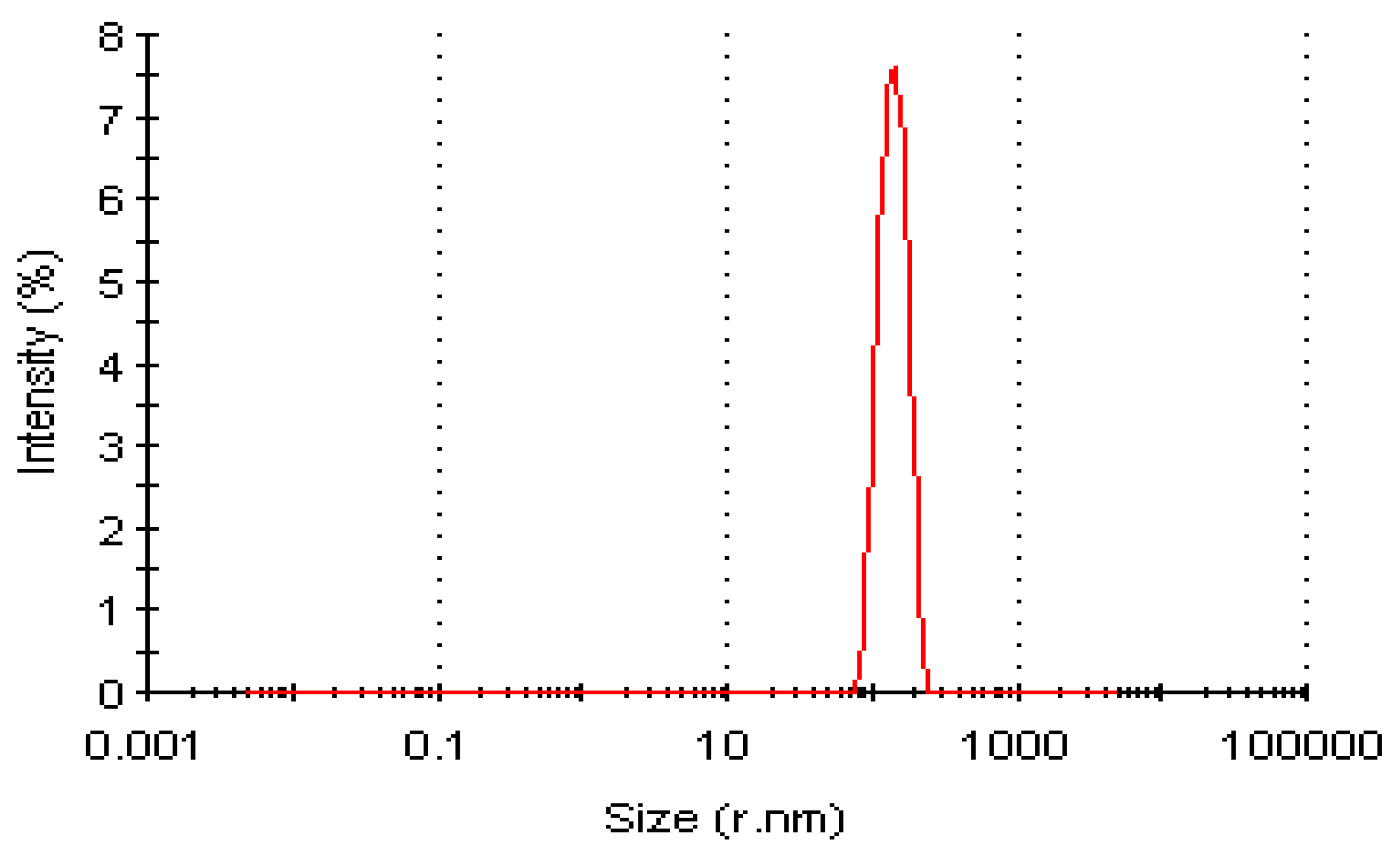
Dynamic Light Scattering Profile of LSK25 Lipase
2.3. Characterization of Purified LSK25 Lipase
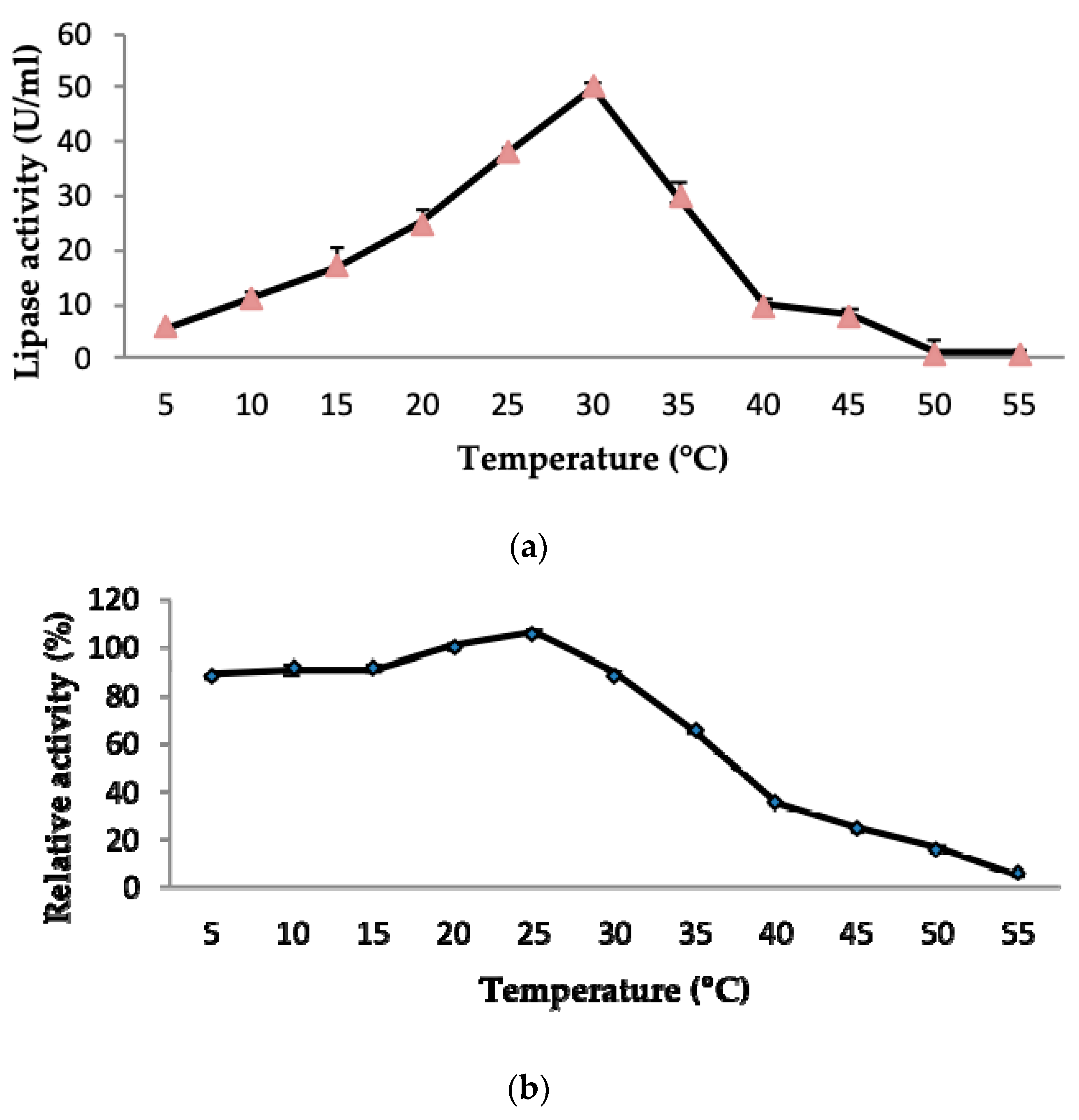
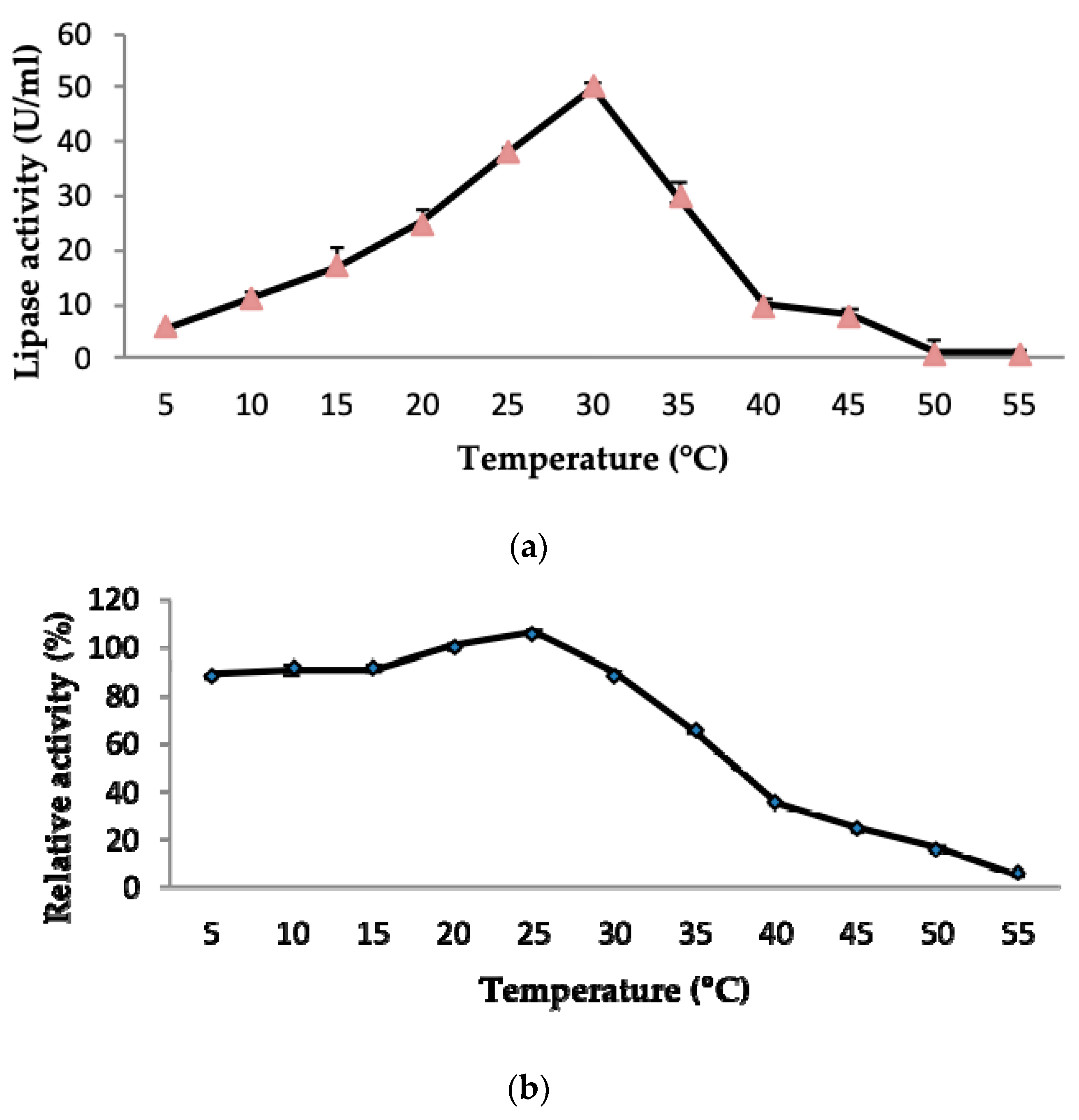
2.3.1. Effect of Temperature on LSK25 Lipase Activity and Stability
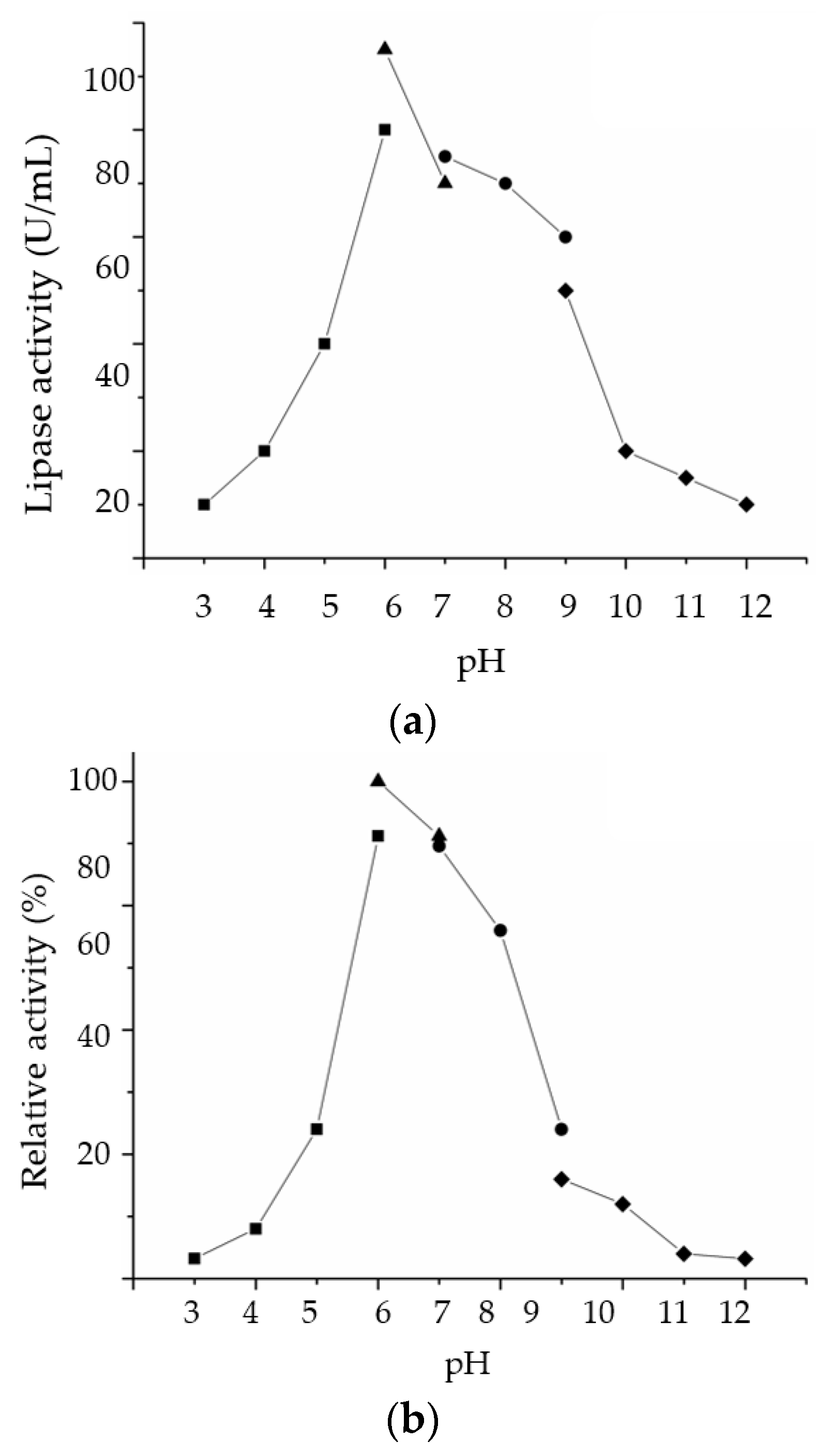
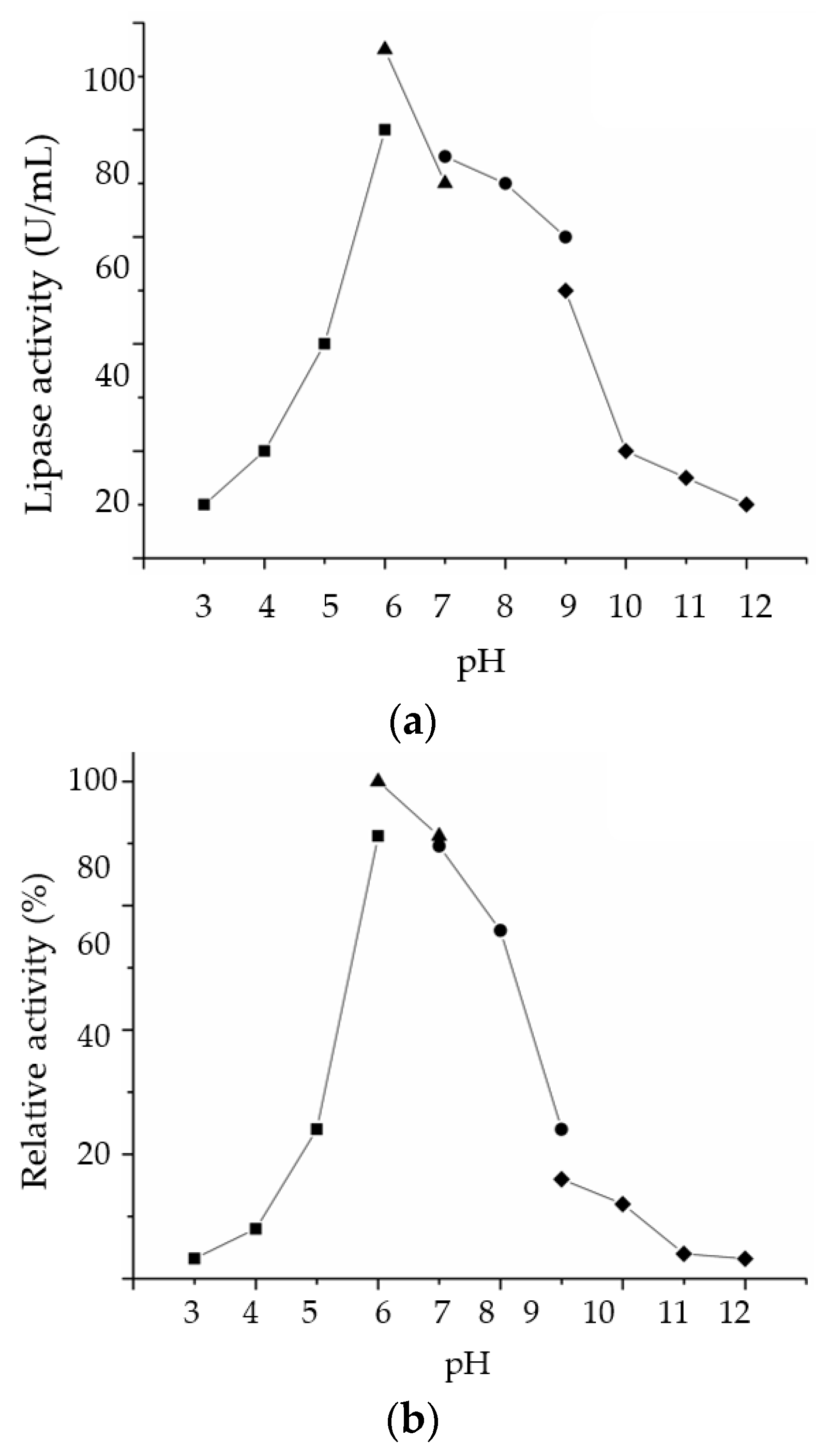
2.3.2. Effect of pH on LSK25 Lipase Activity and Stability
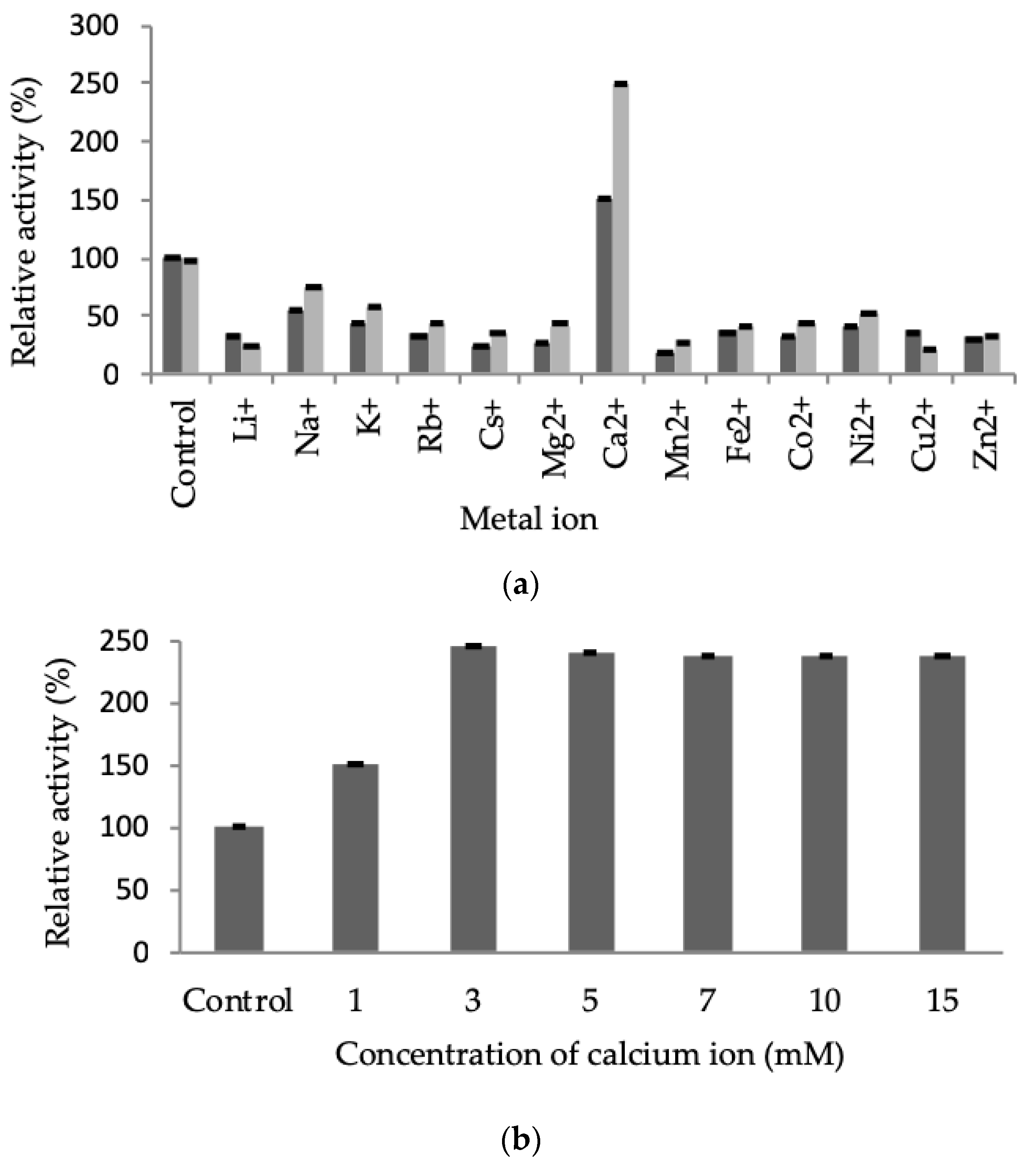
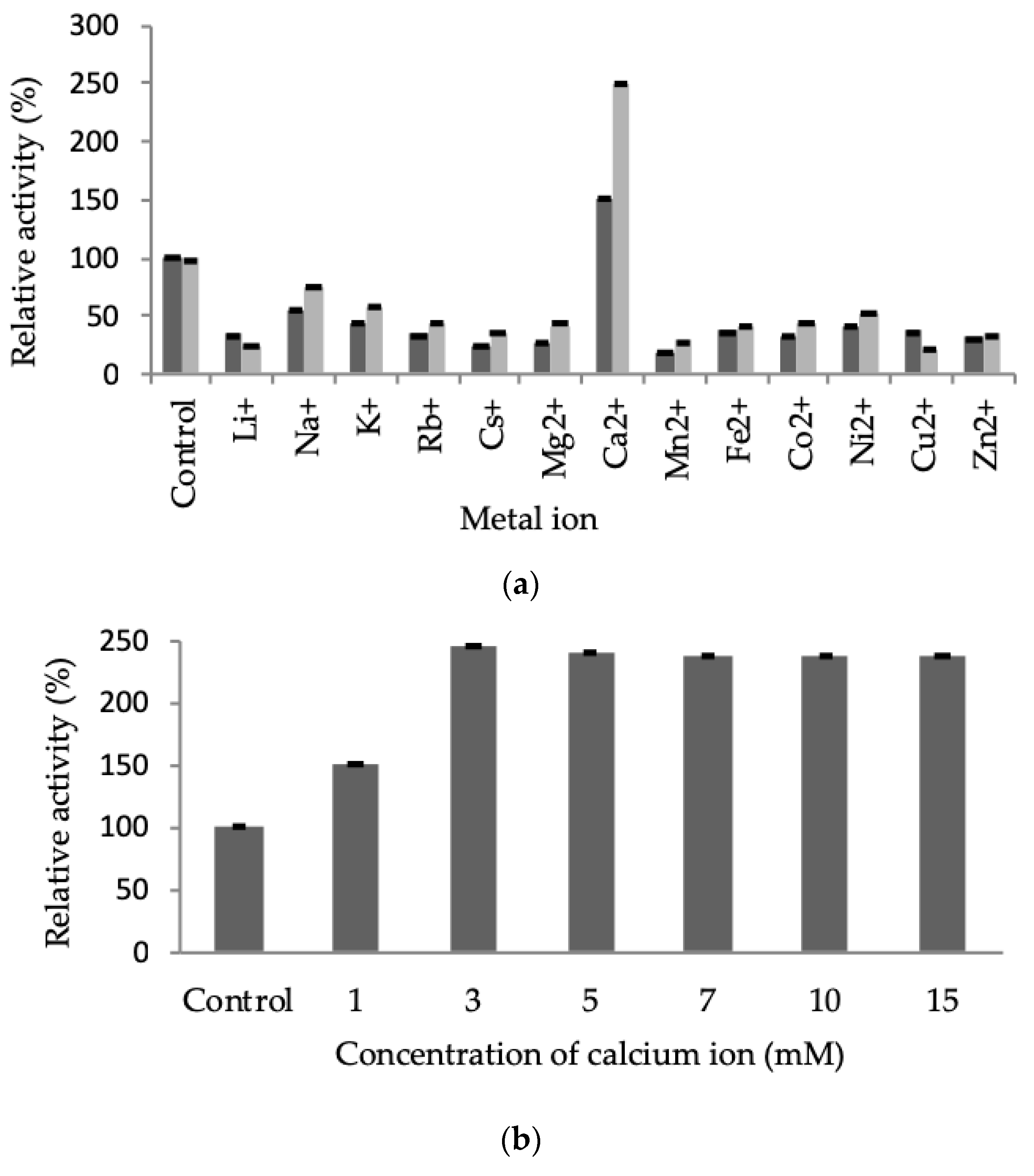
2.3.3. Effect of Metal Ions on LSK25 Lipase Activity
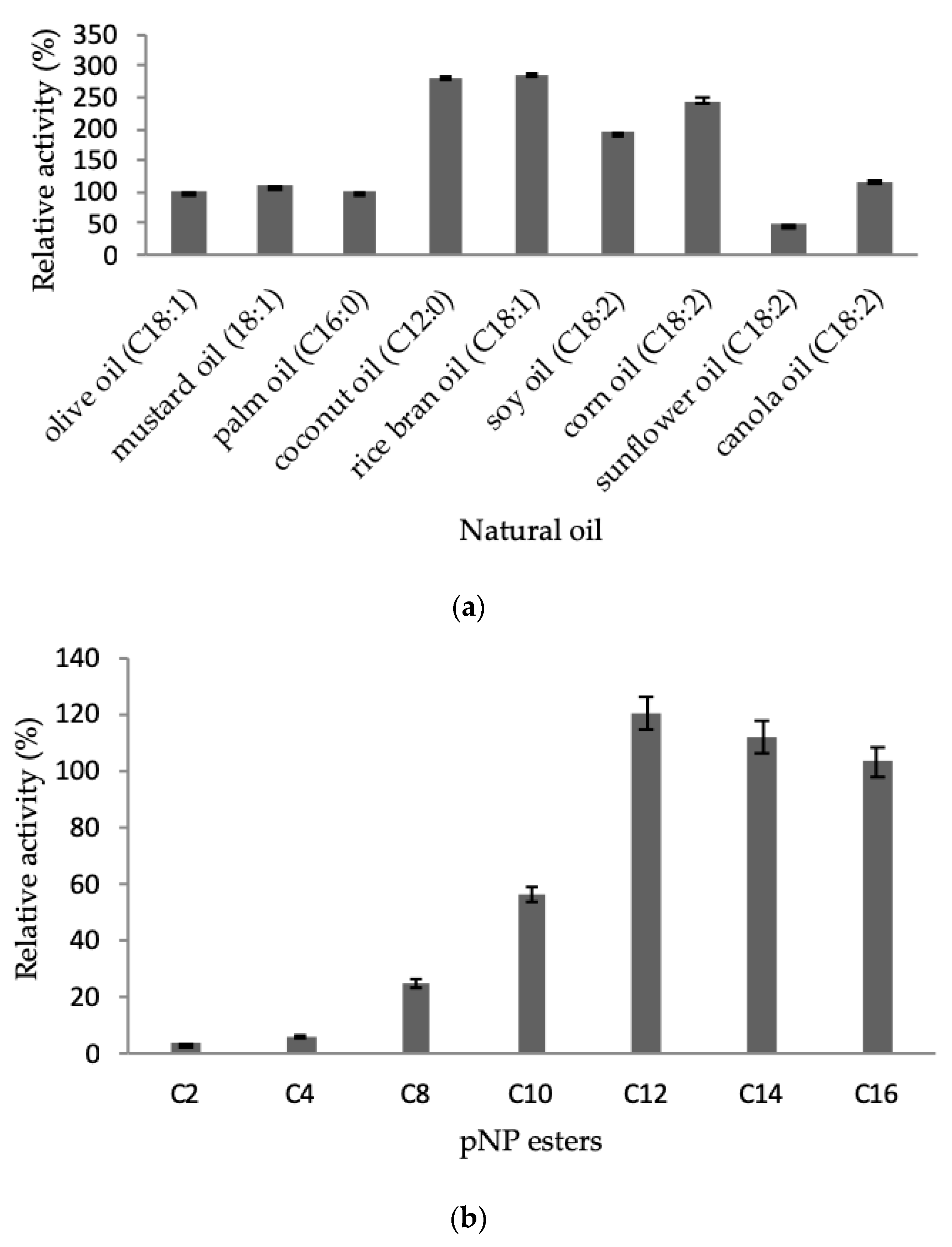
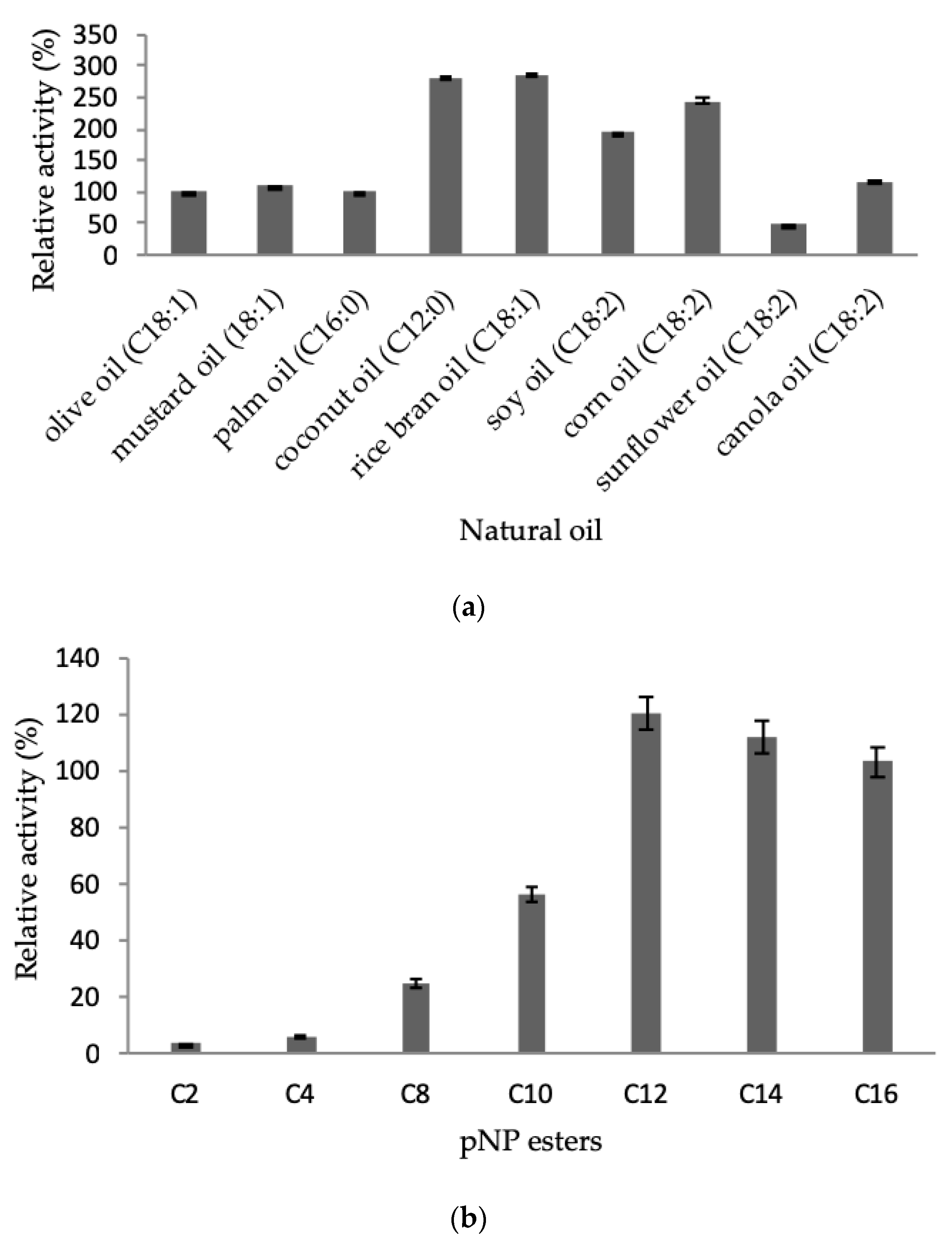
2.3.4. Effect of Natural Oil and Ester Substrates
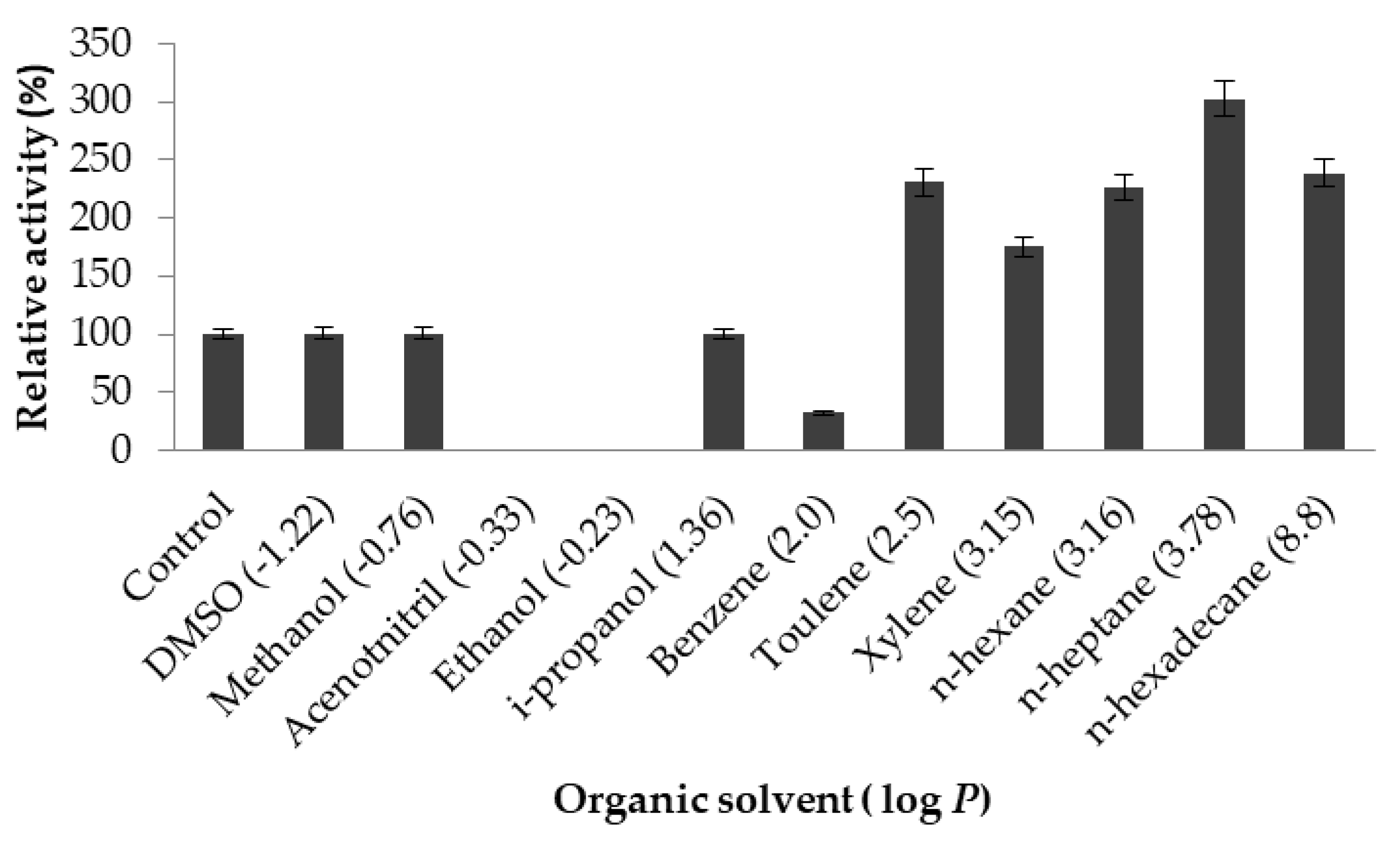
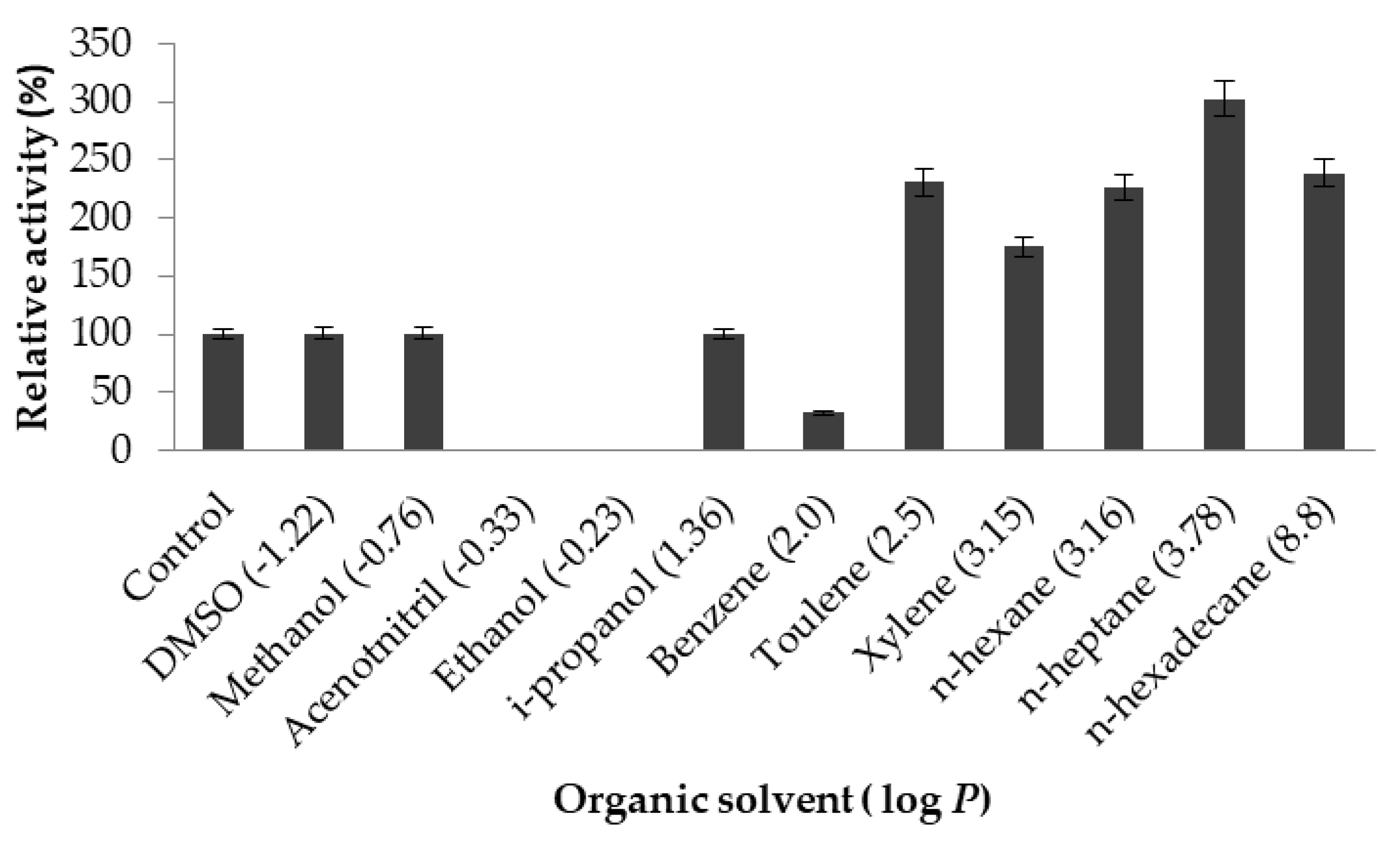
2.3.5. Effect of Organic Solvents on Lipase Activity
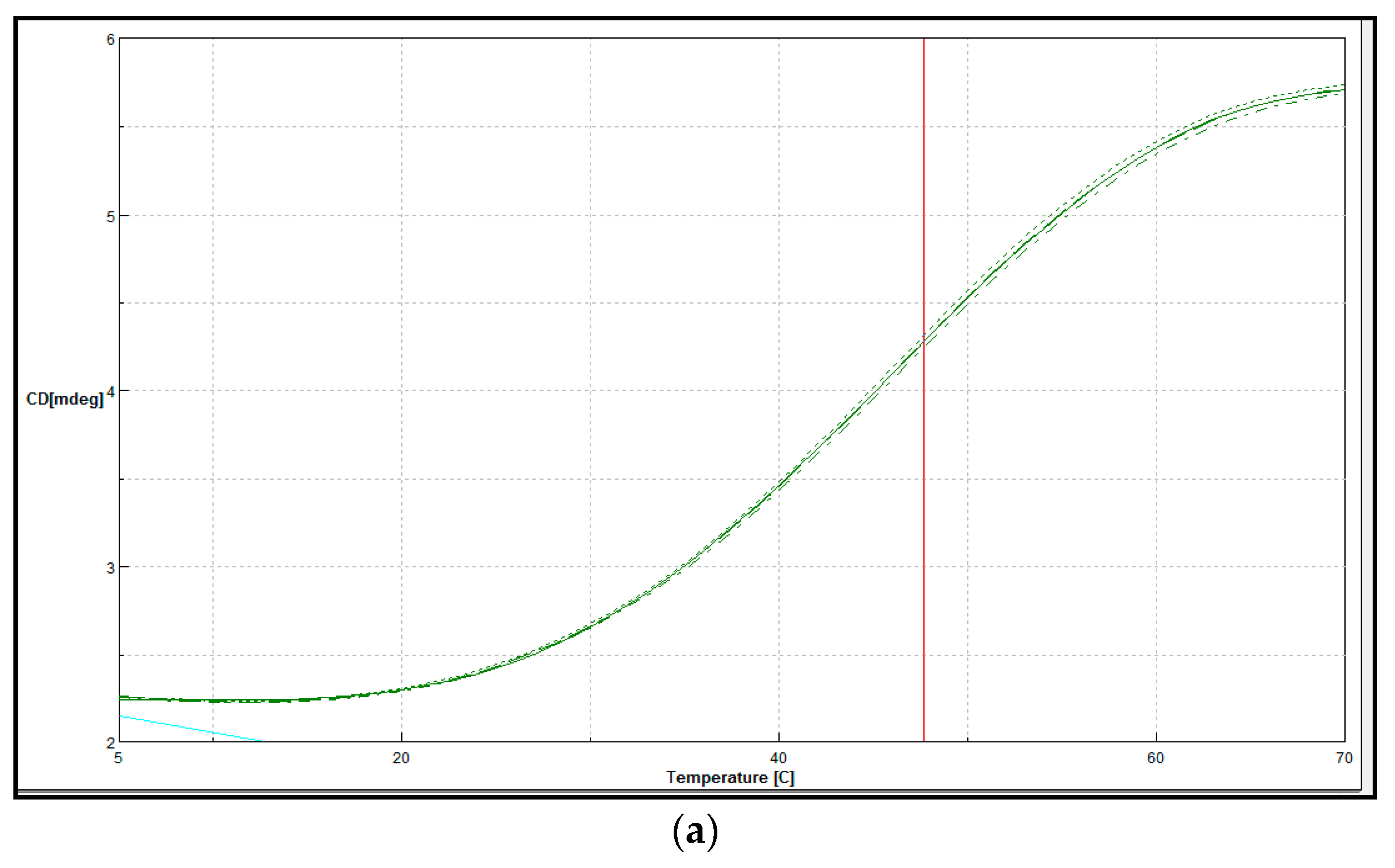
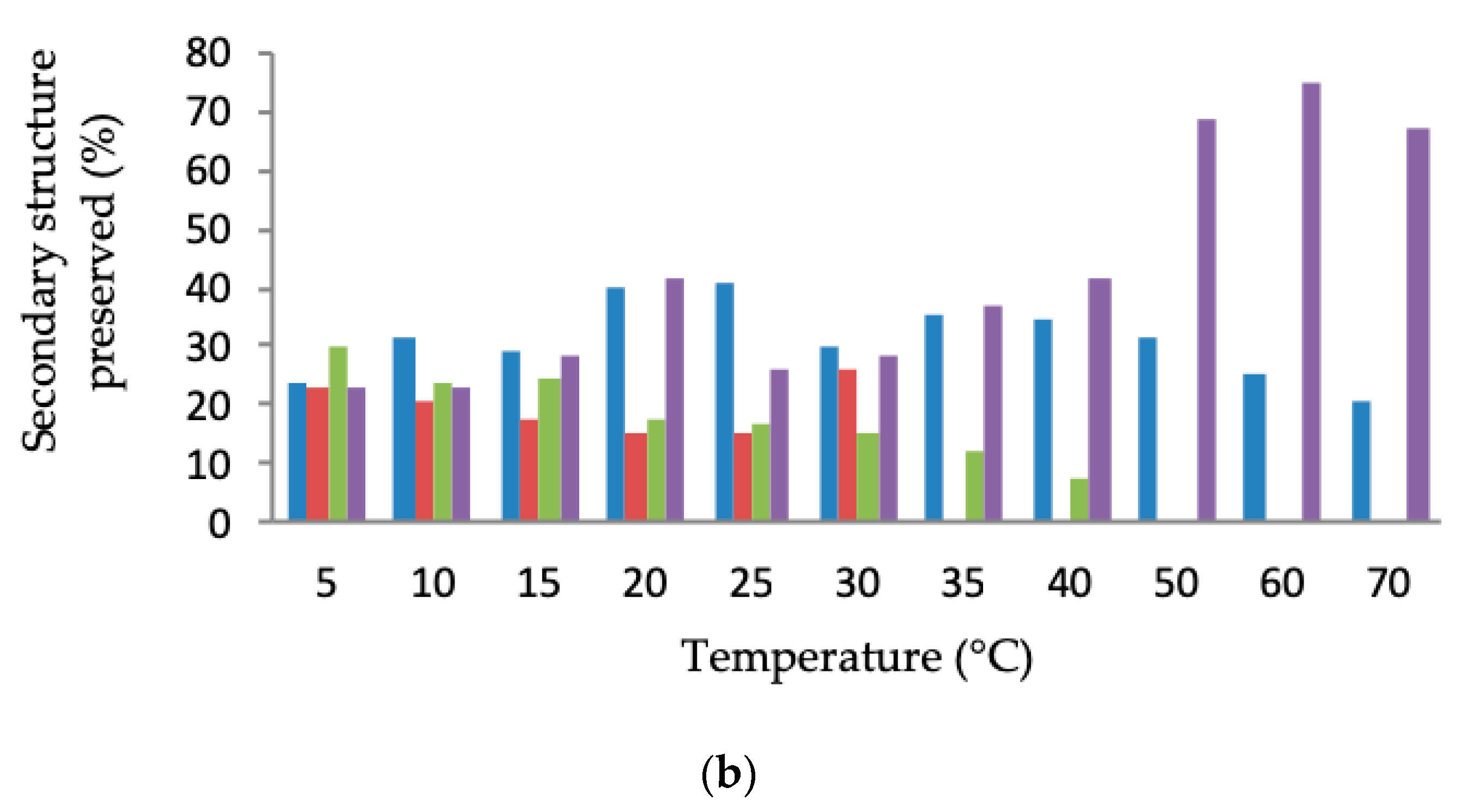
2.4. Biophysical Characterisation of Purified LSK25 Lipase
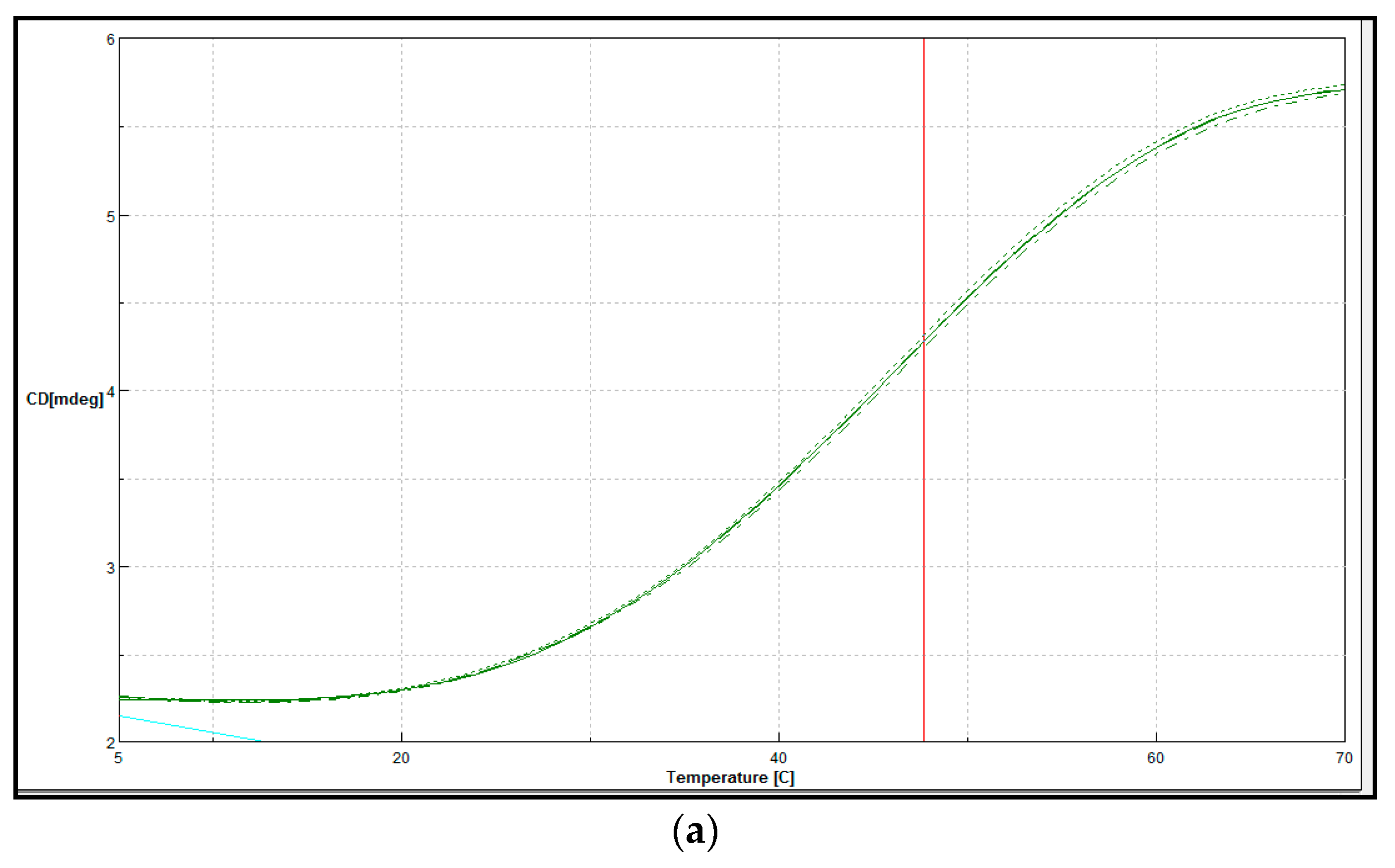
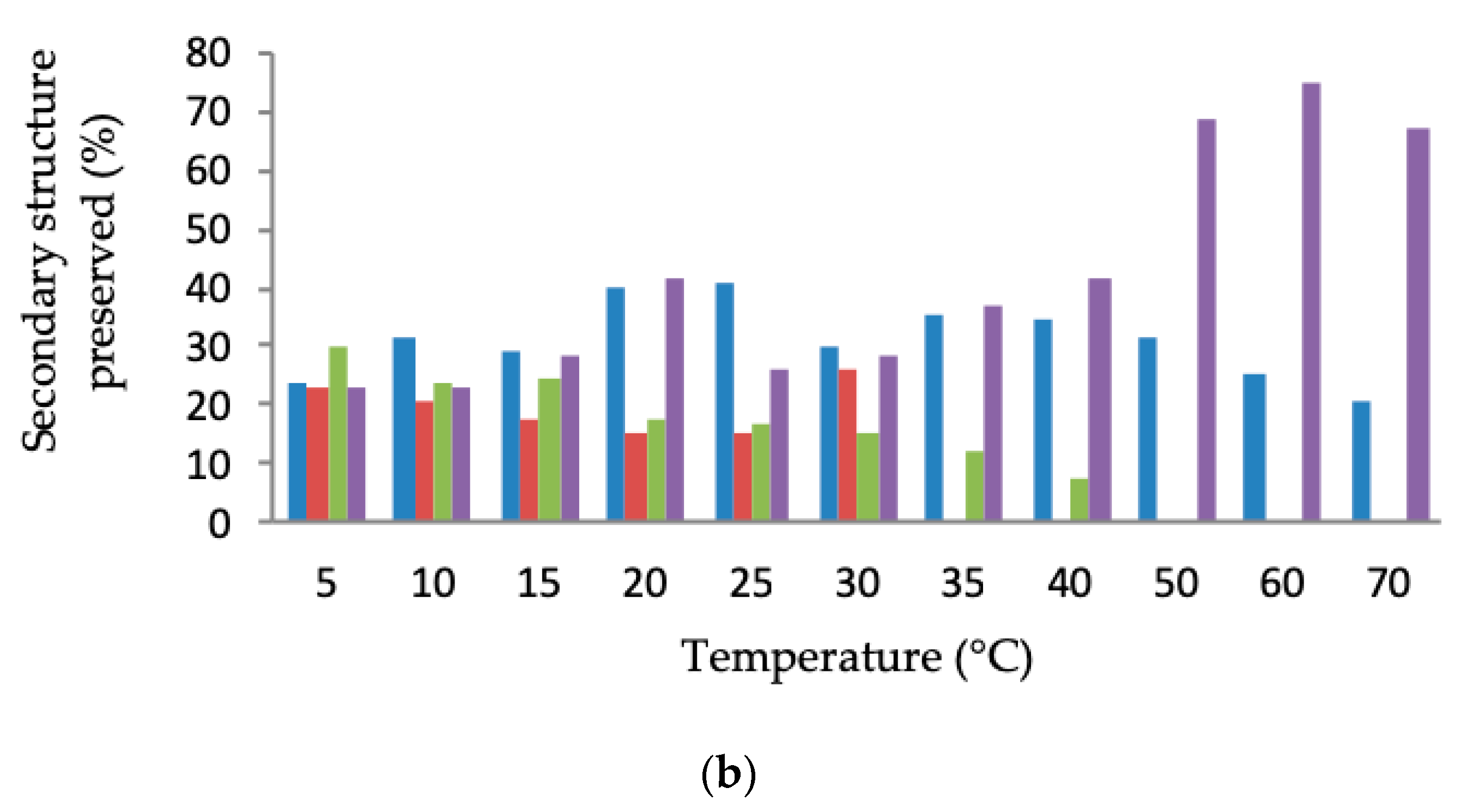
Secondary Structure and Thermal Denaturation
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Plasmids
4.1.1. Qualitative Analyses
4.1.2. Quantitative Analyses
4.2. Recombinant Cold Adapted Lipase Gene
4.3. LSK25 Lipase Sequence Analysis
4.4. Expression of the Lipase Encoding Gene in E. coli Cells
4.5. Solubilisation and Refolding of LSK25 Lipase Inclusion Bodies
4.6. Optimisation of Recombinant Lipase Gene Expression
4.7. Recombinant LSK25 Lipase Purification
4.8. Determination of Protein Content
4.9. Activity Staining
Dynamic Light Scattering (DLS) Analysis of Purified LSK25 Lipase
4.10. Characterization of Purified LSK25 Lipase
4.10.1. Effect of Temperature on Lipase Activity and Stability
4.10.2. Effect of pH on Lipase Activity and Stability
4.10.3. Effect of Metal Ions on Lipase Activity
4.10.4. Effect of Natural Oil and Ester Substrates
4.10.5. Effect of Organic Solvents on Lipase Activity
4.11. Biophysical Characterisation of Purified LSK25 Lipase
Secondary Structure and Thermal Denaturation
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kavitha, M. Cold active lipases—An update. Front. Life Sci. 2016, 9, 226–238. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Siddiqui, K.S. Cold adapted enzymes. In Enzyme Technology; Pandey, A., Webb, C., Soccol, C.R., Larroche, C., Eds.; Asiatech Publishers: New Delhi, India, 2004; pp. 615–638. [Google Scholar]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Joseph, B.; Ramteke, R.W.; Thomas, G. Cold active microbial lipases: Some hot issues and recent developments. Biotechnol. Adv. 2008, 26, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Thakur, S.; Bhatt, A.K. Microbial lipases: Industrial applications and properties. Int. J. Biol. Sci. 2012, 1, 88–92. [Google Scholar]
