Formin-like 1 (FMNL1) Is Associated with Glioblastoma Multiforme Mesenchymal Subtype and Independently Predicts Poor Prognosis

 ,
, 
Abstract

1. Introduction
2. Results
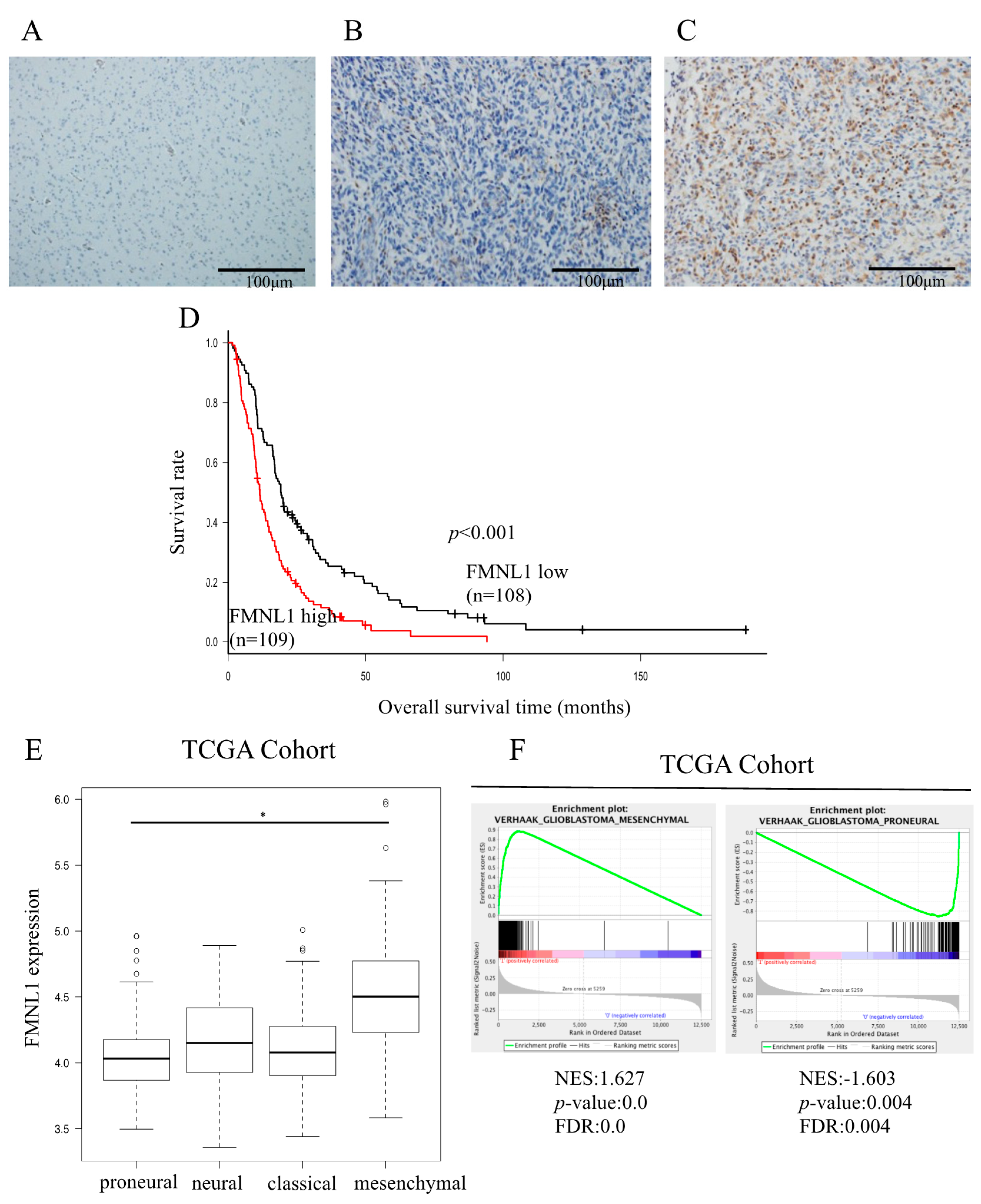
2.1. Relationship between FMNL1 Expression and Clinicopathology
2.2. Relationship between FMNL1 Expression and Prognosis
2.3. Univariate and Multivariate Survival Analyses
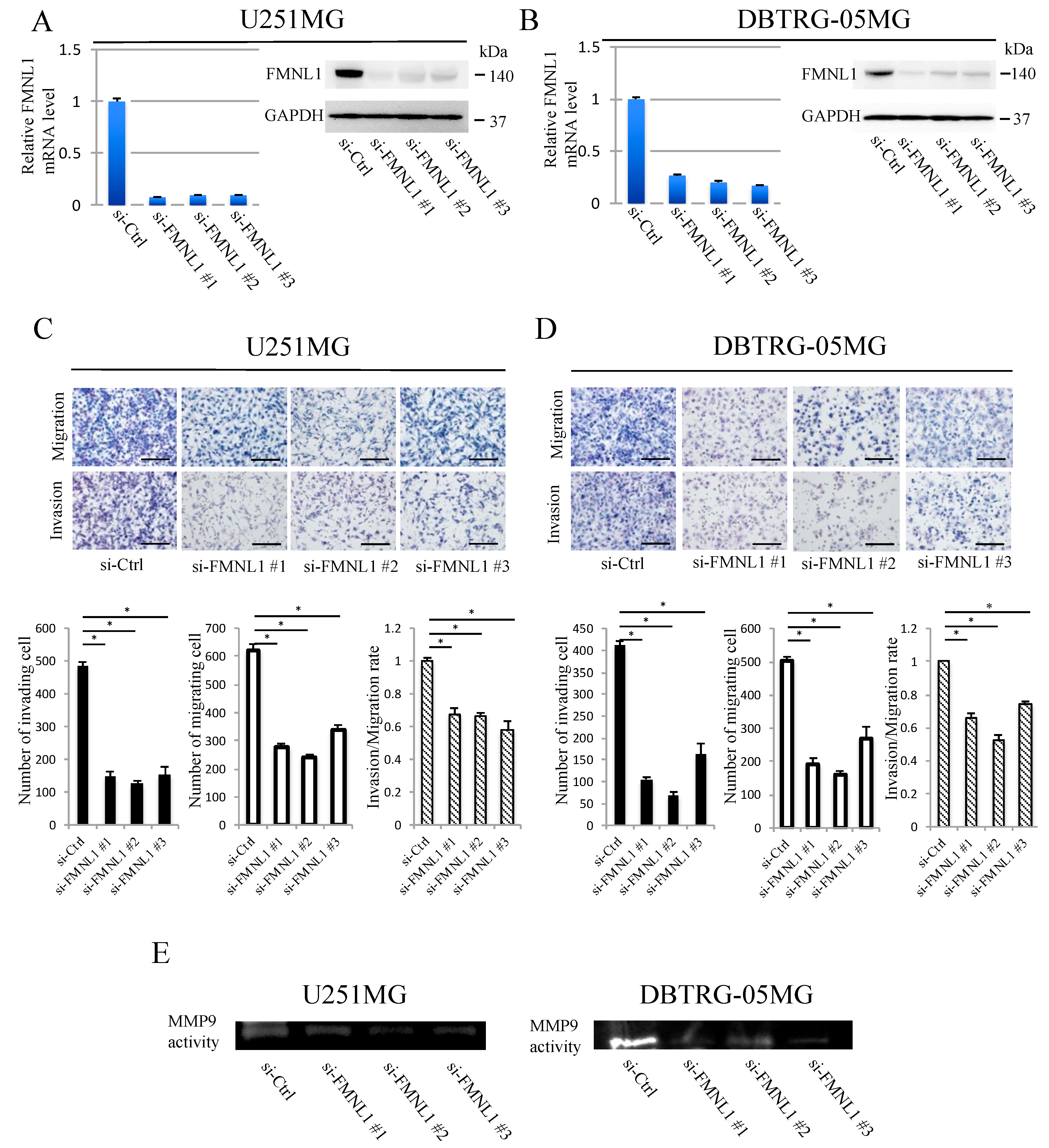
2.4. FMNL1 Knockdown Reduces GBM Migration and Invasion
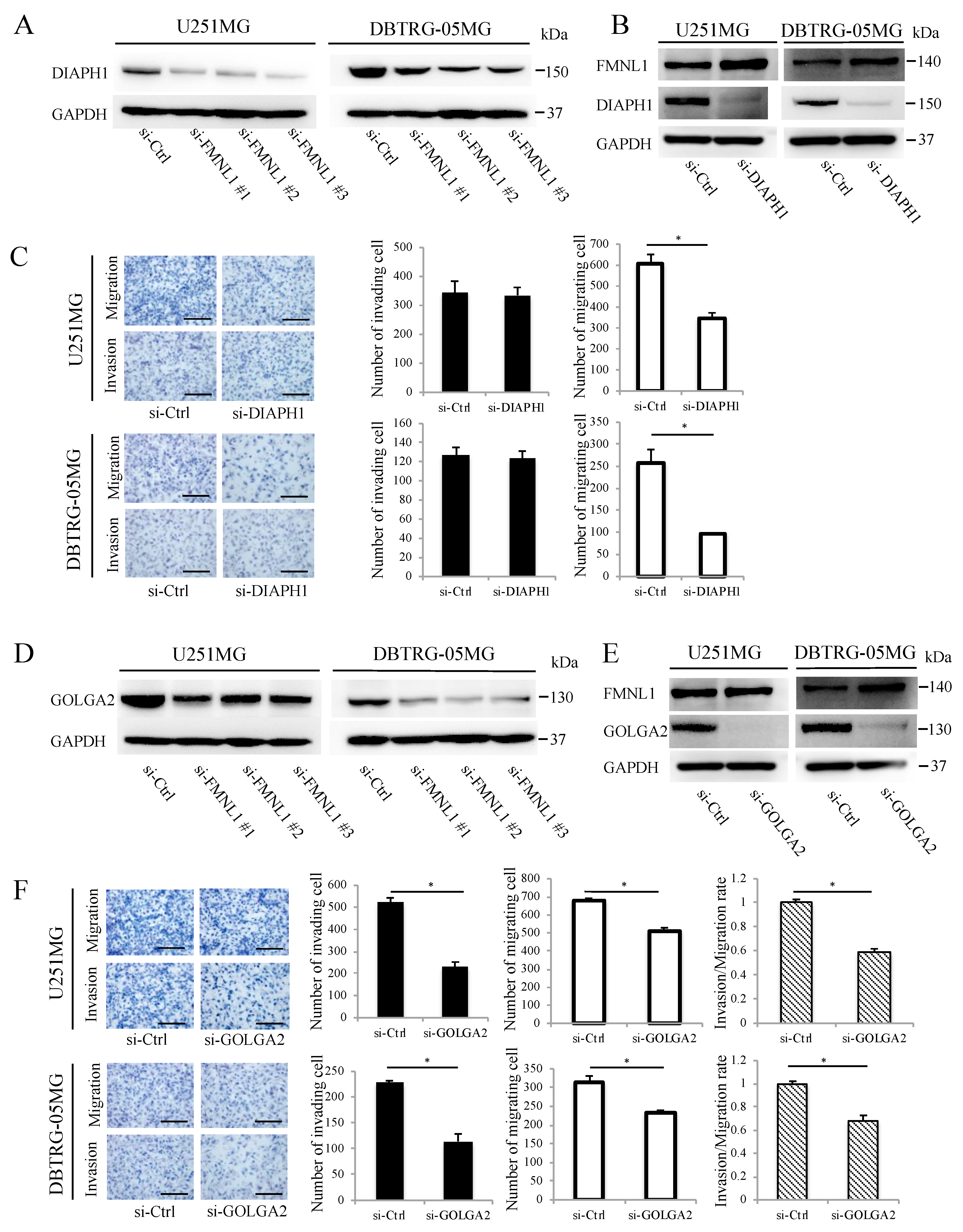
2.5. FMNL1-Mediated GBM Migration Depends on DIAPH1
2.6. FMNL1-Mediated GBM Invasion Depends on GOLGA2
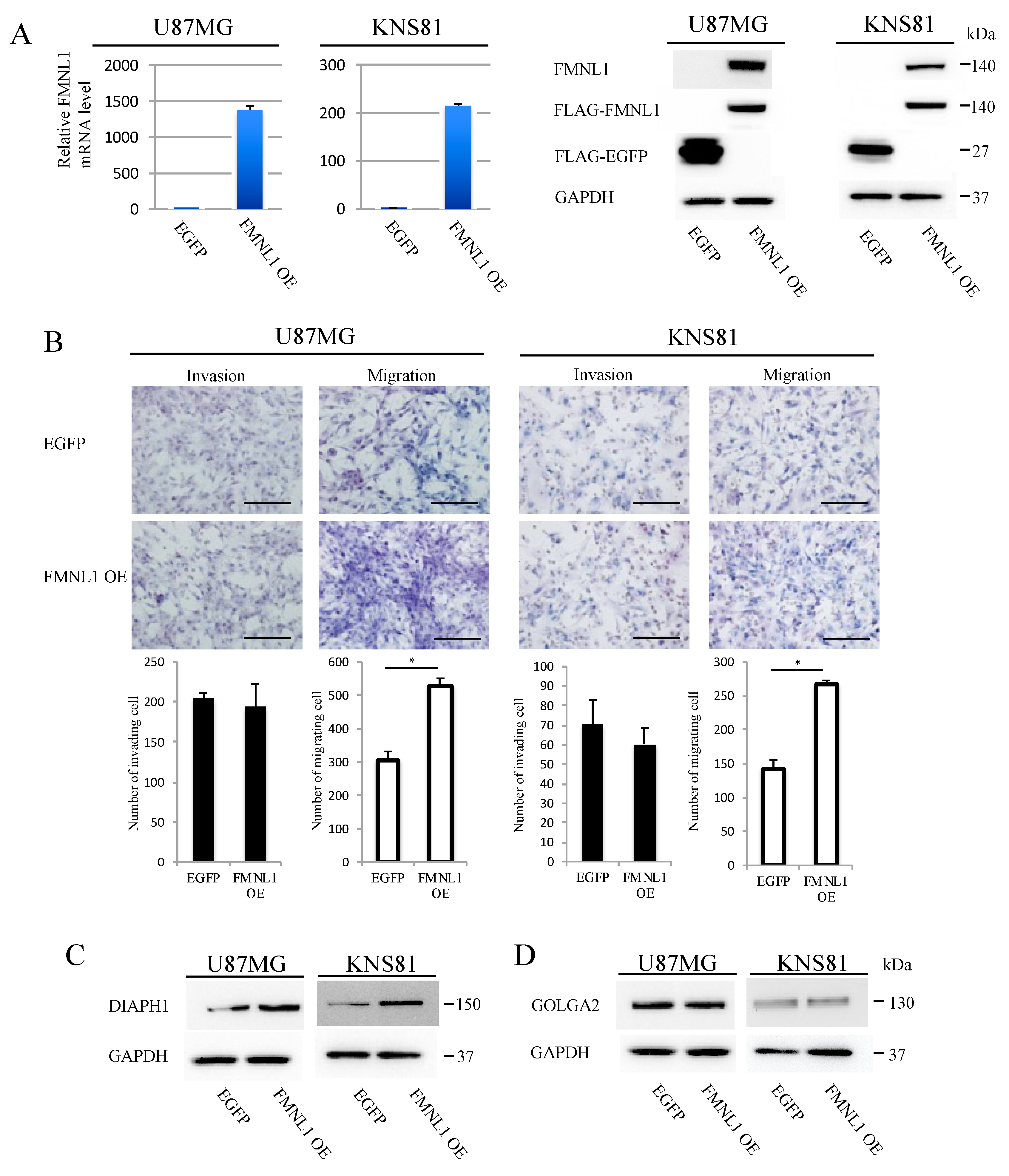
2.7. FMNL1 Overexpression Induces Migration but Not Invasion
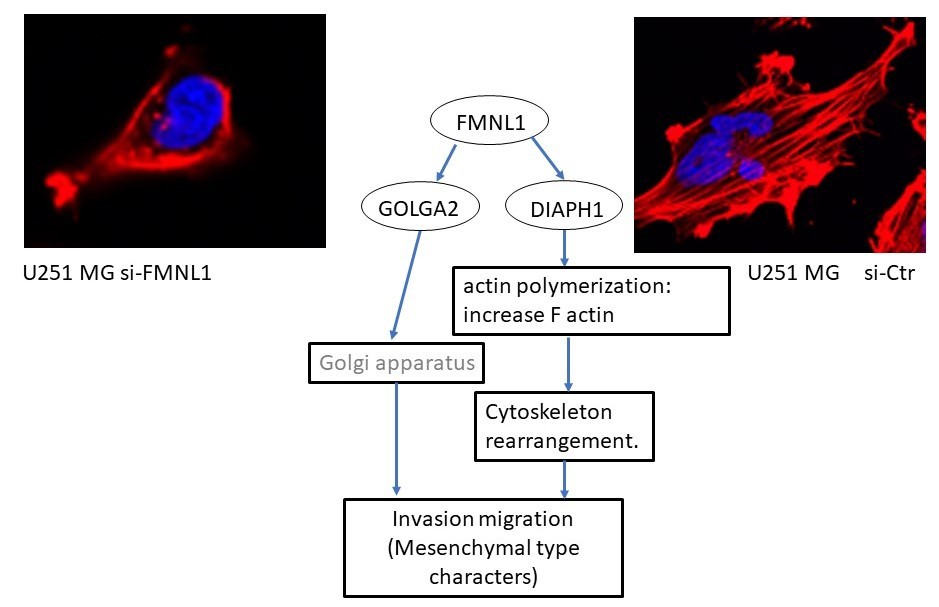
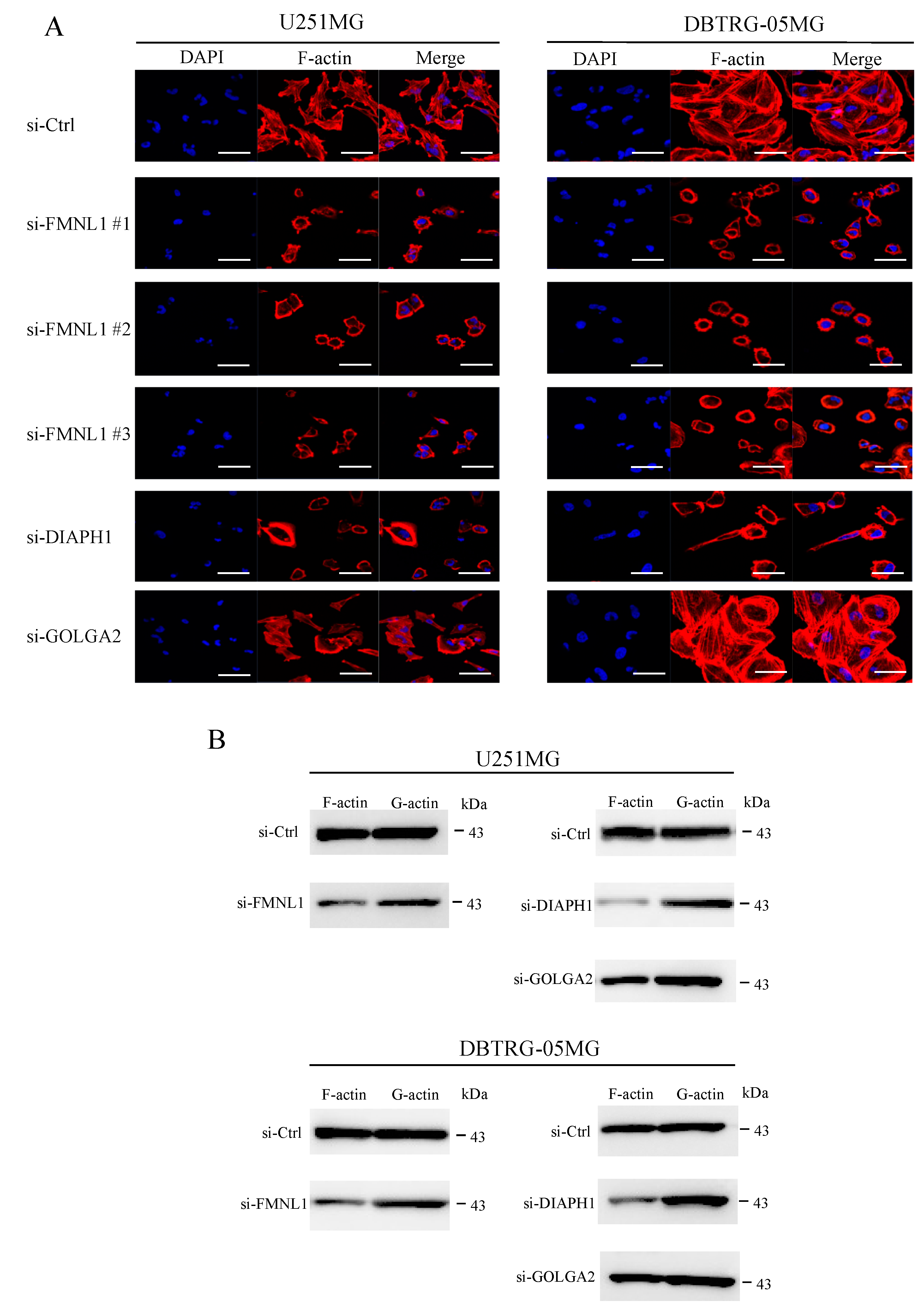
2.8. FMNL1 Knockdown Compromises the Actin Cytoskeleton via DIAPH1 but not GOLGA2
2.9. Identification of FMNL1-Associated Biological Pathways
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Patients and Tumor Samples
4.3. Immunohistochemistry
4.4. Cell Culture
4.5. siRNA Transfection
4.6. FMNL1 Lentiviral Expression Vector
4.7. Transwell Migration and Invasion Assay
4.8. Cell Proliferation Assay
4.9. Confocal Laser Microscopy
4.10. Quantification of Filamentous and Globular Actin
4.11. RNA Isolation and cDNA Synthesis
4.12. RT-PCR
4.13. Protein Extraction and Western Blotting
4.14. In Silico Analyses
4.15. Gelatin Zymography Assay
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FMNL1 | formin-like 1 |
| TCGA | The Cancer Genome Atlas |
| GSEA | gene set enrichment analysis |
| GBM | Glioblastoma multiforme |
| KPS | Karnofsky performance status |
References
- Rick, J.; Chandra, A.; Aghi, M.K. Tumor treating fields: A new approach to glioblastoma therapy. J. Neurooncol. 2018, 137, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Woernle, C.M.; Péus, D.; Hofer, S.; Rushing, E.J.; Held, U.; Bozinov, O.; Krayenbühl, N.; Weller, M.; Regli, L. Efficacy of surgery and further treatment of progressive glioblastoma. World Neurosurg. 