Potential Analysis and Preparation of Chitosan Oligosaccharides as Oral Nutritional Supplements of Cancer Adjuvant Therapy

Abstract
:1. Introduction
2. Results and Discussion
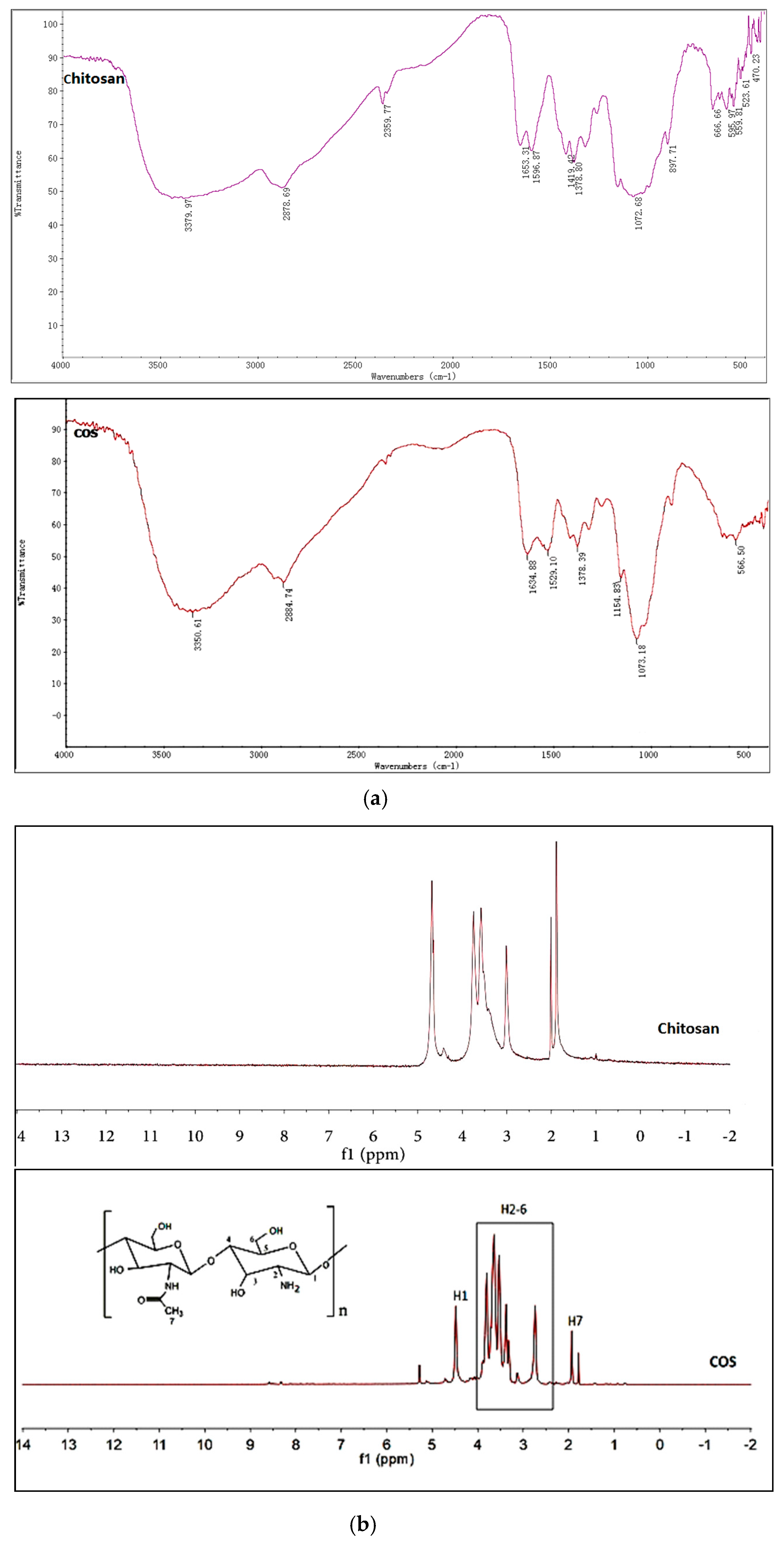
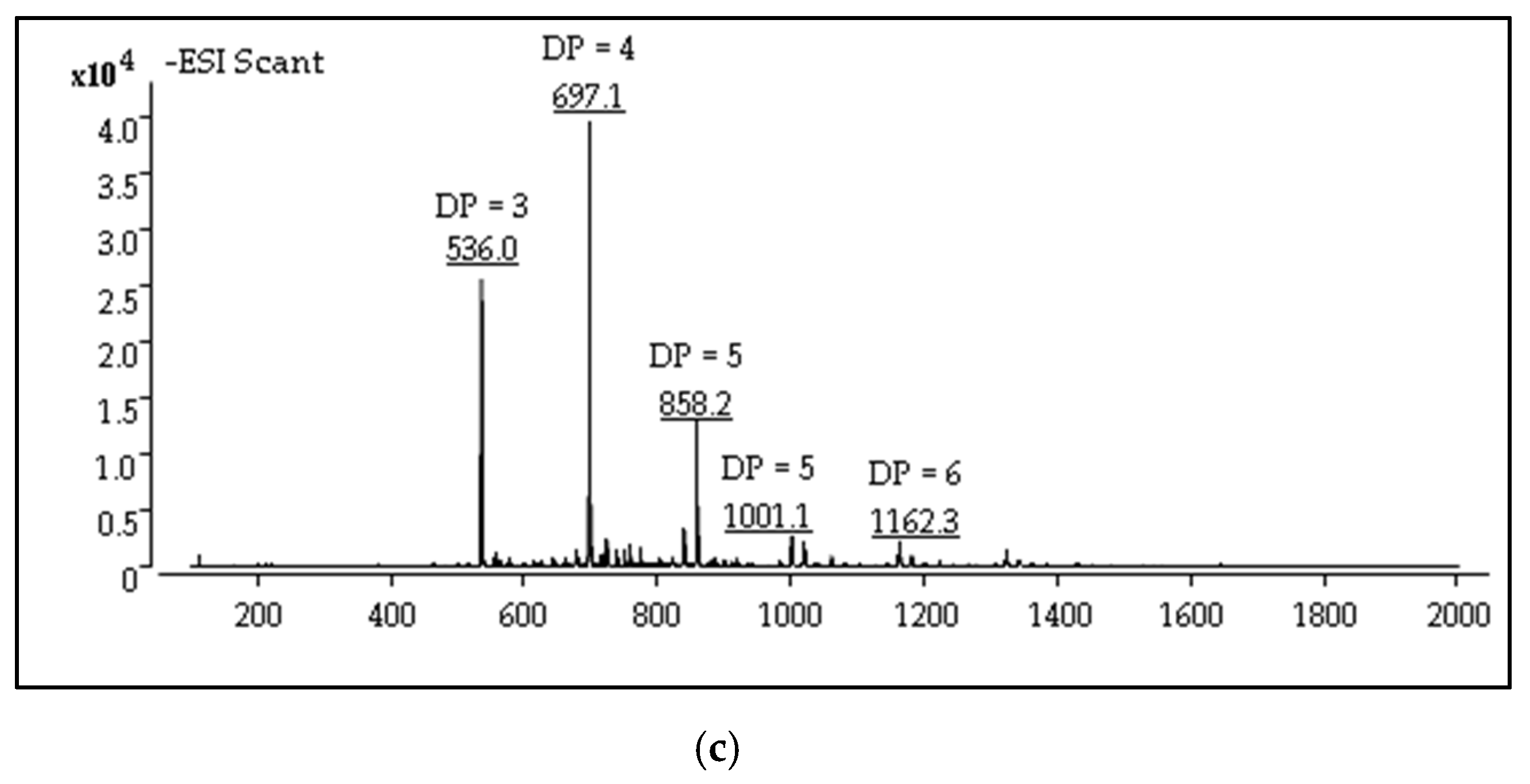
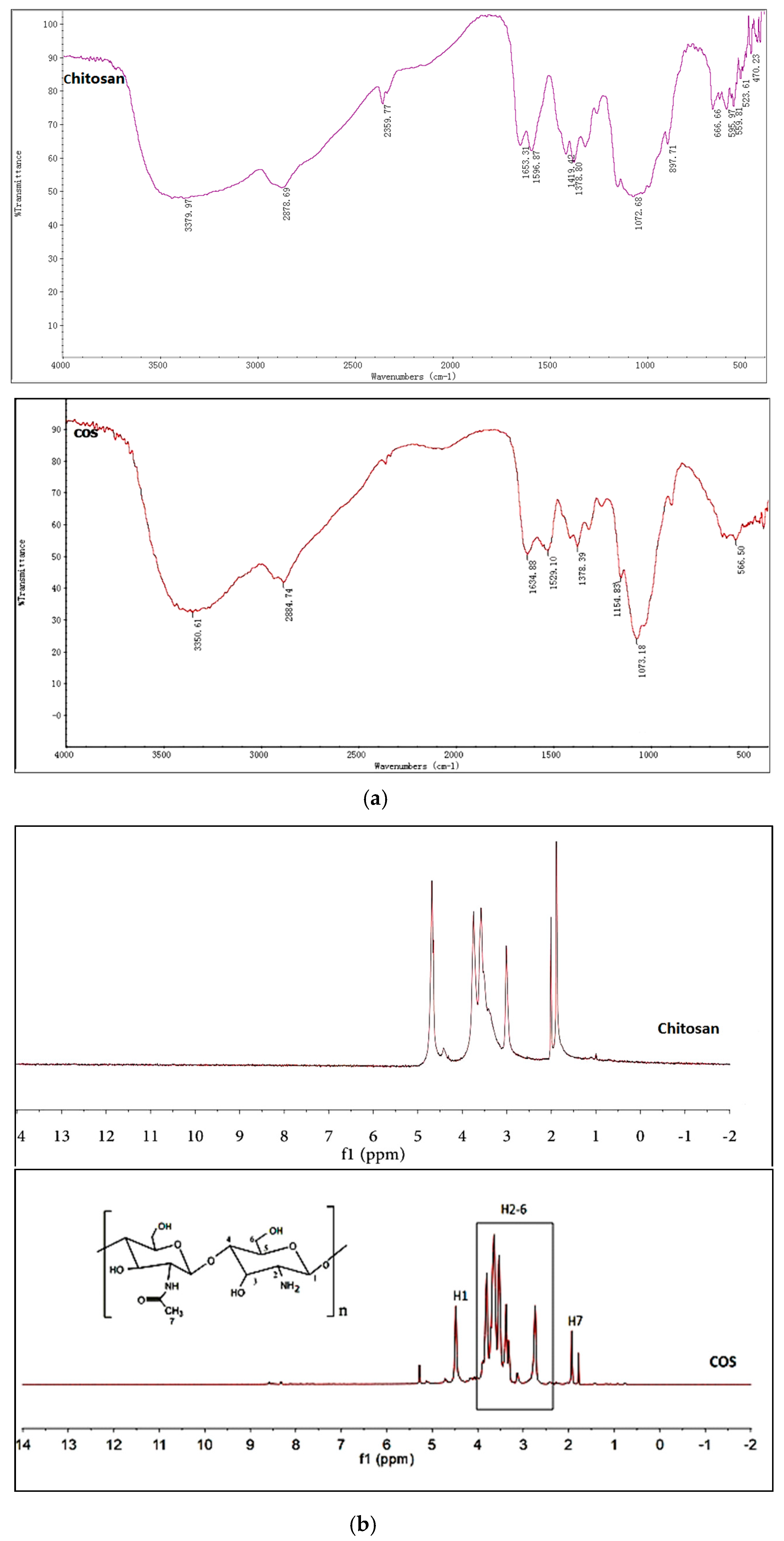
2.1. Structures Characterization of COS
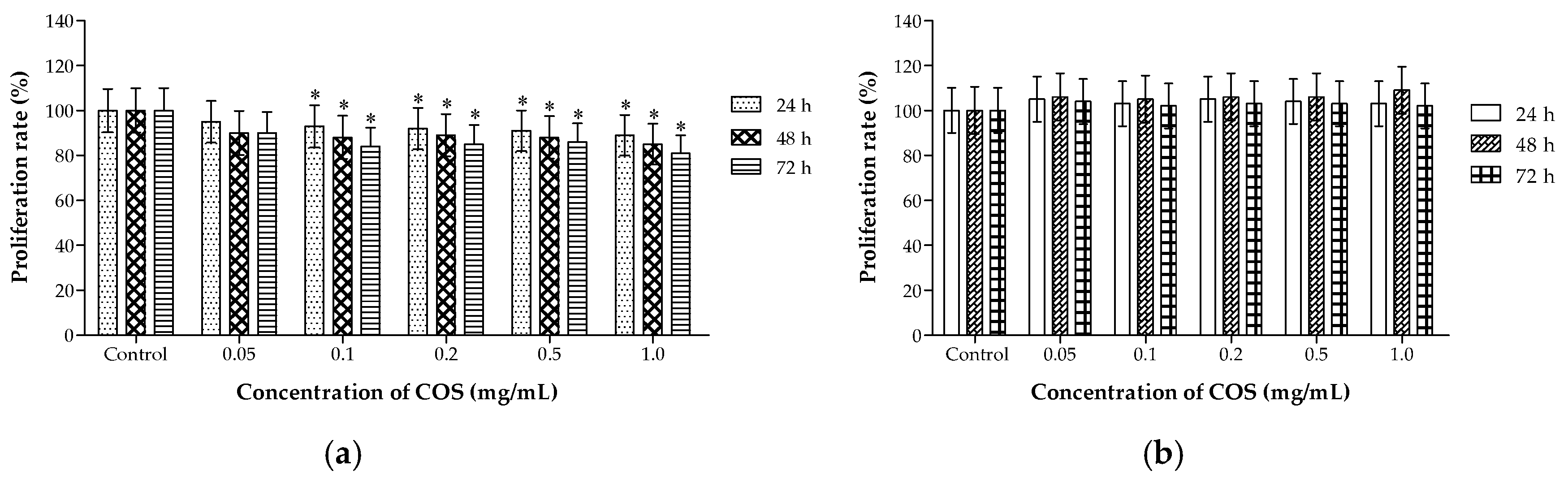
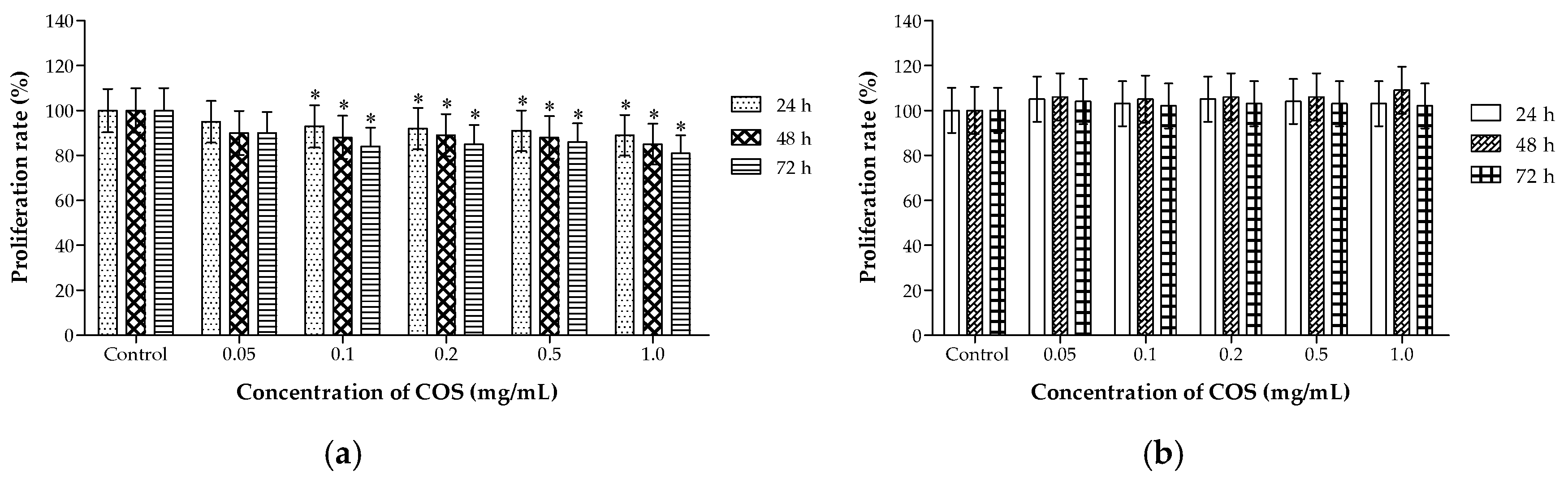
2.2. Different Effects of COS on the Viability of A549 and HL-7702 Cells
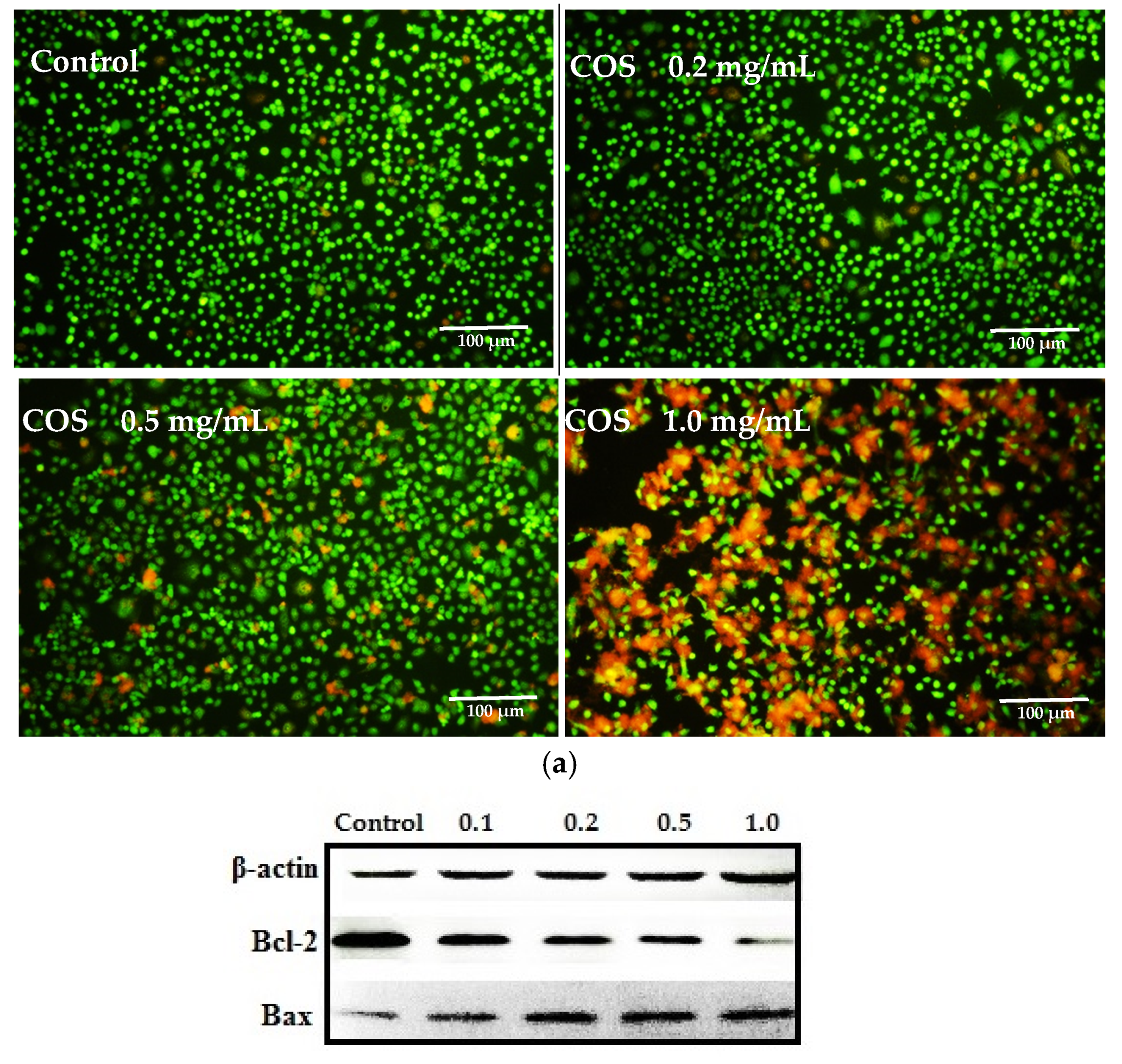
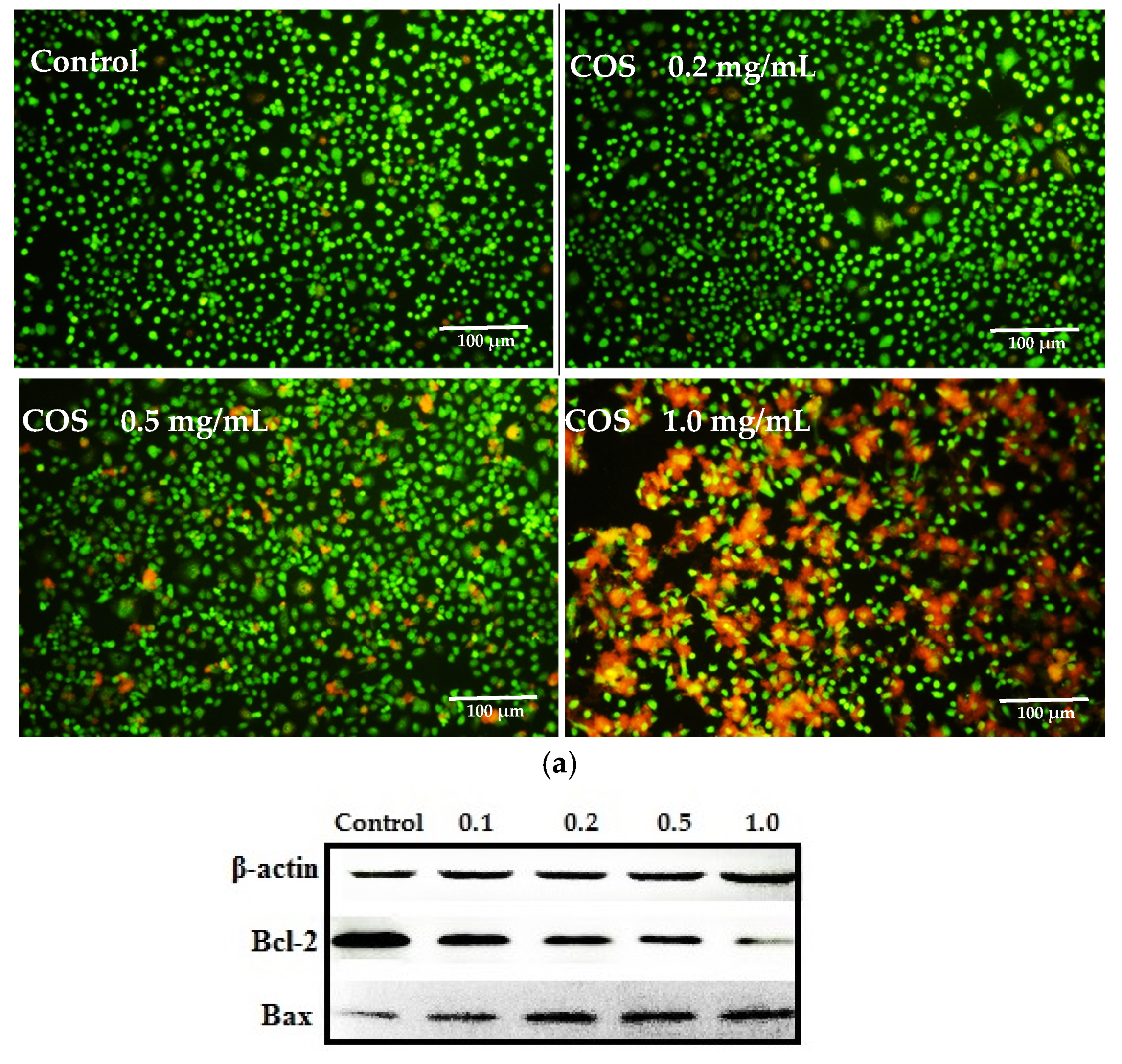
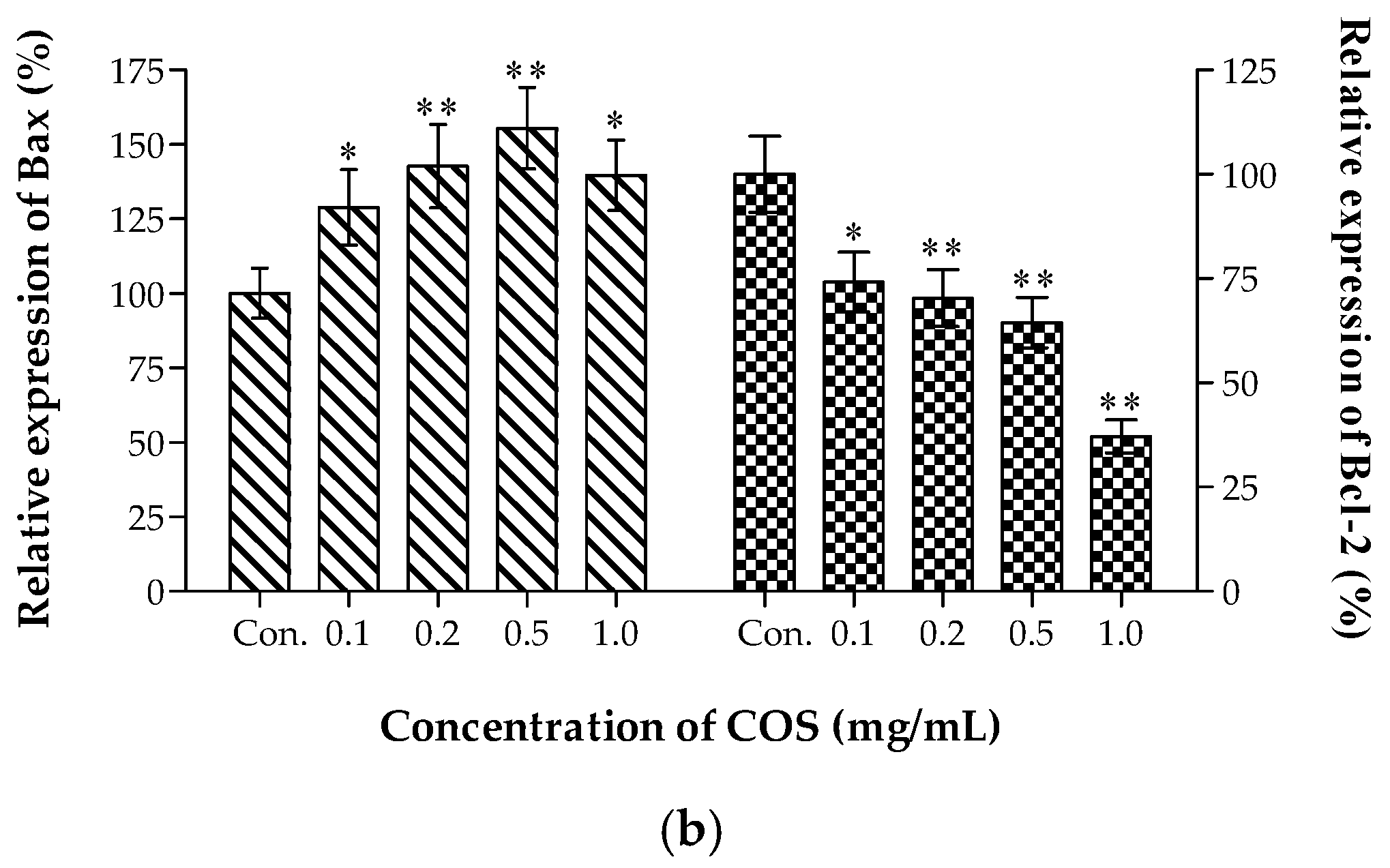
2.3. Growth Inhibiting and Apoptosis Promoting of COS on A549 Cells
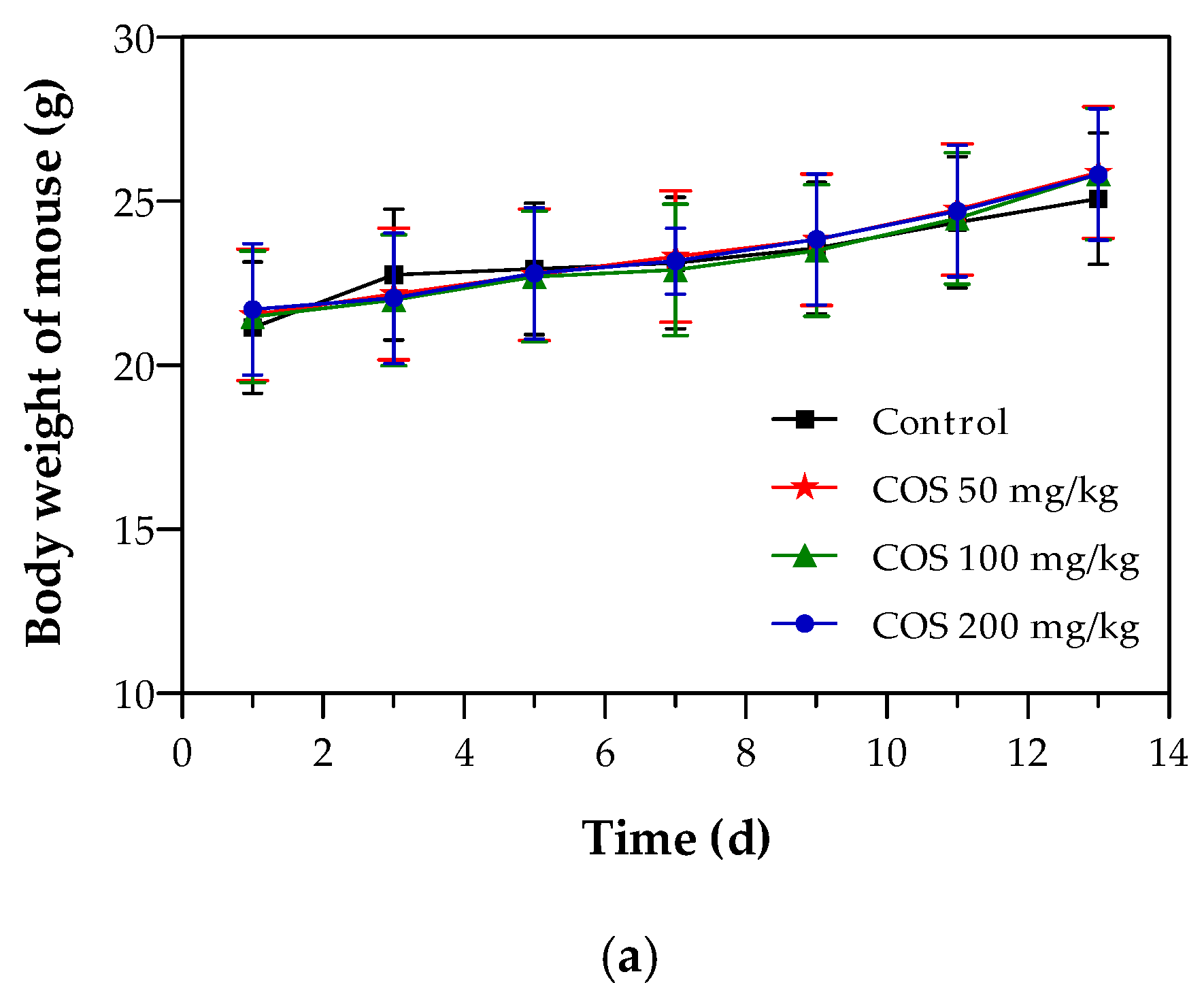
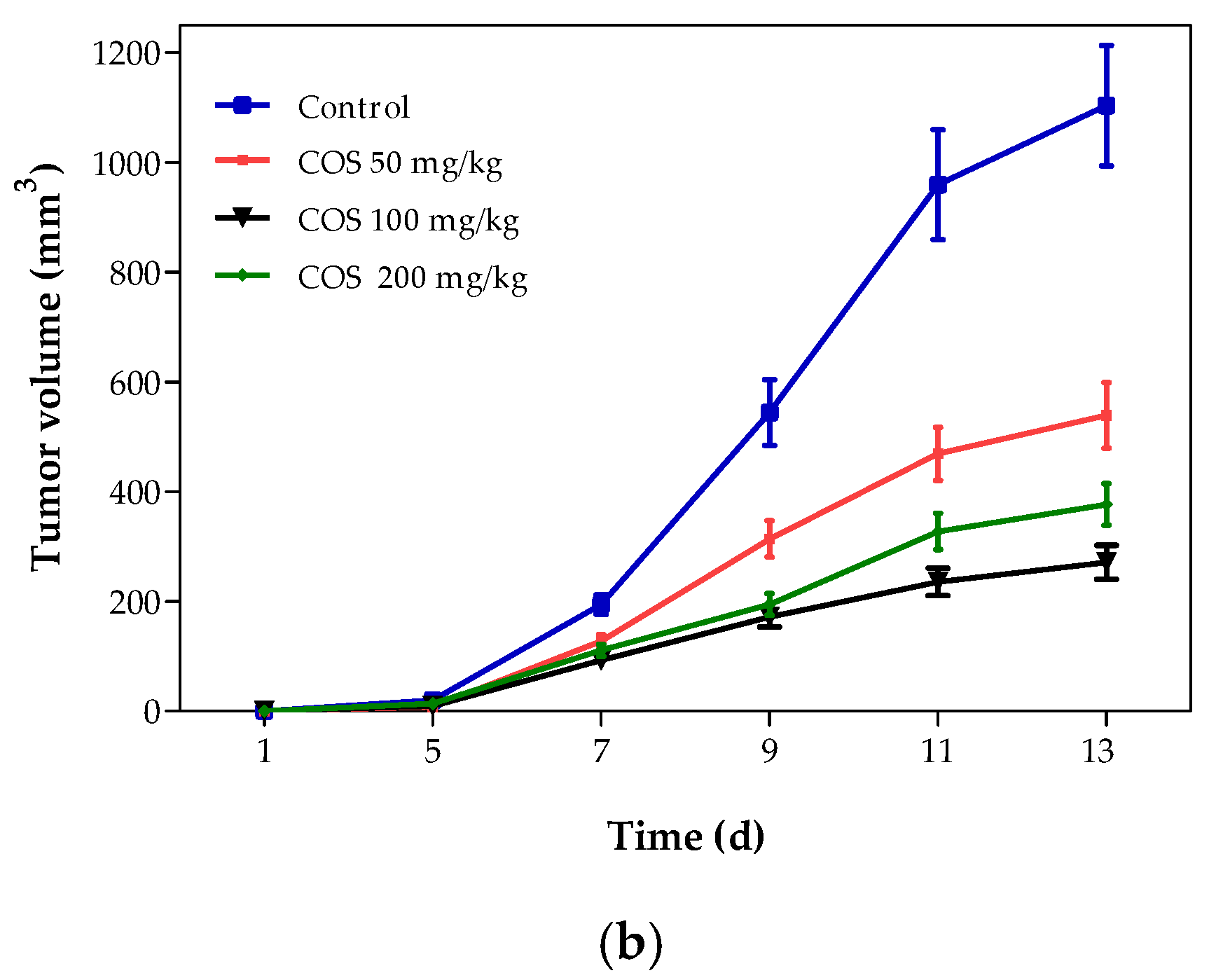
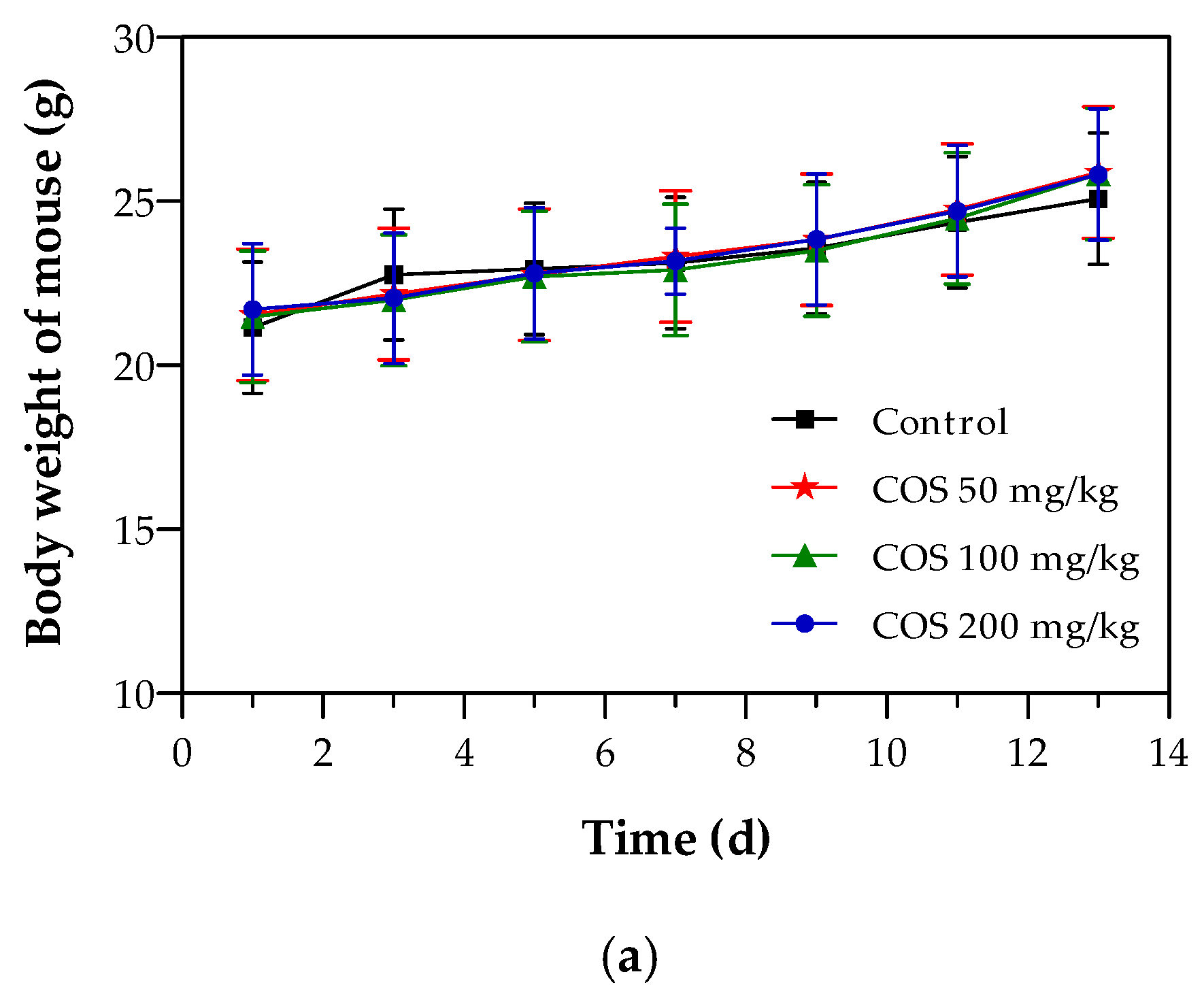
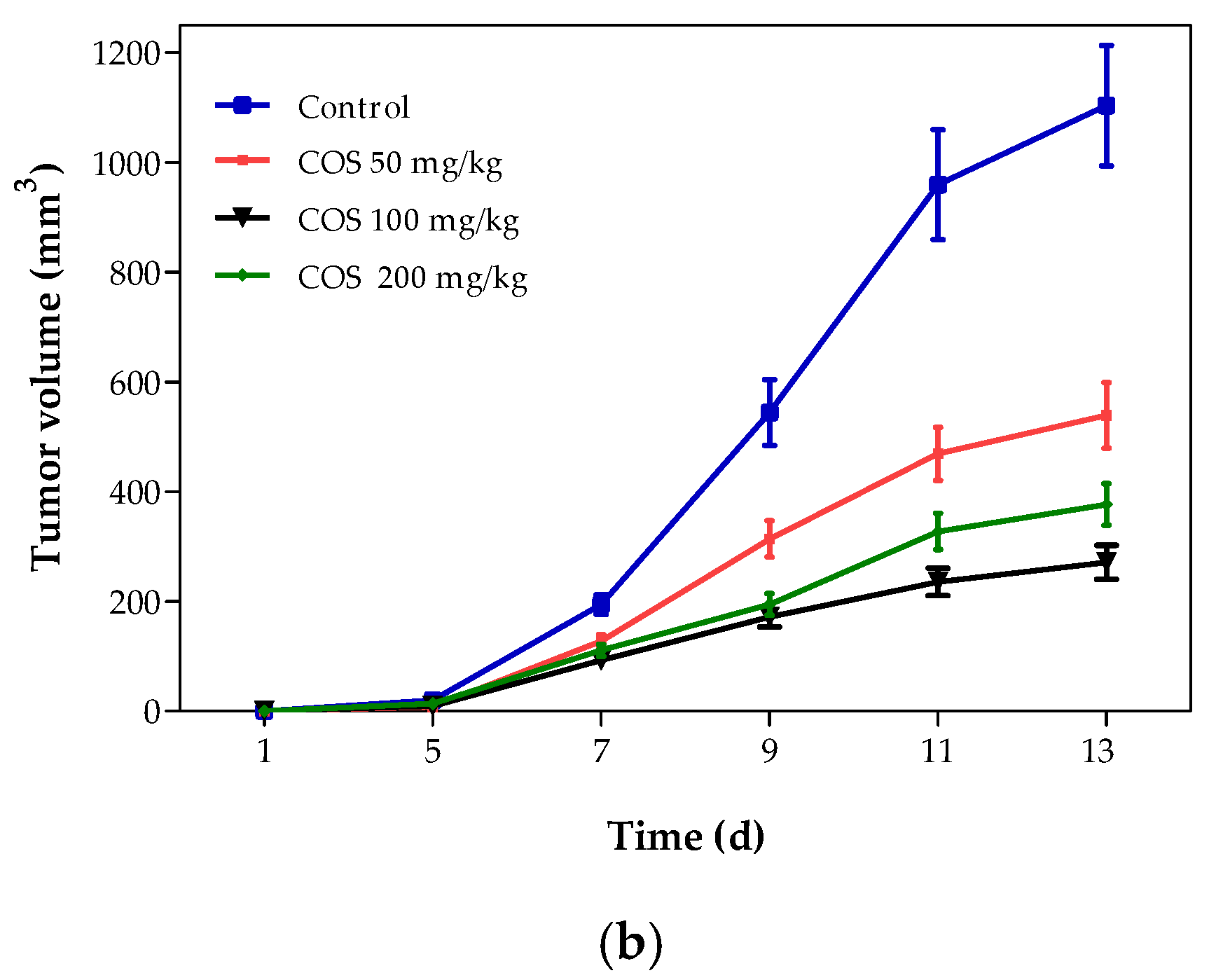
2.4. Effects of COS on Body Weight and Tumor Growth of Mouse
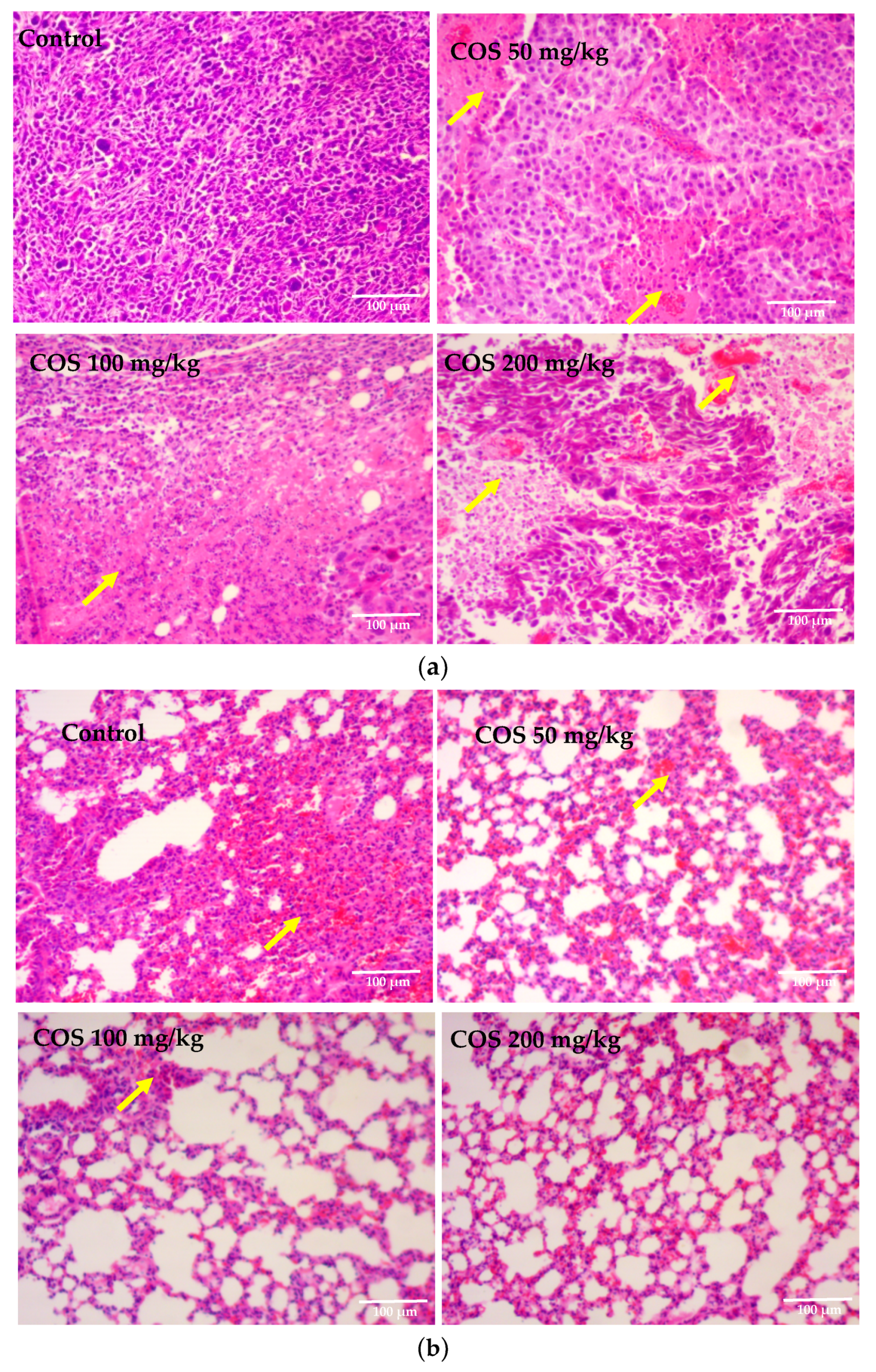
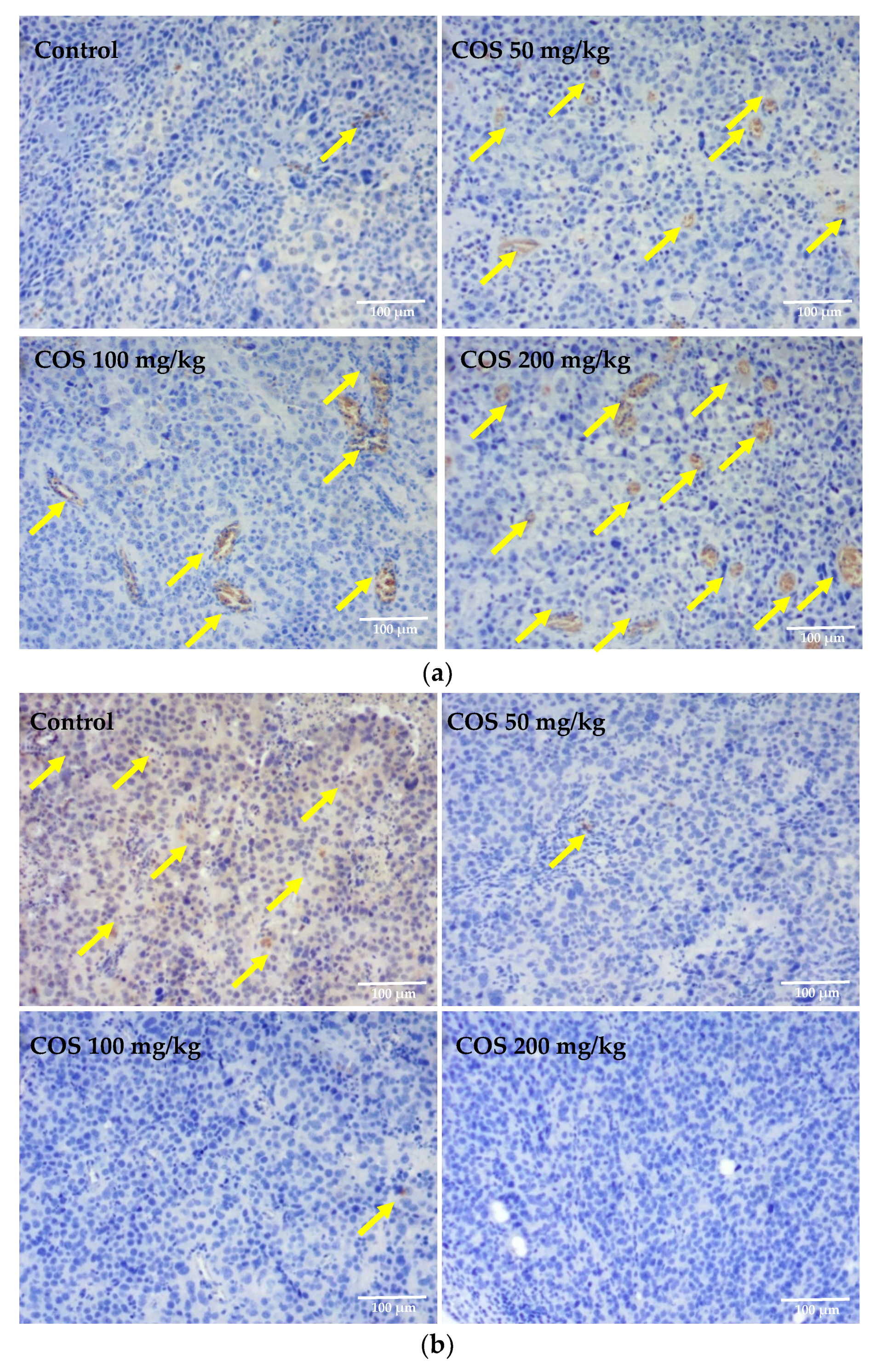
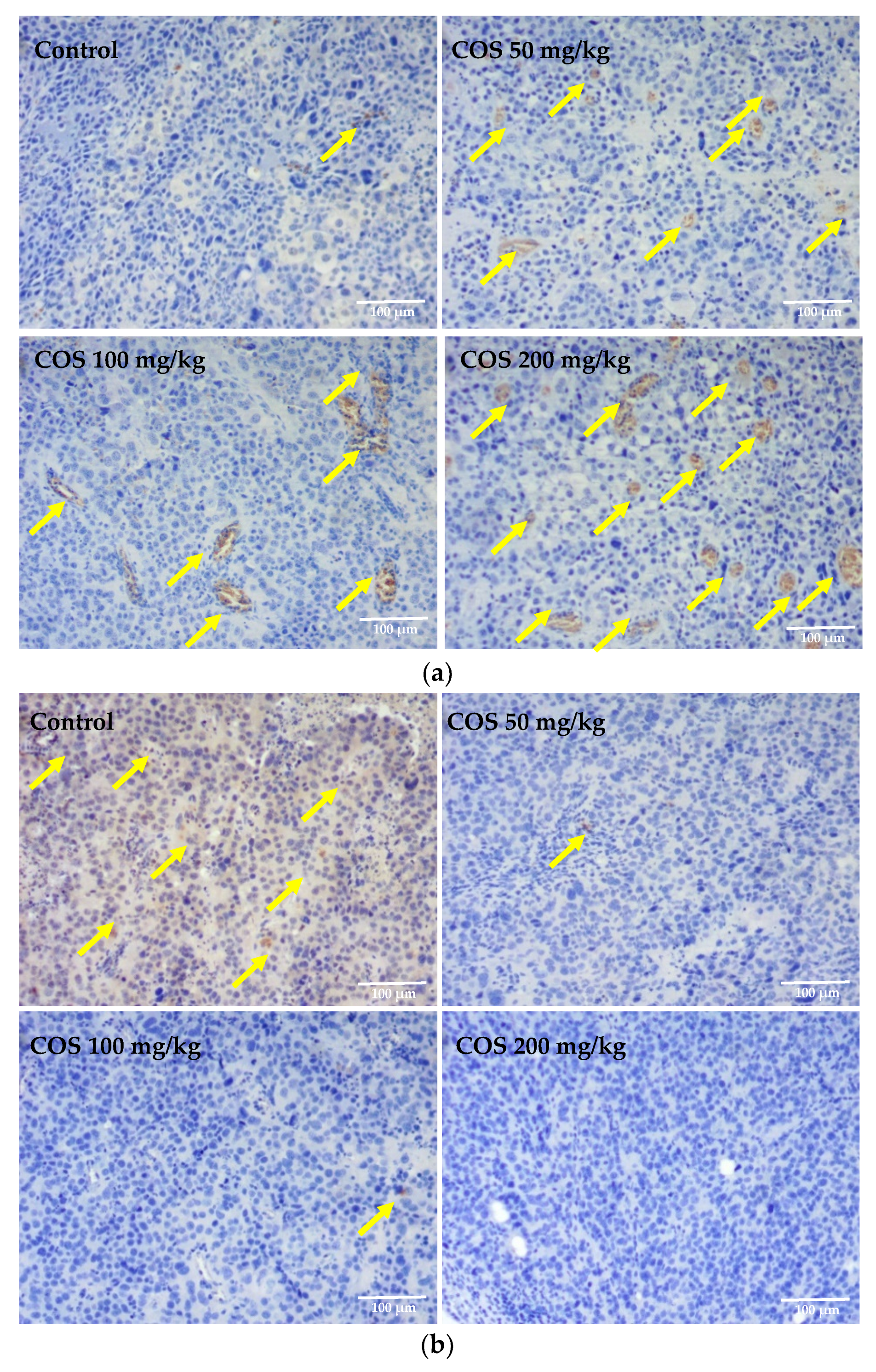
2.5. Effects of COS on Histopathological Changes of Tumor and Lung Tissue
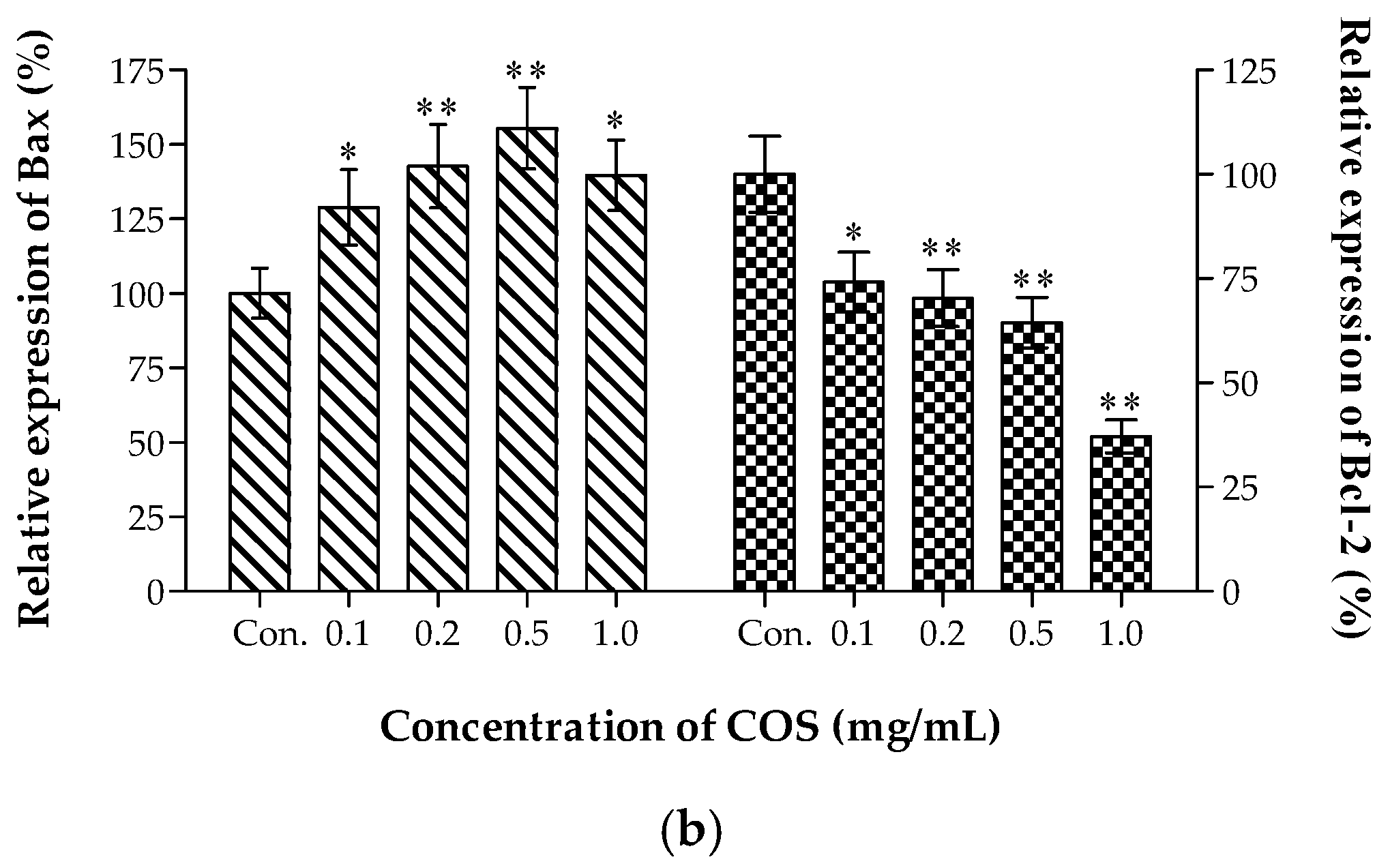
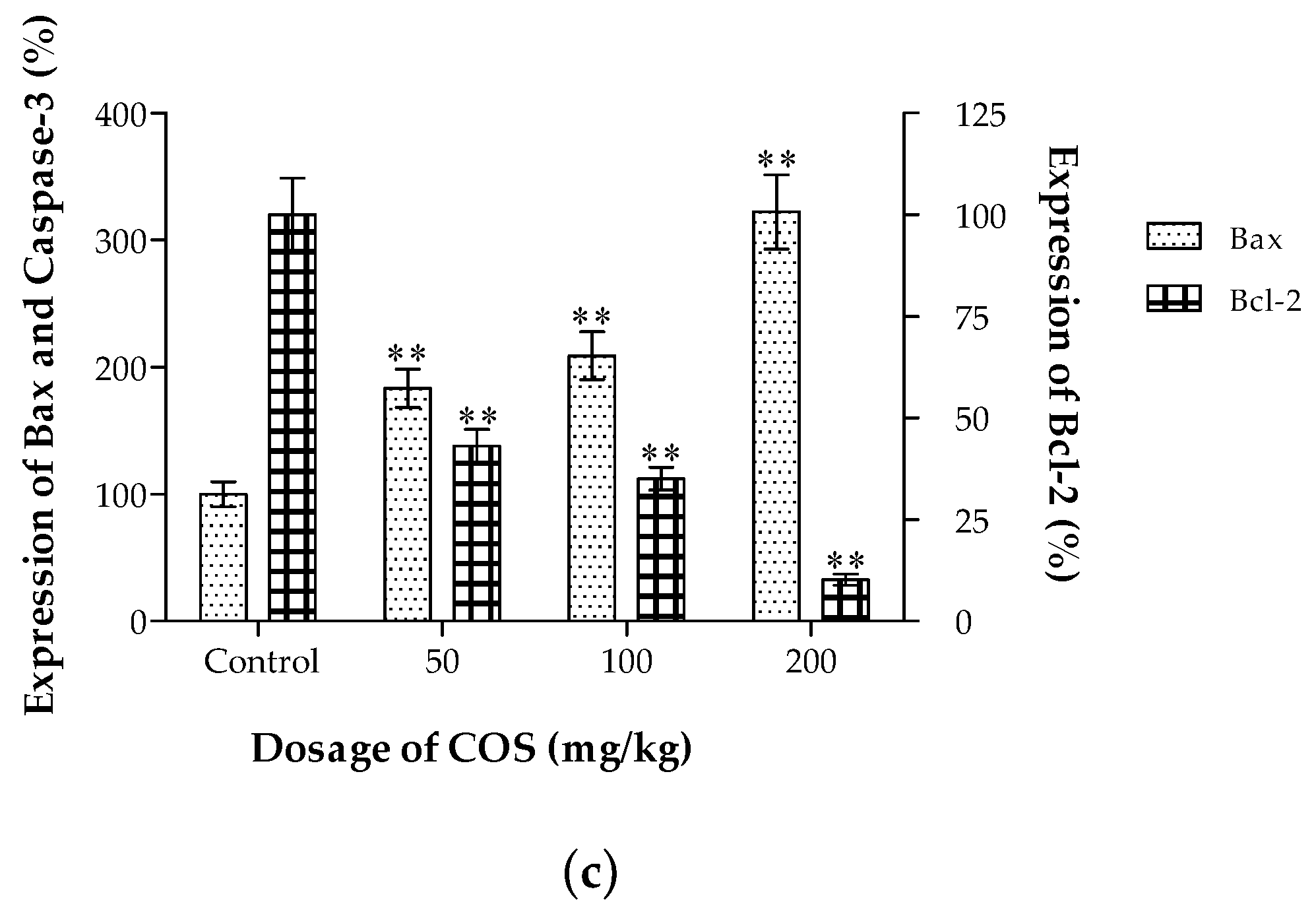
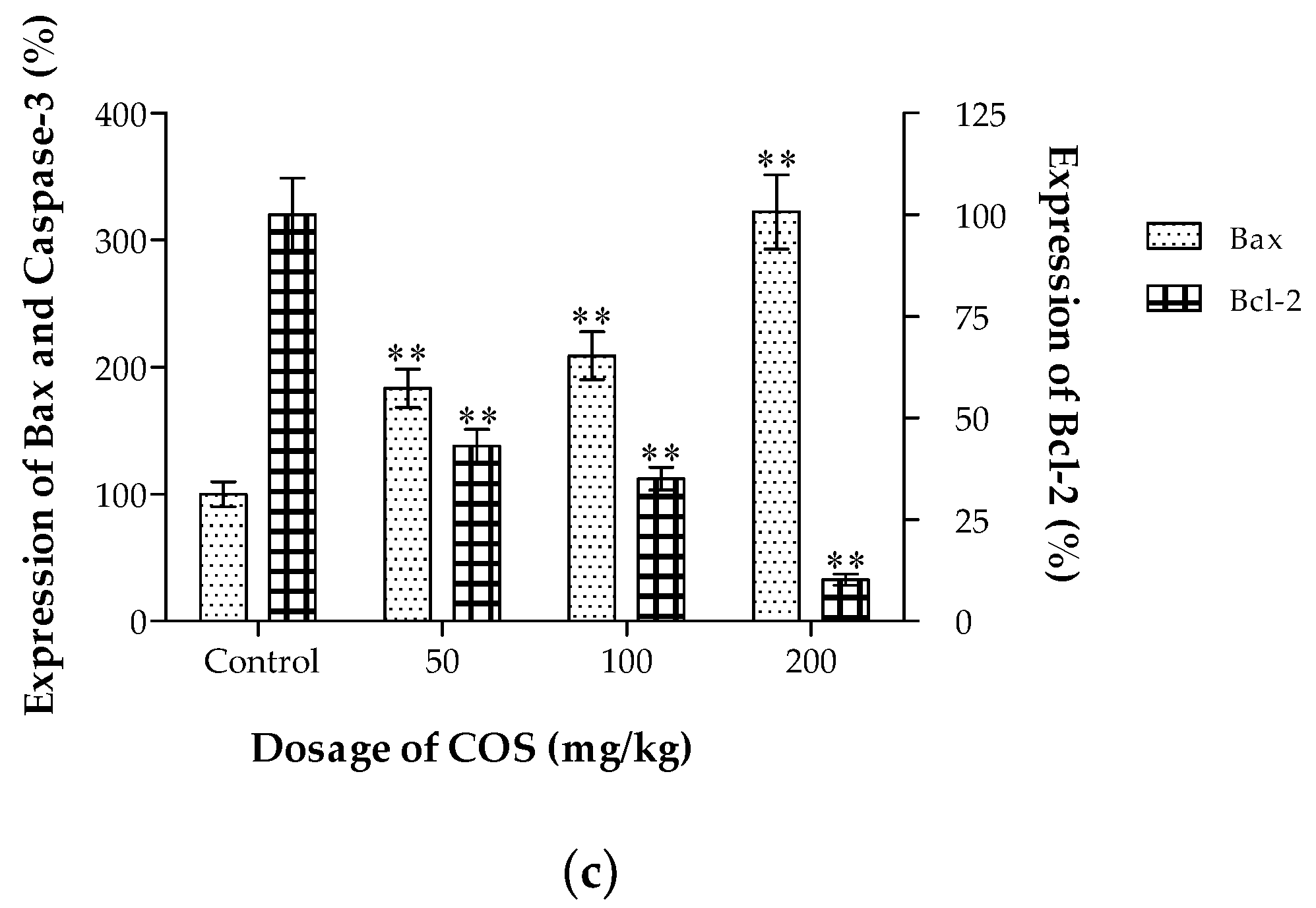
2.6. Effects of COS on the Expression of Bax and Bcl-2 In Vivo
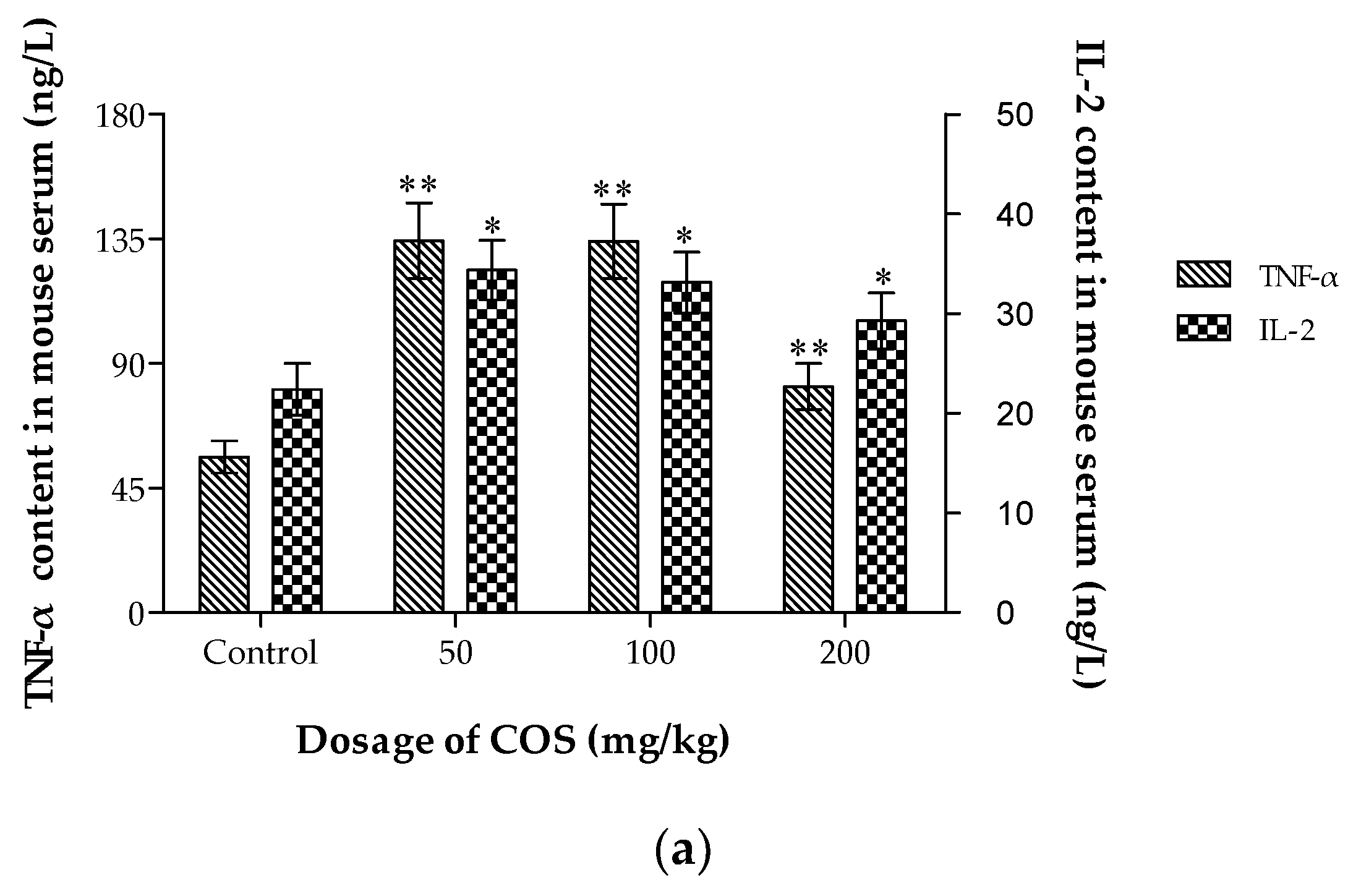
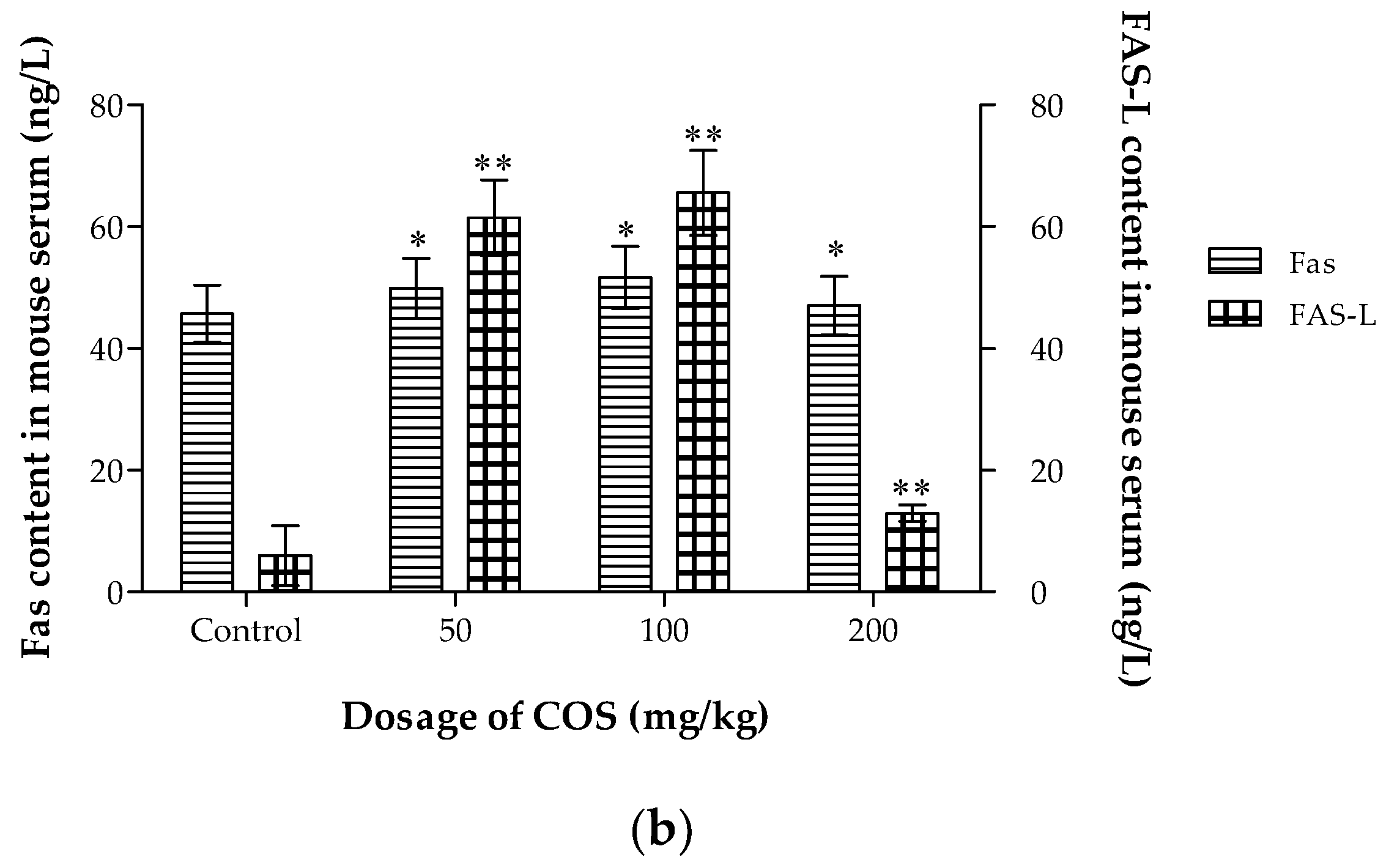
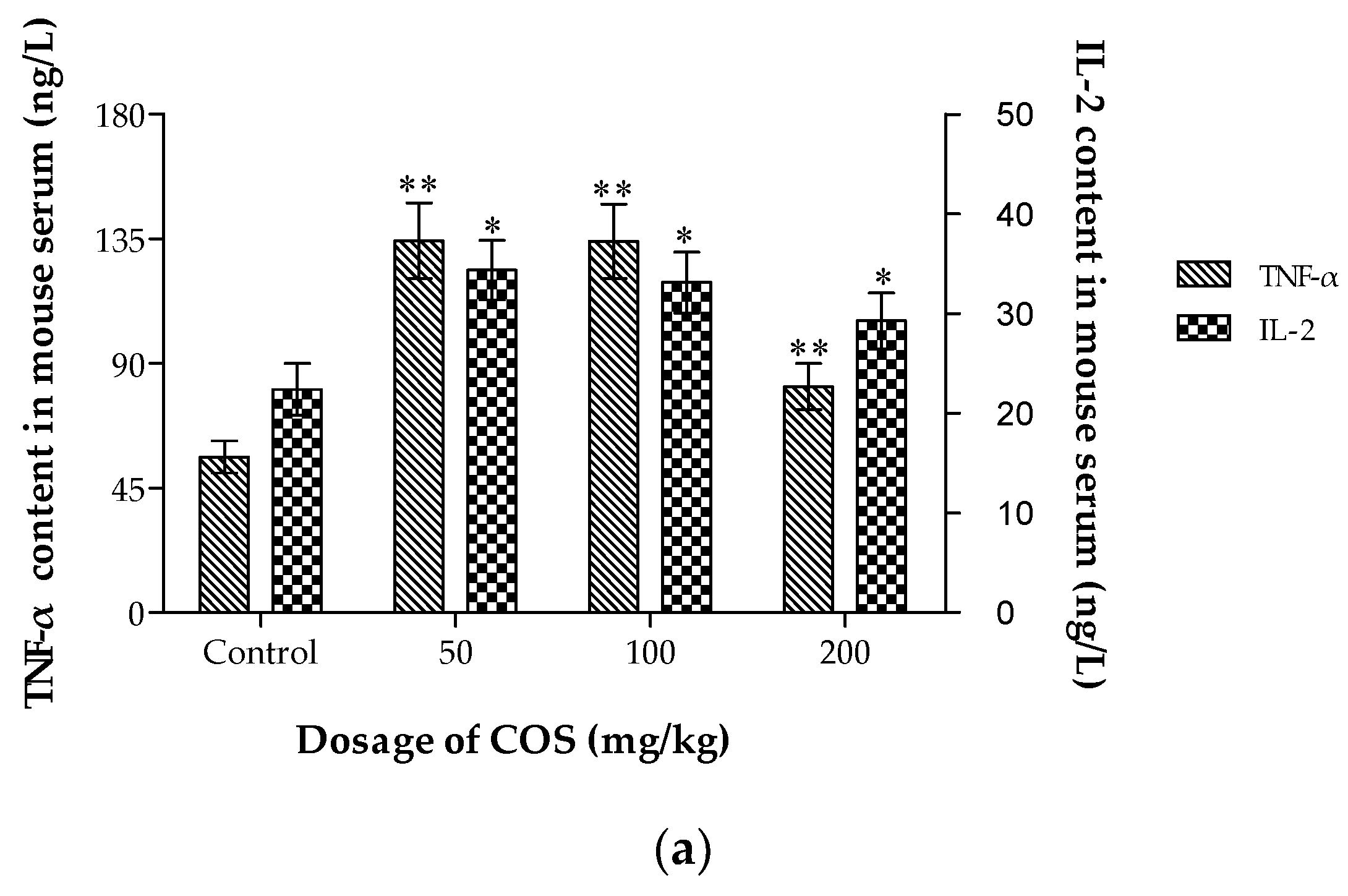
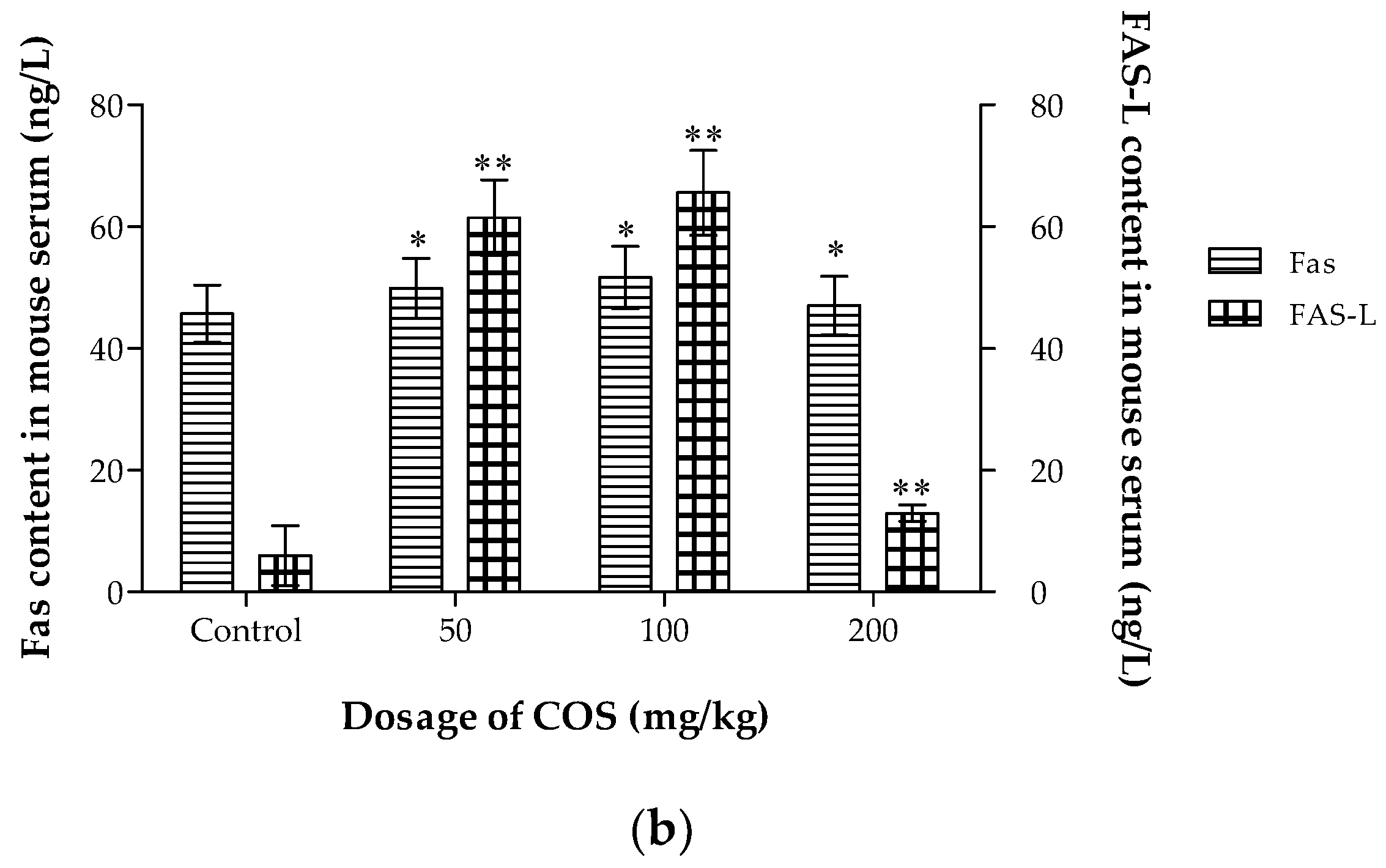
2.7. Effects of COS on TNF-α, IL-2, Fas and Fas-L Levels in Serum
3. Materials and Methods
3.1. Materials and Reagents
3.2. Cell Lines and Animals
3.3. Synthesis and Characterization of Chitosan Oligosaccharide
3.4. Cell Proliferation and Viability Assays In Vitro
3.5. Live-Dead Cell Staining Assay and Apoptosis Analysis
3.6. Anti-Tumor Activities of COS in Lewis Tumor-Bearing Mice
3.7. Histological and Immunohistochemical Analysis
3.8. Measurement of TNF-α, IL-2, Fas and Fas-L Levels in Serum
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, C.; Zhang, Y.; Gao, Y.; Xu, Q.; Ju, X.; Wang, L. Identification and anti-tumor activities of phenolic compounds isolated from defatted adlay (coix lachryma-jobi, l., var. ma-yuen stapf) seed meal. J. Funct. Foods 2016, 26, 394–405. [Google Scholar] [CrossRef]