- Maiangwa, J.; Ali, M.S.M.; Salleh, A.B.; Rahman, R.N.Z.R.A.; Shariff, F.M.; Leow, T.C. Adaptational properties and applications of cold-active lipases from psychrophilic bacteria. Extremophiles 2015, 19, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Salihu, A.; Alam, M.Z. Solvent tolerant lipases: A review. Process Biochem. 2015, 50, 86–96. [Google Scholar] [CrossRef]
- Kamarudin, N.H.A.; Rahman, R.N.Z.R.A.; Ali, M.S.M.; Leow, T.C.; Basri, M.; Salleh, A.B. A new cold-adapted, organic solvent stable lipase from mesophilic Staphylococcus epidermidis AT2. Protein J. 2014, 33, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Gerday, C.; Aittaleb, M.; Arpigny, J.L.; Baise, E.; Chessa, J.P.; Garssoux, G.; Petrescu, I.; Feller, G. Psychrophilic enzymes: A thermodynamic challenge. Biochim. Biophys. Acta. 1997, 1342, 119–131. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Charlton, T.; Ertan, H.; Mohd Omar, S.; Siddiqui, K.S.; Williams, T.J. Biotechnological uses of enzymes from psychrophiles. Microb. Biotechnol. 2011, 4, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Margesin, R.; Gander, S.; Zacke, G.; Gounot, A.M.; Schinner, F. Hydrocarbon degradation and enzyme activities of cold-adapted bacteria and yeasts. Extremophiles 2003, 7, 451–458. [Google Scholar] [CrossRef]
- Thanassi, D.G.; Hultgren, S.J. Multiple pathways allow protein secretion across the bacterial outer membrane. Curr. Opin. Cell Biol. 2000, 12, 420–430. [Google Scholar] [CrossRef]
- Parra, L.P.; Reyes, F.; Acevedo, J.P.; Salazar, O.; Andrews, B.A.; Asenjo, J.A. Cloning and fusion expression of cold-active lipase from marine Antarctic origin. Enzym. Microb. Technol. 2008, 42, 371–377. [Google Scholar] [CrossRef]
- Ganasen, M.; Yaacob, N.; Rahman, R.N.Z.R.A.; Leow, A.T.C.; Basri, M.; Salleh, A.B.; Ali, M.S.M. Cold-adapted organic solvent tolerant alkalophilic family I.3 lipase from an Antarctic Pseudomonas. Int. J. Biol. Macromol. 2016, 92, 1266–1276. [Google Scholar] [CrossRef]
- Latip, W.; Raja Abd Rahman, R.N.Z.; Chor Leow, A.T.; Mohd Shariff, F.; Mohamad Ali, M.S. Expression and characterization of thermotolerant lipase with broad pH profiles isolated from an Antarctic Pseudomonas sp strain AMS3. PeerJ 2016, 4, e2420. [Google Scholar] [CrossRef] [Green Version]
