Expression of Immune Regulatory Genes in the Porcine Internal Genital Tract Is Differentially Triggered by Spermatozoa and Seminal Plasma

 ,
, 

Abstract
1. Introduction
2. Results
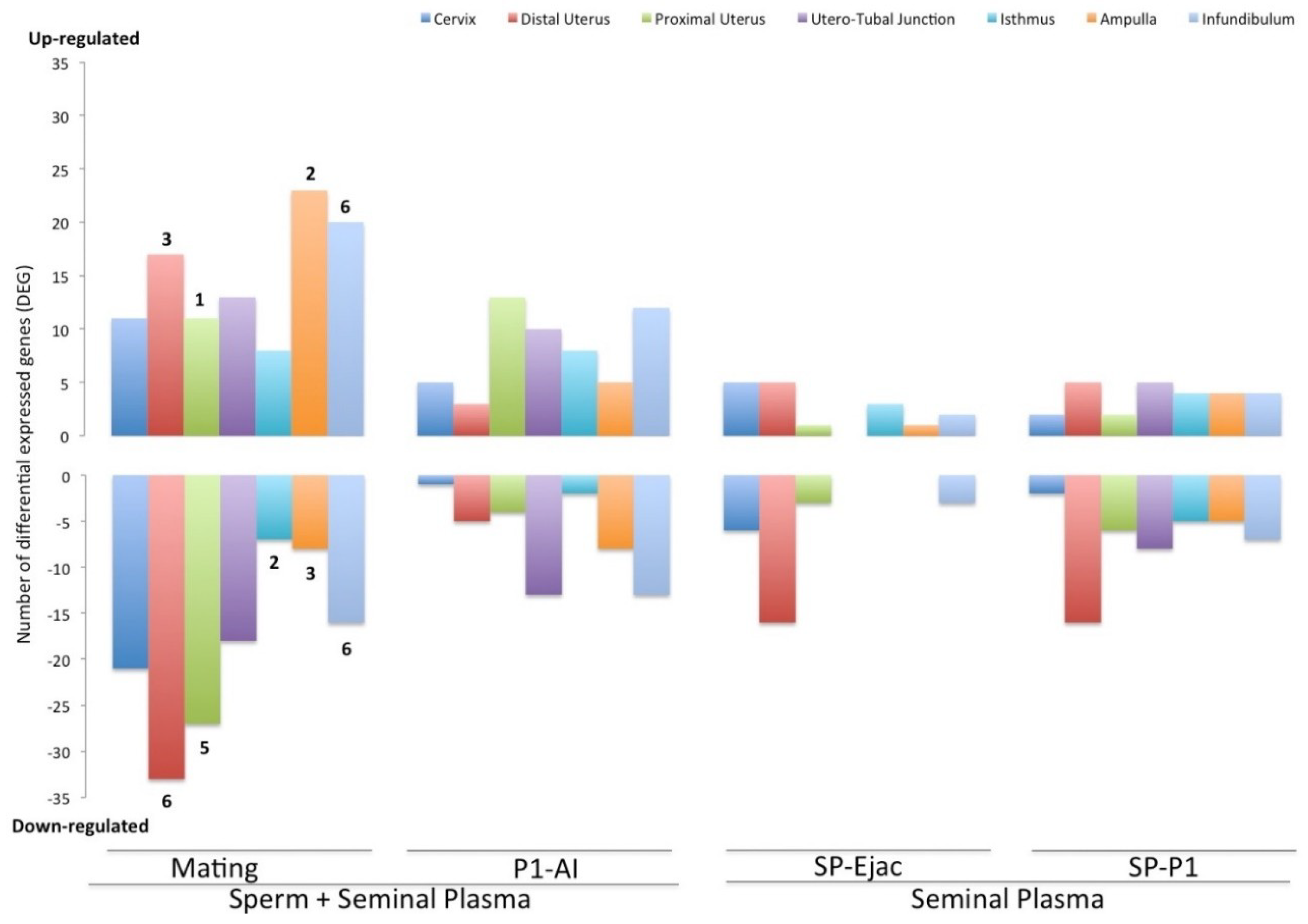
2.1. Entry of Semen-Induced Expression Changes of Genes Involved in Immune System Processes in the Entire Female Genital Tract
2.2. Gene Ontologies of the Immune Functions Hit by the Differentially Expressed Genes
3. Discussion
4. Materials and Methods
4.1. Ethics Statement of the Interventional Study
4.2. Experimental Design
- Control (n = 4), control group, where females were cervically infused with the protein-free extender BTS (50 mL);
- Mating (n = 4), where females were each mated with a single male;
- P1-AI (n = 4), where females were artificially inseminated with ejaculated P1-sperm-peak fractions (extended to 50 mL with BTS);
- SP-Ejac (n = 4), where females were cervically infused with sperm-free SP of the whole ejaculate (50 mL);
- SP-P1 (n = 4), where females were cervically infused with sperm-free SP harvested from ejaculated sperm-peak fractions (P1, pool, 50 mL).
