Pharmacokinetic and Metabolomic Studies with BIO 300, a Nanosuspension of Genistein, in a Nonhuman Primate Model



Abstract
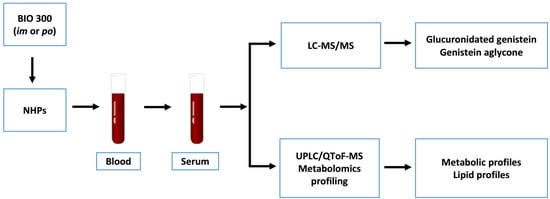
:
1. Introduction
2. Results
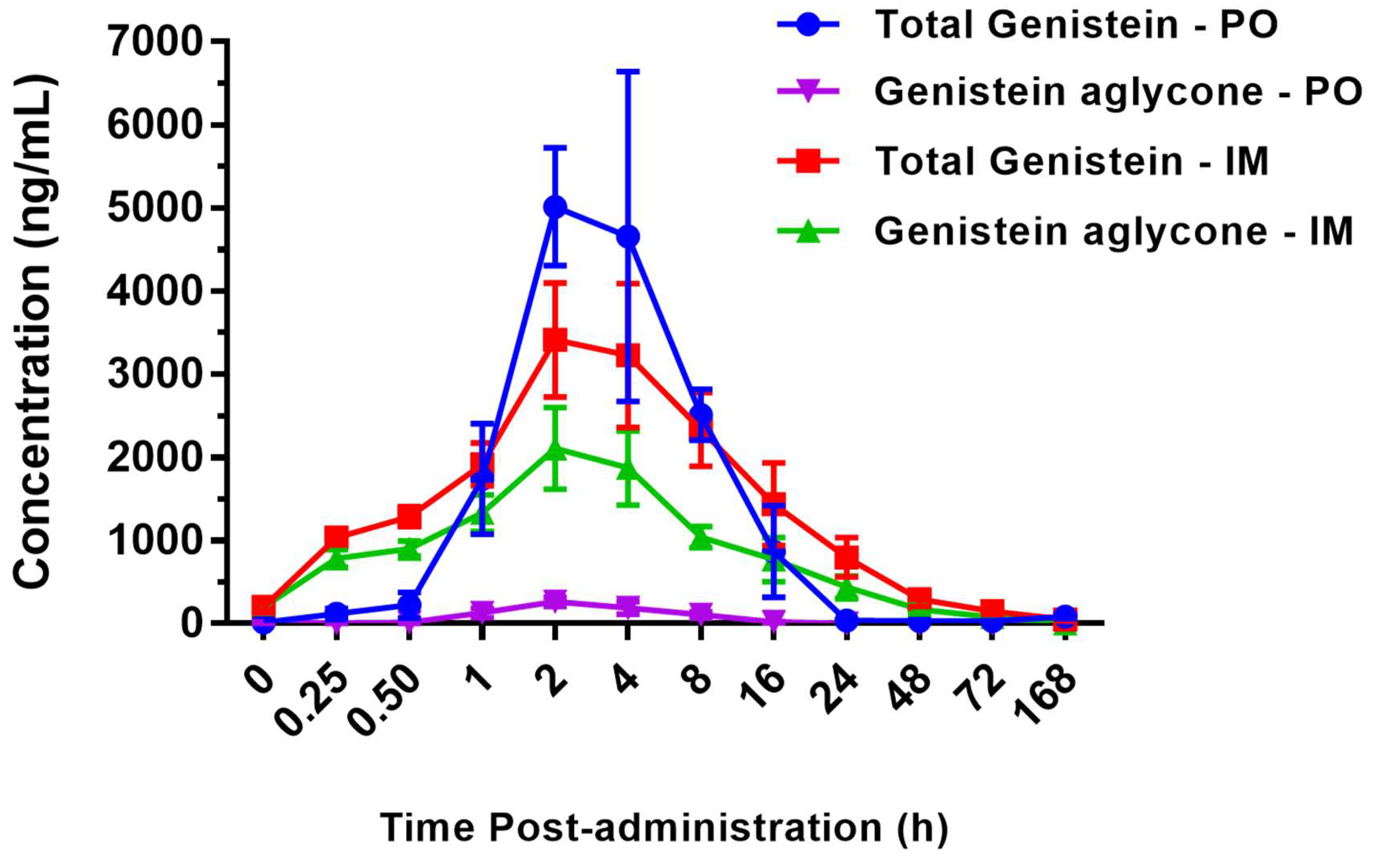
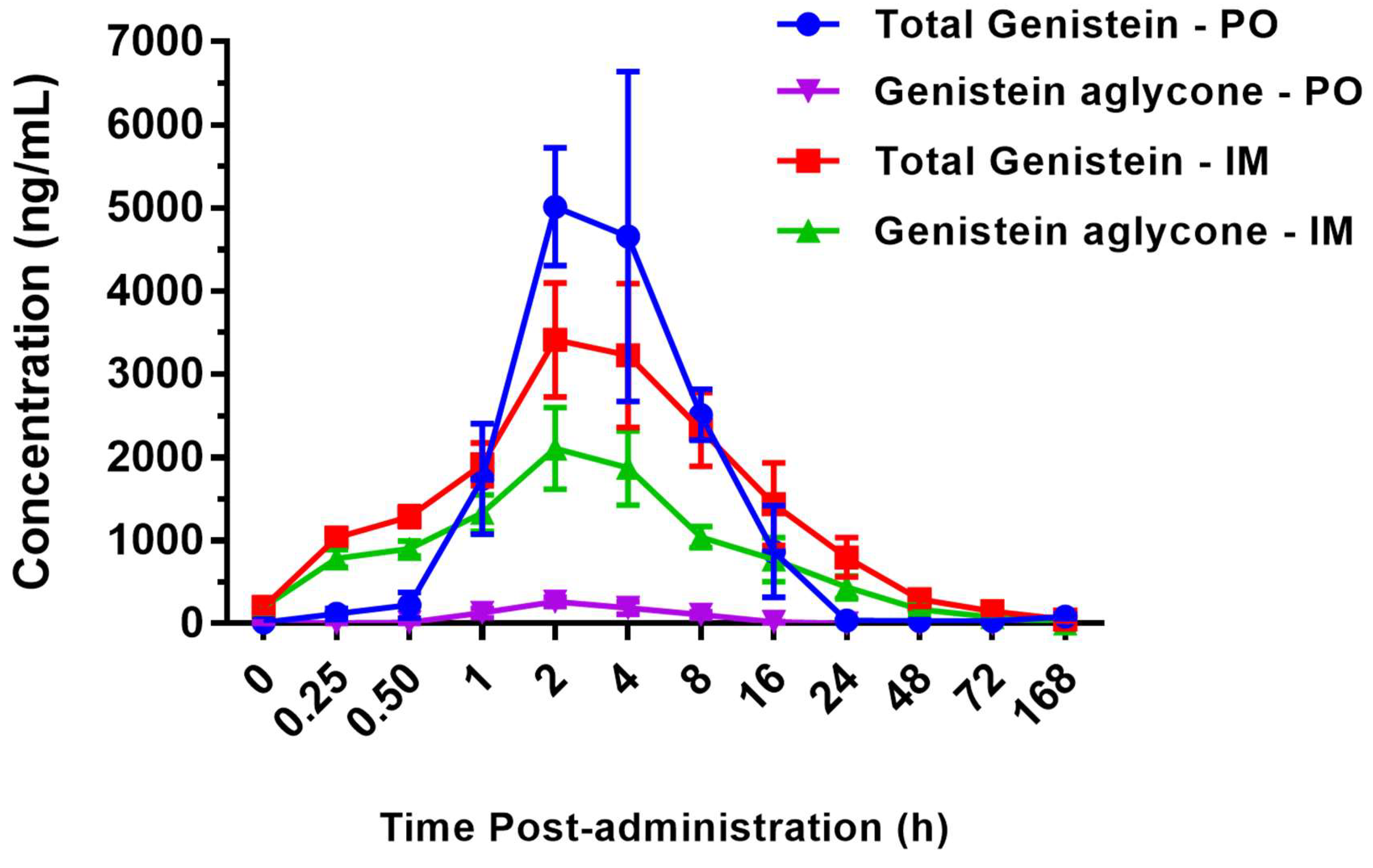
2.1. Pharmacokinetics of BIO 300 Administered po or im
2.2. Analysis for CBC and Vital Signs in NHP Administered BIO 300