2015, 84, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Altieri, R.; Zenga, F.; Fontanella, M.M.; Cofano, F.; Agnoletti, A.; Spena, G.; Crobeddu, E.; Fornaro, R.; Ducati, A.; Garbossa, D. Glioma surgery: Technological advances to achieve a maximal safe resection. Surg. Technol. Int. 2015, 27, 297–302. [Google Scholar]
- Chesarone, M.A.; Dupage, A.G.; Goode, B.L. Unleashing formins to remodel the actin and microtubule cytoskeletons. Nat. Rev. Mol. Cell Biol. 2010, 11, 62–74. [Google Scholar] [CrossRef]
- Higgs, H.N.; Peterson, K.J. Phylogenetic analysis of the formin homology 2 domain. Mol. Biol. Cell 2005, 16, 1–13. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, L.; Duan, X.; Zhang, G.L.; Wang, Z.B.; Wang, Q.; Xiong, B.; Sun, S.C. RhoA-mediated FMNL1 regulates GM130 for actin assembly and phosphorylates MAPK for spindle formation in mouse oocyte meiosis. Cell Cycle 2015, 14, 2835–2843. [Google Scholar] [CrossRef]
- Colon-Franco, J.M.; Gomez, T.S.; Billadeau, D.D. Dynamic remodelling of the actin cytoskeleton by FMNL1 is required for structural maintenance of the Golgi complex. J. Cell Sci. 2011, 124, 3118–3126. [Google Scholar] [CrossRef]
- Seth, A.; Otomo, C.; Rosen, M.K. Autoinhibition regulates cellular localization and actin assembly activity of the diaphanous-related formins FRLα and mDia1. J. Cell Biol. 2006, 174, 701–713. [Google Scholar] [CrossRef]
- Favaro, P.M.; Traina, F.; Vassallo, J.; Brousset, P.; Delsol, G.; Costa, F.F.; Saad, S.T. High expression of FMNL1 protein in T non-Hodgkin’s lymphomas. Leuk. Res. 2006, 30, 735–738. [Google Scholar] [CrossRef]
- Favaro, P.M.; de Souza Medina, S.; Traina, F.; Bassères, D.S.; Costa, F.F.; Saad, S.T. Human leukocyte formin: A novel protein expressed in lymphoid malignancies and associated with Akt. Biochem. Biophys. Res. Commun. 2003, 311, 365–371. [Google Scholar] [CrossRef]
