Placental CX3CL1 is Deregulated by Angiotensin II and Contributes to a Pro-Inflammatory Trophoblast-Monocyte Interaction

 , ,
, , 
Abstract
1. Introduction
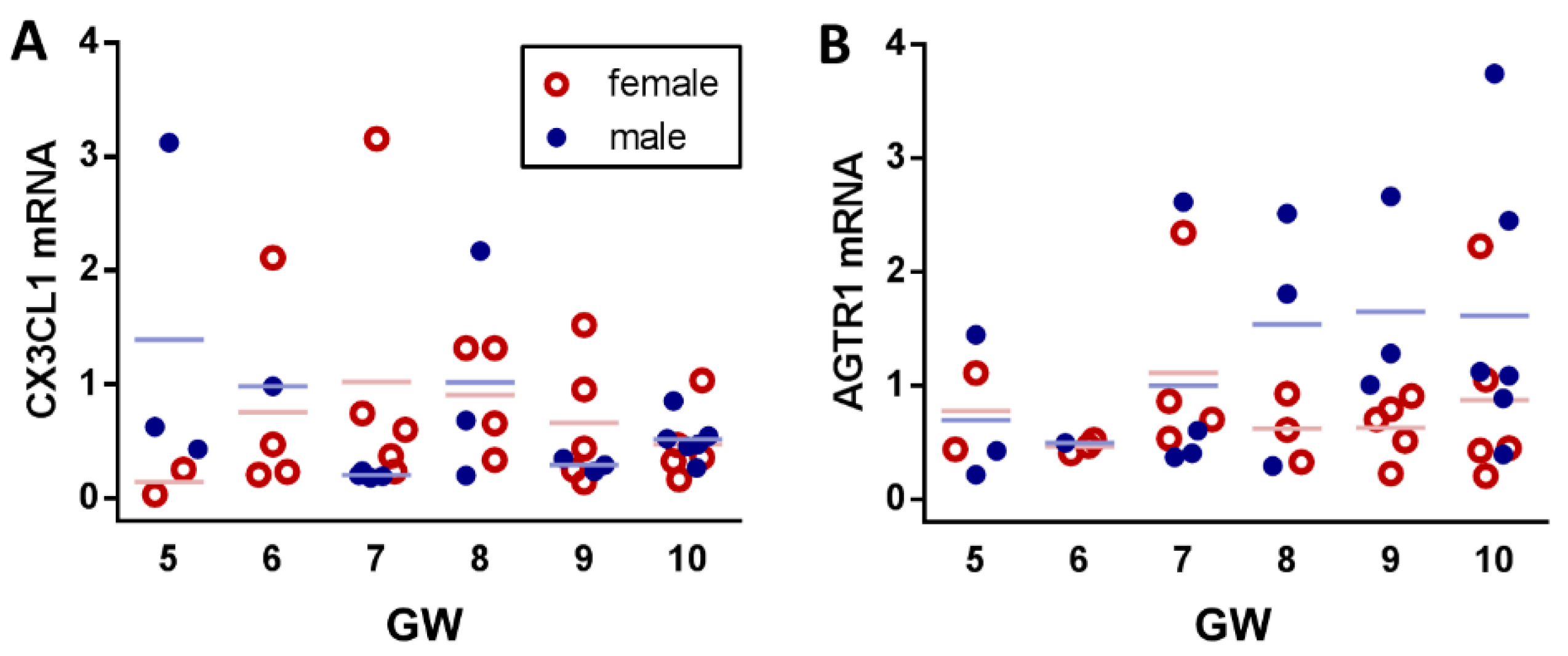
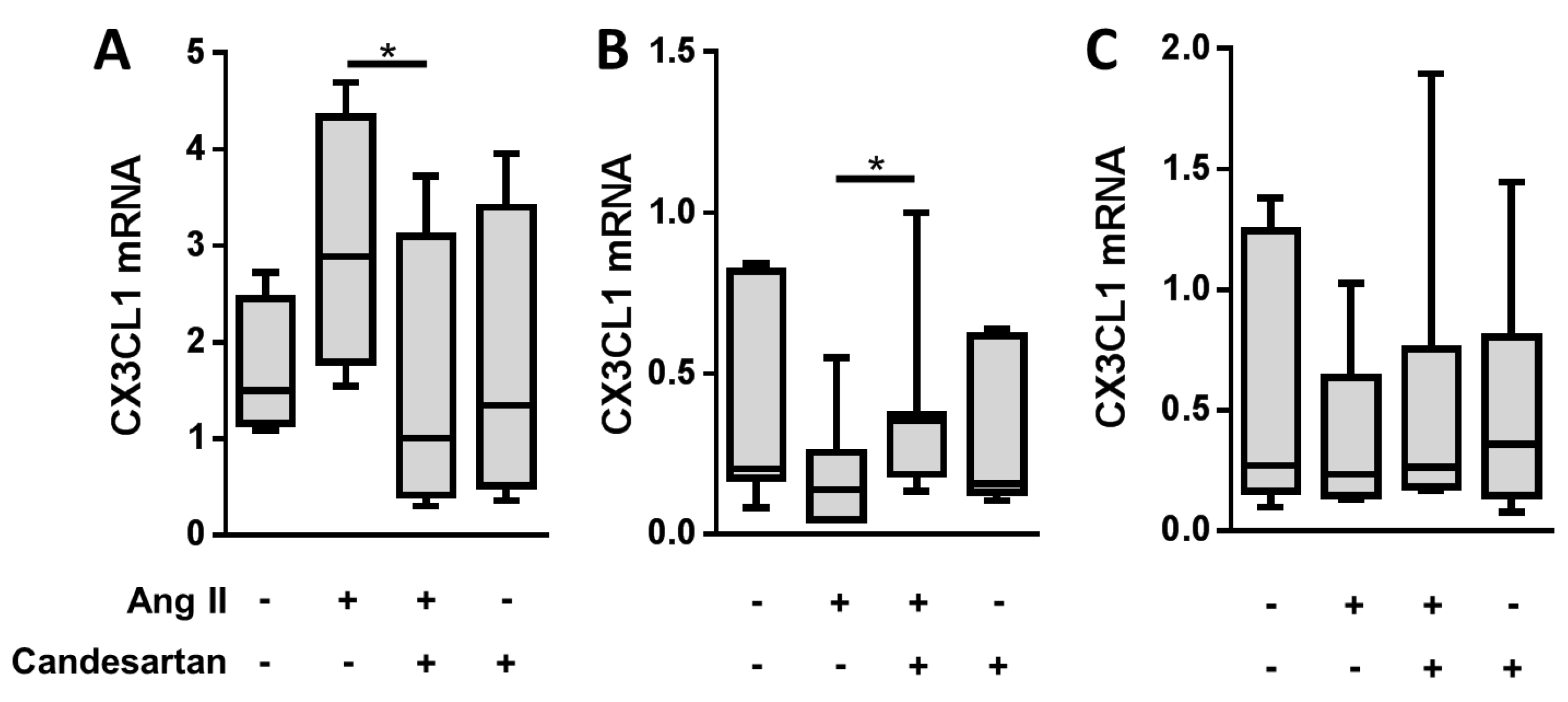
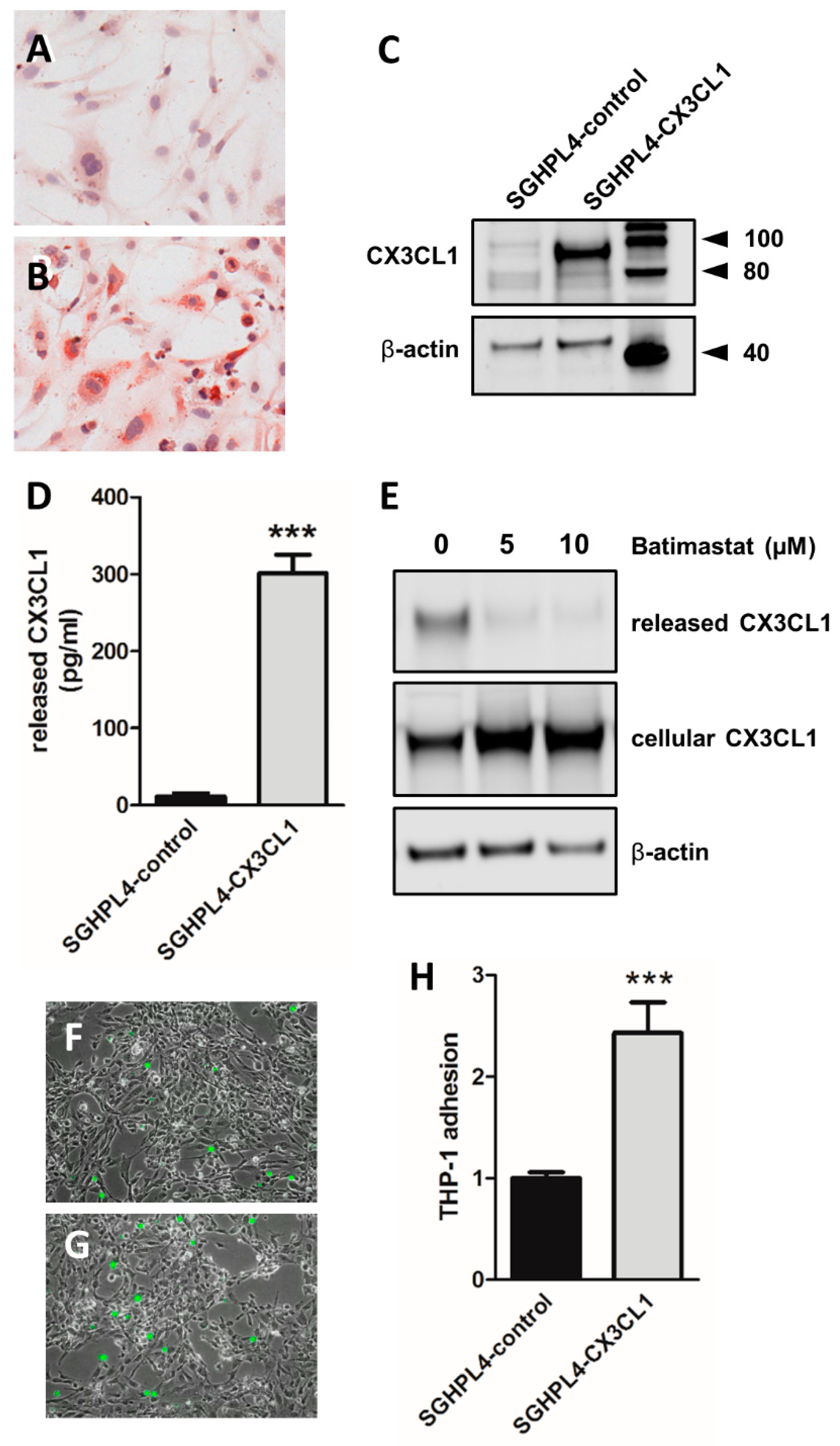
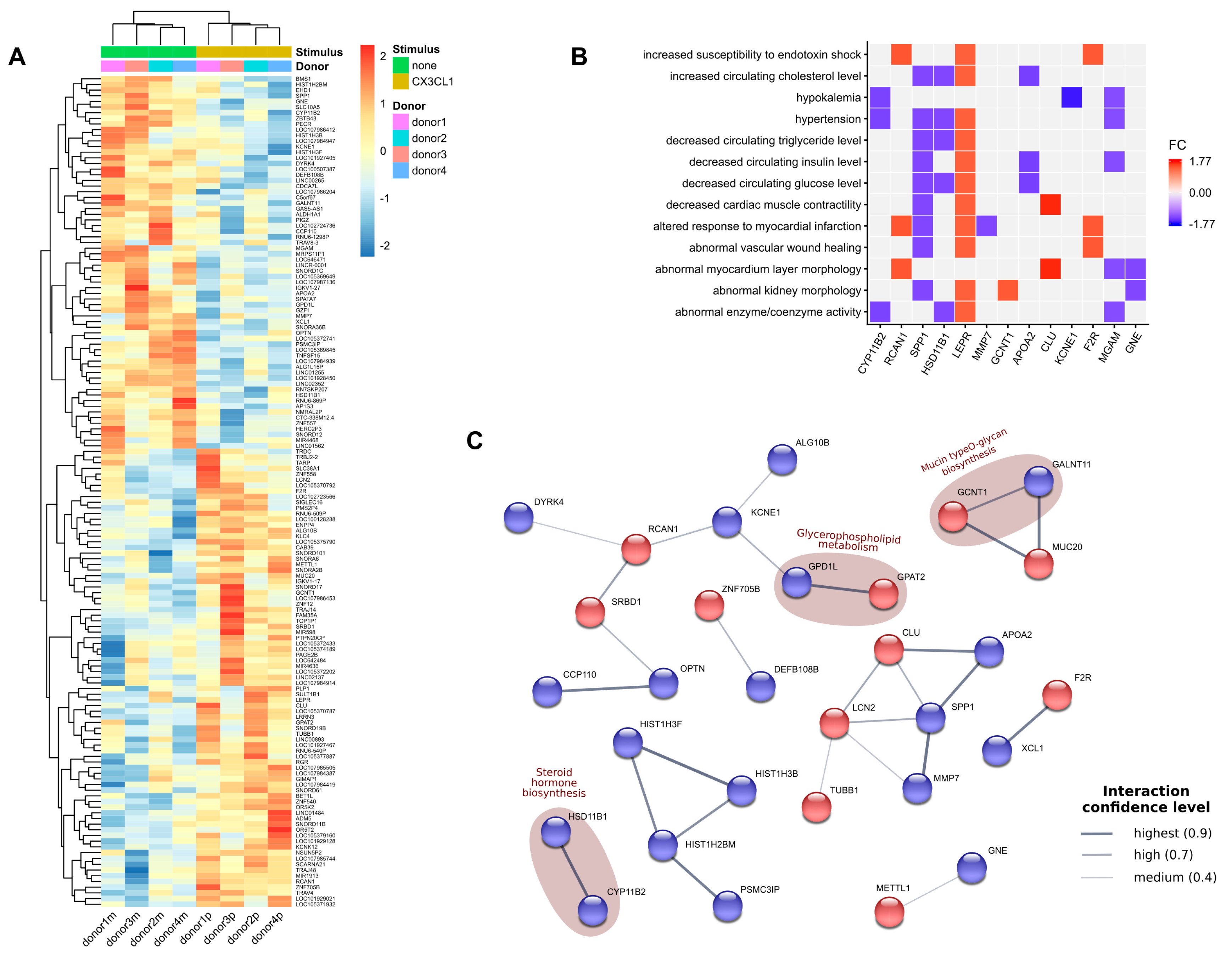
2. Results
3. Discussion
4. Materials and Methods
4.1. Human Specimen
4.2. qPCR Analysis
4.3. Placental Explant Culture
4.4. CX3CL1 Overexpression in SGHPL-4 Cells
4.5. Culture of Monocyte Cell Line THP-1
4.6. THP-1 Adhesion Assay
4.7. Immunocytochemistry
4.8. Immunoblotting
4.9. Isolation of Human Primary Monocytes
4.10. Co-Culture of SGHPL-4 Cells and Human Primary Monocytes
4.11. Multiplex Immunoassays
4.12. Stimulation of Human Primary Monocytes with Recombinant Human CX3CL1
4.13. Microarray Analysis
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benirschke, K.; Burton, G.J.; Baergen, R.N. Pathology of the Human Placenta, 6th ed.; Springer: Heidelberg, Germany, 2012. [Google Scholar]
- Lumbers, E.R.; Pringle, K.G. Roles of the Circulating Renin-Angiotensin-Aldosterone System in Human Pregnancy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R91–R101. [Google Scholar] [CrossRef] [PubMed]
- Tower, C.L.; Lui, S.; Charlesworth, N.R.; Smith, S.D.; Aplin, J.D.; Jones, R.L. Differential Expression of Angiotensin II Type 1 and Type 2 Receptors at the Maternal-Fetal Interface: Potential Roles in Early Placental Development. Reproduction 2010, 140, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Kalenga, M.K.; de Gasparo, M.; Thomas, K.; De Hertogh, R. Angiotensin II Induces Human Placental Lactogen and Pregnancy-Specific Beta 1-Glycoprotein Secretion Via an Angiotensin AT1 Receptor. Eur. J. Pharmacol. 1994, 268, 231–236. [Google Scholar] [CrossRef]
- Kalenga, M.K.; De Gasparo, M.; Thomas, K.; De Hertogh, R. Angiotensin-II Stimulates Estradiol Secretion from Human Placental Explants through AT1 Receptor Activation. J. Clin. Endocrinol. Metab. 1995, 80, 1233–1237. [Google Scholar] [PubMed]
- Zhou, C.C.; Ahmad, S.; Mi, T.; Xia, L.; Abbasi, S.; Hewett, P.W.; Sun, C.; Ahmed, A.; Kellems, R.E.; Xia, Y. Angiotensin II Induces Soluble Fms-Like Tyrosine Kinase-1 Release Via Calcineurin Signaling Pathway in Pregnancy. Circ. Res. 2007, 100, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Dhindsa, S.; Ghanim, H.; Chaudhuri, A. Angiotensin II and Inflammation: The Effect of Angiotensin-Converting Enzyme Inhibition and Angiotensin II Receptor Blockade. J. Hum. Hypertens. 2007, 21, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.N.; Broughton Pipkin, F.; Symonds, E.M. Platelet Angiotensin II Binding and Plasma Renin Concentration, Plasma Renin Substrate and Plasma Angiotensin II in Human Pregnancy. Clin. Sci. (Lond.) 1990, 79, 403–408. [Google Scholar] [CrossRef]
