Long Non-Coding RNA and Acute Leukemia

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Genetic Features of Acute Leukemia
3. LncRNAs Characteristics
4. Biogenesis and Classification
5. LncRNAs in Normal Hematopoiesis
6. LncRNAs in Acute Leukemia
7. LncRNAs in Acute Myeloid Leukemia
8. LncRNAs in Acute Lymphoblastic Leukemia
9. Future Outlooks: Potential Clinical Implications on LncRNAs in Acute Leukemia
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 1. ABL1 | 2. ABL protoconcogene 1 |
| 3. ABL2 | 4. ABL protooncogene 2 |
| 5. AL | 6. Acute leukemia |
| 7. ALL | 8. Acute lymphoblastic leukemia |
| 9. AML | 10. Acute myeloblastic leukemia |
| 11. ANRIL | 12. Antisense non-coding RNA in the INK4-ARF locus B-ALL B cell Acute lymphoblastic leukemia |
| 13. ARID1A | 14. AT-rich interaction domain 1A |
| 15. AURKA | 16. Aurora kinase A gene |
| 17. BALR | 18. B-ALL-associated long non-coding RNAs BL Burkitt Lymphoma |
| 19. CAS9 | 20. CRISPR associated protein 9 |
| 21. CBF | 22. Core-binding factor subunit beta |
| 23. CCAT1 | 24. Colon cancer associated transcript 1 ceRNA Competing endogenous RNA |
| 25. CDKN2A | 26. Cyclin dependent kinase inhibitor 2A |
| 27. CDKN2B | 28. Cyclin dependent kinase inhibitor 2B |
| 29. CDKN2B-AS1 | 30. CDKN2B antisense RNA 1 |
| 31. CEBPA | 32. CCAAT enhancer binding protein alpha |
| 33. CHD6 | 34. Chromodomain helicase DNA binding protein 6 |
| 35. circRNA | 36. Circular RNA |
| 37. CLL | 38. Chronic lymphocytic leukemia |
| 39. CML | 40. Chronic myeloblastic leukemia |
| 41. CN | 42. Cytogenetically normal |
| 43. COLDAIR | 44. COLD assisted intronic non-coding RNA |
| 45. CREBBP | 46. CREB binding protein |
| 47. CRISPR | 48. Clustered regularly interspaced short palindromic repeats |
| 49. CRLF2 | 50. Cytokine receptor like factor 2 |
| 51. CSF1R | 52. Colony stimulating factor 1 receptor |
| 53. DCs | 54. Dendritic Cells |
| 55. DNMT3A | 56. DNA methyltransferase 3α |
| 57. EBF1 | 58. Early B cell factor 1 |
| 59. EGO | 60. Eosinophil granule ontogeny |
| 61. EP300 | 62. E1A binding protein P300 |
| 63. eRNAs | 64. Enhancer RNAs |
| 65. ETS1 | 66. ETS proto-oncogene 1 transcription factor |
| 67. ETV6 | 68. ETS Variant6 |
| 69. FLC | 70. Flowering repressor locus |
| 71. FLT3 | 72. Fms related tyrosine kinase 3 |
| 73. fRNAs | 74. Functional RNAs |
| 75. GAS5 | 76. Growth specific 5 |
| 77. GEO | 78. Gene expression omnibus |
| 79. H19 | 80. Imprinted maternally expressed transcript |
| 81. hnRNP | 82. Heterogenous nuclear ribonucleoprotein |
| 83. HOTAIR | 84. The HOX transcript antisense intergenic RNA |
| 85. HOTTIP | 86. HOXA distal transcript antisense RNA |
| 87. IGFR1 | 88. Insuline-like growth factor type 1 |
| 89. IKZF1 | 90. IKAROS family zinc finger 1 |
| 91. IRAIN | 92. IGFR1 antisense imprinted non protein RNA |
| 93. ITPR1 | 94. Inositol1,4,5-triophosphate receptor type 1 |
| 95. JAK2 | 96. Janus kinase 2 |
| 97. KIT | 98. Tyrosine protein kinase |
| 99. LincRNA | 100. Long intergenic non-coding RNA |
| 101. LncRNA | 102. Long non-coding RNA |
| 103. lnc-DC | 104. Dendritic cell-specifict lncRNA |
| 105. lincRNA-p21 | 106. Large intergenic non-coding RNA p21 |
| 107. lncRNA | 108. Long non-coding RNA |
| 109. LUNAR1 | 110. Leukemia-associated non-coding IGF1R |
| 111. MALAT1 | 112. Metastasis associated lung adenocarcinoma transcript 1 MCL Mantle cell lymphoma |
| 113. MEG3 | 114. Maternally expressed 3 |
| 115. miRNA | 116. MicroRNA |
| 117. mRNA | 118. Messenger RNA |
| 119. NCBI | 120. National center of biotechnology information |
| 121. ncRNA | 122. Non-coding RNA |
| 123. NFAT | 124. Nuclear factor activated T cells |
| 125. nmRNA | 126. Non messengers RNA |
| 127. npcRNA | 128. Non protein-coding RNA |
| 129. NRAS | 130. NRAS proto-oncogene |
| 131. NRON | 132. Non-protein-coding RNA Repressor of NFAT |
| 133. NSD2 | 134. Nuclear receptor binding SET domain protein 2 |
| 135. PANDAR | 136. Promoter of CDKN1A antisense DNA damage activated RNA |
| 137. PAX5 | 138. Paired box 5 |
| 139. PBX1 | 140. PBX Homeobox 1 |
| 141. PCGP | 142. Pediatric cancer genome project |
| 143. PDGFRB | 144. Platelet derived growth factor receptor beta |
| 145. piRNAs | 146. PIWI-interacting RNAs |