- Angkawidjaja, C.; You, D.J.; Matsumura, H.; Kuwahara, K.; Koga, Y.; Takano, K.; Kanaya, S. Crystal structure of a family I.3 lipase from Pseudomonas sp. MIS38 in a closed conformation. FEBS Lett. 2007, 581, 5060–5064. [Google Scholar] [CrossRef]
- Angkawidjaja, C.; Matsumura, H.; Koga, Y.; Takano, K.; Kanaya, S. X-ray crystallographic and MD simulation studies on the mechanism of interfacial activation of a family I.3 lipase with two lids. J. Mol. Biol. 2010, 400, 82–95. [Google Scholar] [CrossRef]
- Meier, R.; Drepper, T.; Svensson, V.; Jaeger, K.E.; Ulrich, B. A Calcium-gated Lid and a Large β-Roll Sandwich Are Revealed by the Crystal Structure of Extracellular Lipase from Serratia marcescens. J. Biol. Chem. 2007, 282, 31477–31483. [Google Scholar] [CrossRef]
- Ali, M.S.M.; Ganasen, M.; Rahman, R.N.Z.R.A.; Chor, A.L.T.; Salleh, A.B.; Basri, M. Cold-adapted RTX lipase from antarctic Pseudomonas sp. strain AMS8: Isolation, molecular modeling and heterologous expression. Protein J. 2013, 32, 317–325. [Google Scholar] [CrossRef]
- Rashid, N.; Shimada, Y.; Ezaki, S.; Atomi, H. Low-Temperature Lipase from Psychrotrophic Pseudomonas sp. Strain Low-Temperature Lipase from Psychrotrophic Pseudomonas sp. Strain KB700A. Appl. Eenviron. Microbiol. 2001, 67, 4064–4069. [Google Scholar] [CrossRef]
- Borgsthal, G.E. How to use dynamic light scattering to improve the likelihood of growing macromolecular crystals. Methods Mol. Biol. 2007, 363, 109–129. [Google Scholar] [CrossRef]
- Talley, K.; Alexov, E. On the pH-optimum of activity and stability of protein. Proteins 2010, 78, 2699–2706. [Google Scholar] [CrossRef] [PubMed]
- Feller, G. Psychrophilic Enzymes: From Folding to Function and Biotechnology. Scientifica 2013, 2013, 512840. [Google Scholar] [CrossRef] [PubMed]
- Yaacob, N.; Mohamad Ali, M.S.; Salleh, A.B.; Rahman, R.N.Z.R.A.; Leow, A.T.C. Toluene promotes lid 2 interfacial activation of cold active solvent tolerant lipase from Pseudomonas fluorescens strain AMS8. J. Mol. Graph. Model. 2016, 68, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.; Goto, Y.; Réfrégiers, M.; Kardos, J. Accurate secondary structure prediction and fold recognition for circular dichroism spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef] [Green Version]
- Arpigny, J.L.; Lamotte, J.; Gerday, C. Molecular adaptation to cold of an antarctic bacterial lipase. J. Mol. Catal. B Enzym. 1997, 3, 29–35. [Google Scholar] [CrossRef]
- Doukyu, N.; Ogino, H. Organic solvent-tolerant enzymes. Biochem. Eng. J. 2010, 48, 270–282. [Google Scholar] [CrossRef]