- Favaro, P.; Traina, F.; Machado-Neto, J.A.; Lazarini, M.; Lopes, M.R.; Pereira, J.K.; Costa, F.F.; Infante, E.; Ridley, A.J.; Saad, S.T. FMNL1 promotes proliferation and migration of leukemia cells. J. Leukoc. Biol. 2013, 94, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Cai, M.Y.; Zhang, J.X.; Wang, F.W.; Tang, L.Q.; Liao, Y.J.; Jin, X.H.; Wang, C.Y.; Guo, L.; Jiang, Y.G.; et al. FMNL1 mediates nasopharyngeal carcinoma cell aggressiveness by epigenetically upregulating MTA1. Oncogene 2018, 37, 6243–6258. [Google Scholar] [CrossRef] [PubMed]
- Péladeau, C.; Heibein, A.; Maltez, M.T.; Copeland, S.J.; Copeland, J.W. A specific FMNL2 isoform is up-regulated in invasive cells. BMC Cell Biol. 2016, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.L.; Liang, L.; Ding, Y.Q. Overexpression of FMNL2 is closely related to metastasis of colorectal cancer. Int. J. Colorectal Dis. 2008, 23, 1041–1047. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, X.; Zeng, Y.; Wang, J.; Zhang, X.; Ding, Y.Q.; Liang, L. FMNL2 enhances invasion of colorectal carcinoma by inducing epithelial-mesenchymal transition. Mol. Cancer Res. 2010, 8, 1579–1590. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, Z.; Wang, K.; Ha, Y.; Lei, H.; Jia, Y.; Ding, R.; Wu, D.; Gan, S.; Li, R.; et al. High FMNL3 expression promotes nasopharyngeal carcinoma cell metastasis: Role in TGF-β1-induced epithelia-to-mesenchymal transition. Sci. Rep. 2010, 7, 1–14. [Google Scholar] [CrossRef]
- Zeng, Y.F.; Xiao, Y.S.; Lu, M.Z.; Luo, X.J.; Hu, G.Z.; Deng, K.Y.; Wu, X.M.; Xin, H.B. Increased expression of formin-like 3 contributes to metastasis and poor prognosis in colorectal carcinoma. Exp. Mol. Pathol. 2015, 98, 260–267. [Google Scholar] [CrossRef]
- Gardberg, M.; Heuser, V.D.; Koskivuo, I.; Koivisto, M.; Carpén, O. FMNL2/FMNL3 formins are linked with oncogenic pathways and predict melanoma outcome. J. Pathol. Clin. Res. 2016, 2, 41–52. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, F.; Niu, Y.J.; Liu, H.L.; Rui, R.; Cui, X.S.; Kim, N.H.; Sun, S.C. Formin mDia1, a downstream molecule of FMNL1, regulates Profilin1 for actin assembly and spindle organization during mouse oocyte meiosis. Biochem. Biophys. Acta Mol. Cell Res. 2015, 1853, 317–327. [Google Scholar] [CrossRef]