4.3. Animals and Tissue Sources
4.4. Semen Collection, Evaluation, and SP Harvesting
4.5. Handling of Sows
4.6. Tissue Sample Preparation
4.7. Determination of Estradiol (E2) and Progesterone (P4) Concentrations
4.8. Microarray Hybridization and Scanning
4.9. Microarray Data Analysis and Bioinformatics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Martínez, H.; Kvist, U.; Saravia, F.; Wallgren, M.; Johannisson, A.; Sanz, L.; Peña, F.J.; Martínez, E.A.; Roca, J.; Vázquez, J.M.; et al. The physiological roles of the boar ejaculate. Soc. Reprod. Fertil. Suppl. 2009, 66, 1–21. [Google Scholar]
- Miller, D.J. The epic journey of sperm through the female genital tract. Animal 2018, 12, S110–S120. [Google Scholar] [CrossRef]
- Mburu, J.N.; Einarsson, S.; Lundeheim, N.; Rodriguez-Martinez, H. Distribution, number and membrane integrity of spermatozoa in the pig oviduct in relation to spontaneous ovulation. Anim. Reprod. Sci. 1996, 45, 109–121. [Google Scholar] [CrossRef]
- Viring, S.; Einarsson, S. Sperm distribution within the genital tract of naturally inseminated gilts. Nord. Vet. Med. 1981, 33, 145–149. [Google Scholar] [PubMed]
- Einarsson, S.; Jones, B.; Larsson, K.; Viring, S. Distribution of small- and medium-sized molecules within the genital tract of artificially inseminated gilts. J. Reprod. Fertil. 1980, 59, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Lovell, J.W.; Getty, R. Fate of semen in the uterus of the sow: Histologic study of endometrium during the 27 hours after natural service. Am. J. Vet. Res. 1968, 29, 609–625. [Google Scholar] [PubMed]
- Rozeboom, K.J.; Troedsson, M.H.; Crabo, B.G. Characterization of uterine leukocyte infiltration in gilts after artificial insemination. J. Reprod. Fertil. 1998, 114, 195–199. [Google Scholar] [CrossRef]
- Rozeboom, K.J.; Troedsson, M.H.; Hodson, H.H.; Shurson, G.C.; Crabo, B.G. The importance of seminal plasma on the fertility of subsequent artificial inseminations in swine. J. Anim. Sci. 2000, 78, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Schuberth, H.J.; Taylor, U.; Zerbe, H.; Waberski, D.; Hunter, R.; Rath, D. Immunological responses to semen in the female genital tract. Theriogenology 2008, 70, 1174–1181. [Google Scholar] [CrossRef]
- Hunter, R.H.; Fléchon, B.; Fléchon, J.E. Pre- and peri-ovulatory distribution of viable spermatozoa in the pig oviduct: A scanning electron microscope study. Tissue Cell 1987, 19, 423–436. [Google Scholar] [CrossRef]
- Rodriguez-Martinez, H.; Nicander, L.; Viring, S.; Einarsson, S.; Larsson, K. Ultrastructure of the uterotubal junction in preovulatory pigs. Anat. Histol. Embryol. 1990, 19, 16–36. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A. Seminal fluid signaling in the female reproductive tract: Lessons from rodents and pigs. J. Anim. Sci. 2007, 85, E36–E44. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martinez, H.; Saravia, F.; Wallgren, M.; Martinez, E.A.; Sanz, L.; Roca, J.; Vazquez, J.M.; Calvete, J.J. Spermadhesin PSP-I/PSP-II heterodimer induces migration of polymorphonuclear neutrophils into the uterine cavity of the sow. J. Reprod. Immunol. 2010, 84, 57–65. [Google Scholar] [CrossRef]
- Barranco, I.; Tvarijonaviciute, A.; Perez-Patiño, C.; Vicente-Carrillo, A.; Parrilla, I.; Ceron, J.J.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Glutathione peroxidase 5 is expressed by the entire pig male genital tract and once in the seminal plasma contributes to sperm survival and in vivo fertility. PLoS ONE 2016, 11, e0162958. [Google Scholar] [CrossRef] [PubMed]