- Tanaka, D.; Yoneda, S.; Yamashiro, Y.; Sakatoku, A.; Kayashima, T.; Yamakawa, K.; Nakamura, S. Characterization of a new cold-adapted lipase from Pseudomonas sp. TK-3. Appl. Biochem. Biotechnol. 2012, 168, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Feller, G.; Narinx, E.; Arpigny, J.L.; Aittaleb, M.; Baise, E.; Geniot, S.; Gerday, C. Enzymes from Psychrophilic Organisms. FEMS Microbiol. Rev. 1996, 18, 189–202. [Google Scholar] [CrossRef]
- Feller, G.; Thiry, M.; Arpigny, J.L.; Gerday, C. Cloning and Expression in Escherichia coli of Three Lipase encoding genes from the Psychrotrophic Antarctic strain Moraxella TA144. Gene 1991, 102, 111–115. [Google Scholar] [CrossRef]
- Panicker, G.; Aislabie, J.; Saul, D.; Bej, A. Cold tolerance of Pseudomonas sp. 30–3 isolated from oil-contaminated soil, Antarctica. Polar Biol. 2002, 25, 5–11. [Google Scholar] [CrossRef]
- Song, J.M.; Nam, K.-W.; Kang, S.G.; Kim, C.-G.; Kwon, S.-T.; Lee, Y.-H. Molecular cloning and characterization of a novel cold-active beta-1,4-D-mannanase from the Antarctic springtail, Cryptopygus antarcticus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 32–40. [Google Scholar] [CrossRef]
- Cieslinski, H.; Kur, J.; Bialkowska, A.; Baran, I.; Makowski, K.; Turkiewicz, M. Cloning, expression, and purification of a recombinant cold-adapted beta-galactosidase from Antarctic bacterium Pseudoalteromonas sp. 22b. Protein Expr. Purif. 2005, 39, 27–34. [Google Scholar] [CrossRef]
- Leow, T.C.; Rahman, R.N.Z.R.A.; Basri, M.; Salleh, A.B. A thermoalkaliphilic lipase of Geobacillus sp. T1. Extremophiles 2007, 11, 527–535. [Google Scholar] [CrossRef]
- Singh, S.M.; Panda, A.K. Solubilization and refolding of bacterial inclusion body proteins. J. Biosci. Bioeng. 2005, 99, 303–310. [Google Scholar] [CrossRef]
- Tsumoto, K.; Ejima, D.; Kumagai, I.; Arakawa, T. Practical considerations in refolding proteins from inclusion bodies. Protein Expr. Purif. 2003, 28, 1–8. [Google Scholar] [CrossRef]
- Park, I.H.; Kim, S.H.; Lee, Y.S.; Lee, S.C.; Zhou, Y.; Kim, C.M.; Ahn, S.C.; Choi, Y.L. Gene cloning, purification, and characterization of a cold-adapted lipase produced by Acinetobacter baumannii BD5. J. Microbiol. Biotechnol. 2009, 19, 128–135. [Google Scholar] [CrossRef]
- Novototskaya-Vlasova, K.; Petrovskaya, L.; Kryukova, E.; Rivkina, E.; Dolgikh, D.; Kirpichnikov, M. Expression and chaperone-assisted refolding of a new cold-active lipase from Psychrobacter cryohalolentis K5T. Protein Expr. Purif. 2013, 91, 96–103. [Google Scholar] [CrossRef]