- Phillips, H.S.; Kharbanda, S.; Chen, R.; Forrest, W.F.; Soriano, R.H.; Wu, T.D.; Misra, A.; Nigro, J.M.; Colman, H.; Soroceanu, L.; et al. Molecular subclasses of high-grade glioma predict prognosis, delineate a pattern of disease progression, and resemble stages in neurogenesis. Cancer Cell 2006, 9, 157–173. [Google Scholar] [CrossRef]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.P.L.; Balasubramaniyan, V.; Vaillant, B.; Ezhilarasan, R.; Hummelink, K.; Hollingsworth, F.; Wani, K.; Heathcock, L.; James, J.D.; Goodman, L.D.; et al. Mesenchymal differentiation mediated by NF-κB promotes radiation resistance in glioblastoma. Cancer Cell 2013, 24, 331–346. [Google Scholar] [CrossRef]
- Carro, M.S.; Lim, W.K.; Alvarez, M.J.; Bollo, R.J.; Zhao, X.; Snyder, E.Y.; Sulman, E.P.; Anne, S.L.; Doetsch, F.; Colman, H.; et al. The transcriptional network for mesenchymal transformation of brain tumours. Nature 2010, 463, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Murata, H.; Yoshimoto, K.; Hatae, R.; Akagi, Y.; Mizoguchi, M.; Hata, N.; Kuga, D.; Nakamizo, A.; Amano, T.; Sayama, T.; et al. Detection of proneural/mesenchymal marker expression in glioblastoma: Temporospatial dynamics and association with chromatin-modifying gene expression. J. Neurooncol. 2015, 125, 33–41. [Google Scholar] [CrossRef]
- Pollard, T.D.; Cooper, J.A. Actin, a central player in cell shape and movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef]
- Gardberg, M.; Heuser, V.D.; Iljin, K.; Kampf, C.; Uhlen, M.; Carpén, O. Characterization of leukocyte formin FMNL1 expression in human tissues. J. Histochem. Cytochem. 2014, 62, 460–470. [Google Scholar] [CrossRef]
- Stark, A.M.; Van de Bergh, J.; Hedderich, J.; Mehdorn, H.M.; Nabavi, A. Glioblastoma: Clinical characteristics, prognostic factors and survival in 492 patients. Clin. Neurol. Neurosurg. 2012, 114, 840–845. [Google Scholar] [CrossRef]