- Browatzki, M.; Larsen, D.; Pfeiffer, C.A.; Gehrke, S.G.; Schmidt, J.; Kranzhofer, A.; Katus, H.A.; Kranzhofer, R. Angiotensin II Stimulates Matrix Metalloproteinase Secretion in Human Vascular Smooth Muscle Cells Via Nuclear Factor-kappaB and Activator Protein 1 in a Redox-Sensitive Manner. J. Vasc. Res. 2005, 42, 415–423. [Google Scholar] [CrossRef]
- Ruiz-Ortega, M.; Lorenzo, O.; Ruperez, M.; Blanco, J.; Egido, J. Systemic Infusion of Angiotensin II into Normal Rats Activates Nuclear Factor-kappaB and AP-1 in the Kidney: Role of AT(1) and AT(2) Receptors. Am. J. Pathol. 2001, 158, 1743–1756. [Google Scholar] [CrossRef]
- Ruiz-Ortega, M.; Lorenzo, O.; Egido, J. Angiotensin III Increases MCP-1 and Activates NF-kappaB and AP-1 in Cultured Mesangial and Mononuclear Cells. Kidney Int. 2000, 57, 2285–2298. [Google Scholar] [CrossRef]
- Zlotnik, A.; Yoshie, O. Chemokines: A New Classification System and their Role in Immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef]
- Garton, K.J.; Gough, P.J.; Blobel, C.P.; Murphy, G.; Greaves, D.R.; Dempsey, P.J.; Raines, E.W. Tumor Necrosis Factor-Alpha-Converting Enzyme (ADAM17) Mediates the Cleavage and Shedding of Fractalkine (CX3CL1). J. Biol. Chem. 2001, 276, 37993–38001. [Google Scholar] [PubMed]
- Hundhausen, C.; Misztela, D.; Berkhout, T.A.; Broadway, N.; Saftig, P.; Reiss, K.; Hartmann, D.; Fahrenholz, F.; Postina, R.; Matthews, V.; et al. The Disintegrin-Like Metalloproteinase ADAM10 is Involved in Constitutive Cleavage of CX3CL1 (Fractalkine) and Regulates CX3CL1-Mediated Cell-Cell Adhesion. Blood 2003, 102, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Kervancioglu Demirci, E.; Salamonsen, L.A.; Gauster, M. The Role of CX3CL1 in Fetal-Maternal Interaction during Human Gestation. Cell Adhes. Migr. 2016, 10, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Szukiewicz, D.; Kochanowski, J.; Mittal, T.K.; Pyzlak, M.; Szewczyk, G.; Cendrowski, K. Chorioamnionitis (ChA) Modifies CX3CL1 (Fractalkine) Production by Human Amniotic Epithelial Cells (HAEC) Under Normoxic and Hypoxic Conditions. J. Inflamm. (Lond.) 2014, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Szukiewicz, D.; Kochanowski, J.; Pyzlak, M.; Szewczyk, G.; Stangret, A.; Mittal, T.K. Fractalkine (CX3CL1) and its Receptor CX3CR1 may Contribute to Increased Angiogenesis in Diabetic Placenta. Mediators Inflamm. 2013, 2013, 437576. [Google Scholar] [CrossRef] [PubMed]