- Baig, M.H.; Adil, M.; Khan, R.; Dhadi, S.; Ahmad, K.; Rabbani, G.; Bashir, T.; Imran, M.A.; Husain, F.M.; Lee, E.J.; et al. Enzyme targeting strategies for prevention and treatment of cancer: Implications for cancer therapy. Semin. Cancer Biol. 2017, 30101–30103. [Google Scholar] [CrossRef] [PubMed]
- Mou, H.; Zheng, Y.; Zhao, P.; Bao, H.; Fang, W.; Xu, N. Celastrol induces apoptosis in non-small-cell lung cancer a549 cells through activation of mitochondria- and fas/fasl-mediated pathways. Toxicol. In Vitro 2011, 25, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Q.; Shu, Y.; Wang, H.; Zheng, Z.; Wang, J.; Wang, K. Induction of apoptosis in s180 tumor bearing mice by polysaccharide from lentinus edodes via mitochondria apoptotic pathway. J. Funct. Foods 2015, 15, 151–159. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Zhou, F.; Doughty, A.; Hoover, A.R.; Nordquist, R.E.; Chen, W.R. Nanotechnology-based photoimmunological therapies for cancer. Cancer Lett. 2019, 442, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Sethi, G. Bioactive natural products in cancer prevention and therapy: Progress and promise. Semin. Cancer Biol. 2016, 40, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.T.; Luo, M.N.; Li, C.C. Chitosan oligosaccharides reduce acetaminophen-induced hepatotoxicity by suppressing cyp-mediated bioactivation. J. Funct. Foods 2015, 12, 262–270. [Google Scholar] [CrossRef]
- Jiang, Z.; Li, Z.; Zhang, W.; Yang, Y.; Han, B.; Liu, W.; Peng, Y. Dietary Natural N-Acetyl-d-Glucosamine Prevents Bone Loss in Ovariectomized Rat Model of Postmenopausal Osteoporosis. Molecules 2018, 23, 2302. [Google Scholar] [CrossRef] [PubMed]
- Sashiwa, H.; Aiba, S.I. Chemically modified chitin and chitosan as biomaterials. Prog. Polym. Sci. 2004, 29, 887–908. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Regenstein, J.M. Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): A review. Trend. Food Sci. Technol. 2016, 48, 40–50. [Google Scholar] [CrossRef]
- Lodhi, G.; Kim, Y.S.; Hwang, J.W.; Kim, S.K.; Jeon, Y.J.; Je, J.Y.; Ahn, C.B.; Moon, S.H.; Jeon, B.T.; Park, P.J. Chitooligosaccharide and its derivatives: Preparation and biological applications. BioMed. Res. Int. 2014, 2014, 654913. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xia, W.; Liu, P.; Cheng, Q.; Tahirou, T.; Gu, W.; Li, B. Chitosan modification and pharmaceutical/biomedical applications. Mar. Drugs 2010, 8, 1962–1987. [Google Scholar] [CrossRef] [PubMed]