- Barranco, I.; Perez-Patiño, C.; Tvarijonaviciute, A.; Parrilla, I.; Vicente-Carrillo, A.; Alvarez-Rodriguez, M.; Ceron, J.J.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Active paraoxonase 1 is synthesised throughout the internal boar genital organs. Reproduction 2017, 154, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Perez-Patiño, C.; Barranco, I.; Parrilla, I.; Valero, M.L.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Characterization of the porcine seminal plasma proteome comparing ejaculate portions. J. Proteomics 2016, 142, 15–23. [Google Scholar] [CrossRef]
- Perez-Patiño, C.; Parrilla, I.; Li, J.; Barranco, I.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. The proteome of pig spermatozoa is remodeled during ejaculation. Mol. Cell. Proteomics 2018. [Google Scholar] [CrossRef]
- O’Leary, S.; Armstrong, D.T.; Robertson, S.A. Transforming growth factor-β (TGFβ) in porcine seminal plasma. Reprod. Fertil. Dev. 2011, 23, 748–758. [Google Scholar] [CrossRef]
- Jiwakanon, J.; Dalin, A.-M. Short communication: Concentration of TGF-β1, IL-10 and IL-6 in boar seminal plasma and TGF-β1 level in different fractions of ejaculates. Anim. Reprod. Sci. 2012, 131, 194–198. [Google Scholar] [CrossRef]
- Barranco, I.; Rubér, M.; Perez-Patiño, C.; Atikuzzaman, M.; Martinez, E.A.; Roca, J.; Rodriguez-Martinez, H. The seminal plasma of the boar is rich in cytokines, with significant individual and intra-ejaculate variation. Am. J. Reprod. Immunol. 2015, 74, 523–532. [Google Scholar] [CrossRef]
- Claus, R.; Schopper, D.; Wagner, H.G. Seasonal effect on steroids in blood plasma and seminal plasma of boars. J. Steroid Biochem. 1983, 19, 725–729. [Google Scholar] [CrossRef]
- Zduńczyk, S.; Janowski, T.; Raś, A.; Barański, W. Concentrations of oestrogens in blood plasma and seminal plasma of boars during the postpuberal period. Pol. J. Vet. Sci. 2011, 14, 539–544. [Google Scholar] [CrossRef]
- Piehl, L.L.; Fischman, M.L.; Hellman, U.; Cisale, H.; Miranda, P.V. Boar seminal plasma exosomes: Effect on sperm function and protein identification by sequencing. Theriogenology 2013, 79, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
- Žura Žaja, I.; Samardžija, M.; Vince, S.; Vilić, M.; Majić-Balić, I.; Đuričić, D.; Milinković-Tur, S. Differences in seminal plasma and spermatozoa antioxidative systems and seminal plasma lipid and protein levels among boar breeds and hybrid genetic traits. Anim. Reprod. Sci. 2016, 170, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, H.; Shen, D.; Wang, S.; Zhang, L.; Wang, X.; Gao, B.; Wu, T.; Li, B.; Li, K.; et al. Comparative profiling of small RNAs of pig seminal plasma and ejaculated and epididymal sperm. Reproduction 2017, 153, 785–796. [Google Scholar] [CrossRef]
- Fazeli, A.; Affara, N.A.; Hubank, M.; Holt, W.V. Sperm-induced modification of the oviductal gene expression profile after natural insemination in mice. Biol. Reprod. 2004, 71, 60–65. [Google Scholar] [CrossRef]
- Georgiou, A.S.; Snijders, A.P.L.; Sostaric, E.; Aflatoonian, R.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; Martinez, E.A.; Wright, P.C.; Fazeli, A. Modulation of the oviductal environment by gametes. J. Proteome Res. 2007, 6, 4656–4666. [Google Scholar] [CrossRef]
- Almiñana, C.; Caballero, I.; Heath, P.R.; Maleki-Dizaji, S.; Parrilla, I.; Cuello, C.; Gil, M.A.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; et al. The battle of the sexes starts in the oviduct: Modulation of oviductal transcriptome by X and Y-bearing spermatozoa. BMC Genomics 2014, 15, 293. [Google Scholar] [CrossRef] [PubMed]
- López-Úbeda, R.; García-Vázquez, F.A.; Romar, R.; Gadea, J.; Muñoz, M.; Hunter, R.H.F.; Coy, P. Oviductal Transcriptome Is Modified after Insemination during Spontaneous Ovulation in the Sow. PLoS ONE 2015, 10, e0130128. [Google Scholar] [CrossRef] [PubMed]