- Siwetz, M.; Dieber-Rotheneder, M.; Cervar-Zivkovic, M.; Kummer, D.; Kremshofer, J.; Weiss, G.; Herse, F.; Huppertz, B.; Gauster, M. Placental Fractalkine is Up-Regulated in Severe Early-Onset Preeclampsia. Am. J. Pathol. 2015, 185, 1334–1343. [Google Scholar] [CrossRef]
- Usta, A.; Turan, G.; Sancakli Usta, C.; Avci, E.; Adali, E. Placental Fractalkine Immunoreactivity in Preeclampsia and its Correlation with Histopathological Changes in the Placenta and Adverse Pregnancy Outcomes. J. Matern. Fetal. Neonatal Med. 2018. [Google Scholar] [CrossRef]
- Hildemann, S.K.; Schulz, C.; Fraccarollo, D.; Schopp, C.; Flierl, U.; Wissel, K.; Pelisek, J.; Massberg, S.; Bauersachs, J.; Schafer, A. Fractalkine Promotes Platelet Activation and Vascular Dysfunction in Congestive Heart Failure. Thromb. Haemost. 2014, 111, 725–735. [Google Scholar] [CrossRef]
- Flierl, U.; Bauersachs, J.; Schafer, A. Modulation of Platelet and Monocyte Function by the Chemokine Fractalkine (CX3 CL1) in Cardiovascular Disease. Eur. J. Clin. Investig. 2015, 45, 624–633. [Google Scholar] [CrossRef]
- Siwetz, M.; Blaschitz, A.; Kremshofer, J.; Bilic, J.; Desoye, G.; Huppertz, B.; Gauster, M. Metalloprotease Dependent Release of Placenta Derived Fractalkine. Mediat. Inflamm. 2014, 2014, 839290. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.; Tacke, R.; Hedrick, C.C.; Hanna, R.N. Nonclassical Patrolling Monocyte Function in the Vasculature. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.F.; Wang, H. Monocyte and Macrophage Differentiation: Circulation Inflammatory Monocyte as Biomarker for Inflammatory Diseases. Biomark. Res. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Rius, C.; Piqueras, L.; Gonzalez-Navarro, H.; Albertos, F.; Company, C.; Lopez-Gines, C.; Ludwig, A.; Blanes, J.I.; Morcillo, E.J.; Sanz, M.J. Arterial and Venous Endothelia Display Differential Functional Fractalkine (CX3CL1) Expression by Angiotensin-II. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Cao, Y.; Liu, S.; Peng, Z.; Zhang, S. Neferine Inhibits Angiotensin II-Induced Rat Aortic Smooth Muscle Cell Proliferation Predominantly by Downregulating Fractalkine Gene Expression. Exp. Ther. Med. 2014, 8, 1545–1550. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, B. Placental Origins of Preeclampsia: Challenging the Current Hypothesis. Hypertension 2008, 51, 970–975. [Google Scholar] [CrossRef]