- Gerday, C.; Aittaleb, M.; Bentahir, M.; Chessa, J.P.; Claverie, P.; Collins, T.; D’Amico, S.; Dumont, J.; Garsoux, G.; Georlette, D.; et al. Cold-adapted enzymes: From fundamentals to biotechnology. Trends Biotechnol. 2000, 18, 103–107. [Google Scholar] [CrossRef]
- Kumar, D.; Kumar, L.; Nagar, S.; Raina, C.; Parshad, R.; Gupta, V.K. Screening, isolation and production of lipase/esterase producing Bacillus sp. strain DVL2 and its potential evaluation in esterification and resolution reactions. Arch. Appl. Sci. Res. 2012, 4, 1763–1770. [Google Scholar]
- Oliveira, C.; Domingues, L. Guidelines to reach high-quality purified recombinant proteins. Appl. Microbiol. Biotechnol. 2018, 102, 81–92. [Google Scholar] [CrossRef]
- Geoffry, K.; Achur, R.N. Screening and production of lipase from fungal organisms. Biocatal. Agric. Biotechnol. 2018, 14, 241–253. [Google Scholar] [CrossRef]
- Wu, G.; Zhang, X.; Wei, L.; Wu, G.; Kumar, A.; Mao, T.; Liu, Z. A cold-adapted, solvent and salt tolerant esterase from marine bacterium Psychrobacter pacificensis. Int. J. Biol. Macromol. 2015, 81, 180–187. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, H.; Wu, Y.; Zeng, J.; Guo, Z.; Wang, L.; Shen, C.; Qiao, D.; Cao, Y. A new cold-adapted, alkali-stable and highly salt-tolerant esterase from Bacillus licheniformis. Int. J. Biol. Macromol. 2018, 111, 1183–1193. [Google Scholar] [CrossRef]
- Kuddus, M. Cold-active enzymes in food biotechnology: An updated mini review. J. Appl. Biol. Biotechnol. 2018, 6, 58–63. [Google Scholar] [CrossRef]
- Alquati, C.; De Gioia, L.; Santarossa, G.; Alberghina, L.; Fantucci, P.; Lotti, M. The cold-active lipase of Pseudomonas fragi. Heterologous expression, biochemical characterization and molecular modeling. Eur. J. Biochem. 2002, 269, 3321–3328. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, S.; Zeng, R. Cloning, expression, and characterization of a cold-adapted lipase gene from an Antarctic deep-sea psychrotrophic bacterium, Psychrobacter sp. 7195. J. Microbiol. Biotechnol. 2007, 17, 604–610. [Google Scholar]
- Tian, J.; Lei, Z.; Qiu, P.; Wang, L.; Tian, Y. Purification and characterization of a cold-adapted lipase from Oceanobacillus strain PT-11. PLoS ONE 2014, 9, e101343. [Google Scholar] [CrossRef]
- Cheng, Y.Y.; Qian, Y.K.; Li, Z.F.; Wu, Z.H.; Liu, H.; Li, Y.Z. A novel cold-adapted lipase from Sorangium cellulosum strain So0157-2: Gene cloning, expression and enzymatic characterization. Int. J. Mol. Sci. 2011, 12, 6765–6780. [Google Scholar] [CrossRef]
- Zhang, J.W.; Zeng, R.Y. Molecular cloning and expression of a cold-adapted lipase gene from an Antarctic deep sea psychrotrophic bacterium Pseudomonas sp. 7323. Mar. Biotechnol. 2008, 10, 612–621. [Google Scholar] [CrossRef]