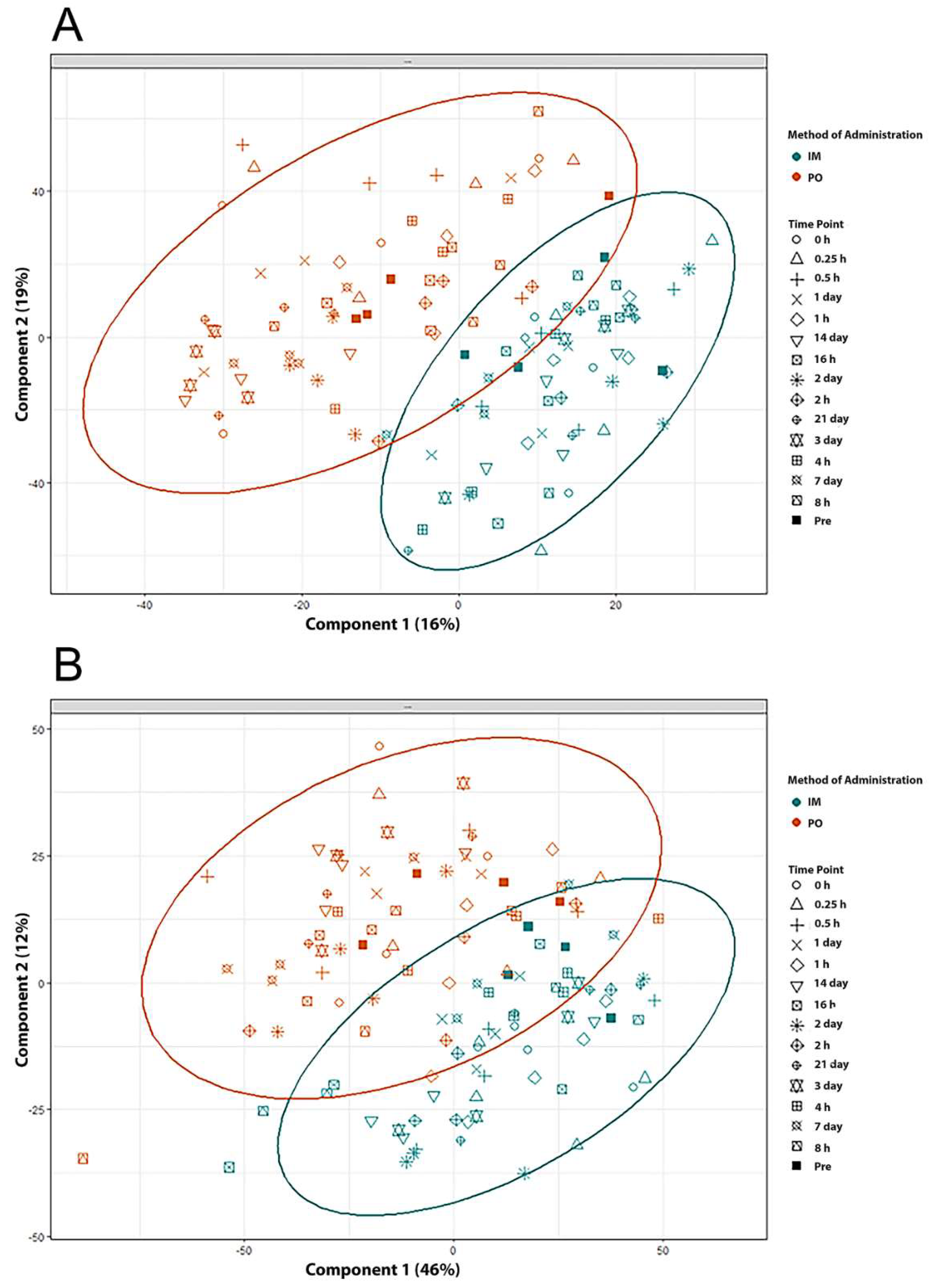
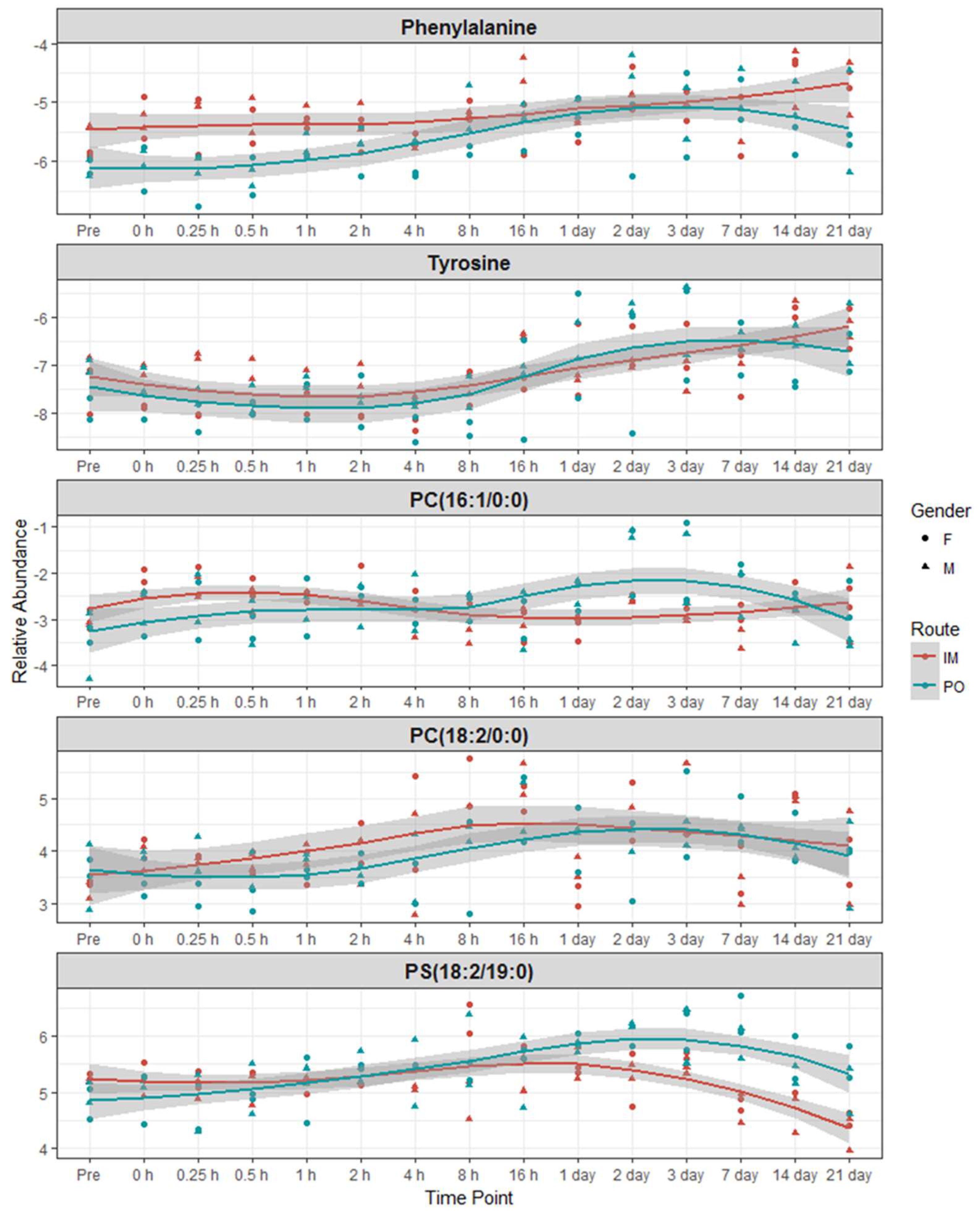
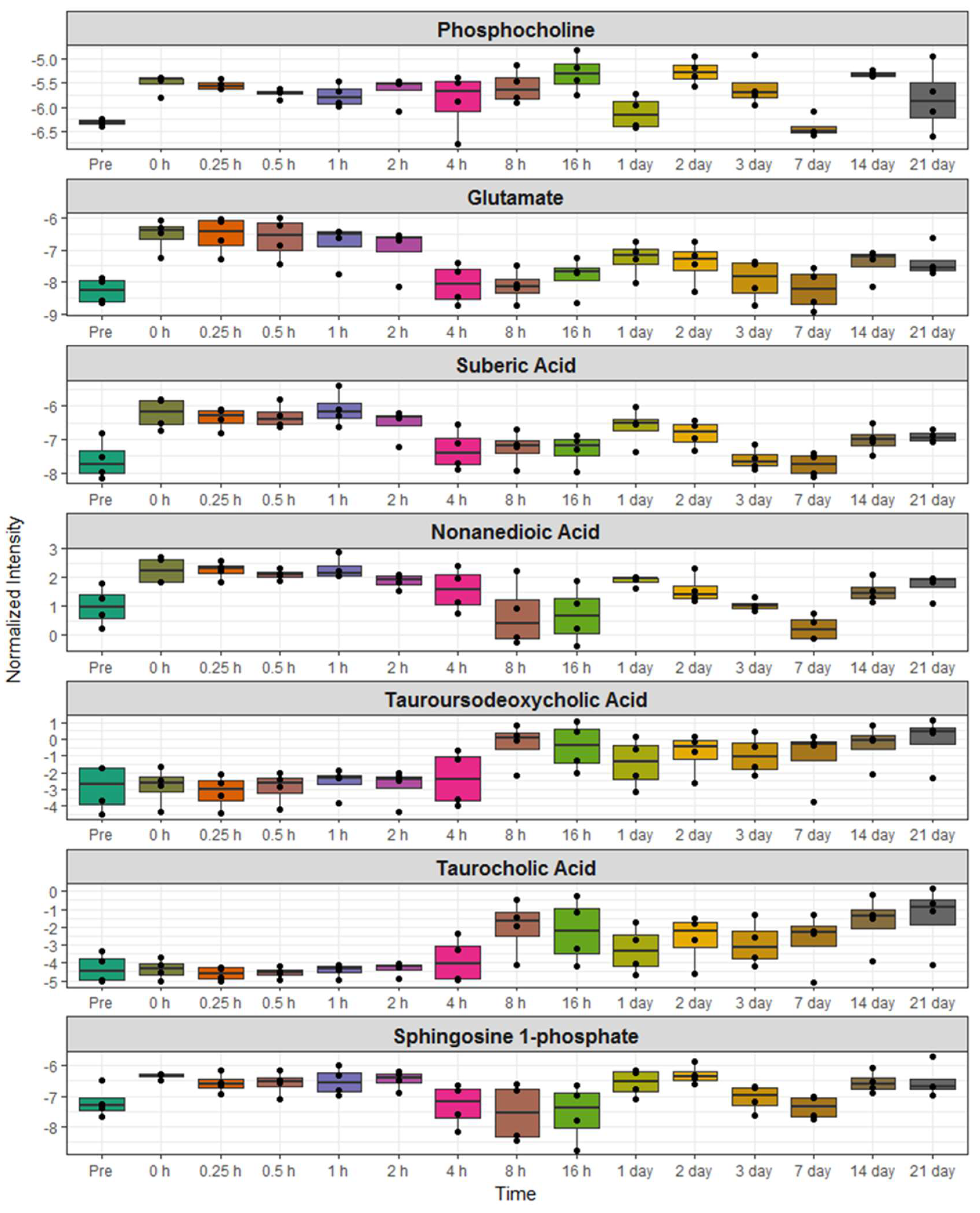
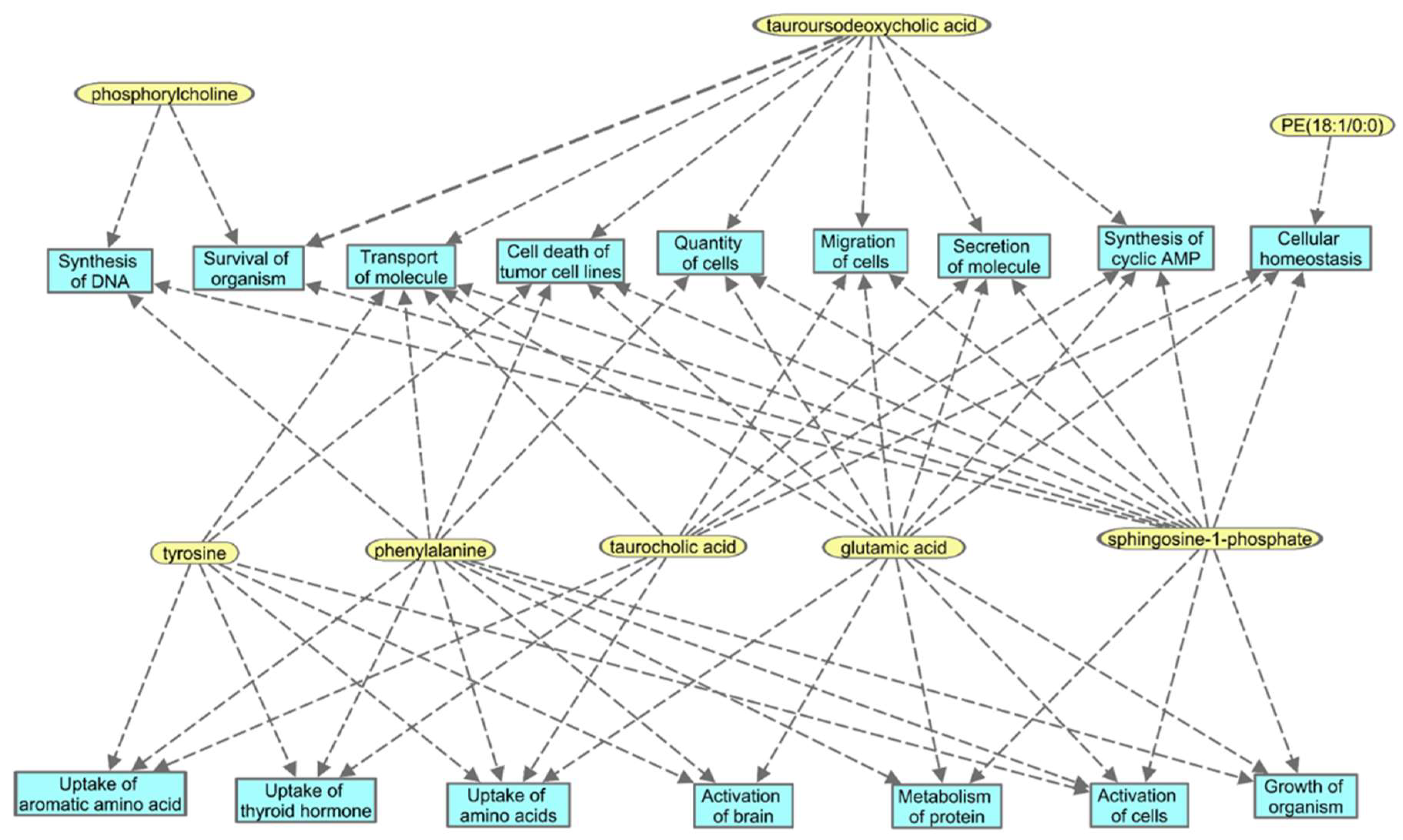
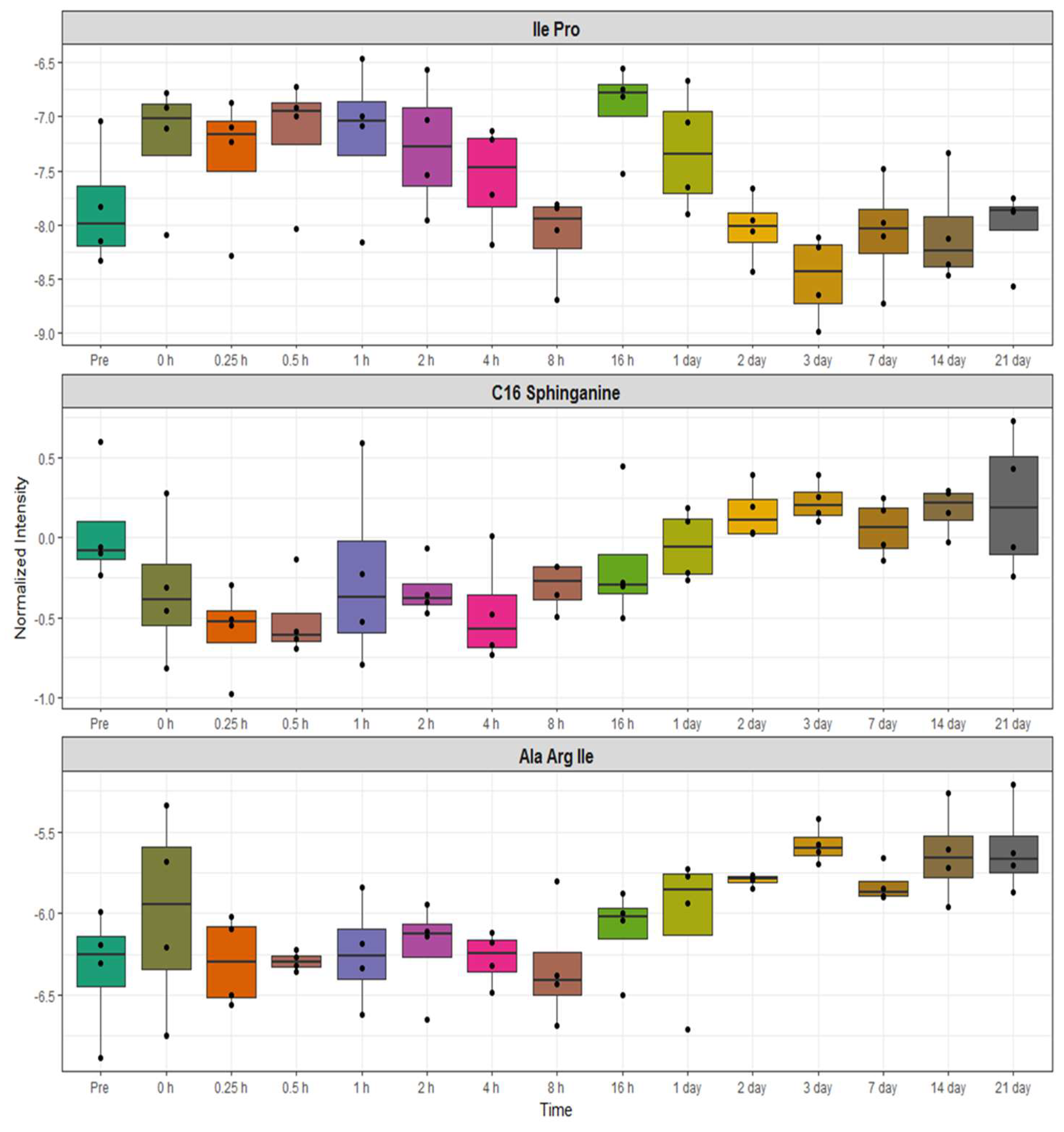
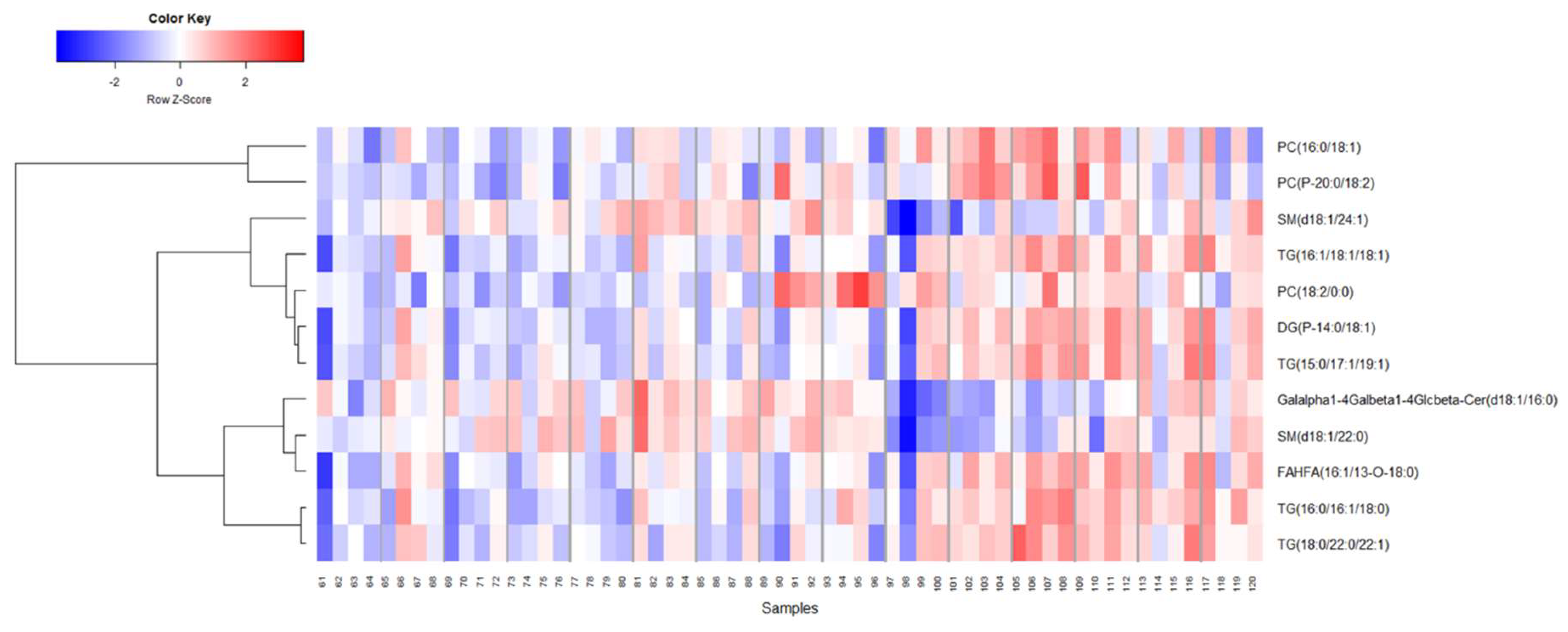
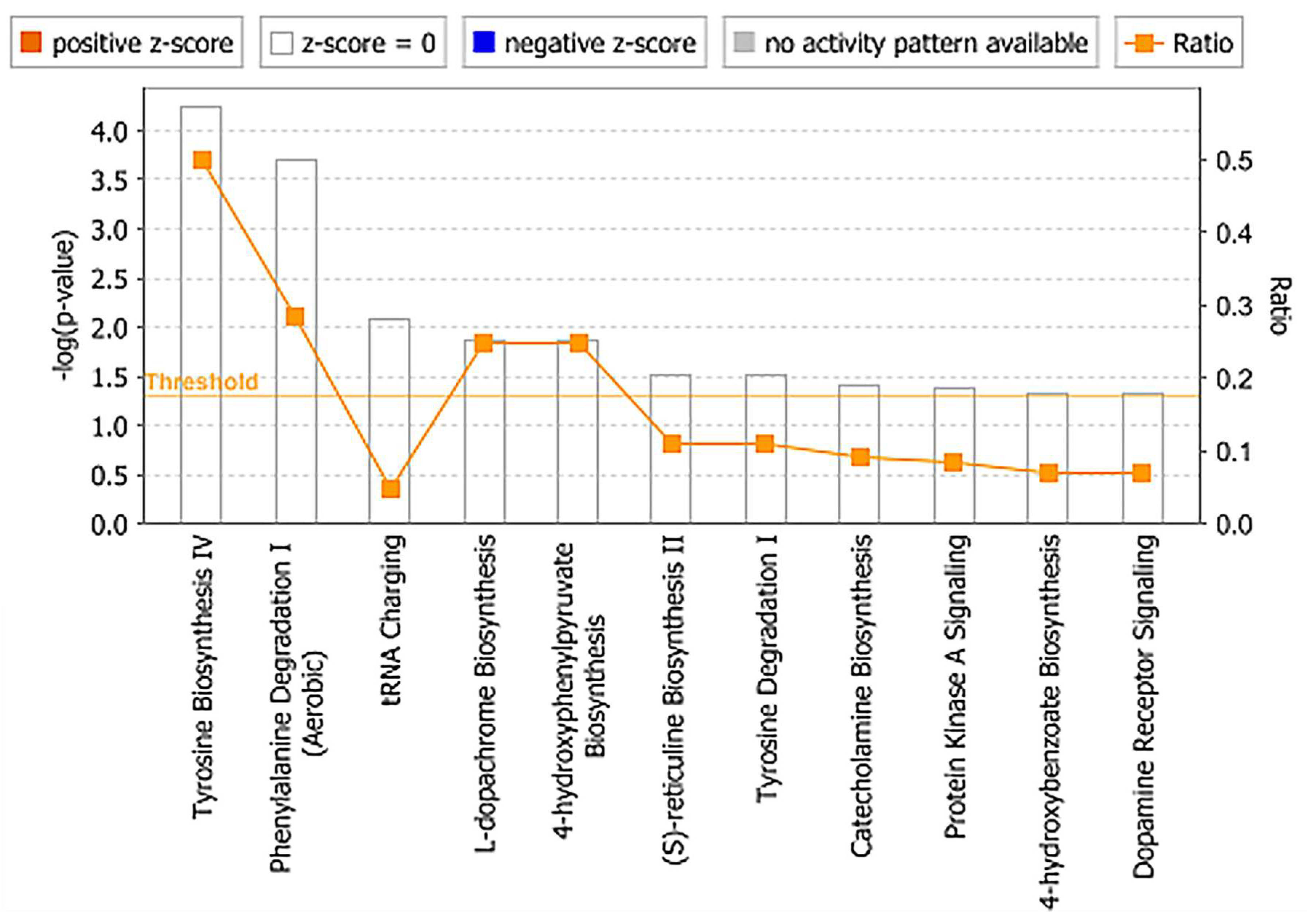
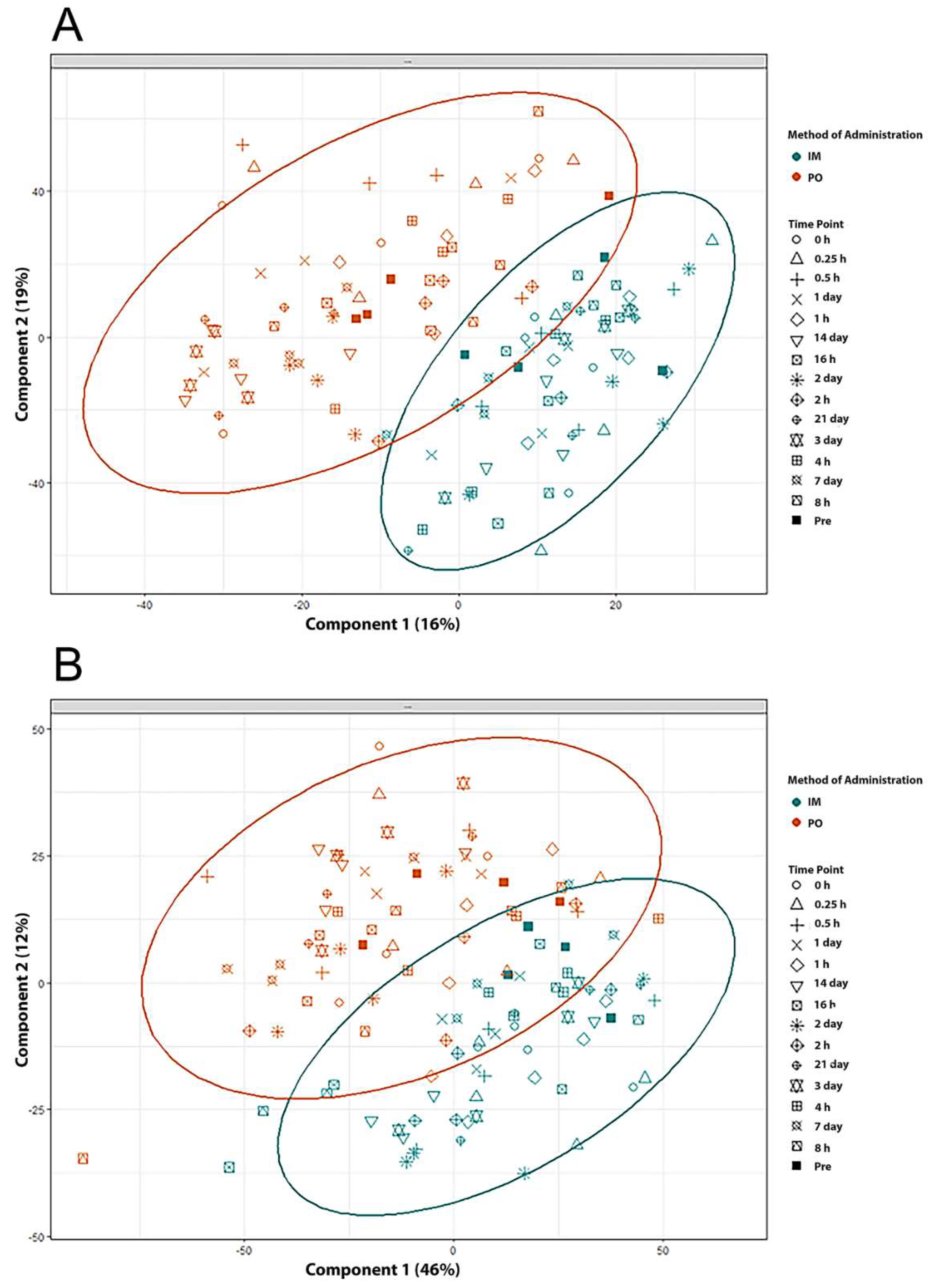