- Herse, F.; Staff, A.C.; Hering, L.; Muller, D.N.; Luft, F.C.; Dechend, R. AT1-Receptor Autoantibodies and Uteroplacental RAS in Pregnancy and Pre-Eclampsia. J. Mol. Med. (Berl) 2008, 86, 697–703. [Google Scholar] [CrossRef]
- Wenzel, K.; Rajakumar, A.; Haase, H.; Geusens, N.; Hubner, N.; Schulz, H.; Brewer, J.; Roberts, L.; Hubel, C.A.; Herse, F.; et al. Angiotensin II Type 1 Receptor Antibodies and Increased Angiotensin II Sensitivity in Pregnant Rats. Hypertension 2011, 58, 77–84. [Google Scholar] [CrossRef]
- Herse, F.; Dechend, R.; Harsem, N.K.; Wallukat, G.; Janke, J.; Qadri, F.; Hering, L.; Muller, D.N.; Luft, F.C.; Staff, A.C. Dysregulation of the Circulating and Tissue-Based Renin-Angiotensin System in Preeclampsia. Hypertension 2007, 49, 604–611. [Google Scholar] [CrossRef]
- Herse, F.; LaMarca, B. Angiotensin II Type 1 Receptor Autoantibody (AT1-AA)-Mediated Pregnancy Hypertension. Am. J. Reprod. Immunol. 2013, 69, 413–418. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Placental Oxidative Stress: From Miscarriage to Preeclampsia. J. Soc. Gynecol. Investig. 2004, 11, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Cindrova-Davies, T. Gabor than Award Lecture 2008: Pre-Eclampsia-from Placental Oxidative Stress to Maternal Endothelial Dysfunction. Placenta 2009, 30, S55–S65. [Google Scholar] [CrossRef] [PubMed]
- Lamarca, B. Endothelial Dysfunction. an Important Mediator in the Pathophysiology of Hypertension during Pre-Eclampsia. Minerva Ginecol. 2012, 64, 309–320. [Google Scholar]
- Jauniaux, E.; Poston, L.; Burton, G.J. Placental-Related Diseases of Pregnancy: Involvement of Oxidative Stress and Implications in Human Evolution. Hum. Reprod. Update 2006, 12, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E.; Murray, A.J. Oxygen and Placental Development; Parallels and Differences with Tumour Biology. Placenta 2017, 56, 14–18. [Google Scholar] [CrossRef]
- George, A.J.; Thomas, W.G.; Hannan, R.D. The Renin-Angiotensin System and Cancer: Old Dog, New Tricks. Nat. Rev. Cancer 2010, 10, 745–759. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, X.; Cai, L.; Yang, Y.; Sui, G.; Fu, S. Angiotensin II/angiotensin II Type I Receptor (AT1R) Signaling Promotes MCF-7 Breast Cancer Cells Survival Via PI3-kinase/Akt Pathway. J. Cell. Physiol. 2010, 225, 168–173. [Google Scholar] [CrossRef]