- Mourya, V.K.; Inamdar, N.N.; Choudhari, Y.M. Chitooligosaccharides: Synthesis, characterization and applications. Polym. Sci. Ser. A 2011, 53, 583–612. [Google Scholar] [CrossRef]
- Wu, G.J.; Wu, C.H.; Tsai, G.J. Chitooligosaccharides from the shrimp chitosan hydrolysate induces differentiation of murine raw264.7 macrophages into dendritic-like cells. J. Funct. Foods 2015, 12, 70–79. [Google Scholar] [CrossRef]
- Xie, C.; Xin, W.; Long, C.; Wang, Q.; Fan, Z.; Li, S.; Yin, Y. Chitosan oligosaccharide affects antioxidant defense capacity and placental amino acids transport of sows. BMC Vet. Res. 2016, 12, 243–250. [Google Scholar] [CrossRef]
- Qiao, Y.; Bai, X.F.; Du, Y.G. Chitosan oligosaccharides protect mice from lps challenge by attenuation of inflammation and oxidative stress. Int. Immunopharm. 2011, 11, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.S.; Shon, Y.H. Suppression of metastasis of human breast cancer cells by chitosan oligosaccharides. J. Microbiol. Biotechnol. 2009, 19, 629–633. [Google Scholar]
- Kumar, S.G.; Rahman, M.A.; Lee, S.H.; Hwang, H.S.; Kim, H.A.; Yun, J.W. Plasma proteome analysis for anti-obesity and anti-diabetic potentials of chitosan oligosaccharides in ob/ob mice. Proteomics 2009, 9, 2149–2162. [Google Scholar] [CrossRef]
- Yin, H.; Zhao, X.; Du, Y. Oligochitosan: A plant diseases vaccine-a review. Carbohyd. Polym. 2010, 82, 1–8. [Google Scholar] [CrossRef]
- Choi, E.H.; Yang, H.P.; Chun, H.S. Chitooligosaccharide ameliorates dietinduced obesity in mice and affects adipose gene expression involved in adipogenesis and inflammation. Nutr. Res. 2012, 32, 218–228. [Google Scholar] [CrossRef]
- Guminska, M.; Ignacak, J.; Wojcik, E. In vitro inhibitory effect of chitosan and its degradation products on energy metabolism in Ehrlich ascites tumour cells (EAT). Pol. J. Pharmacol. 1996, 48, 495–501. [Google Scholar] [PubMed]