- Filippini, G.; Falcone, C.; Boiardi, A.; Broggi, G.; Bruzzone, M.G.; Caldiroli, D.; Farina, R.; Farinotti, M.; Fariselli, L.; Finocchiaro, G.; et al. Prognostic factors for survival in 676 consecutive patients with newly diagnosed primary glioblastoma. Neuro-Oncology 2008, 10, 79–87. [Google Scholar] [CrossRef]
- Lamborn, K.R.; Chang, S.M.; Prados, M.D. Prognostic factors for survival of patients with glioblastoma: Recursive partitioning analysis. Neuro-Oncology 2004, 6, 227–235. [Google Scholar] [CrossRef]
- Cheng, W.Y.; Kandel, J.J.; Yamashiro, D.J.; Canoll, P.; Anastassiou, D. A multi-cancer mesenchymal transition gene expression signature is associated with prolonged time to recurrence in glioblastoma. PLoS ONE 2012, 7, e34705. [Google Scholar] [CrossRef]
- Bezanilla, M.; Gladfelter, A.S.; Kovar, D.R.; Lee, W.L. Cytoskeletal dynamics: A view from the membrane. J. Cell Biol. 2015, 209, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Kühn, S.; Geyer, M. Formins as effector proteins of Rho GTPases. Small GTPases 2014, 5, e983876. [Google Scholar] [CrossRef] [PubMed]
- Rezania, S.; Kammerer, S.; Li, C.; Steinecker-Frohnwieser, B.; Gorischek, A.; DeVaney, T.T.J.; Verheyen, S.; Passegger, C.A.; Tabrizi-Wizsy, N.G.; Hackl, H.; et al. Overexpression of KCNJ3 gene splice variants affects vital parameters of the malignant breast cancer cell line MCF-7 in an opposing manner. BMC Cancer 2016, 16, 628. [Google Scholar] [CrossRef] [PubMed]
- Srebrow, A.; Kornblihtt, A.R. The connection between splicing and cancer. J. Cell Sci. 2006, 119, 2635–2641. [Google Scholar] [CrossRef]
- Narumiya, S.; Tanji, M.; Ishizaki, T. Rho signaling, ROCK and mDia1, in transformation, metastasis and invasion. Cancer Metastasis Rev. 2009, 28, 65–76. [Google Scholar] [CrossRef]
- Yamana, N.; Arakawa, Y.; Nishino, T.; Kurokawa, K.; Tanji, M.; Itoh, R.E.; Monypenny, J.; Ishizaki, T.; Bito, H.; Nozaki, K.; et al. The Rho-mDia1 pathway regulates cell polarity and focal adhesion turnover in migrating cells through mobilizing Apc and c-Src. Mol. Cell. Biol. 2006, 26, 6844–6858. [Google Scholar] [CrossRef]