- Kurlak, L.O.; Mistry, H.D.; Cindrova-Davies, T.; Burton, G.J.; Broughton Pipkin, F. Human Placental Renin-Angiotensin System in Normotensive and Pre-Eclamptic Pregnancies at High Altitude and After Acute Hypoxia-Reoxygenation Insult. J. Physiol. 2016, 594, 1327–1340. [Google Scholar] [CrossRef]
- Granger, J.P.; Alexander, B.T.; Llinas, M.T.; Bennett, W.A.; Khalil, R.A. Pathophysiology of Hypertension during Preeclampsia Linking Placental Ischemia with Endothelial Dysfunction. Hypertension 2001, 38, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Kulasingam, V.; Alexander, R.T.; Touret, N.; Fong, A.M.; Patel, D.D.; Robinson, L.A. Recycling of the Membrane-Anchored Chemokine, CX3CL1. J. Biol. Chem. 2005, 280, 19858–19866. [Google Scholar] [CrossRef] [PubMed]
- Siwetz, M.; Sundl, M.; Kolb, D.; Hiden, U.; Herse, F.; Huppertz, B.; Gauster, M. Placental Fractalkine Mediates Adhesion of THP-1 Monocytes to Villous Trophoblast. Histochem. Cell Biol. 2015, 143, 565–574. [Google Scholar] [CrossRef]
- Przybyl, L.; Haase, N.; Golic, M.; Rugor, J.; Solano, M.E.; Arck, P.C.; Gauster, M.; Huppertz, B.; Emontzpohl, C.; Stoppe, C.; et al. CD74-Downregulation of Placental Macrophage-Trophoblastic Interactions in Preeclampsia. Circ. Res. 2016, 119, 55–68. [Google Scholar] [CrossRef]
- Haskell, C.A.; Cleary, M.D.; Charo, I.F. Unique Role of the Chemokine Domain of Fractalkine in Cell Capture. Kinetics of Receptor Dissociation Correlate with Cell Adhesion. J. Biol. Chem. 2000, 275, 34183–34189. [Google Scholar] [CrossRef]
- Li, C.; He, J.; Zhong, X.; Gan, H.; Xia, Y. CX3CL1/CX3CR1 Axis Contributes to Angiotensin II-Induced Vascular Smooth Muscle Cell Proliferation and Inflammatory Cytokine Production. Inflammation 2018, 41, 824–834. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, M.H.; Yi, H.S.; Kim, S.Y.; Kim, H.H.; Kim, J.H.; Yeon, J.E.; Byun, K.S.; Byun, J.S.; Jeong, W.I. CX3CR1 Differentiates F4/80(Low) Monocytes into Pro-Inflammatory F4/80(High) Macrophages in the Liver. Sci. Rep. 2018, 8, 15076. [Google Scholar] [CrossRef]
- Florentin, J.; Coppin, E.; Vasamsetti, S.B.; Zhao, J.; Tai, Y.Y.; Tang, Y.; Zhang, Y.; Watson, A.; Sembrat, J.; Rojas, M.; et al. Inflammatory Macrophage Expansion in Pulmonary Hypertension Depends upon Mobilization of Blood-Borne Monocytes. J. Immunol. 2018, 200, 3612–3625. [Google Scholar] [CrossRef]
- Bellamy, L.; Casas, J.P.; Hingorani, A.D.; Williams, D.J. Pre-Eclampsia and Risk of Cardiovascular Disease and Cancer in Later Life: Systematic Review and Meta-Analysis. BMJ 2007, 335, 974. [Google Scholar] [CrossRef]