- Atikuzzaman, M.; Mehta Bhai, R.; Fogelholm, J.; Wright, D.; Rodriguez-Martinez, H. Mating induces the expression of immune- and pH-regulatory genes in the utero-vaginal junction containing mucosal sperm-storage tubuli of hens. Reproduction 2015, 150, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Das, S.C.; Isobe, N.; Nishibori, M.; Yoshimura, Y. Expression of transforming growth factor-beta isoforms and their receptors in utero-vaginal junction of hen oviduct in presence or absence of resident sperm with reference to sperm storage. Reproduction 2006, 132, 781–790. [Google Scholar] [CrossRef]
- Das, S.C.; Isobe, N.; Yoshimura, Y. Changes in the expression of interleukin-1beta and lipopolysaccharide-induced TNF factor in the oviduct of laying hens in response to artificial insemination. Reproduction 2009, 137, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Isobe, N.; Obitsu, T.; Yoshimura, Y. Expression of lipases and lipid receptors in sperm storage tubules and possible role of fatty acids in sperm survival in the hen oviduct. Theriogenology 2016, 85, 1334–1342. [Google Scholar] [CrossRef]
- Atikuzzaman, M.; Alvarez-Rodriguez, M.; Vicente-Carrillo, A.; Johnsson, M.; Wright, D.; Rodriguez-Martinez, H. Conserved gene expression in sperm reservoirs between birds and mammals in response to mating. BMC Genomics 2017, 18, 98. [Google Scholar] [CrossRef] [PubMed]
- Bauersachs, S.; Blum, H.; Mallok, S.; Wenigerkind, H.; Rief, S.; Prelle, K.; Wolf, E. Regulation of ipsilateral and contralateral bovine oviduct epithelial cell function in the postovulation period: A transcriptomics approach. Biol. Reprod. 2003, 68, 1170–1177. [Google Scholar] [CrossRef]
- Calvete, J.J.; Ensslin, M.; Mburu, J.; Iborra, A.; Martínez, P.; Adermann, K.; Waberski, D.; Sanz, L.; Töpfer-Petersen, E.; Weitze, K.F.; et al. Monoclonal antibodies against boar sperm zona pellucida-binding protein AWN-1. Characterization of a continuous antigenic determinant and immunolocalization of AWN epitopes in inseminated sows. Biol. Reprod. 1997, 57, 735–742. [Google Scholar] [CrossRef]
- Caballero, I.; Vázquez, J.M.; Rodríguez-Martínez, H.; Gill, M.A.; Calvete, J.J.; Sanz, L.; Garcia, E.M.; Roca, J.; Martínez, E.A. Influence of seminal plasma PSP-I/PSP-II spermadhesin on pig gamete interaction. Zygote 2005, 13, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martinez, H.; Iborra, A.; Martínez, P.; Calvete, J.J. Immunoelectronmicroscopic imaging of spermadhesin AWN epitopes on boar spermatozoa bound in vivo to the zona pellucida. Reprod. Fertil. Dev. 1998, 10, 491–497. [Google Scholar] [CrossRef]
- Jolliffe, I.T.; Cadima, J. Principal component analysis: A review and recent developments. Philos. Trans. Ann. Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef]
- Lavon, U.; Boursnell, J.C. The split ejaculate of the boar: Contributions of the epididymides and seminal vesicles. J. Reprod. Fertil. 1975, 42, 541–552. [Google Scholar] [CrossRef]
- Mann, T.; Lutwak-Mann, C. Male Reproductive Function and Semen, 1st ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1981; p. 495. [Google Scholar]
- Einarsson, S. Transport of boar semen in the female reproductive tract. In Proceedings of the 1st Conference on Deep Freezing Boar Semen, Uppsala, Sweden, 25–27 August 1985; pp. 189–197. [Google Scholar]