- Chen, R.; Guo, L.; Dang, H. Gene cloning, expression, and characterization of a cold-adapted lipase from a psychrophilic deep-sea bacterium Psychrobacter sp. C18. World J. Microb. Biot. 2011, 27, 431–441. [Google Scholar] [CrossRef]
- Priyanka, P.; Kinsella, G.; Henehan, G.T.; Ryan, B.J. Isolation, purification and characterization of a novel solvent stable lipase from Pseudomonas reinekei. Protein Expr. Purif. 2019, 153, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Chu, X.; Zhang, W.; Wu, N.; Fan, Y. A novel cold-adapted lipase from Acinobacter sp. XMZ-26: Gene cloning and characterization. Appl. Microbiol. Biotechnol. 2011, 90, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Yadav, K.N.S.; Adsul, M.G.; Bastawde, K.B.; Jadhav, D.D.; Thulasiram, H.V.; Gokhale, D.V. Differential induction, purification and characterization of cold active lipase from Yarrowia lipolytica NCIM 3639. Bioresour. Technol. 2011, 102, 10663–10670. [Google Scholar] [CrossRef]
- Ramani, K.; Chockalingam, E.; Sekaran, G. Production of a novel extracellular acidic lipase from Pseudomonas gessardii using slaughterhouse waste as a substrate. J. Ind. Microbiol. Biotechnol. 2010, 37, 531–535. [Google Scholar] [CrossRef]
- Almeida, A.F.; Tauk-Tornisielo, S.; Carmona, E.C. Acid lipase from Candida viswanathii: Production, biochemical properties, and potential application. BioMed Res. Int. 2013, 2013, 435818. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Zhang, W.H.; Wang, W.D.; Dai, Y.J.; Zhang, H.T.; Wang, Y.; Wang, H.K.; Lu, F.P. A high-detergent-performance, cold-adapted lipase from Pseudomonas stutzeri PS59 suitable for detergent formulation. J. Mol. Catal. B Enzym. 2014, 102, 16–24. [Google Scholar] [CrossRef]
- Litantra, R.; Lobionda, S.; Yim, J.H.; Kim, H.K. Expression and biochemical characterization of cold-adapted lipases from Antarctic Bacillus pumilus strains. J. Microbiol. Biotechnol. 2013, 23, 1221–1228. [Google Scholar] [CrossRef]
- Sharma, R.; Chisti, Y.; Banerjee, U.C. Production, purification, characterization, and applications of lipases. Biotechnol. Adv. 2001, 19, 627–662. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.A.V.; Le, T.D.; Phan, H.N.; Tran, L.B. Hydrolysis Activity of Virgin Coconut Oil Using Lipase from Different Sources. Scientifica 2018, 2018, 9120942. [Google Scholar] [CrossRef]
- Rahman, R.N.Z.R.A.; Baharum, S.N.; Basri, M.; Salleh, A.B. High-yield purification of an organic solvent-tolerant lipase from Pseudomonas sp. strain S5. Anal. Biochem. 2005, 341, 267–274. [Google Scholar] [CrossRef]
