Proteome Alterations in Equine Osteochondrotic Chondrocytes


 , ,
, , 

Abstract
1. Introduction
2. Results
2.1. 2DE Analysis of Proteins Extracted from OC and Normal Chondrocytes
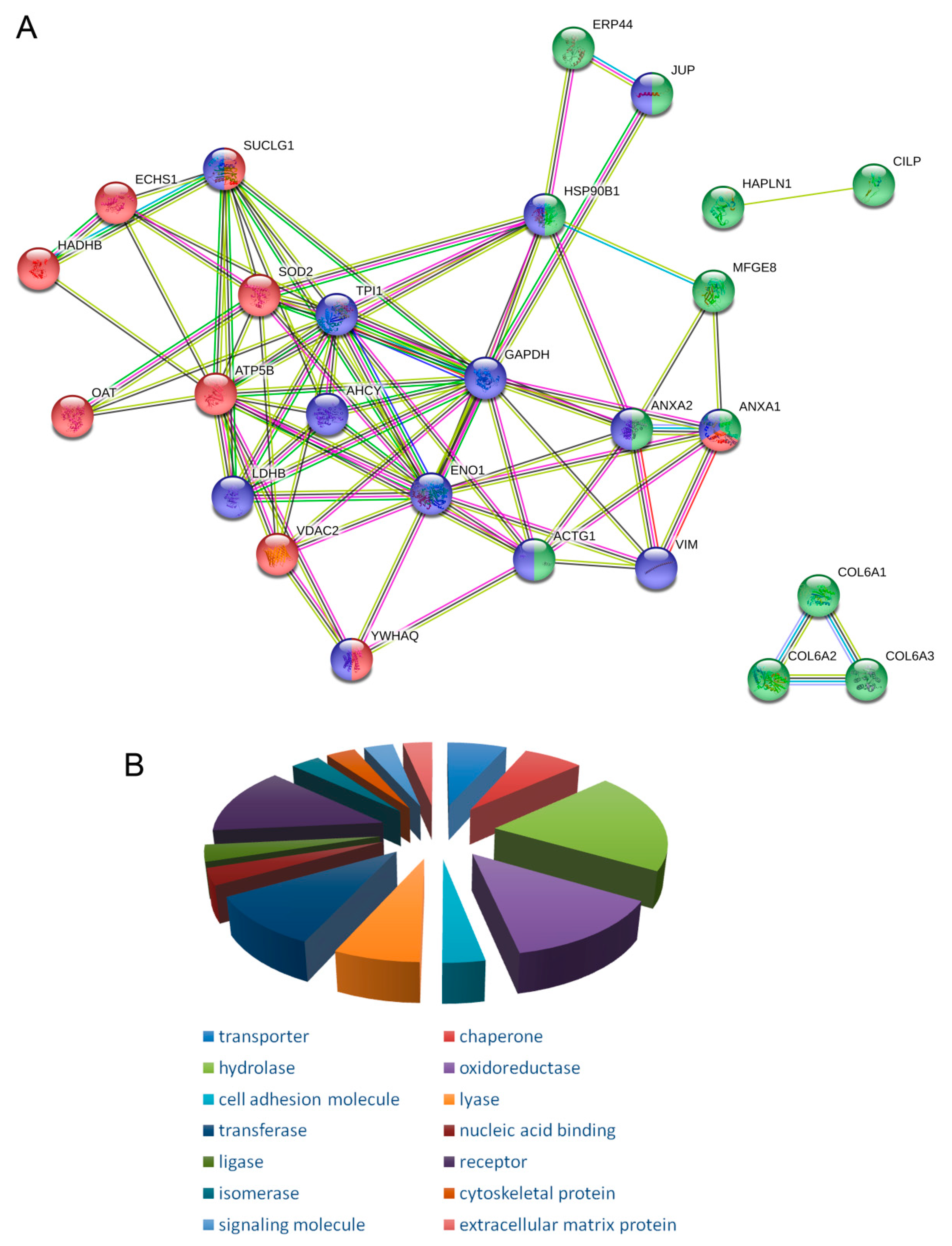