| 147. PML | 148. Promyelocytic Leukemia gene |
| 149. PROMPTs | 150. Promoter-associated long RNAs |
| 151. RB1 | 152. RB transcriptional corepressor 1 |
| 153. RBPs | 154. RNA-binding proteins |
| 155. RUNX1 | 156. Runt related transcription factor 1 |
| 157. SHP1 | 158. Scr homology region 2 domain containing phosphatase-1 |
| 159. siRNA | 160. Small interference RNA |
| 161. snRNAs | 162. Small nuclear RNA |
| 163. snoRNAs | 164. Small nucleolar RNA |
| 165. STAT3 | 166. Signal transducer and activator of transcription 3 |
| 167. TCF3 | 168. Transcription Factor 3Ç |
| 169. TERC | 170. Telomerase RNA component |
| 171. TET2 | 172. Tet methylcytosine dioxygenase 2 |
| 173. TLR | 174. Tool-like receptor |
| 175. TP53 | 176. Tumor protein P53 |
| 177. TP53COR1 | 178. Tumor protein P53 pathway corepressor 1 |
| 179. TUG1 | 180. Taurine-up regulated gene 1 |
| 181. UCA1 | 182. Urothelial carcinoma associated 1 |
| 183. vlincRNA | 184. Very long intergenic RNA |
| 185. XIST | 186. X inactive specific transcript |
References
- Mejia-Arangure, J.M.; McNally, R.J.Q. Acute Leukemia in Children. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Linet, M.S.; Brown, L.M.; Mbulaiteye, S.M.; Check, D.; Ostroumova, E.; Landgren, A.; Devesa, S.S. International long-term trends and recent patterns in the incidence of leukemias and lymphomas among children and adolescents ages 0–19 years. Int. J. Cancer 2016, 138, 1862–1874. [Google Scholar] [CrossRef] [PubMed]
- Schuz, J.; Erdmann, F. Environmental Exposure and Risk of Childhood Leukemia: An Overview. Arch. Med. Res. 2016, 47, 607–614. [Google Scholar] [CrossRef]
- Beltran-Anaya, F.O.; Cedro-Tanda, A.; Hidalgo-Miranda, A.; Romero-Cordoba, S.L. Insights into the Regulatory Role of Non-coding RNAs in Cancer Metabolism. Front. Physiol. 2016, 7, 342. [Google Scholar] [CrossRef] [PubMed]
- Perez-Saldivar, M.L.; Fajardo-Gutierrez, A.; Bernaldez-Rios, R.; Martinez-Avalos, A.; Medina-Sanson, A.; Espinosa-Hernandez, L.; Flores-Chapa, J.D.; Amador-Sanchez, R.; Penaloza-Gonzalez, J.G.; Alvarez-Rodriguez, F.J.; et al. Childhood acute leukemias are frequent in Mexico City: Descriptive epidemiology. BMC Cancer 2011, 11, 355. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.; Bruford, E.A. Naming ‘junk’: Human non-protein coding RNA (ncRNA) genome nomenclature. Hum. Genom. 2011, 5, 90–98. [Google Scholar] [CrossRef]
- Connelly, C.M.; Moon, M.H.; Schneekloth, J.S. The Emerging Role of RNA as a Therapeutic Target for Small Molecules. Cell Chem. Biol. 2016, 23, 1077–1090. [Google Scholar] [CrossRef]
- Greaves, M. In utero origins of childhood leukaemia. Early Hum. Dev. 2005, 81, 123–129. [Google Scholar] [CrossRef]
- Mullighan, C.G. Genomic profiling of B-progenitor acute lymphoblastic leukemia. Best Pract. Res. Clin. Haematol. 2011, 24, 489–503. [Google Scholar] [CrossRef]
- Janczar, S.; Janczar, K.; Pastorczak, A.; Harb, H.; Paige, A.J.W.; Zalewska-Szewczyk, B.; Danilewicz, M.; Mlynarski, W. The Role of Histone Protein Modifications and Mutations in Histone Modifiers in Pediatric B-Cell Progenitor Acute Lymphoblastic Leukemia. Cancers 2017, 9, 2. [Google Scholar] [CrossRef]
- Roberts, K.G.; Gu, Z.H.; Payne-Turner, D.; McCastlain, K.; Harvey, R.C.; Chen, I.M.; Pei, D.Q.; Iacobucci, I.; Valentine, M.; Pounds, S.B.; et al. High Frequency and Poor Outcome of Philadelphia Chromosome-Like Acute Lymphoblastic Leukemia in Adults. J. Clin. Oncol. 2017, 35, 394. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, J.; Kiialainen, A.; Karlberg, O.; Berglund, E.C.; Goransson-Kultima, H.; Sonderkaer, M.; Nielsen, K.L.; Gustafsson, M.G.; Behrendtz, M.; Forestier, E.; et al. Digital gene expression profiling of primary acute lymphoblastic leukemia cells. Leukemia 2012, 26, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.K.; Ma, J.; Wang, J.M.; Chen, X.; Gedman, A.L.; Dang, J.J.; Nakitandwe, J.; Holmfeldt, L.; Parker, M.; Easton, J.; et al. The landscape of somatic mutations in infant MLL-rearranged acute lymphoblastic leukemias. Nat. Genet. 2015, 47, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Rastogi, P.; Shah, B.; Zhang, L. B lymphoblastic leukemia/lymphoma: New insights into genetics, molecular aberrations, subclassification and targeted therapy. Oncotarget 2017, 8, 66728–66741. [Google Scholar] [CrossRef] [PubMed]