- Cao, Y.; Zhuang, Y.; Yao, C.; Wu, B.; He, B. Purification and characterization of an organic solvent-stable lipase from Pseudomonas stutzeri LC2-8 and its application for efficient resolution of (R, S)-1-phenylethanol. Biochem. Eng. J. 2012, 64, 55–60. [Google Scholar] [CrossRef]
- Gurung, N.; Ray, S.; Bose, S.; Rai, V. A broader view: Microbial enzymes and their relevance in industries, medicine, and beyond. BioMed Res. Int. 2013, 2013, 329121. [Google Scholar] [CrossRef] [PubMed]
- Parwata, I.P.; Asyari, M.; Hertadi, R. Organic solvent-stable lipase from moderate halophilic bacteria Pseudomonas stutzeri isolated from the mud crater of bleduk kuwu, central java, Indonesia. J. Pure Appl. Microbiol. 2014, 8, 31–40. [Google Scholar]
- Javed, S.; Azeem, F.; Hussain, S.; Rasul, I.; Siddique, M.H.; Riaz, M.; Afzal, M.; Kouser, A.; Nadeem, H. Bacterial lipases: A review on purification and characterization. Prog. Biophys. Mol. Biol. 2018, 132, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Mander, P.; Cho, S.S.; Simkhada, J.R.; Choi, Y.H.; Park, D.J.; Ha, J.W.; Yoo, J.C. An organic solvent-tolerant alkaline lipase from Streptomyces sp. CS268 and its application in biodiesel production. Biotechnol. Bioprocess Eng. 2012, 17, 67–75. [Google Scholar] [CrossRef]
- Bose, A.; Keharia, H. Production, characterization and applications of organic solvent tolerant lipase by Pseudomonas aeruginosa AAU2. Biocatal. Agric. Biotechnol. 2013, 2, 255–266. [Google Scholar] [CrossRef]
- Jung, J.W.; Lee, W. Structure-based functional discovery of proteins: Structural proteomics. J. Biochem. Mol. Biol. 2004, 37, 28–34. [Google Scholar] [CrossRef]
- Wang, Y. The Lid Domain in Lipases: Structural and Functional Determinant of enzymatic Properties. Bioeng. Biotechnol. 2017, 5, 16. [Google Scholar] [CrossRef]
- Raussens, V.; Ruysschaert, J.M.; Goormaghtigh, E. Protein concentration is not an absolute prerequisite for the determination of secondary structure from circular dichroism spectra: A new scaling method. Anal. Biochem. 2003, 319, 114–121. [Google Scholar] [CrossRef]
- Salwoom, L.; Raja Abd Rahman, R.N.Z.; Salleh, A.B.; Mohd Shariff, F.; Convey, P.; Pearce, D.; Mohamad Ali, M.S. Isolation, Characterisation, and Lipase Production of a Cold-Adapted Bacterial Strain Pseudomonas sp. LSK25 Isolated from Signy Island, Antarctica. Molecules 2019, 24, 715. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Rhee, J.S. A simple and rapid colorimetric method for determination of free fatty acids for lipase assay. J. Am. Oil Chem. Soc. 1986, 63, 89–92. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microb. 2014, 5, 172. [Google Scholar] [CrossRef]