- Cheng, L.; Xu, J.; Qian, Y.Y.; Pan, H.Y.; Yang, H.; Shao, M.Y.; Cheng, R.; Hu, T. Interaction between mDia1 and ROCK in Rho-induced migration and adhesion of human dental pulp cells. Int. Endod. J. 2017, 50, 15–23. [Google Scholar] [CrossRef]
- Zilberman, Y.; Alieva, N.O.; Miserey-Lenkei, S.; Lichtenstein, A.; Kam, Z.; Sabanay, H.; Bershadsky, A. Involvement of the Rho-mDia1 pathway in the regulation of Golgi complex architecture and dynamics. Mol. Biol. Cell 2011, 22, 2900–2911. [Google Scholar] [CrossRef]
- Lin, Y.N.; Izbicki, J.R.; König, A.; Habermann, J.K.; Blechner, C.; Lange, T.; Schumacher, U.; Windhorst, S. Expression of DIAPH1 is up-regulated in colorectal cancer and its down-regulation strongly reduces the metastatic capacity of colon carcinoma cells. Int. J. Cancer 2014, 134, 1571–1582. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, L.; Chen, J.; Liang, J.; Xu, Y.; Li, Z.; Chen, F.; Du, D. Knockdown of Diaph1 expression inhibits migration and decreases the expression of MMP2 and MMP9 in human glioma cells. Biomed. Pharmacother. 2017, 96, 596–602. [Google Scholar] [CrossRef]
- Lizárraga, F.; Poincloux, R.; Romao, M.; Montagnac, G.; Le Dez, G.; Bonne, I.; Rigaill, G.; Raposo, G.; Chavrier, P. Diaphanous-related formins are required for invadopodia formation and invasion of breast tumor cells. Cancer Res. 2009, 69, 2792–2800. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N. Emerging new roles of GM130, a cis-Golgi matrix protein, in higher order cell functions. J. Pharmacol. Sci. 2010, 112, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Rabouille, C.; Watson, R.; Nilsson, T.; Hui, N.; Slusarewicz, P.; Kreis, T.E.; Warren, G. Characterization of a cis-Golgi matrix protein, GM130. J. Cell Biol. 1995, 131, 1715–1726. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Hong, S.H.; Jiang, H.L.; Minai-Tehrani, A.; Yu, K.N.; Lee, J.H.; Kim, J.E.; Shin, J.Y.; Kang, B.; Park, S.; et al. GOLGA2/GM130, cis-Golgi matrix protein, is a novel target of anticancer gene therapy. Mol. Ther. 2012, 20, 2052–2063. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yang, C.; Guo, S.; Wu, Y. GM130 regulates epithelial-to-mesenchymal transition and invasion of gastric cancer cells via snail. Int. J. Clin. Exp. Pathol. 2015, 8, 10784–10791. [Google Scholar] [PubMed]
- Hirano, H.; Yonezawa, H.; Yunoue, S.; Habu, M.; Uchida, H.; Yoshioka, T.; Kishida, S.; Kishida, M.; Oyoshi, T.; Fujio, S.; et al. Immunoreactivity of Wnt5a, Fzd2, Fzd6, and Ryk in glioblastoma: Evaluative methodology for DAB chromogenic immunostaining. Brain Tumor Pathol. 2014, 31, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Shinsato, Y.; Furukawa, T.; Yunoue, S.; Yonezawa, H.; Minami, K.; Nishizawa, Y.; Ikeda, R.; Kawahara, K.; Yamamoto, M.; Hirano, H.; et al. Reduction of MLH1 and PMS2 confers temozolomide resistance and is associated with recurrence of glioblastoma. Oncotarget 2013, 4, 2261–2270. [Google Scholar] [CrossRef]
- Che, X.F.; Zheng, C.L.; Owatari, S.; Mutoh, M.; Gotanda, T.; Jeung, H.C.; Furukawa, T.; Ikeda, R.; Yamamoto, M.; Haraguchi, M.; et al. Overexpression of survivin in primary ATL cells and sodium arsenite induces apoptosis by down-regulating survivin expression in ATL cell lines. Blood 2006, 107, 4880–4887. [Google Scholar] [CrossRef]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Univariate Analysis | Multivariate Analysis | |||||
|---|---|---|---|---|---|---|
| HR | 95% CI | p-Value | HR | 95% CI | p-Value | |
| Sex | 1.081 | 0.82–1.43 | 0.589 | 0.957 | 0.71–1.29 | 0.773 |
| Age | 1.008 | 1.01–1.03 | < 0.001 | 1.013 | 1.00–1.02 | 0.009 |
| KPS (> 70) | 0.522 | 0.39–0.70 | < 0.001 | 0.749 | 0.55–1.03 | 0.072 |
| Ki-67 (> 30%) | 0.986 | 0.74–1.31 | 0.919 | 0.794 | 0.59–1.07 | 0.132 |
| Extent of surgical resection (Total + Subtotal/Partial + Biopsy) | 0.466 | 0.35–0.62 | < 0.001 | 0.631 | 0.46–0.86 | 0.004 |