- Aziz, H.; Ping, C.Y.; Alias, H.; Ab Mutalib, N.S.; Jamal, R. Gene Mutations as Emerging Biomarkers and Therapeutic Targets for Relapsed Acute Myeloid Leukemia. Front. Pharmacol. 2017, 8, 897. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and Functions of Long Noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.T.; Song, X.Y.; Glass, C.K.; Rosenfeld, M.G. The Long Arm of Long Noncoding RNAs: Roles as Sensors Regulating Gene Transcriptional Programs. Cold Spring Harb. Perspect. Biol. 2011, 3, a003756. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; Du, X. The long non-coding RNAs, a new cancer diagnostic and therapeutic gold mine. Mod. Pathol. 2013, 26, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Kapranov, P.; St Laurent, G.; Raz, T.; Ozsolak, F.; Reynolds, C.P.; Sorensen, P.H.B.; Reaman, G.; Milos, P.; Arceci, R.J.; Thompson, J.F.; et al. The majority of total nuclear-encoded non-ribosomal RNA in a human cell is ‘dark matter’ un-annotated RNA. BMC Biol. 2010, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermuller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [PubMed]
- Dieci, G.; Fiorino, G.; Castelnuovo, M.; Teichmann, M.; Pagano, A. The expanding RNA polymerase III transcriptome. Trends Genet. 2007, 23, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.F.; Yang, L.; Zhang, Y.; Xiang, J.F.; Wu, Y.W.; Carmichael, G.G.; Chen, L.L. Long Noncoding RNAs with snoRNA Ends. Mol. Cell 2012, 48, 219–230. [Google Scholar] [CrossRef]
- Hung, T.; Wang, Y.L.; Lin, M.F.; Koegel, A.K.; Kotake, Y.; Grant, G.D.; Horlings, H.M.; Shah, N.; Umbricht, C.; Wang, P.; et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nat. Genet. 2011, 43, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Orom, U.A.; Derrien, T.; Beringer, M.; Gumireddy, K.; Gardini, A.; Bussotti, G.; Lai, F.; Zytnicki, M.; Notredame, C.; Huang, Q.H.; et al. Long Noncoding RNAs with Enhancer-like Function in Human Cells. Cell 2010, 143, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs Are the Predominant Transcript Isoform from Hundreds of Human Genes in Diverse Cell Types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef]
- Yang, L.; Duff, M.O.; Graveley, B.R.; Carmichael, G.G.; Chen, L.L. Genomewide characterization of non-polyadenylated RNAs. Genome Biol. 2011, 12, R16. [Google Scholar] [CrossRef]
- St Laurent, G.; Wahlestedt, C.; Kapranov, P. The Landscape of long noncoding RNA classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef]
- Han, D.; Wang, M.; Ma, N.; Xu, Y.; Jiang, Y.T.; Gao, X. Long noncoding RNAs: Novel players in colorectal cancer. Cancer Lett. 2015, 361, 13–21. [Google Scholar] [CrossRef]
- Di Gesualdo, F.; Capaccioli, S.; Lulli, M. A pathophysiological view of the long non-coding RNA world. Oncotarget 2014, 5, 10976–10996. [Google Scholar] [CrossRef]
- Morlando, M.; Ballarino, M.; Fatica, A. Long Non-Coding RNAs: New Players in Hematopoiesis and Leukemia. Front. Med. 2015, 2, 23. [Google Scholar] [CrossRef] [PubMed]
- Ulitsky, I.; Bartel, D.P. lincRNAs: Genomics, Evolution, and Mechanisms. Cell 2013, 154, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.; Lee, J.T. YY1 Tethers Xist RNA to the Inactive X Nucleation Center. Cell 2011, 146, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Binder, J.; Frankild, S.; Tsafou, K.; Stolte, C.; O’Donoghue, S.; Schneider, R.; Jensen, L.J. COMPARTMENTS. Available online: https://compartments.jensenlab.org/Search (accessed on 11 July 2018).
- Fang, K.; Han, B.W.; Chen, Z.H.; Lin, K.Y.; Zeng, C.W.; Li, X.J.; Li, J.H.; Luo, X.Q.; Chen, Y.Q. A distinct set of long non-coding RNAs in childhood MLL-rearranged acute lymphoblastic leukemia: Biology and epigenetic target. Hum. Mol. Genet. 2014, 23, 3278–3288. [Google Scholar] [CrossRef]
- Chen, L.L. Linking Long Noncoding RNA Localization and Function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef]
- Xia, F.; Dong, F.L.; Yang, Y.; Huang, A.F.; Chen, S.; Sun, D.; Xiong, S.D.; Zhang, J.P. Dynamic Transcription of Long Non-Coding RNA Genes during CD4+ T Cell Development and Activation. PLoS ONE 2014, 9, e101588. [Google Scholar] [CrossRef]