- Laemmli, U.K. Most commonly used discontinuous buffer system for SDS electrophoresis. Nature 1970, 227, 680–686. [Google Scholar] [CrossRef]
- Arndt, C.; Koristha, S.; Bartsch, H.; Bachmann, M. Native Polyacrylamide Gels. Protein Electrophoresis 2012, 869, 49–53. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sommer, P.; Bormann, C.; Götz, F. Genetic and biochemical characterization of a new extracellular lipase from Streptomyces cinnamomeus. Appl. Environ. Microbiol. 1997, 63, 3553–3560. [Google Scholar]
- Frisken, B.J. Revisiting the Method of Cumulants for the Analysis of Dynamic Light-Scattering Data. Appl. Opt. 2001, 40, 4087–4091. [Google Scholar] [CrossRef]
- Comer, J.; Tam, K. Lipophilicity Profiles: Theory and Measurement. In Pharmacokinetic Optimization in Drug Research: Biological, Physicochemical, and Computational Strategies; Wiley-VCH: Weinheim, Germany, 2001; pp. 275–304. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salwoom, L.; Raja Abd. Rahman, R.N.Z.; Salleh, A.B.; Mohd. Shariff, F.; Convey, P.; Mohamad Ali, M.S. New Recombinant Cold-Adapted and Organic Solvent Tolerant Lipase from Psychrophilic Pseudomonas sp. LSK25, Isolated from Signy Island Antarctica. Int. J. Mol. Sci. 2019, 20, 1264. https://doi.org/10.3390/ijms20061264
Salwoom L, Raja Abd. Rahman RNZ, Salleh AB, Mohd. Shariff F, Convey P, Mohamad Ali MS. New Recombinant Cold-Adapted and Organic Solvent Tolerant Lipase from Psychrophilic Pseudomonas sp. LSK25, Isolated from Signy Island Antarctica. International Journal of Molecular Sciences. 2019; 20(6):1264. https://doi.org/10.3390/ijms20061264
Chicago/Turabian StyleSalwoom, Leelatulasi, Raja Noor Zaliha Raja Abd. Rahman, Abu Bakar Salleh, Fairolniza Mohd. Shariff, Peter Convey, and Mohd Shukuri Mohamad Ali. 2019. "New Recombinant Cold-Adapted and Organic Solvent Tolerant Lipase from Psychrophilic Pseudomonas sp. LSK25, Isolated from Signy Island Antarctica" International Journal of Molecular Sciences 20, no. 6: 1264. https://doi.org/10.3390/ijms20061264
APA StyleSalwoom, L., Raja Abd. Rahman, R. N. Z., Salleh, A. B., Mohd. Shariff, F., Convey, P., & Mohamad Ali, M. S. (2019). New Recombinant Cold-Adapted and Organic Solvent Tolerant Lipase from Psychrophilic Pseudomonas sp. LSK25, Isolated from Signy Island Antarctica. International Journal of Molecular Sciences, 20(6), 1264. https://doi.org/10.3390/ijms20061264