- McDonald, S.D.; Malinowski, A.; Zhou, Q.; Yusuf, S.; Devereaux, P.J. Cardiovascular Sequelae of preeclampsia/eclampsia: A Systematic Review and Meta-Analyses. Am. Heart J. 2008, 156, 918–930. [Google Scholar] [CrossRef]
- Ostuni, M.A.; Guellec, J.; Hermand, P.; Durand, P.; Combadiere, C.; Pincet, F.; Deterre, P. CX3CL1, a Chemokine Finely Tuned to Adhesion: Critical Roles of the Stalk Glycosylation and the Membrane Domain. Biol. Open 2014, 3, 1173–1182. [Google Scholar] [CrossRef]
- Harrison, J.K.; Fong, A.M.; Swain, P.A.; Chen, S.; Yu, Y.R.; Salafranca, M.N.; Greenleaf, W.B.; Imai, T.; Patel, D.D. Mutational Analysis of the Fractalkine Chemokine Domain. Basic Amino Acid Residues Differentially Contribute to CX3CR1 Binding, Signaling, and Cell Adhesion. J. Biol. Chem. 2001, 276, 21632–21641. [Google Scholar] [CrossRef]
- Barnett-Norris, J.; Lynch, D.; Reggio, P.H. Lipids, Lipid Rafts and Caveolae: Their Importance for GPCR Signaling and their Centrality to the Endocannabinoid System. Life Sci. 2005, 77, 1625–1639. [Google Scholar] [CrossRef]
- Drewlo, S.; Levytska, K.; Kingdom, J. Revisiting the Housekeeping Genes of Human Placental Development and Insufficiency Syndromes. Placenta 2012, 33, 952–954. [Google Scholar] [CrossRef]
- Siwetz, M.; Blaschitz, A.; El-Heliebi, A.; Hiden, U.; Desoye, G.; Huppertz, B.; Gauster, M. TNF-Alpha Alters the Inflammatory Secretion Profile of Human First Trimester Placenta. Lab. Invest. 2016, 96, 428. [Google Scholar] [CrossRef]
- Gauster, M.; Maninger, S.; Siwetz, M.; Deutsch, A.; El-Heliebi, A.; Kolb-Lenz, D.; Hiden, U.; Desoye, G.; Herse, F.; Prokesch, A. Downregulation of p53 Drives Autophagy during Human Trophoblast Differentiation. Cell Mol. Life Sci. 2017, 75, 1839–1855. [Google Scholar] [CrossRef]
- Binger, K.J.; Gebhardt, M.; Heinig, M.; Rintisch, C.; Schroeder, A.; Neuhofer, W.; Hilgers, K.; Manzel, A.; Schwartz, C.; Kleinewietfeld, M.; et al. High Salt Reduces the Activation of IL-4- and IL-13-Stimulated Macrophages. J. Clin. Invest. 2015, 125, 4223–4238. [Google Scholar] [CrossRef]
- Kolesnikov, N.; Hastings, E.; Keays, M.; Melnichuk, O.; Tang, Y.A.; Williams, E.; Dylag, M.; Kurbatova, N.; Brandizi, M.; Burdett, T.; et al. ArrayExpress Update—Simplifying Data Submissions. Nucleic Acids Res. 2015, 43, D1113-6. [Google Scholar] [CrossRef]
- Brazma, A.; Hingamp, P.; Quackenbush, J.; Sherlock, G.; Spellman, P.; Stoeckert, C.; Aach, J.; Ansorge, W.; Ball, C.A.; Causton, H.C.; et al. Minimum Information about a Microarray Experiment (MIAME)-Toward Standards for Microarray Data. Nat. Genet. 2001, 29, 365–371. [Google Scholar] [CrossRef]