- Paralkar, V.R.; Mishra, T.; Luan, J.; Yao, Y.; Kossenkov, A.V.; Anderson, S.M.; Dunagin, M.; Pimkin, M.; Gore, M.; Sun, D.; et al. Lineage and species-specific long noncoding RNAs during erythro-megakaryocytic development. Blood 2014, 123, 1927–1937. [Google Scholar] [CrossRef]
- Wagner, L.A.; Christensen, C.J.; Dunn, D.M.; Spangrude, G.J.; Georgelas, A.; Kelley, L.; Esplin, M.S.; Weiss, R.B.; Gleich, G.J. EGO, a novel, noncoding RNA gene, regulates eosinophil granule protein transcript expression. Blood 2007, 109, 5191–5198. [Google Scholar] [CrossRef]
- Imperato, M.R.; Cauchy, P.; Obier, N.; Bonifer, C. The RUNX1-PU.1 axis in the control of hematopoiesis. Int. J. Hematol. 2015, 101, 319–329. [Google Scholar] [CrossRef]
- Wang, P.; Xue, Y.Q.; Han, Y.M.; Lin, L.; Wu, C.; Xu, S.; Jiang, Z.P.; Xu, J.F.; Liu, Q.Y.; Cao, X.T. The STAT3-Binding Long Noncoding RNA lnc-DC Controls Human Dendritic Cell Differentiation. Science 2014, 344, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, A.; Emmrich, S.; Schmidt, F.; Beck, D.; Ng, M.; Reimer, C.; Adams, F.F.; Grasedieck, S.; Witte, D.; Kabler, S.; et al. The non-coding RNA landscape of human hematopoiesis and leukemia. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Hon, C.C.; Ramilowski, J.A.; Harshbarger, J.; Bertin, N.; Rackham, O.J.; Gough, J.; Denisenko, E.; Schmeier, S.; Poulsen, T.M.; Severin, J.; et al. An atlas of human long non-coding RNAs with accurate 5′ ends. Nature 2017, 543, 199–204. [Google Scholar] [CrossRef]
- Berg, J.S.; Lin, K.K.; Sonnet, C.; Boles, N.C.; Weksberg, D.C.; Nguyen, H.; Holt, L.J.; Rickwood, D.; Daly, R.J.; Goodell, M.A. Imprinted Genes That Regulate Early Mammalian Growth Are Coexpressed in Somatic Stem Cells. PLoS ONE 2011, 6, e26410. [Google Scholar] [CrossRef] [PubMed]
- Ghazavi, F.; De Moerloose, B.; Van Loocke, W.; Wallaert, A.; Helsmoortel, H.H.; Ferster, A.; Bakkus, M.; Plat, G.; Delabesse, E.; Uyttebroeck, A.; et al. Unique long non-coding RNA expression signature in ETV6/RUNX1-driven B-cell precursor acute lymphoblastic leukemia. Oncotarget 2016, 7, 73769–73780. [Google Scholar] [CrossRef]
- Fernando, T.R.; Contreras, J.R.; Zampini, M.; Rodriguez-Malave, N.I.; Alberti, M.O.; Anguiano, J.; Tran, T.M.; Palanichamy, J.K.; Gajeton, J.; Ung, N.M.; et al. The lncRNA CASC15 regulates SOX4 expression in RUNX1-rearranged acute leukemia. Mol. Cancer 2017, 16, 126. [Google Scholar] [CrossRef]
- Sun, J.N.; Li, W.; Sun, Y.P.; Yu, D.H.; Wen, X.; Wang, H.; Cui, J.W.; Wang, G.J.; Hoffman, A.R.; Hu, J.F. A novel antisense long noncoding RNA within the IGF1R gene locus is imprinted in hematopoietic malignancies. Nucleic Acids Res. 2014, 42, 9588–9601. [Google Scholar] [CrossRef]
- Chen, S.Y.; Liang, H.R.; Yang, H.; Zhou, K.R.; Xu, L.M.; Liu, J.X.; Lai, B.; Song, L.; Luo, H.; Peng, J.M.; et al. Long non-coding RNAs: The novel diagnostic biomarkers for leukemia. Environ. Toxicol. Pharmacol. 2017, 55, 81–86. [Google Scholar] [CrossRef]
- Hughes, J.M.; Legnini, I.; Salvatori, B.; Masciarelli, S.; Marchioni, M.; Fazi, F.; Morlando, M.; Bozzoni, I.; Fatica, A. C/EBPα-p30 protein induces expression of the oncogenic long non-coding RNA UCA1 in acute myeloid leukemia. Oncotarget 2015, 6, 18534–18544. [Google Scholar] [CrossRef]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [PubMed]
- Benetatos, L.; Hatzimichael, E.; Dasoula, A.; Dranitsaris, G.; Tsiara, S.; Syrrou, M.; Georgiou, I.; Bourantas, K.L. CpG methylation analysis of the MEG3 and SNRPN imprinted genes in acute myeloid leukemia and myelodysplastic syndromes. Leuk. Res. 2010, 34, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, W.; Guo, R.; Sun, J.; Cui, J.; Wang, G.; Hoffman, A.R.; Hu, J.F. An intragenic long noncoding RNA interacts epigenetically with the RUNX1 promoter and enhancer chromatin DNA in hematopoietic malignancies. Int. J. Cancer 2014, 135, 2783–2794. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Xu, Y.; Xu, L.; Yu, X.; Cheng, J.; Yang, L.; Chen, S.; Li, Y. Inhibition of long non-coding RNA NEAT1 impairs myeloid differentiation in acute promyelocytic leukemia cells. BMC Cancer 2014, 14, 693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lian, Z.; Padden, C.; Gerstein, M.B.; Rozowsky, J.; Snyder, M.; Gingeras, T.R.; Kapranov, P.; Weissman, S.M.; Newburger, P.E. A myelopoiesis-associated regulatory intergenic noncoding RNA transcript within the human HOXA cluster. Blood 2009, 113, 2526–2534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Weissman, S.M.; Newburger, P.E. Long intergenic non-coding RNA HOTAIRM1 regulates cell cycle progression during myeloid maturation in NB4 human promyelocytic leukemia cells. RNA Biol. 2014, 11, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.H.; Wang, W.T.; Huang, W.; Fang, K.; Sun, Y.M.; Liu, S.R.; Luo, X.Q.; Chen, Y.Q. The lncRNA HOTAIRM1 regulates the degradation of PML-RARA oncoprotein and myeloid cell differentiation by enhancing the autophagy pathway. Cell Death Differ. 2017, 24, 212–224. [Google Scholar] [CrossRef]
- Wang, X.Q.; Dostie, J. Reciprocal regulation of chromatin state and architecture by HOTAIRM1 contributes to temporal collinear HOXA gene activation. Nucleic Acids Res. 2017, 45, 1091–1104. [Google Scholar] [CrossRef]
- Díaz-Beyá, M.; Brunet, S.; Nomdedéu, J.; Pratcorona, M.; Cordeiro, A.; Gallardo, D.; Escoda, L.; Tormo, M.; Heras, I.; Ribera, J.M.; et al. The lincRNA HOTAIRM1, located in the HOXA genomic region, is expressed in acute myeloid leukemia, impacts prognosis in patients in the intermediate-risk cytogenetic category, and is associated with a distinctive microRNA signature. Oncotarget 2015, 6, 31613–31627. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, X.; Frazão, J.B.; Condino-Neto, A.; Newburger, P.E. HOX antisense lincRNA HOXA-AS2 is an apoptosis repressor in all trans retinoic acid treated NB4 promyelocytic leukemia cells. J. Cell. Biochem. 2013, 114, 2375–2383. [Google Scholar] [CrossRef]
- Ebralidze, A.K.; Guibal, F.C.; Steidl, U.; Zhang, P.; Lee, S.; Bartholdy, B.; Jorda, M.A.; Petkova, V.; Rosenbauer, F.; Huang, G.; et al. PU.1 expression is modulated by the balance of functional sense and antisense RNAs regulated by a shared cis-regulatory element. Genes Dev. 2008, 22, 2085–2092. [Google Scholar] [CrossRef]
- McCarty, G.; Loeb, D.M. Hypoxia-sensitive epigenetic regulation of an antisense-oriented lncRNA controls WT1 expression in myeloid leukemia cells. PLoS ONE 2015, 10, e0119837. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Kang, Q.; Zhu, X.; Chen, Q.; Wang, X.; Chen, Y.; Ouyang, J.; Zhang, L.; Tan, H.; Chen, R.; et al. A long noncoding RNA critically regulates Bcr-Abl-mediated cellular transformation by acting as a competitive endogenous RNA. Oncogene 2015, 34, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, W.; Cao, L.; Li, Z.; Wang, X. Long Non-Coding RNA CCAT1 Acts as a Competing Endogenous RNA to Regulate Cell Growth and Differentiation in Acute Myeloid Leukemia. Mol. Cells 2016, 39, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Yoshikawa, R.; Harada, H.; Harada, Y.; Ishida, A.; Yamazaki, T. Long noncoding RNA, CCDC26, controls myeloid leukemia cell growth through regulation of KIT expression. Mol. Cancer 2015, 14, 90. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.Y.; Hu, X.Q.; Xie, F.Y.; Yu, Z.J.; Li, H.Y.; Bin-Zhou; Wu, J.B.; Tang, L.Y.; Gao, S.M. Long non-coding RNA HOTAIR modulates c-KIT expression through sponging miR-193a in acute myeloid leukemia. FEBS Lett. 2015, 589, 1981–1987. [Google Scholar] [CrossRef]
- Zeng, C.; Yu, X.; Lai, J.; Yang, L.; Chen, S.; Li, Y. Overexpression of the long non-coding RNA PVT1 is correlated with leukemic cell proliferation in acute promyelocytic leukemia. J. Hematol. Oncol. 2015, 8, 126. [Google Scholar] [CrossRef]
- Pan, J.Q.; Zhang, Y.Q.; Wang, J.H.; Xu, P.; Wang, W. lncRNA co-expression network model for the prognostic analysis of acute myeloid leukemia. Int. J. Mol. Med. 2017, 39, 663–671. [Google Scholar] [CrossRef]
- Fernando, T.R.; Rodriguez-Malave, N.I.; Waters, E.V.; Yan, W.H.; Casero, D.; Basso, G.; Pigazzi, M.; Rao, D.S. LncRNA Expression Discriminates Karyotype and Predicts Survival in B-Lymphoblastic Leukemia. Mol. Cancer Res. 2015, 13, 839–851. [Google Scholar] [CrossRef]
- Lajoie, M.; Drouin, S.; Caron, M.; St-Onge, P.; Ouimet, M.; Gioia, R.; Lafond, M.H.; Vidal, R.; Richer, C.; Oualkacha, K.; et al. Specific expression of novel long non-coding RNAs in high-hyperdiploid childhood acute lymphoblastic leukemia. PLoS ONE 2017, 12, e174124. [Google Scholar] [CrossRef]