- Hasegawa, M.; Yagi, K.; Iwakawa, S.; Hirai, M. Chitosan induces apoptosis via caspase-3 activation in bladder tumor cells. Jpn. J. Cancer Res. 2001, 92, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Mendis, E.; Rajapakse, N.; Kim, S.K. Strong electronic charge as an important factor for anticancer activity of chitooligosaccharides (COS). Life Sci. 2006, 78, 2399–2408. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Song, W.; Cao, D.; Liu, X.; Jia, Y. Growth inhibition and apoptosis of human leukemia K562 cells induced by seleno-short-chain chitosan. Methods Find Exp. Clin. Pharmacol. 2008, 30, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.S.; Kim, M.K.; Shon, Y.H. Chemopreventive effect of chitosan oligosaccharide against colon carcinogenesis. J. Microbiol. Biotechnol. 2007, 17, 1546–1549. [Google Scholar] [PubMed]
- Park, J.K.; Chung, M.J.; Choi, H.N.; Park, Y.I. Effects of the molecular weight and the degree of deacetylation of chitosan oligosaccharides on antitumor activity. Int. J. Mol. Sci. 2011, 12, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.T.; Chen, M.H.; Chan, H.Y.; Jeng, J.H.; Wang, Y.J. Inhibitory effects of chitooligosaccharides on tumor growth and metastasis. Food Chem. Toxicol. 2009, 47, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.; Sereno, J.; Garrido, P.; Parada, B.; Cunha, M.F.; Reis, F.; Pintado, M.E.; Santos-Silva, A. Inhibition of bladder tumor growth by chitooligosaccharides in an experimental carcinogenesis model. Mar. Drugs 2012, 10, 2661–2675. [Google Scholar] [CrossRef]
- Maeda, Y.; Kimura, Y. Antitumor effects of various low-molecular-weight chitosans are due to increased natural killer activity of intestinal intraepithelial lymphocytes in sarcoma 180-bearing mice. J. Nutr. 2004, 134, 945–950. [Google Scholar] [CrossRef]
- Mattaveewong, T.; Wongkrasant, P.; Chanchai, S.; Pichyangkura, R.; Chatsudthipong, V.; Muanprasat, C. Chitosan oligosaccharide suppresses tumor progression in a mouse model of colitis-associated colorectal cancer through AMPK activation and suppression of NF-kappaB and mTOR signaling. Carbohydr. Polym. 2016, 145, 30–36. [Google Scholar] [CrossRef]
- Muanprasat, C.; Wongkrasant, P.; Satitsri, S.; Moonwiriyakit, A.; Pongkorpsakol, P.; Mattaveewong, T.; Pichyangkura, R.; Chatsudthipong, V. Activation of AMPK by chitosan oligosaccharide in intestinal epithelial cells: Mechanism of action and potential applications in intestinal disorders. Biochem. Pharmacol. 2015, 96, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Je, J.Y.; Lee, S.J.; Kim, Y.S.; Hwang, J.W.; Sung, S.H.; Moon, S.H.; Jeon, B.T.; Kim, S.K.; Jeon, Y.J.; Park, P.J. Chitooligosaccharides induce apoptosis in human myeloid leukemia hl-60 cells. Bioorg. Med. Chem. Lett. 2012, 22, 6136–6138. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Shi, Y.; Wang, S.; Yang, Y.; Han, B.; Liu, W.; Jones, D.N.; Liu, W. Structural and biochemical insights into the degradation mechanism of chitosan by chitosanase ou01. BBA Gen. Subj. 2015, 1850, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Wang, S.; Xu, W.; Han, B.; Liu, W.; Jones, D.N.; Liu, W. Structural insights into the substrate-binding mechanism for a novel chitosanase. Biochem. J. 2015, 461, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Aam, B.B.; Heggset, E.B.; Norberg, A.L.; Sørlie, M.; Vårum, K.M.; Eijsink, V.G. Production of chitooligosaccharides and their potential applications in medicine. Mar. Drugs 2010, 8, 1482–1517. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Liu, P.; Zhang, J.; Chen, J. Biological activities of chitosan and chitooligosaccharides. Food Hydrocoll. 2011, 25, 170–179. [Google Scholar] [CrossRef]
- Jiang, Y.; Fu, C.; Wu, S.; Liu, G.; Guo, J.; Su, Z. Determination of the deacetylation degree of chitooligosaccharides. Mar. Drugs 2017, 15, 332. [Google Scholar] [CrossRef]
- Liaqat, F.; Eltem, R. Chitooligosaccharides and their biological activities: A comprehensive review. Carbohyd. Polym. 2018, 184, 243. [Google Scholar] [CrossRef]
- Yin, X.; Zhou, J.; Jie, C.; Xing, D.; Zhang, Y. Anticancer activity and mechanism of scutellaria barbata extract on human lung cancer cell line A549. Life Sci. 2004, 75, 2233–2244. [Google Scholar] [CrossRef]
- Huang, S.T.; Yang, R.C.; Yang, L.J.; Lee, P.N.; Pang, J.H.S. Phyllanthus urinaria, triggers the apoptosis and bcl-2 down-regulation in lewis lung carcinoma cells. Life Sci. 2003, 72, 1705–1716. [Google Scholar] [CrossRef]
- Thuret, G.; Pipparelli, A.; Duband, S.; Piselli, S.; Muraine, M.; Peoc’H, M.; Acquart, S.; Zhao, M.; Prades, J.M.; Gain, P.G. Corneal graft endothelial viability assessment using the triple labeling hoechst/ethidium homodimer/calcein-am: Technical improvements usind 3d microscopy. Acta Ophthalmol. 2010, 85, s240. [Google Scholar] [CrossRef]