- Sandberg, R.; Larsson, O. Improved Precision and Accuracy for Microarrays using Updated Probe Set Definitions. BMC Bioinform. 2007, 8, 48. [Google Scholar] [CrossRef]
- Kauffmann, A.; Gentleman, R.; Huber, W. ArrayQualityMetrics—A Bioconductor Package for Quality Assessment of Microarray Data. Bioinformatics 2009, 25, 415–416. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- SEQC/MAQC-III Consortium. A Comprehensive Assessment of RNA-Seq Accuracy, Reproducibility and Information Content by the Sequencing Quality Control Consortium. Nat. Biotechnol. 2014, 32, 903–914. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-Protein Interaction Networks, Integrated Over the Tree of Life. Nucleic Acids Res. 2015, 43, D447-52. [Google Scholar] [CrossRef]
- Gel, B.; Serra, E. KaryoploteR: An R/Bioconductor Package to Plot Customizable Genomes Displaying Arbitrary Data. Bioinformatics 2017, 33, 3088–3090. [Google Scholar] [CrossRef]
- Durinck, S.; Spellman, P.T.; Birney, E.; Huber, W. Mapping Identifiers for the Integration of Genomic Datasets with the R/Bioconductor Package biomaRt. Nat. Protoc. 2009, 4, 1184–1191. [Google Scholar] [CrossRef]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating High-Throughput Genomic Analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef]
- Melgert, B.N.; Spaans, F.; Borghuis, T.; Klok, P.A.; Groen, B.; Bolt, A.; de Vos, P.; van Pampus, M.G.; Wong, T.Y.; van Goor, H.; et al. Pregnancy and Preeclampsia Affect Monocyte Subsets in Humans and Rats. PLoS ONE 2012, 7, e45229. [Google Scholar] [CrossRef]
- White, G.E.; McNeill, E.; Channon, K.M.; Greaves, D.R. Fractalkine Promotes Human Monocyte Survival Via a Reduction in Oxidative Stress. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2554–2562. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Female (n = 25) | Male (n = 20) | p Value | ||
|---|---|---|---|---|---|
| Gestational age (days) | 56.84 | ± 12.62 | 56.85 | ± 13.55 | 0.998 |
| Fetal crown-rump length (CRL) | 1.74 | ± 1.27 | 1.84 | ± 1.43 | 0.797 |
| Placental volume | 2.20 | ± 1.74 | 1.86 | ± 0.79 | 0.625 |
| Maternal age (years) | 26.68 | ± 4.61 | 25.30 | ± 5.45 | 0.362 |
| Maternal weight (kg) | 58.32 | ± 7.40 | 57.25 | ± 7.89 | 0.642 |
| Maternal height (cm) | 164.84 | ± 4.57 | 165.25 | ± 6.80 | 0.811 |
| Maternal BMI | 21.44 | ± 2.34 | 20.90 | ± 1.85 | 0.399 |
| Cytokine/Chemokine | Aliases | Control (pg/mL) | CX3CL1 (pg/mL) | Fold Change | p Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| MIF | 5993.0 | ± | 928.5 | 5740.5 | ± | 653.6 | 0.96 | 0.6857 | |