- Matthijs, A.; Engel, B.; Woelders, H. Neutrophil recruitment and phagocytosis of boar spermatozoa after artificial insemination of sows, and the effects of inseminate volume, sperm dose and specific additives in the extender. Reproduction 2003, 125, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, D.J.; Macpherson, A.M.; Tremellen, K.P.; Mottershead, D.G.; Gilchrist, R.B.; Robertson, S.A. TGF-β mediates proinflammatory seminal fluid signaling in human cervical epithelial cells. J. Immunol. 2012, 189, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Jiwakanon, J.; Persson, E.; Dalin, A.-M. The influence of pre- and post-ovulatory insemination and early pregnancy on the infiltration by cells of the immune system in the sow oviduct. Reprod. Domest. Anim. 2006, 41, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A.; Mau, V.J.; Tremellen, K.P.; Seamark, R.F. Role of high molecular weight seminal vesicle proteins in eliciting the uterine inflammatory response to semen in mice. J. Reprod. Fertil. 1996, 107, 265–277. [Google Scholar] [CrossRef]
- Schjenken, J.E.; Robertson, S.A. Seminal fluid signalling in the female reproductive tract: Implications for reproductive success and offspring health. Adv. Exp. Med. Biol. 2015, 868, 127–158. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Ji, L.; Zhang, Y.; Zhen, Y.; Zhang, Q.; Xu, X.; Liu, B. Transcriptome differences in porcine alveolar macrophages from Tongcheng and Large White pigs in response to highly pathogenic porcine reproductive and respiratory syndrome virus (PRRSV) infection. Int. J. Mol. Sci. 2017, 18, 1475. [Google Scholar] [CrossRef] [PubMed]
- Fung, K.Y.; Mangan, N.E.; Cumming, H.; Horvat, J.C.; Mayall, J.R.; Stifter, S.A.; De Weerd, N.; Roisman, L.C.; Rossjohn, J.; Robertson, S.A.; et al. Interferon-ε protects the female reproductive tract from viral and bacterial infection. Science 2013, 339, 1088–1092. [Google Scholar] [CrossRef]
- Sharkey, D.J.; Glynn, D.J.; Schjenken, J.E.; Tremellen, K.P.; Robertson, S.A. Interferon-gamma inhibits seminal plasma induction of colony-stimulating factor 2 in mouse and human reproductive tract epithelial cells. Biol. Reprod. 2018, 99, 514–526. [Google Scholar] [CrossRef]
- Su, D.; Zhou, Y.; Hu, S.; Guan, L.; Shi, C.; Wang, Q.; Chen, Y.; Lu, C.; Li, Q.; Ma, X. Role of GAB1/PI3K/AKT signaling high glucose-induced cardiomyocyte apoptosis. Biomed. Pharmacother. 2017, 93, 1197–1204. [Google Scholar] [CrossRef]
- Phillips, T.A.; Ni, J.; Pan, G.; Ruben, S.M.; Wei, Y.F.; Pace, J.L.; Hunt, J.S. TRAIL (Apo-2L) and TRAIL receptors in human placentas: Implications for immune privilege. J. Immunol. 1999, 162, 6053–6059. [Google Scholar]
- Kaczmarek, M.M.; Krawczynski, K.; Blitek, A.; Kiewisz, J.; Schams, D.; Ziecik, A.J. Seminal plasma affects prostaglandin synthesis in the porcine oviduct. Theriogenology 2010, 74, 1207–1220. [Google Scholar] [CrossRef] [PubMed]
- Kiewisz, J.; Kaczmarek, M.M.; Morawska, E.; Blitek, A.; Kapelanski, W.; Ziecik, A.J. Estrus synchronization affects WNT signaling in the porcine reproductive tract and embryos. Theriogenology 2011, 76, 1684–1694. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, C.; Mjösberg, J.; Matussek, A.; Geffers, R.; Matthiesen, L.; Berg, G.; Sharma, S.; Buer, J.; Ernerudh, J. Gene expression profiling of human decidual macrophages: Evidence for immunosuppressive phenotype. PLoS ONE 2008, 3, e2078. [Google Scholar] [CrossRef]
- Lee, S.T.; Feng, M.; Wei, Y.; Li, Z.; Qiao, Y.; Guan, P.; Jiang, X.; Wong, C.H.; Huynh, K.; Wang, J.; et al. Protein tyrosine phosphatase UBASH3B is overexpressed in triple-negative breast cancer and promotes invasion and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 11121–11126. [Google Scholar] [CrossRef]
- Giuffrida, V.; Pezzino, F.M.; Romano, F.; Litrico, L.; Garofalo, M.R.; Nicotra, G.; Libra, M.; D’Amico, F.; Castrogiovanni, P.; Imbesi, R.; et al. Gene expression in mouse spermatogenesis during ontogenesis. Int. J. Mol. Med. 2006, 17, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Sirot, L.K.; Findlay, G.D.; Sitnik, J.L.; Frasheri, D.; Avila, F.W.; Wolfner, M.F. Molecular characterization and evolution of a gene family encoding both female- and male-specific reproductive proteins in Drosophila. Mol. Biol. Evol. 2014, 31, 1554–1567. [Google Scholar] [CrossRef]