- Ashkenazi, A.; Fairbrother, W.J.; Leverson, J.D.; Souers, A.J. From basic apoptosis discoveries to advanced selective bcl-2 family inhibitors. Nat. Rev. Drug. Discov. 2017, 16, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Alves, S.; Neiri, L.; Chaves, S.R.; Vieira, S.; Trindade, D.; Manon, S.; Dominguez, V.; Pintado, B.; Jonckheere, V.; Van Damme, P.; Silva, R.D.; Aldabe, R.; Côrte-Real, M. N-terminal acetylation modulates bax targeting to mitochondria. Int. J. Biochem. Cell B. 2018, 95, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.H.; Ho, Y.S.; Lai, C.S.; Hsieh, S.C.; Chen, L.H.; Lin, E.; Ho, C.T.; Pan, M.H. Hexahydro-β-acids potently inhibit 12-O-tetradecanoylphorbol 13-acetate-induced skin inflammation and tumor promotion in mice. J. Agric. Food Chem. 2013, 61, 11541–11549. [Google Scholar] [CrossRef] [PubMed]
- Reguiga, M.B.; Bouquet, C.; Farinotti, R.; Bonhomme-Faivre, L. Interferon-alpha improves docetaxel antitumoral and antimetastatic efficiency in lewis lung carcinoma bearing mice. Life Sci. 2012, 91, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Janker, F.; Weder, W.; Jang, J.H.; Jungraithmayr, W. Preclinical, non-genetic models of lung adenocarcinoma: A comparative survey. Oncotarget 2018, 9, 30527–30538. [Google Scholar] [CrossRef] [PubMed]
- Kokawa, K.; Shikone, T.; Otani, T.; Nakano, R. Apoptosis and the expression of bax and bcl-2 in squamous cell carcinoma and adenocarcinoma of the uterine cervix. Cancer 2015, 85, 1799–1809. [Google Scholar] [CrossRef]
- Zhu, L.; Han, M.B.; Gao, Y.; Wang, H.; Dai, L.; Wen, Y.; Na, L.X. Curcumin triggers apoptosis via upregulation of bax/bcl-2 ratio and caspase activation in sw872 human adipocytes. Mol. Med. Rep. 2015, 12, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Placzek, W.J.; Wei, J.; Kitada, S.; Zhai, D.; Reed, J.C.; Pellecchia, M. A survey of the anti-apoptotic bcl-2 subfamily expression in cancer types provides a platform to predict the efficacy of bcl-2 antagonists in cancer therapy. Cell Death Dis. 2010, 1, e40. [Google Scholar] [CrossRef]
- Xie, Z.; Xiao, Z.; Wang, F. Hepatitis C Virus Nonstructural 5A Protein (HCV-NS5A) Inhibits Hepatocyte Apoptosis through the NF-κb/miR-503/bcl-2 Pathway. Mol. Cells 2017, 40, 202–210. [Google Scholar]
- Sowdhamini, R.; Perumalsamy, L.R.; Vaigundan, D.; Caroline, K.; Neha, P.; Apurva, S. The n-terminus and alpha-5, alpha-6 helices of the pro-apoptotic protein bax, modulate functional interactions with the anti-apoptotic protein Bcl-xl. BMC Cell Biol. 2007, 8, 16. [Google Scholar]