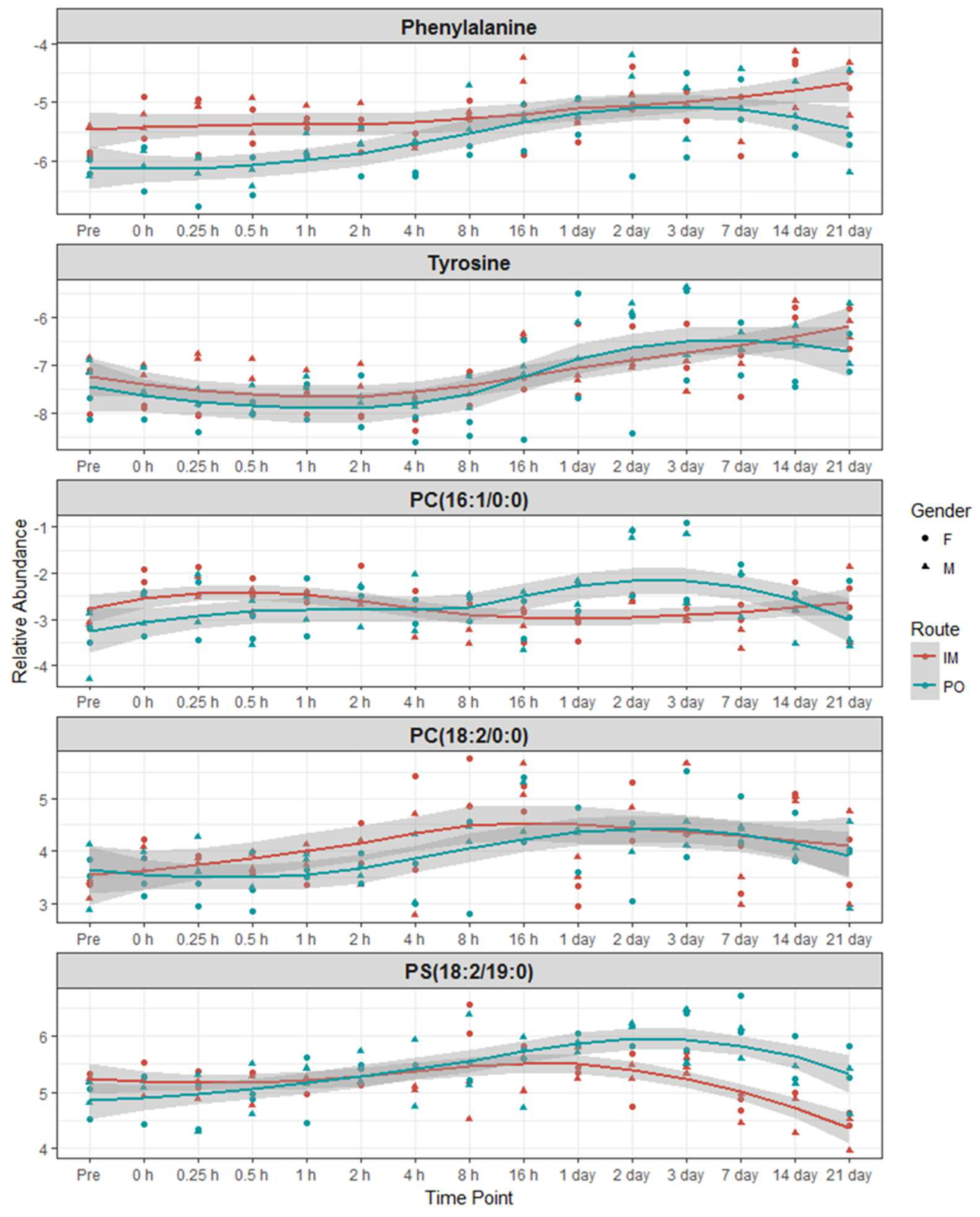
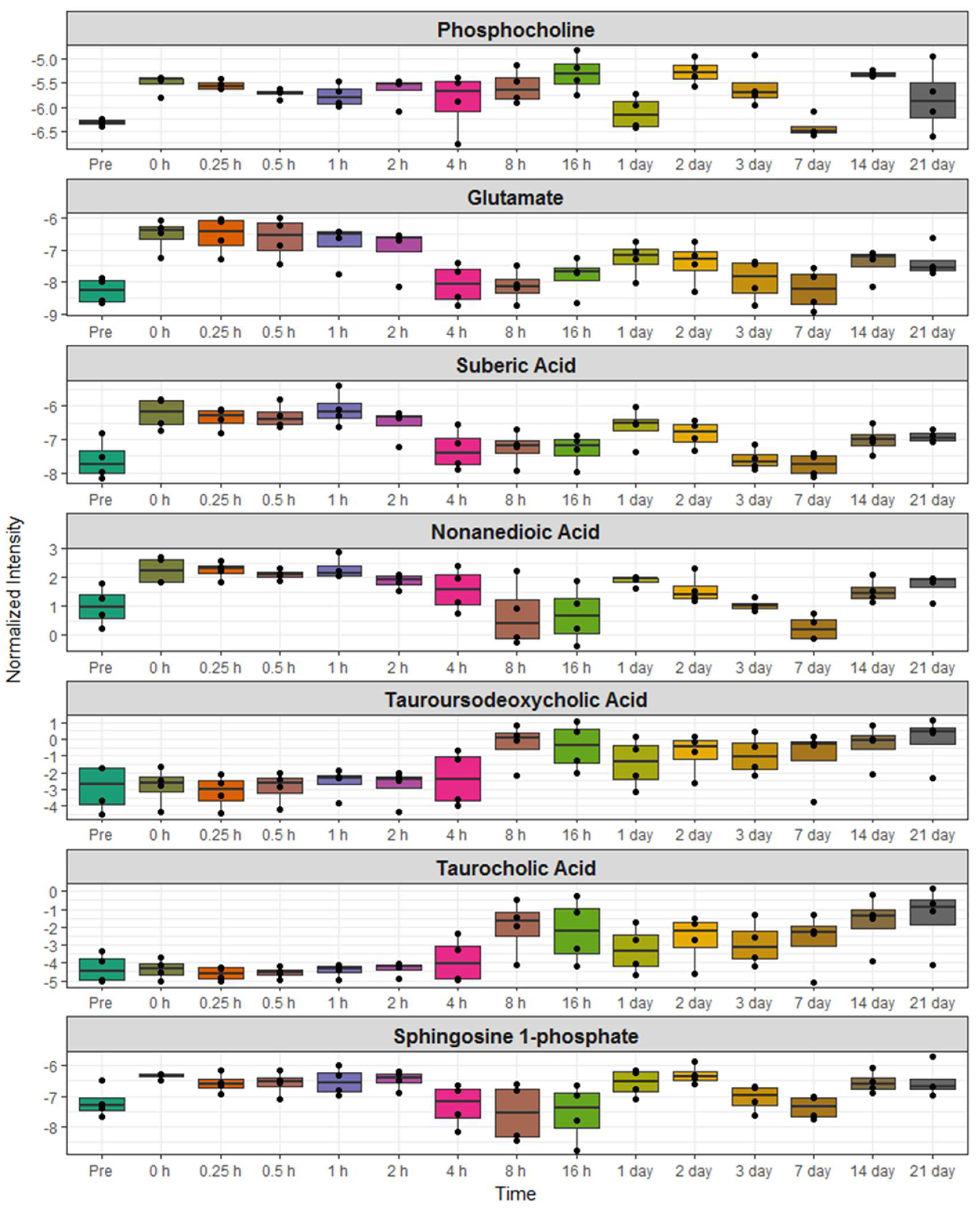
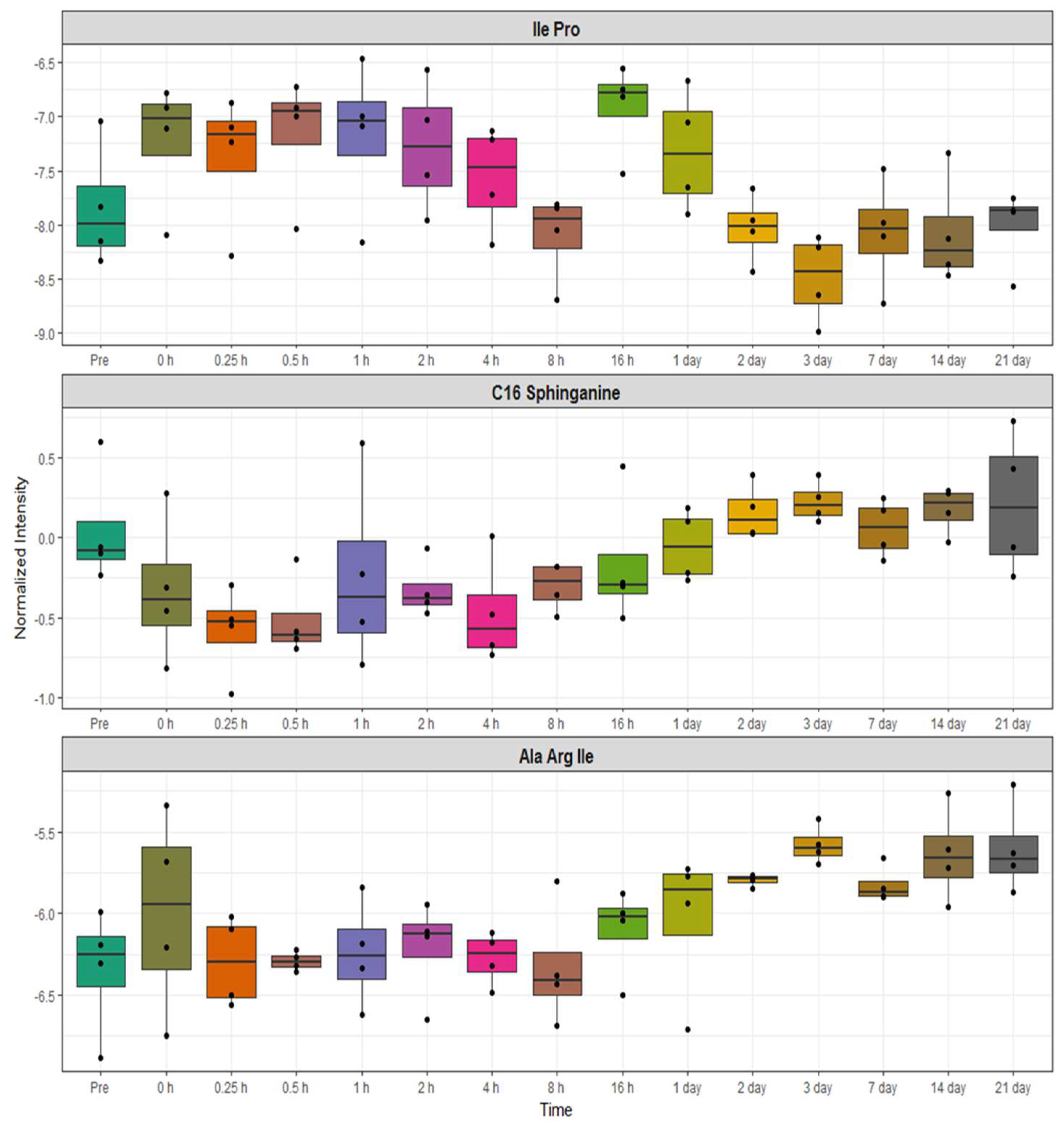
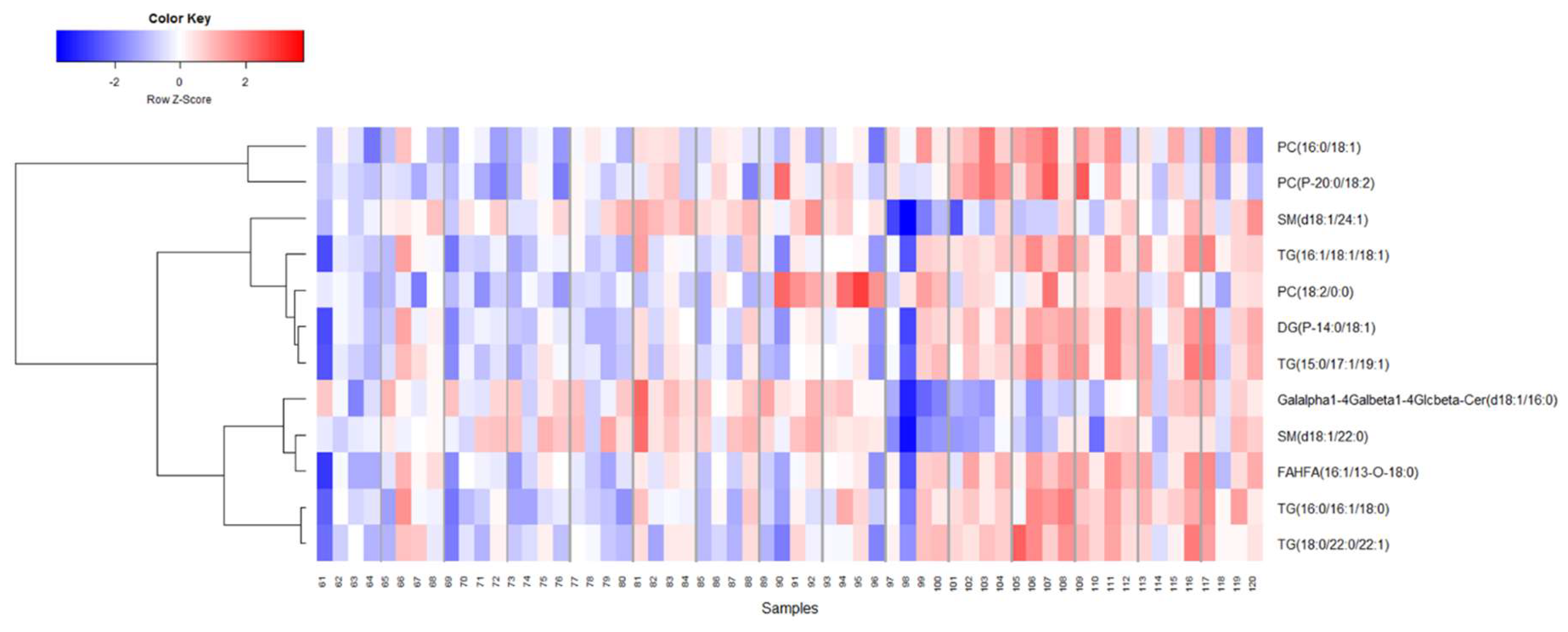
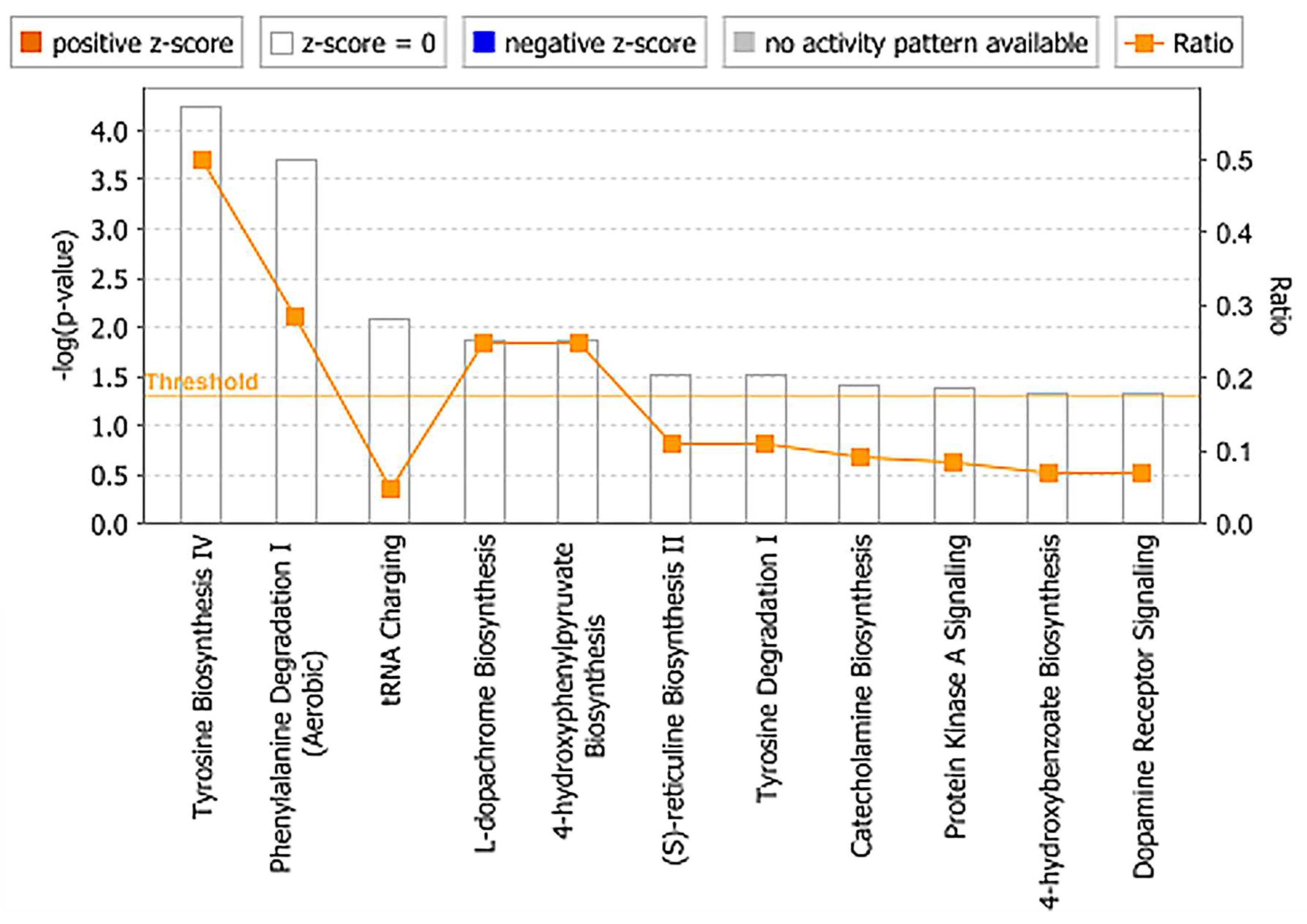
2.3. Identification of Metabolite Signatures of BIO 300 Administration in NHPs
3. Discussion
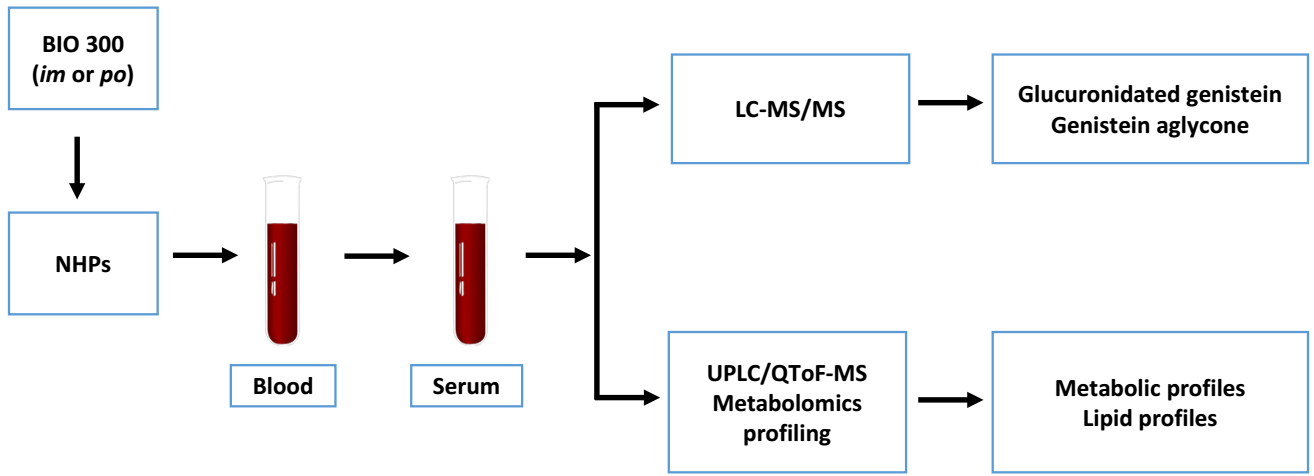
4. Materials and Methods
4.1. Animals and Animal Care
4.2. Drug Preparation and Administration
4.3. Serum Sample Collection
4.4. Pharmacokinetic Analysis
4.5. Serum Metabolomics Using UPLC QTOF Analysis
4.6. Analysis for CBC
4.7. Data Processing and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gosden, C.; Gardener, D. Weapons of mass destruction -threats and responses. BMJ 2005, 331, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, G.C.; McGeary, M.; McCutchen, S.R. Assessing Medical Preparedness to Respond to a Terrorist Nuclear Event: Workshop Report; The National Academies Press: Washington, DC, USA, 2009. [Google Scholar]
- Singh, V.K.; Seed, T.M. A review of radiation countermeasures focusing on injury-specific medicinals and regulatory approval status: Part I. Radiation sub-syndromes, animal models and FDA-approved countermeasures. Int. J. Radiat. Biol. 2017, 93, 851–869. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. FDA Approves Leukine for Acute Radiation Syndrome. Available online: https://www.fda.gov/downloads/EmergencyPreparedness/Counterterrorism/MedicalCountermeasures/AboutMCMi/UCM603226.pdf (accessed on 1 April 2018).