- Ouimet, M.; Drouin, S.; Lajoie, M.; Caron, M.; St-Onge, P.; Gioia, R.; Richer, C.; Sinnett, D. A childhood acute lymphoblastic leukemia-specific lncRNA implicated in prednisolone resistance, cell proliferation, and migration. Oncotarget 2017, 8, 7477–7488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xu, H.G.; Lu, C. A novel long non-coding RNA T-ALL-R-LncR1 knockdown and Par-4 cooperate to induce cellular apoptosis in T-cell acute lymphoblastic leukemia cells. Leuk. Lymphoma 2014, 55, 1373–1382. [Google Scholar] [CrossRef] [PubMed]
- Melo, C.P.D.; Campos, C.B.; Rodrigues, J.D.; Aguirre-Neto, J.C.; Atalla, A.; Pianovski, M.A.D.; Carbone, E.K.; Lares, L.B.Q.; Moraes-Souza, H.; Octacilio-Silva, S.; et al. Long non-coding RNAs: Biomarkers for acute leukaemia subtypes. Br. J. Haematol. 2016, 173, 318–320. [Google Scholar] [CrossRef] [PubMed]
- Romero-Barrios, N.; Legascue, M.F.; Benhamed, M.; Ariel, F.; Crespi, M. Splicing regulation by long noncoding RNAs. Nucleic Acids Res. 2018, 46, 2169–2184. [Google Scholar] [CrossRef] [PubMed]
- Akbari Moqadam, F.; Lange-Turenhout, E.A.; Ariës, I.M.; Pieters, R.; Den Boer, M.L. MiR-125b, miR-100 and miR-99a co-regulate vincristine resistance in childhood acute lymphoblastic leukemia. Leuk. Res. 2013, 37, 1315–1321. [Google Scholar] [CrossRef]
- Zhang, X.; Hamblin, M.H.; Yin, K.J. The long noncoding RNA Malat1: Its physiological and pathophysiological functions. RNA Biol. 2017, 14, 1705–1714. [Google Scholar] [CrossRef]
- Fan, Y.; Shen, B.; Tan, M.; Mu, X.; Qin, Y.; Zhang, F.; Liu, Y. Long non-coding RNA UCA1 increases chemoresistance of bladder cancer cells by regulating Wnt signaling. FEBS J. 2014, 281, 1750–1758. [Google Scholar] [CrossRef]
- Han, Y.; Yang, Y.N.; Yuan, H.H.; Zhang, T.T.; Sui, H.; Wei, X.L.; Liu, L.; Huang, P.; Zhang, W.J.; Bai, Y.X. UCA1, a long non-coding RNA up-regulated in colorectal cancer influences cell proliferation, apoptosis and cell cycle distribution. Pathology 2014, 46, 396–401. [Google Scholar] [CrossRef]
- Sun, M.D.; Zheng, Y.Q.; Wang, L.P.; Zhao, H.T.; Yang, S. Long noncoding RNA UCA1 promotes cell proliferation, migration and invasion of human leukemia cells via sponging miR-126. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2233–2245. [Google Scholar]
- Zhang, Y.; Liu, Y.; Xu, X. Knockdown of LncRNA-UCA1 suppresses chemoresistance of pediatric AML by inhibiting glycolysis through the microRNA-125a/hexokinase 2 pathway. J. Cell. Biochem. 2018, 119, 6296–6308. [Google Scholar] [CrossRef]
- Miyoshi, N.; Wagatsuma, H.; Wakana, S.; Shiroishi, T.; Nomura, M.; Aisaka, K.; Kohda, T.; Surani, M.A.; Kaneko-Ishino, T.; Ishino, F. Identification of an imprinted gene, Meg3/Gtl2 and its human homologue MEG3, first mapped on mouse distal chromosome 12 and human chromosome 14q. Genes Cells 2000, 5, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Ren, Z.; Sun, P. Overexpression of the long non-coding RNA MEG3 impairs in vitro glioma cell proliferation. J. Cell. Biochem. 2012, 113, 1868–1874. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Beya, M.; Navarro, A.; Cordeiro, A.; Pratcorona, M.; Castellano, J.; Torrente, M.A.; Nomdedeu, M.; Risueño, R.; Rozman, M.; Monzo, M.; et al. Exploring the Expression Profile of Long Non-Coding RNA (lncRNA) in Different Acute Myeloid Leukemia (AML) Subtypes: t(8;16)(p11;p13)/MYST3-Crebbp AML Harbors a Distinctive LncRNA Signature. Blood 2015, 126, 1397. [Google Scholar]
- Garzon, R.; Volinia, S.; Papaioannou, D.; Nicolet, D.; Kohlschmidt, J.; Yan, P.S.; Mrozek, K.; Bucci, D.; Carroll, A.J.; Baer, M.R.; et al. Expression and prognostic impact of lncRNAs in acute myeloid leukemia. Proc. Natl. Acad. Sci. USA 2014, 111, 18679–18684. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, L.; Zhao, F.; Xu, R.; Jiang, J.; Zhang, C.; Liu, H.; Huang, H. Long non-coding RNA taurine-upregulated gene 1 correlates with poor prognosis, induces cell proliferation, and represses cell apoptosis via targeting aurora kinase A in adult acute myeloid leukemia. Ann. Hematol. 2018, 97, 1375–1389. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.L.; Ren, T.Y.; Cao, S.W.; Zheng, S.H.; Hu, X.M.; Hu, Y.W.; Lin, L.; Chen, J.; Zheng, L.; Wang, Q. HBx-related long non-coding RNA DBH-AS1 promotes cell proliferation and survival by activating MAPK signaling in hepatocellular carcinoma. Oncotarget 2015, 6, 33791–33804. [Google Scholar] [CrossRef] [PubMed]