- Ohno, T.; Nakano, T.; Niibe, Y.; Tsujii, H.; Oka, K. Bax protein expression correlates with radiation-induced apoptosis in radiation therapy for cervical carcinoma. Cancer 2015, 83, 103–110. [Google Scholar] [CrossRef]
- Benhur, H.; Gurevich, P.; Huszar, M.; Benarie, A.; Berman, V.; Tendler, Y.; Zinder, O.; Tchanishev, R.; Gershon, S.; Mor, G.; et al. Apoptosis and apoptosis-related proteins (fas, fas ligand, blc-2, p53) in lymphoid elements of human ovarian tumors. Eur. J. Gynaecol. Oncol. 2000, 21, 53–57. [Google Scholar]
- Chang, L.; Kamata, H.; Solinas, G.; Luo, J.; Maeda, S.; Venuprasad, K.; Liu, Y.C.; Karin, M. The E3 Ubiquitin Ligase Itch Couples JNK Activation to TNFα-induced Cell Death by Inducing c-FLIPL Turnover. Cell 2006, 124, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Jackaman, C.; Bundell, C.S.; Kinnear, B.F.; Smith, A.M.; Filion, P.; Van, H.D.; Robinson, B.W.; Nelson, D.J. Il-2 intratumoral immunotherapy enhances cd8+ t cells that mediate destruction of tumor cells and tumor-associated vasculature: A novel mechanism for il-2. J. Immunol. 2003, 171, 5051–5063. [Google Scholar] [CrossRef]
- Zhang, B.; Sun, T.; Xue, L.; Han, X.; Zhang, B.; Lu, N.; Shi, Y.; Tan, W.; Zhou, Y.; Zhao, D.; et al. Functional polymorphisms in fas and fasl contribute to increased apoptosis of tumor infiltration lymphocytes and risk of breast cancer. Carcinogenesis 2007, 28, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Suda, T.; Takahashi, T.; Golstein, P.; Nagata, S. Molecular cloning and expression of the fas ligand, a novel member of the tumor necrosis factor family. Cell 1993, 75, 1169–1178. [Google Scholar] [CrossRef]
- Beidler, D.R.; Tewari, M.; Friesen, P.D.; Poirier, G.; Dixit, V.M. The baculovirus p35 protein inhibits fas- and tumor necrosis factor-induced apoptosis. J. Biol. Chem. 1995, 270, 16526–16528. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, W.; Peng, Y.; Han, B.; Yang, Y. Toll like receptor 4 (TLR4) mediates the stimulating activities of chitosan oligosaccharide on macrophages. Int. Immunopharm. 2014, 23, 254–261. [Google Scholar] [CrossRef]
- Jiang, Z.; Han, B.; Liu, W.; Peng, Y. Evaluation on biological compatibility of carboxymethyl chitosan as biomaterials for antitumor drug delivery. J. Biomater. Appl. 2017, 31, 985–994. [Google Scholar] [CrossRef]