- U.S. Food and Drug Administration. FDA Approves Neupogen for Treatment of Patients with Radiation-Induced Myelosuppression Following a Radiological/Nuclear Incident. Available online: http://www.fda.gov/EmergencyPreparedness/Counterterrorism/MedicalCountermeasures/AboutMCMi/ucm443245.htm (accessed on 6 July 2016).
- National Institute of Allergic and Infectious Diseases. Pegfilgrastim Approved for Treating Acute Radiation Syndrome. Available online: https://www.niaid.nih.gov/topics/radnuc/Pages/pegfilgrastim.aspx (accessed on 18 August 2016).
- Singh, V.K.; Seed, T.M. An update on sargramostim for treatment of acute radiation syndrome. Drugs Today 2018, 54, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Farese, A.M.; MacVittie, T.J. Filgrastim for the treatment of hematopoietic acute radiation syndrome. Drugs Today 2015, 51, 537–548. [Google Scholar] [PubMed]
- Landauer, M.R.; Srinivasan, V.; Seed, T.M. Genistein treatment protects mice from ionizing radiation injury. J. Appl. Toxicol. 2003, 23, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Mungunsukh, O.; Zins, S.; Day, R.M.; Landauer, M.R. Genistein induces radioprotection by hematopoietic stem cell quiescence. Int. J. Radiat. Biol. 2008, 84, 713–726. [Google Scholar] [CrossRef]
- Day, R.; Barshishat-Kupper, M.; Mog, S.; McCart, E.; Prasanna, P.; Davis, T.; Landauer, M. Genistein protects against biomarkers of delayed lung sequelae in mice surviving high dose total body irradiation. J. Radiat. Res. 2008, 49, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Clarke, T.K.; Mog, S.R.; Landauer, M.R. Subcutaneous administration of genistein prior to lethal irradiation supports multilineage, hematopoietic progenitor cell recovery and survival. Int. J. Radiat. Biol. 2007, 83, 141–151. [Google Scholar] [CrossRef]
- Singh, V.K.; Grace, M.B.; Parekh, V.I.; Whitnall, M.H.; Landauer, M.R. Effects of genistein administration on cytokine induction in whole-body gamma irradiated mice. Int. Immunopharmacol. 2009, 9, 1401–1410. [Google Scholar] [CrossRef]
- Yang, Z.; Kulkarni, K.; Zhu, W.; Hu, M. Bioavailability and pharmacokinetics of genistein: Mechanistic studies on its ADME. Anticancer Agents Med. Chem. 2012, 12, 1264–1280. [Google Scholar] [CrossRef]
- Jackson, I.L.; Zodda, A.; Gurung, G.; Pavlovic, R.; Kaytor, M.D.; Kuskowski, M.A.; Vujaskovic, Z. BIO 300, a nanosuspension of genistein, mitigates pneumonitis/fibrosis following high-dose radiation exposure in the C57L/J murine model. Br. J. Pharmacol. 2017, 174, 4738–4750. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.T.; Li, X.H.; Fu, D.; Xiao, M.; Landauer, M.R. Genistein nanoparticles protect mouse hematopoietic system and prevent proinflammatory factors after gamma irradiation. Radiat. Res. 2013, 180, 316–325. [Google Scholar] [CrossRef]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Strom, A.; Treuter, E.; Warner, M.; et al. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; van der Saag, P.T.; van der Burg, B.; Gustafsson, J.K. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef] [PubMed]
- Raffoul, J.J.; Wang, Y.; Kucuk, O.; Forman, J.D.; Sarkar, F.H.; Hillman, G.G. Genistein inhibits radiation-induced activation of NF-kappaB in prostate cancer cells promoting apoptosis and G2/M cell cycle arrest. BMC Cancer 2006, 6, 107. [Google Scholar] [CrossRef]
- Sierens, J.; Hartley, J.A.; Campbell, M.J.; Leathem, A.J.; Woodside, J.V. Effect of phytoestrogen and antioxidant supplementation on oxidative DNA damage assessed using the comet assay. Mutat. Res. 2001, 485, 169–176. [Google Scholar] [CrossRef]
- Sierens, J.; Hartley, J.A.; Campbell, M.J.; Leathem, A.J.; Woodside, J.V. In vitro isoflavone supplementation reduces hydrogen peroxide-induced DNA damage in sperm. Teratog. Carcinog. Mutagen. 2002, 22, 227–234. [Google Scholar] [CrossRef]
- Jackson, I.L.; Xu, P.; Hadley, C.; Katz, B.P.; McGurk, R.; Down, J.D.; Vujaskovic, Z. A preclinical rodent model of radiation-induced lung injury for medical countermeasure screening in accordance with the FDA animal rule. Health Phys. 2012, 103, 463–473. [Google Scholar] [CrossRef]
- Jackson, I.L.; Vujaskovic, Z.; Down, J.D. Revisiting strain-related differences in radiation sensitivity of the mouse lung: Recognizing and avoiding the confounding effects of pleural effusions. Radiat. Res. 2010, 173, 10–20. [Google Scholar] [CrossRef]
- Jackson, I.L.; Vujaskovic, Z.; Down, J.D. A further comparison of pathologies after thoracic irradiation among different mouse strains: Finding the best preclinical model for evaluating therapies directed against radiation-induced lung damage. Radiat. Res. 2011, 175, 510–518. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Guidance for Industry: Product Development under the Animal Rule. Available online: http://www.fda.gov/downloads/Drugs/GuidanceComplianceRegulatoryInformation/Guidances/UCM399217.pdf (accessed on 5 July 2018).