| IL8 | CXCL8 | 181.7 | ± | 40.3 | 347.1 | ± | 23.1 | 1.91 | 0.0286 |
| CCL2 | MCP-1 | 37.5 | ± | 6.3 | 47.3 | ± | 14.6 | 1.26 | 0.8857 |
| IL6 | 26.9 | ± | 7.7 | 24.1 | ± | 1.5 | 0.89 | 1.0000 | |
| CCL24 | eotaxin-2 | 18.9 | ± | 12.3 | 34.9 | ± | 13.5 | 1.85 | 0.3005 |
| IL16 | 16.4 | ± | 3.4 | 23.7 | ± | 4.1 | 1.44 | 0.3429 | |
| CCL19 | MIP-3β | 7.1 | ± | 2.5 | 14.5 | ± | 1.7 | 2.03 | 0.0294 |
| CCL13 | MCP-4 | 3.4 | ± | 0.5 | 5.8 | ± | 0.3 | 1.69 | 0.0286 |
| CXCL16 | SCYB16 | 3.1 | ± | 0.6 | 3.8 | ± | 0.6 | 1.25 | 0.4857 |
| CCL20 | MIP-3α | 2.8 | ± | 1.2 | 9.8 | ± | 4.6 | 3.47 | 0.3429 |
| CCL3 | MIP-1α | 1.7 | ± | 0.4 | 1.8 | ± | 0.1 | 1.06 | 0.8857 |
| CCL8 | MCP-2 | 0.9 | ± | 0.2 | 0.4 | ± | 0.2 | 0.45 | 0.3094 |
| CXCL11 | I-TAC | 0.6 | ± | 0.4 | 0.8 | ± | 0.3 | 1.26 | 0.8794 |
| CXCL5 | ENA-78 | n.d. | 198.3 | ± | 67.5 | ||||
| CXCL1 | Gro-α | n.d. | 62.0 | ± | 36.6 | ||||
| CCL21 | 6Ckine | n.d. | 49.4 | ± | 9.9 | ||||
| CXCL6 | GCP-2 | n.d. | 13.4 | ± | 7.8 | ||||
| IL10 | n.d. | 6.3 | ± | 0.6 | |||||
| IL1β | n.d. | 0.5 | ± | 0.3 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nonn, O.; Güttler, J.; Forstner, D.; Maninger, S.; Zadora, J.; Balogh, A.; Frolova, A.; Glasner, A.; Herse, F.; Gauster, M. Placental CX3CL1 is Deregulated by Angiotensin II and Contributes to a Pro-Inflammatory Trophoblast-Monocyte Interaction. Int. J. Mol. Sci. 2019, 20, 641. https://doi.org/10.3390/ijms20030641
Nonn O, Güttler J, Forstner D, Maninger S, Zadora J, Balogh A, Frolova A, Glasner A, Herse F, Gauster M. Placental CX3CL1 is Deregulated by Angiotensin II and Contributes to a Pro-Inflammatory Trophoblast-Monocyte Interaction. International Journal of Molecular Sciences. 2019; 20(3):641. https://doi.org/10.3390/ijms20030641
Chicago/Turabian StyleNonn, Olivia, Jacqueline Güttler, Désirée Forstner, Sabine Maninger, Julianna Zadora, András Balogh, Alina Frolova, Andreas Glasner, Florian Herse, and Martin Gauster. 2019. "Placental CX3CL1 is Deregulated by Angiotensin II and Contributes to a Pro-Inflammatory Trophoblast-Monocyte Interaction" International Journal of Molecular Sciences 20, no. 3: 641. https://doi.org/10.3390/ijms20030641
APA StyleNonn, O., Güttler, J., Forstner, D., Maninger, S., Zadora, J., Balogh, A., Frolova, A., Glasner, A., Herse, F., & Gauster, M. (2019). Placental CX3CL1 is Deregulated by Angiotensin II and Contributes to a Pro-Inflammatory Trophoblast-Monocyte Interaction. International Journal of Molecular Sciences, 20(3), 641. https://doi.org/10.3390/ijms20030641