- Wang, X.; Chang, Q.; Wang, Y.; Su, F.; Zhang, S. Late-onset temperature reduction can retard the aging process in aged fish via a combined action of an anti-oxidant system and the insulin/insulin-like growth factor 1 signaling pathway. Rejuv. Res. 2014, 17, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Han, B.; Li, H.; Yang, Y.; Liu, W. Carboxymethyl chitosan represses tumor angiogenesis in vitro and in vivo. Carbohyd. Polym. 2015, 129, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Sun, Y.; Zheng, S.L.; Qin, Y.; Mcclements, D.J.; Hu, J.N.; Deng, Z. Antitumor and immunomodulatory effects of ginsenoside rh2 and its octyl ester derivative in h22 tumor-bearing mice. J. Funct. Foods 2017, 32, 382–390. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group and Dosage | Weight of Tumor (g) | Inhibition on Tumor Weight |
|---|---|---|
| Control | 1.71 ± 0.24 | – |
| COS (50 mg/kg) | 1.11 ± 0.37 * | 35.32% |
| COS (100 mg/kg) | 0.81 ± 0.18 ** | 52.53% |
| COS (200 mg/kg) | 0.88 ± 0.21 ** | 48.44% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.; Li, H.; Qiao, J.; Yang, Y.; Wang, Y.; Liu, W.; Han, B. Potential Analysis and Preparation of Chitosan Oligosaccharides as Oral Nutritional Supplements of Cancer Adjuvant Therapy. Int. J. Mol. Sci. 2019, 20, 920. https://doi.org/10.3390/ijms20040920
Jiang Z, Li H, Qiao J, Yang Y, Wang Y, Liu W, Han B. Potential Analysis and Preparation of Chitosan Oligosaccharides as Oral Nutritional Supplements of Cancer Adjuvant Therapy. International Journal of Molecular Sciences. 2019; 20(4):920. https://doi.org/10.3390/ijms20040920
Chicago/Turabian StyleJiang, Zhiwen, Hui Li, Jing Qiao, Yan Yang, Yanting Wang, Wanshun Liu, and Baoqin Han. 2019. "Potential Analysis and Preparation of Chitosan Oligosaccharides as Oral Nutritional Supplements of Cancer Adjuvant Therapy" International Journal of Molecular Sciences 20, no. 4: 920. https://doi.org/10.3390/ijms20040920
APA StyleJiang, Z., Li, H., Qiao, J., Yang, Y., Wang, Y., Liu, W., & Han, B. (2019). Potential Analysis and Preparation of Chitosan Oligosaccharides as Oral Nutritional Supplements of Cancer Adjuvant Therapy. International Journal of Molecular Sciences, 20(4), 920. https://doi.org/10.3390/ijms20040920