- Dainiak, N.; Waselenko, J.K.; Armitage, J.O.; MacVittie, T.J.; Farese, A.M. The hematologist and radiation casualties. Hematol. Am. Soc. Hematol. Educ. Program 2003, 2003, 473–496. [Google Scholar] [CrossRef]
- Weisdorf, D.; Chao, N.; Waselenko, J.K.; Dainiak, N.; Armitage, J.O.; McNiece, I.; Confer, D. Acute radiation injury: Contingency planning for triage, supportive care, and transplantation. Biol. Blood Marrow Transplant. 2006, 12, 672–682. [Google Scholar] [CrossRef]
- Waselenko, J.K.; MacVittie, T.J.; Blakely, W.F.; Pesik, N.; Wiley, A.L.; Dickerson, W.E.; Tsu, H.; Confer, D.L.; Coleman, C.N.; Seed, T.; et al. Medical management of the acute radiation syndrome: Recommendations of the Strategic National Stockpile Radiation Working Group. Ann. Intern. Med. 2004, 140, 1037–1051. [Google Scholar] [CrossRef]
- Kaneko, A.; Matsumoto, T.; Matsubara, Y.; Sekiguchi, K.; Koseki, J.; Yakabe, R.; Aoki, K.; Aiba, S.; Yamasaki, K. Glucuronides of phytoestrogen flavonoid enhance macrophage function via conversion to aglycones by beta-glucuronidase in macrophages. Immun. Inflamm. Dis. 2017, 5, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Raabe, B.M.; Lovaglio, J.; Grover, G.S.; Brown, S.A.; Boucher, J.F.; Yuan, Y.; Civil, J.R.; Gillhouse, K.A.; Stubbs, M.N.; Hoggatt, A.F.; et al. Pharmacokinetics of cefovecin in cynomolgus macaques (Macaca fascicularis), olive baboons (Papio anubis), and rhesus macaques (Macaca mulatta). J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 389–395. [Google Scholar] [PubMed]
- Papp, R.; Popovic, A.; Kelly, N.; Tschirret-Guth, R. Pharmacokinetics of Cefovecin in squirrel monkey (Saimiri sciureus), rhesus macaques (Macaca mulatta), and cynomolgus macaques (Macaca fascicularis). J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 805–808. [Google Scholar]
- Vile, G.F.; Tyrrell, R.M. UVA radiation-induced oxidative damage to lipids and proteins in vitro and in human skin fibroblasts is dependent on iron and singlet oxygen. Free Radic. Biol. Med. 1995, 18, 721–730. [Google Scholar] [CrossRef]
- Gulbahar, O.; Aricioglu, A.; Akmansu, M.; Turkozer, Z. Effects of radiation on protein oxidation and lipid peroxidation in the brain tissue. Transplant. Proc. 2009, 41, 4394–4396. [Google Scholar] [CrossRef]
- Ó Broin, P.; Vaitheesvaran, B.; Saha, S.; Hartil, K.; Chen, E.I.; Goldman, D.; Fleming, W.H.; Kurland, I.J.; Guha, C.; Golden, A. Intestinal microbiota-derived metabolomic blood plasma markers for prior radiation injury. Int. J. Radiat. Oncol. Biol. Phys. 2015, 91, 360–367. [Google Scholar] [CrossRef]
- Arab, K.; Rossary, A.; Flourie, F.; Tourneur, Y.; Steghens, J.P. Docosahexaenoic acid enhances the antioxidant response of human fibroblasts by upregulating gamma-glutamyl-cysteinyl ligase and glutathione reductase. Br. J. Nutr. 2006, 95, 18–26. [Google Scholar] [CrossRef]
- Jones, J.W.; Jackson, I.L.; Vujaskovic, Z.; Kaytor, M.D.; Kane, M.A. Targeted metabolomics identifies pharmacodynamic biomarkers for BIO 300 mitigation of radiation-induced lung injury. Pharm. Res. 2017, 34, 2698–2709. [Google Scholar] [CrossRef]
- Singh, V.K.; Kulkarni, S.; Fatanmi, O.O.; Wise, S.Y.; Newman, V.L.; Romaine, P.L.; Hendrickson, H.; Gulani, J.; Ghosh, S.P.; Kumar, K.S.; et al. Radioprotective efficacy of gamma-tocotrienol in nonhuman primates. Radiat. Res. 2016, 185, 285–298. [Google Scholar] [CrossRef] [PubMed]
- National Research Council of the National Academy of Sciences. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Kaur, P.; Rizk, N.; Ibrahim, S.; Luo, Y.; Younes, N.; Perry, B.; Dennis, K.; Zirie, M.; Luta, G.; Cheema, A.K. Quantitative metabolomic and lipidomic profiling reveals aberrant amino acid metabolism in type 2 diabetes. Mol. Biosyst. 2013, 9, 307–317. [Google Scholar] [CrossRef] [PubMed]
- LaConti, J.J.; Laiakis, E.C.; Mays, A.D.; Peran, I.; Kim, S.E.; Shay, J.W.; Riegel, A.T.; Fornace, A.J.; Wellstein, A. Distinct serum metabolomics profiles associated with malignant progression in the Kras(G12D) mouse model of pancreatic ductal adenocarcinoma. BMC Genom. 2015, 16, S1. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 9 October 2018).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| im Free Genistein (Aglycone) (50 mg/kg) | |||||
| NHP | Tmax (hr) | Cmax (ng/mL) | T1/2 (h) | AUC0–48 (ng.h/mL) | AUC0–∞ (ng.h/mL) |
| NHP 1 | 2 | 3550 | 5.47 | 24,159 | 24,234 |
| NHP 2 | 1 | 1980 | 15.26 | 44,783 | 50,024 |
| NHP 3 | 2 | 1410 | 18.15 | 28,194 | 33,667 |
| NHP 4 | 2 | 1580 | 20.17 | 30,202 | 36,953 |
| Average | 1.75 | 2130 | 14.76 | 31,834 | 36,219 |
| SD | 0.50 | 976 | 6.51 | 8991 | 10,666 |
| Med | 2.00 | 1780 | 16.70 | 29,188 | 35,310 |
| Max | 2.00 | 3550 | 20.17 | 44,783 | 50,024 |
| Min | 1.00 | 1410 | 5.47 | 24,159 | 24,234 |
| %CV | 28.57 | 45.84 | 44.11 | 28.24 | 29.45 |
| im Total Genistein (50 mg/kg) | |||||
| NHP | Tmax (hr) | Cmax (ng/mL) | T1/2 (h) | AUC0–48 (ng.h/mL) | AUC0–∞ (ng.h/mL) |
| NHP 1 | 4 | 5760 | 6.05 | 49,046 | 49,358 |
| NHP 2 | 8 | 3570 | 15.33 | 83,671 | 91,588 |
| NHP 3 | 2 | 2950 | 18.91 | 53,329 | 64,788 |
| NHP 4 | 2 | 2360 | 19.92 | 49,330 | 60,051 |
| Average | 4.00 | 3660 | 15.05 | 58,844 | 66,446 |
| SD | 2.83 | 1485 | 6.31 | 16,666 | 17,961 |
| Med | 3.00 | 3260 | 17.12 | 51,330 | 62,419 |
| Max | 8.00 | 5760 | 19.92 | 83,671 | 91,588 |
| Min | 2.00 | 2360 | 6.05 | 49,046 | 49,358 |
| %CV | 70.71 | 40.56 | 41.94 | 28.32 | 27.03 |
| po Free Genistein (Aglycone) (100 mg/kg) | |||||
| NHP | Tmax (hr) | Cmax (ng/mL) | T1/2 (h) | AUC0–48 (ng.h/mL) | AUC0–∞ (ng.h/mL) |
| NHP 1 | 4 | 398 | 2.57 | 3041 | 3048 |
| NHP 2 | 2 | 273 | 2.08 | 1195 | 1201 |
| NHP 3 | 2 | 162 | 4.22 | 1304 | 1316 |
| NHP 4 | 2 | 411 | 1.84 | 2127 | 2133 |
| Average | 2.50 | 311 | 2.68 | 1917 | 1924 |
| SD | 1.00 | 117 | 1.07 | 857 | 856 |
| Med | 2.00 | 336 | 2.32 | 1715 | 1724 |
| Max | 4.00 | 411 | 4.22 | 3041 | 3048 |
| Min | 2.00 | 162 | 1.84 | 1195 | 1201 |
| %CV | 40.00 | 37.69 | 40.01 | 44.73 | 44.50 |
| po total genistein (100 mg/kg) | |||||
| NHP | Tmax (hr) | Cmax (ng/mL) | T1/2 (h) | AUC0–48 (ng.h/mL) | AUC0–∞ (ng.h/mL) |
| NHP 1 | 4 | 10,400 | 2.37 | 60,788 | 60,805 |
| NHP 2 | 2 | 6330 | 2.88 | 38,775 | 38,912 |
| NHP 3 | 2 | 5150 | 4.09 | 54,492 | 54,860 |
| NHP 4 | 2 | 5560 | 2.19 | 29,703 | 29,789 |
| Average | 2.50 | 6860 | 2.88 | 45,939 | 46,092 |
| SD | 1.00 | 2410 | 0.86 | 14,243 | 14,267 |
| Med | 2.00 | 5945 | 2.63 | 46,633 | 46,886 |
| Max | 4.00 | 10,400 | 4.09 | 60,788 | 60,805 |
| Min | 2.00 | 5150 | 2.19 | 29,703 | 29,789 |
| %CV | 40.00 | 35.13 | 29.70 | 31.00 | 30.95 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheema, A.K.; Mehta, K.Y.; Santiago, P.T.; Fatanmi, O.O.; Kaytor, M.D.; Singh, V.K. Pharmacokinetic and Metabolomic Studies with BIO 300, a Nanosuspension of Genistein, in a Nonhuman Primate Model. Int. J. Mol. Sci. 2019, 20, 1231. https://doi.org/10.3390/ijms20051231
Cheema AK, Mehta KY, Santiago PT, Fatanmi OO, Kaytor MD, Singh VK. Pharmacokinetic and Metabolomic Studies with BIO 300, a Nanosuspension of Genistein, in a Nonhuman Primate Model. International Journal of Molecular Sciences. 2019; 20(5):1231. https://doi.org/10.3390/ijms20051231
Chicago/Turabian StyleCheema, Amrita K., Khyati Y. Mehta, Paola T. Santiago, Oluseyi O. Fatanmi, Michael D. Kaytor, and Vijay K. Singh. 2019. "Pharmacokinetic and Metabolomic Studies with BIO 300, a Nanosuspension of Genistein, in a Nonhuman Primate Model" International Journal of Molecular Sciences 20, no. 5: 1231. https://doi.org/10.3390/ijms20051231
APA StyleCheema, A. K., Mehta, K. Y., Santiago, P. T., Fatanmi, O. O., Kaytor, M. D., & Singh, V. K. (2019). Pharmacokinetic and Metabolomic Studies with BIO 300, a Nanosuspension of Genistein, in a Nonhuman Primate Model. International Journal of Molecular Sciences, 20(5), 1231. https://doi.org/10.3390/ijms20051231