- Gioia, R.; Drouin, S.; Ouimet, M.; Caron, M.; St-Onge, P.; Richer, C.; Sinnett, D. LncRNAs downregulated in childhood acute lymphoblastic leukemia modulate apoptosis, cell migration, and DNA damage response. Oncotarget 2017, 8, 80645–80650. [Google Scholar] [CrossRef] [PubMed]
- Garitano-Trojaola, A.; José-Enériz, E.S.; Ezponda, T.; Unfried, J.P.; Carrasco-León, A.; Razquin, N.; Barriocanal, M.; Vilas-Zornoza, A.; Sangro, B.; Segura, V.; et al. Deregulation of linc-PINT in acute lymphoblastic leukemia is implicated in abnormal proliferation of leukemic cells. Oncotarget 2018, 9, 12842–12852. [Google Scholar] [CrossRef]
- Ngoc, P.C.T.; Tan, S.H.; Tan, T.K.; Chan, M.M.; Li, Z.; Yeoh, A.E.J.; Tenen, D.G.; Sanda, T. Identification of novel lncRNAs regulated by the TAL1 complex in T-cell acute lymphoblastic leukemia. Leukemia 2018, 32, 2138–2151. [Google Scholar] [CrossRef]
- Ransohoff, J.D.; Wei, Y.N.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Sayad, A.; Hajifathali, A.; Hamidieh, A.A.; Roshandel, E.; Taheri, M. HOTAIR Long Noncoding RNA is not a Biomarker for Acute Myeloid Leukemia (AML) in Iranian Patients. Asian Pac. J. Cancer Prev. 2017, 18, 1581–1584. [Google Scholar] [PubMed]
- Li, J.; Sun, C.K. Long noncoding RNA SNHG5 is up-regulated and serves as a potential prognostic biomarker in acute myeloid leukemia. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3342–3347. [Google Scholar]
- Gao, S.; Zhou, B.; Li, H.; Huang, X.; Wu, Y.; Xing, C.; Yu, X.; Ji, Y. Long noncoding RNA HOTAIR promotes the self-renewal of leukemia stem cells through epigenetic silencing of p15. Exp. Hematol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Zheng, C.P.; Chen, S.Y.; Cai, X.P.; Shi, Y.J.; Lin, B.J.; Chen, Y.M. Overexpression of long non-coding RNA HOTAIR predicts a poor prognosis in patients with acute myeloid leukemia. Oncol. Lett. 2015, 10, 2410–2414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Huang, S.H.; Zhou, H.R.; Chen, C.J.; Tian, L.H.; Shen, J.Z. Role of HOTAIR in the diagnosis and prognosis of acute leukemia. Oncol. Rep. 2016, 36, 3113–3122. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Q.; Tang, S.; Li, M.; Feng, A.; Qin, L.; Liu, Z.; Wang, X. The role of long noncoding RNA HOTAIR in the acquired multidrug resistance to imatinib in chronic myeloid leukemia cells. Hematology 2017, 22, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Shang, C.; Guo, Y.; Zhang, H.; Xue, Y.X. Long noncoding RNA HOTAIR is a prognostic biomarker and inhibits chemosensitivity to doxorubicin in bladder transitional cell carcinoma. Cancer Chemother. Pharmacol. 2016, 77, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, N.; Verma, V.; Kumar, M.; Kumar, A.; Singh, B. Nanomedicine in cancer stem cell therapy: From fringe to forefront. Cell Tissue Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, T.; Yamamoto, T. Acceleration of cancer science with genome editing and related technologies. Cancer Sci. 2018. [Google Scholar] [CrossRef] [PubMed]


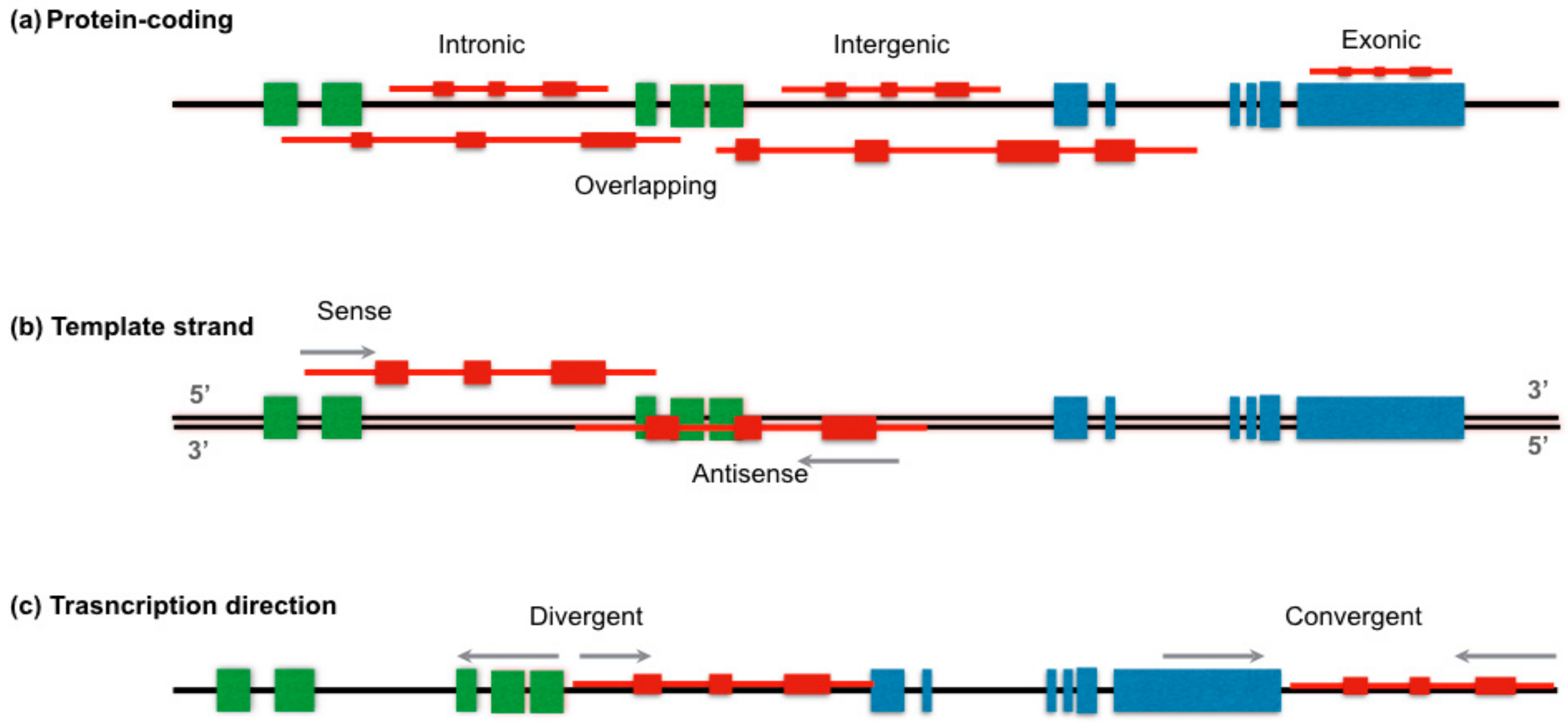{kind=link}
| Functional Type | Cellular Location | Mechanism of Action | Examples | Reference |
|---|---|---|---|---|
| Guide | Nucleus | Essential for the proper localization of proteins to their site-specific reaction. | XIST, ANRIL | [34] |
| Decoys | Plasma membrane, nucleus and cytosol | Sequestering regulatory factors (transcription factors, catalytic proteins subunits, chromatin modifiers, etc.) to modulate transcription | GAS5, MALAT1 | [35,36,37] |
| Scaffold | Nucleus | Providing platforms for assembly of multiple-component complexes such as the polycomb repressive complexes and ribonucleoprotein complex. | CDKN2B-AS1, HOTAIR | [35,36] |
| Signaling | Nucleus | Serving as a molecular signal to regulate transcription in response to various stimuli | TP53COR1, PANDAR | [35,36] |
| Enhancer | Nucleus | Binding with mediator complex to enhance transcription | HOTTIP, CCAT1-L, LUNAR1 | [25,33] |
| LncRNAs | Classification | Function | Target Genes | Expression Level in Leukemia | Reference |