| Number of surgeries (Single/Multiple) | 1.685 | 1.14–2.50 | 0.009 | 1.329 | 0.85–2.07 | 0.208 |
| Chemotherapy (Yes/No) | 0.202 | 0.13–0.31 | < 0.001 | 0.356 | 0.20–0.64 | < 0.001 |
| Bevacizumab (Yes/No) | 0.716 | 0.51–1.00 | 0.051 | 0.743 | 0.53–1.04 | 0.087 |
| Radiation therapy | 0.979 | 0.97–0.98 | < 0.001 | 0.987 | 0.98–0.99 | < 0.001 |
| FMNL1 expression (High/Low) | 0.534 | 0.40–0.71 | < 0.001 | 0.631 | 0.46–0.86 | < 0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Higa, N.; Shinsato, Y.; Kamil, M.; Hirano, T.; Takajo, T.; Shimokawa, M.; Minami, K.; Yamamoto, M.; Kawahara, K.; Yonezawa, H.; et al. Formin-like 1 (FMNL1) Is Associated with Glioblastoma Multiforme Mesenchymal Subtype and Independently Predicts Poor Prognosis. Int. J. Mol. Sci. 2019, 20, 6355. https://doi.org/10.3390/ijms20246355
Higa N, Shinsato Y, Kamil M, Hirano T, Takajo T, Shimokawa M, Minami K, Yamamoto M, Kawahara K, Yonezawa H, et al. Formin-like 1 (FMNL1) Is Associated with Glioblastoma Multiforme Mesenchymal Subtype and Independently Predicts Poor Prognosis. International Journal of Molecular Sciences. 2019; 20(24):6355. https://doi.org/10.3390/ijms20246355
Chicago/Turabian StyleHiga, Nayuta, Yoshinari Shinsato, Muhammad Kamil, Takuro Hirano, Tomoko Takajo, Michiko Shimokawa, Kentaro Minami, Masatatsu Yamamoto, Kohichi Kawahara, Hajime Yonezawa, and et al. 2019. "Formin-like 1 (FMNL1) Is Associated with Glioblastoma Multiforme Mesenchymal Subtype and Independently Predicts Poor Prognosis" International Journal of Molecular Sciences 20, no. 24: 6355. https://doi.org/10.3390/ijms20246355
APA StyleHiga, N., Shinsato, Y., Kamil, M., Hirano, T., Takajo, T., Shimokawa, M., Minami, K., Yamamoto, M., Kawahara, K., Yonezawa, H., Hirano, H., Furukawa, T., Yoshimoto, K., & Arita, K. (2019). Formin-like 1 (FMNL1) Is Associated with Glioblastoma Multiforme Mesenchymal Subtype and Independently Predicts Poor Prognosis. International Journal of Molecular Sciences, 20(24), 6355. https://doi.org/10.3390/ijms20246355