- Mariani, L.-L.; Lozeron, P.; Théaudin, M.; Mincheva, Z.; Signate, A.; Ducot, B.; Algalarrondo, V.; Denier, C.; Adam, C.; Nicolas, G.; et al. Genotype-phenotype correlation and course of transthyretin familial amyloid polyneuropathies in France. Ann. Neurol. 2015, 78, 901–916. [Google Scholar] [CrossRef] [PubMed]
- Monestier, O.; Blanquet, V. WFIKKN1 and WFIKKN2: “Companion” proteins regulating TGFB activity. Cytokine Growth Factor Rev. 2016, 32, 75–84. [Google Scholar] [CrossRef]
- Robertson, S.A.; Ingman, W.V.; O’Leary, S.; Sharkey, D.J.; Tremellen, K.P. Transforming growth factor beta--a mediator of immune deviation in seminal plasma. J. Reprod. Immunol. 2002, 57, 109–128. [Google Scholar] [CrossRef]
- Tremellen, K.P.; Russell, P. The distribution of immune cells and macrophages in the endometrium of women with recurrent reproductive failure. II: Adenomyosis and macrophages. J. Reprod. Immunol. 2012, 93, 58–63. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, H.; Kvist, U.; Ernerudh, J.; Sanz, L.; Calvete, J.J. Seminal plasma proteins: what role do they play? Am. J. Reprod. Immunol. 2011, 66 (Suppl. 1), 11–22. [Google Scholar] [CrossRef] [PubMed]
- Barranco, I.; Tvarijonaviciute, A.; Perez-Patiño, C.; Parrilla, I.; Ceron, J.J.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. High total antioxidant capacity of the porcine seminal plasma (SP-TAC) relates to sperm survival and fertility. Sci. Rep. 2015, 5, 18538. [Google Scholar] [CrossRef] [PubMed]
- Jiwakanon, J.; Persson, E.; Kaeoket, K.; Dalin, A.-M. The sow endosalpinx at different stages of the oestrous cycle and at anoestrus: Studies on morphological changes and infiltration by cells of the immune system. Reprod. Domest. Anim. 2005, 40, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Jiwakanon, J.; Berg, M.; Persson, E.; Fossum, C.; Dalin, A.-M. Cytokine expression in the gilt oviduct: Effects of seminal plasma, spermatozoa and extender after insemination. Anim. Reprod. Sci. 2010, 119, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Latifi, Z.; Kusama, K.; Nakamura, K.; Shimada, M.; Imakawa, K. Induction of immune-related gene expression by seminal exosomes in the porcine endometrium. Biochem. Biophys. Res. Commun. 2018, 495, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Patiño, C.; Parrilla, I.; Barranco, I.; Vergara-Barberán, M.; Simó-Alfonso, E.F.; Herrero-Martínez, J.M.; Rodriguez-Martínez, H.; Martínez, E.A.; Roca, J. New in-depth analytical approach of the porcine seminal plasma proteome reveals potential fertility biomarkers. J. Proteome Res. 2018, 17, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Simonsson, A. Näringsrekommendationer och Fodermedelstabeller till svin (Nutrient and Metabolizable Energy Recommendations for Swine, in Swedish). Swedish University of Agricultural Sciences, SLU Info Printer, Uppsala, Sweden. 1994; 75. [Google Scholar]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- UniProt Consortium UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Treatment | Target | Up-Regulated Genes (log Fold Change, p < 0.05) | Down-Regulated Genes (log Fold Change, p < 0.05) |
|---|---|---|---|
| Mating | Cvx | VEGFB, GAB1, TMEM131, CELSR1, DPP4, SPATA13, CDIP1, ACSL3, BPIFC, GIMAP8, CD244 | STAT2, APOH, DLL4, FAP, GPX8, HSPA4, LITAF, LOC100518083, MARCH3, PLEKHG3, SELP, SLA-7, SLC11A1, STAT1, STAT5A, UBASH3B |
| DistUt | GAB1, CD209, LOC102161621, WEE1, HSPA4L, ACSS2, C4BPA, C4BPA, AHSA1, PPP1CC, PPP1CA, ACSL3, MARCH2, BPIFC, VEGFB, SLAMF7, GPX2 | DAB2, LOC100517400, CEBPB, STAT2, STAT1, STAT3, UBASH3B, LOC100518083, LOC100626135, TNS4, ACSL4, CEBPB, LITAF, GAB2, STAT5A, OASL, CREB3L2, TNFSF10, GPX8, LOC100623062, ACER2, MARCH3, ALOX12, LOC100156301, CEBPD, PLEKHG3, APOH, STAT2, FAP | |