|---|---|---|---|---|---|
| Myeloblastic Leukemia | |||||
| IRAIN | Intronic | Intrachromosomal interactions | IGF1R | Downregulated in leukemia cell lines and in patients with high risk AML | [49] |
| UCA1 | Intergenic | Proliferation of AML cells. Oncofetal gene | CDKN1B | Upregulated | [50,51,52] |
| MEG3 | Intergenic | Tumor suppressor gene | P53 | Downregulated | [52,53] |
| RUNXOR | Sense | Chromosomal translocations | RUNX1 | Upregulated | [54] |
| NEAT1 | Intergenic | Myeloid differentiation cells | Unknown in AML | Downregulated | [50,52,55] |
| LLEST | Tumor suppressor | BCL-2 | Downregulated or even not expressed. | ||
| HOTAIRM1 | Antisense | Myeloid differentiation cells, autophagy mechanisms, chromatin remodeling and architecture | HOXA1, HOXA4, CD11b and CD18 | Upregulated | [52,56,57,58,59,60] |
| HOXA-AS2 | Antisense | Apoptotic repressor in NB4 promyelocytic leukemia cells | Unknown | Upregulated | [61] |
| PU.1-AS | Antisense | Involved in the translation of PU.1 | PU.1 | Downregulated | [62] |
| WT1-AS | Antisense | WT1 expression | WT1 | [63] | |
| EGO | Intronic | MBP and EDN expression | [41] | ||
| BGL3 | Intergenic | Apoptosis and DNA methylation | miR-17, miR-93, miR-20a, miR-20b, miR-106a and miR-106b | Upregulated | [50,52,64] |
| CCAT1 | Intergenic | Monocytic cell differentiation | miR-155 | [9,52,65] | |
| CCDC26 | Intergenic | AML cell proliferation | c-Kit | [66] | |
| HOTAIR | Intergenic | Apoptosis inhibitor | miR-193a and c-Kit | Upregulated | [67] |
| PVT1 | Intergenic | Proliferation of promyelocytes | MYC | Upregulated | [52,68] |
| ZNF571-AS1 | Antisense | Regulator of JAK/STAT signaling pathway | KIT and STAT5 | [69] | |
| Lymphoblastic Leukemia | |||||
| BALR-2 | Uncharacterized | Unknown | Unknown | Overexpressed in prednisone-resistant B-ALL patients | [70] |
| BALR-1 | Unknown | Unknown | Unknown | Upregulated | [70] |
| BARL-6 | Unknown | Promotes cell survival and inhibits apoptosis | Unknown | Upregulated | [70] |
| LINC00958 | Intergenic | Unknown | Unknown | Upregulated in t(12;21) preB cALL | [70,71] |
| DBH-AS1 | Antisense | Cell proliferation and cell survival | Unknown | Upregulated | |
| RP11-137H2.4 | Uncharacterized | Apoptosis, proliferation, cell migration | Unknown | Upregulated. Glucocorticoids resistance | [72] |
| ANRIL | Antisense | Cellular proliferation and apoptosis | CDKN2A/B. CBX7, SUZ12 | Upregulated | [52] |
| T-ALL-R-LncR1 | Unknown | Promotor of the formation of Par-4/THAP1 protein complex, and the activity of caspase-3 | Unknown | Upregulated in children with T-ALL | [73] |
| LUNAR1 | Enhancer-like | Promotor of T-ALL proliferation by inducing IGF1R expression. | IGF1R | Downregulated | [50,52,74] |
| MALAT1 | Intergenic | Alternative splicing and epigenetic modification | Unknown | Upregulated Downregulated in vincristine-resistant ALL | [50,52,75,76,77] |
| CASC15 | Intergenic | Cellular survival and proliferation | SOX4 | Upregulated | [48] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Miranda, G.M.; Hidalgo-Miranda, A.; Bárcenas-López, D.A.; Núñez-Enríquez, J.C.; Ramírez-Bello, J.; Mejía-Aranguré, J.M.; Jiménez-Morales, S. Long Non-Coding RNA and Acute Leukemia. Int. J. Mol. Sci. 2019, 20, 735. https://doi.org/10.3390/ijms20030735
Cruz-Miranda GM, Hidalgo-Miranda A, Bárcenas-López DA, Núñez-Enríquez JC, Ramírez-Bello J, Mejía-Aranguré JM, Jiménez-Morales S. Long Non-Coding RNA and Acute Leukemia. International Journal of Molecular Sciences. 2019; 20(3):735. https://doi.org/10.3390/ijms20030735
Chicago/Turabian StyleCruz-Miranda, Gabriela Marisol, Alfredo Hidalgo-Miranda, Diego Alberto Bárcenas-López, Juan Carlos Núñez-Enríquez, Julian Ramírez-Bello, Juan Manuel Mejía-Aranguré, and Silvia Jiménez-Morales. 2019. "Long Non-Coding RNA and Acute Leukemia" International Journal of Molecular Sciences 20, no. 3: 735. https://doi.org/10.3390/ijms20030735
APA StyleCruz-Miranda, G. M., Hidalgo-Miranda, A., Bárcenas-López, D. A., Núñez-Enríquez, J. C., Ramírez-Bello, J., Mejía-Aranguré, J. M., & Jiménez-Morales, S. (2019). Long Non-Coding RNA and Acute Leukemia. International Journal of Molecular Sciences, 20(3), 735. https://doi.org/10.3390/ijms20030735