| ProxUt | GAB1, HSPB1, HAGHL, HSPH1, LMX1A, LMX1B, CD1D, PTGS1, MARCH2, PLA2G4D, VEGFB | DAB2, STAT2, STAT1, MCF2, STAT3, UBASH3B, LOC100518083, ACSL4, LITAF, STAT5B, STAT5A, OASL, CREB3L2, CD38, TNFSF10, GPX8, LOC100623062, SELP, MARCH3, SLA-7, PLEKHG3, FAP | |
| UTJ | DUOX2, LOC100518524, VEGFB, GAB1, CD84, SLAMF6, AIF1, CD2, ACSL3, HVCN1, GIMAP8 | CEBPB, CD209, PRKAB2, ACSBG1, TNS4, CEBPB, LITAF, STAT5A, CREB3L2, GPX8, ADGRB2, TRAF2, PLEKHG3, VEGF, FAM43A, FAP | |
| Isth | GAB1, SLAMF7, ACSS1, SIT1, MARCH2, HVCN1, WFIKKN2, SLA-6 | SLC27A4, ACSBG1, LITAF, STAT5A, GPX8 | |
| Amp | DUOX2, LOC100518524, VEGFB, LOC100517400, GAB1, HSPB1, STAT3, DPP4, LOC100626135, TNS4, SIT1, ADGRD2, ACER2, CD1E, DLL3, LOC100156301, PTGS1, CEBPD, CEBPE, GIMAP8, WFIKKN2, SLA-6 | KIAA0319, LOC100518083, WEE1, HSPA4L, PGF, GPX8, AIFM3, ADGRB2 | |
| Inf | LOC100517400, GAB1, SLAMF7, LY9, STAT3, CELSR1, LOC102161621, LOC100626135, TNS4, ACSS2, GAB2, TNFSF10, LOC100156301, CEBPD, VEGFA, WFIKKN2, LOC100518524, VEGFB, CD84, SLAMF6 | MAPK8IP1, ACSBG1, HSPA4L, AHSA1, STAT5A, PGF, GPX8, AIFM3, ADGRB2, CSRP2, MARCH2, PRDX4, TRAF3, DLL4 | |
| P1-AI | Cvx | LOC100739854, GPX5, ACER2, GIMAP8, DAB1 | HSPA4L |
| DistUt | LMX1A, BPIFC, SLA-6 | TNS4, CREB3L2, ALOX12, PTGS1, PLEKHG3 | |
| ProxUt | DUOX2, SLAMF6, TNFSF11, LMO4, SLC11A1 , SIT1, LOC100513601, CD1D, BPIFC, HVCN1, CD244, SLA-6 | MCF2, UBASH3B, STAT5B, GPX8 | |
| UTJ | LOC100518524, VEGFB, GAB1, GPX5, CRYAB, DPP4, ACSL1, CDIP1, MARCH2, WFIKKN2 | CXCL16, CELSR1, LOC100518083, TNS4, ACSL4, HSPA4L, VAV2, CD38, ACER2, CSRP2, PLEKHG3, VEGFA, VAV1 | |
| Isth | VEGFB, GAB1, LOC100626135, GAB2, LOC100156301, BPIFC, SLA-6, FAP | HSPH1, PPP1CA | |
| Amp | VEGFB, HEPACAM, LOC100513601, DLL3, BPIFC, ADGRB2, STAT1 | HSPA4, LOC100518083, HSPH1, OASL | |
| Inf | VEGFB, LOC100517400, GAB1, LOC102161621, HEPACAM, ACSS2, C4BPA, GAB2, C4BPA, ACER2, LOC100513601, BPIFC | DUOX1, HSPA4, HSPH1, AHSA1, SLC11A1, OASL, PGF, GPX8, ADGRB2, CSRP2, CEBPE, LTA | |
| SP-Ejac | Cvx | ACSS2, C4BPA, ACER2, DAB1 | UBASH3B, CREM, ACSBG1, TNS4, MARCH3, APOH |
| DistUt | HSPA4L, PPP1CC, PRDX4, BPIFC, DAB1 | CEBPB, SLC27A4, GPX5, UBASH3B, TNS4, CEBPA, SIT1, STAT5A, MARCH3, LOC100156301, CEBPD, PLEKHG3, APOH | |
| ProxUt | PRKAB2 | DUOX2, MARCH3 | |
| UTJ | CEBPB, ACSBG1, TNS4, CEBPB, ADGRD2, TRAF2 | ||
| Isth | PCD1B, TNS4, PLA2G4D | ||
| Amp | UBASH3B | ||
| Inf | SELP, MZB1 | GPX2, HAGHL, CEBPE | |
| SP-P1 | Cvx | ACER2, LOC100513601 | GAB2 APOH |
| DistUt | LOC102161621, HSPA4L, LMX1A, PPP1CC, BPIFC | CEBPB, LY9, SLC27A4, TNFSF14, LOC100626135, LTB, TNS4, CEBPB LMO2, LTB, SIT1, MARCH3, LOC100156301, PTGS1, CEBPD, PLEKHG3 | |
| ProxUt | LOC100513601, CD1D | SELE, LMO2, SELE, GPX8, ADGRB2, MARCH3 | |
| UTJ | TNFSF4, LOC100513601, BPIFC, GIMAP8, WFIKKN2 | CEBPB, STAT3, TNS4, ADGRD2, TRAF2, CEBPD, CEBPE | |
| Isth | PCD1B, DPP4, DPP10, FAP | SLC27A4, MAPK8IP1, LOC100518083, PPP1CA, VAV1 | |
| Amp | LMX1A, LOC100513601, APOH, BPIFC | STAT1, LOC100518083, OASL | |
| Inf | LOC102161621, SELP, LOC100513601, FAP | ACSBG1, STAT5A, OASL, ADGRB2, FAM43A |
| Gene | KEGG-Pathway | FDR q < 0.05 | PCA p < 0.003 | Up-Regulated (red) or Down-Regulated (green) per Tissue Segment after Treatment (mating, P1-AI, SP-Ejac, SP-P1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cvx | DistUt | ProxUt | UTJ | Isth | Amp | Inf | ||||
| AIFM3 | Endocrine resistance | YES | YES | Mating * | Mating * | |||||
| APOH | Cholesterol metabolism | YES* | YES | Mating * | SP-P1 | |||||
| CD209 | C-type lectin receptor signaling pathway | - | YES | Mating | ||||||
| CD244 | Natural killer cell mediated cytotoxicity | - | YES | P1-AI | ||||||
| CREB3L2 | TNF, AMPK, cAMP, PI3K-Akt and cGMP-PKG signaling pathways | YES | - | Mating * | Mating * | |||||
| CRYAB | Protein processing in endoplasmic reticulum | - | YES | P1-AI | P1-AI | |||||
| DPP4 | Protein digestion and absorption | - | YES | SP-P1 | ||||||
| FAP | JAK-STAT signaling pathway, apoptosis | YES* | YES | Mating | Mating * | |||||
| GAB1 | Bacterial invasion of epithelial cells, Ras signaling pathway | YES* | YES | P1-AI | Mating * | |||||
| GAB2 | Fc epsilon RI and Fc gamma R-signaling pathways | - | YES | P1-AI | ||||||
| GPX2 | Arachidonic acid and Glutathione metabolism | YES | - | Mating * | ||||||
| GPX5 | Arachidonic acid and Glutathione metabolism | - | YES | Mating | ||||||
| GPX8 | Arachidonic acid and Glutathione metabolism | YES | YES | Mating * | Mating * | Mating * | Mating | |||
| HSPA4L | Protein processing in endoplasmic reticulum | YES* | YES | Mating */ SP-P1♯ | ||||||
| LOC100156301 | --- | YES | - | Mating | ||||||
| LOC100518524 | --- | - | YES | P1-AI | ||||||
| ADGRB2 | Cell surface receptor signaling and G-protein coupled receptor activity | YES* | YES | Mating * | Mating */ P1-AI | |||||
| LOC100739854 | --- | - | YES | P1-AI | ||||||
| LOC102161621 | --- | YES | YES | Mating | ||||||
| OASL | Human papillomavirus infection | - | YES | Mating | ||||||
| PGF | MAPK, PI3K-Akt, Rap1 and Ras signaling pathways | YES | YES | Mating * | ||||||
| PLEKHG3 | --- | YES | - | Mating * | ||||||
| PRDX4 | --- | YES | - | Mating * | ||||||
| STAT5A | JAK-STAT signaling pathway, Th1, Th2 and Th17cell differentiation | YES | - | Mating * | Mating * | |||||
| TNFSF10 | Cytokine-cytokine receptor interaction, Apoptosis, FoxO signaling pathway, Natural killer cell mediated cytotoxicity | - | YES | Mating/ P1-AI | ||||||
| TNFSF11 | Cytokine-cytokine receptor interaction, NF-kappa B signaling pathway | - | YES | Mating | ||||||
| UBASH3B | --- | YES | YES | Mating * | Mating * | Mating * | ||||
| VAV2 | Leukocyte transendothelial migration, B cell receptor, Chemokine, cAMP, Rap1, Fc epsilon RI signaling pathways, Fc gamma R-mediated phagocytosis, Natural killer cell mediated cytotoxicity, T cell receptor signaling pathway | - | YES | P1-AI | ||||||
| WFIKKN2 | Prolactin signalling pathway | - | YES | P1-AI | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez-Rodriguez, M.; Atikuzzaman, M.; Venhoranta, H.; Wright, D.; Rodriguez-Martinez, H. Expression of Immune Regulatory Genes in the Porcine Internal Genital Tract Is Differentially Triggered by Spermatozoa and Seminal Plasma. Int. J. Mol. Sci. 2019, 20, 513. https://doi.org/10.3390/ijms20030513
Alvarez-Rodriguez M, Atikuzzaman M, Venhoranta H, Wright D, Rodriguez-Martinez H. Expression of Immune Regulatory Genes in the Porcine Internal Genital Tract Is Differentially Triggered by Spermatozoa and Seminal Plasma. International Journal of Molecular Sciences. 2019; 20(3):513. https://doi.org/10.3390/ijms20030513
Chicago/Turabian StyleAlvarez-Rodriguez, Manuel, Mohammad Atikuzzaman, Heli Venhoranta, Dominic Wright, and Heriberto Rodriguez-Martinez. 2019. "Expression of Immune Regulatory Genes in the Porcine Internal Genital Tract Is Differentially Triggered by Spermatozoa and Seminal Plasma" International Journal of Molecular Sciences 20, no. 3: 513. https://doi.org/10.3390/ijms20030513
APA StyleAlvarez-Rodriguez, M., Atikuzzaman, M., Venhoranta, H., Wright, D., & Rodriguez-Martinez, H. (2019). Expression of Immune Regulatory Genes in the Porcine Internal Genital Tract Is Differentially Triggered by Spermatozoa and Seminal Plasma. International Journal of Molecular Sciences, 20(3), 513. https://doi.org/10.3390/ijms20030513