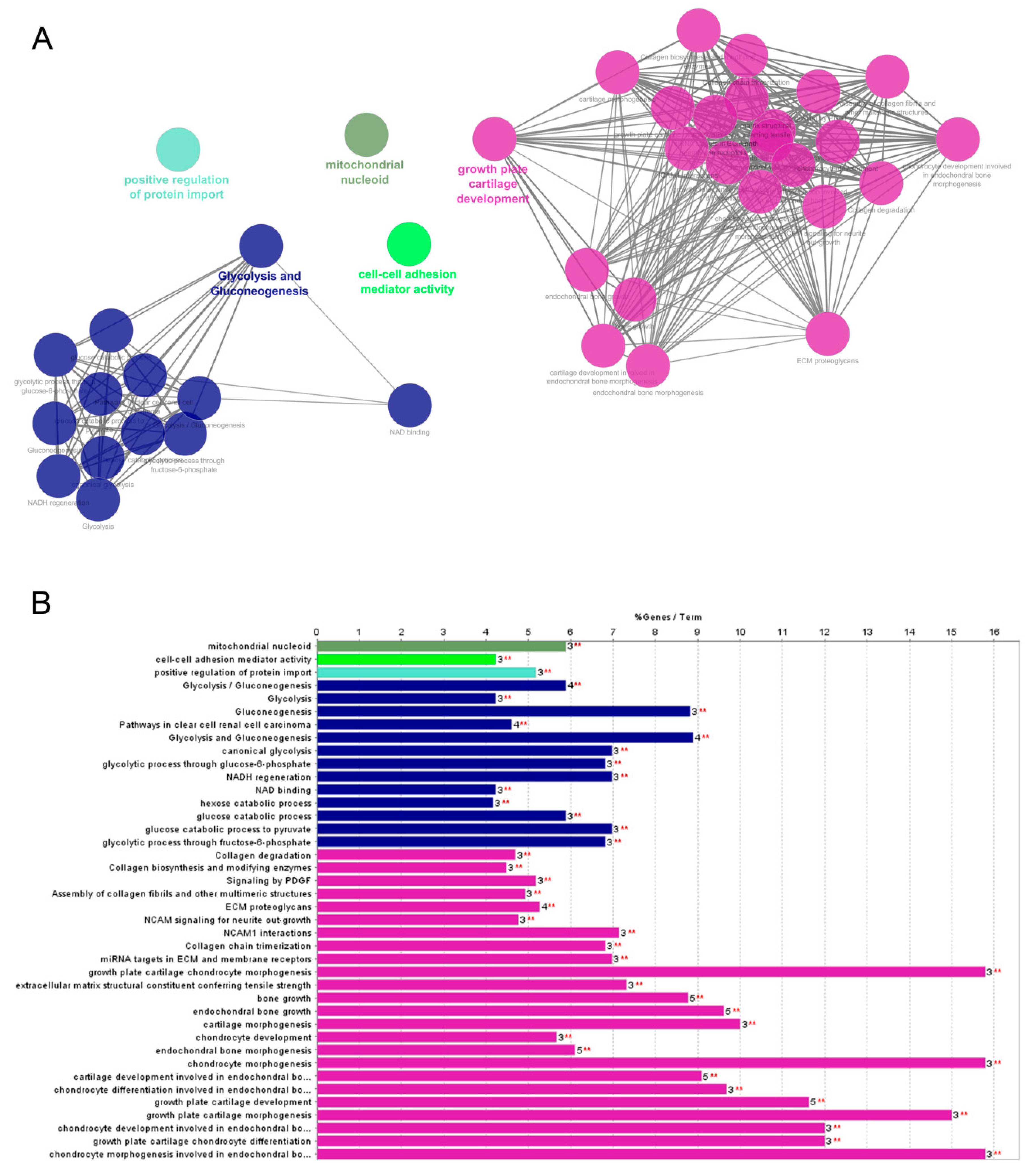
2.2. Protein Enrichment Analysis
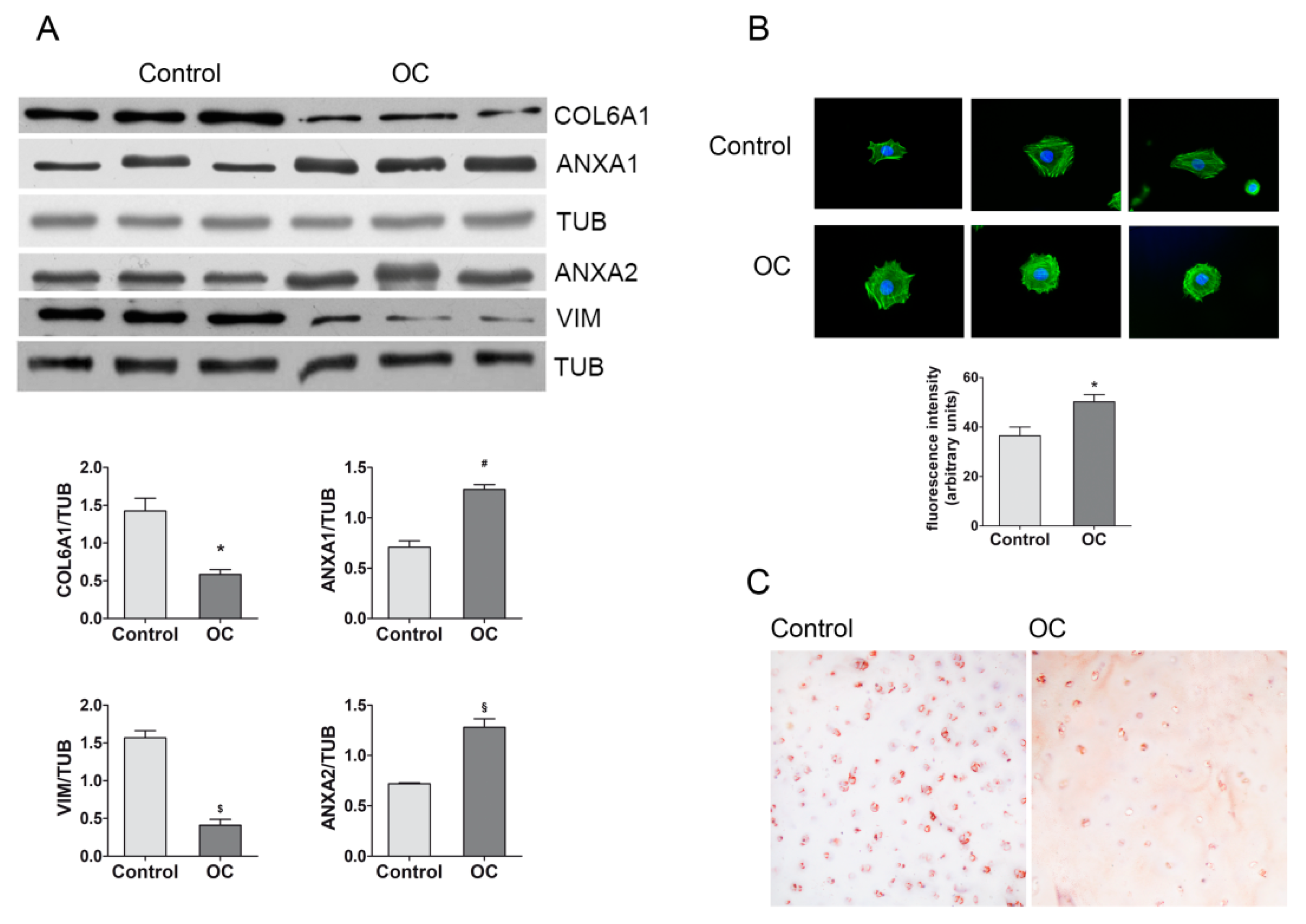
2.3. Validation of Proteomics Results
3. Discussion
4. Materials and Methods
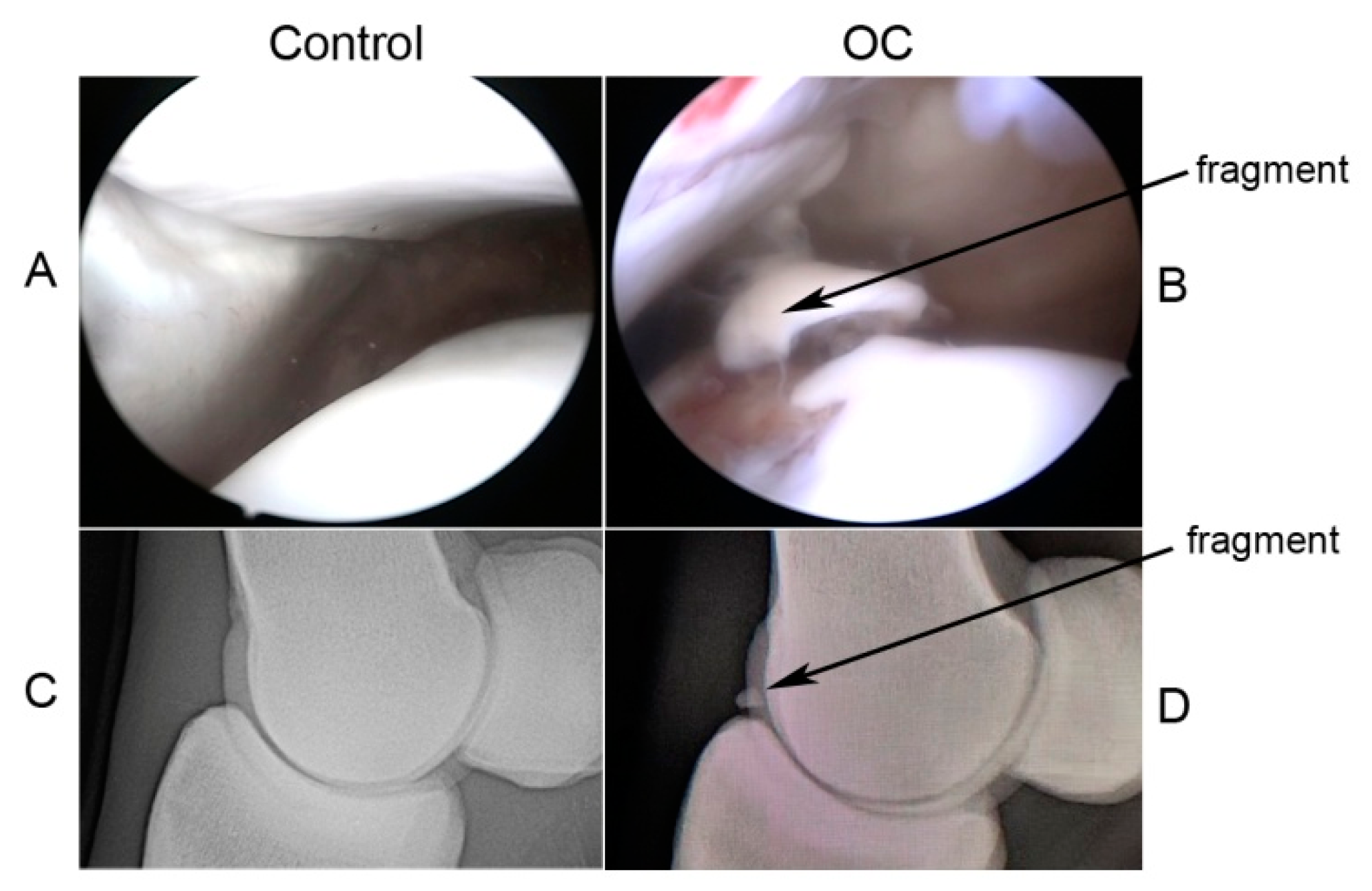
4.1. Cartilage Sampling and Processing
4.2. 2D-Gel Electrophoresis
4.3. Protein Digestion and MS Analysis
4.4. Protein Enrichment Analysis
4.5. Validation of Proteomics Results
4.5.1. Western Blotting
4.5.2. Actin Labeling by Fluorescently Labeled Phalloidin
4.5.3. Immunohistochemical Evaluation of COL6A1 in Cartilage
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| YWHAQ | 14-3-3 protein theta |
| ACTG1 | Actin, cytoplasmic 1 |
| AHCY | Adenosylhomocysteinase |
| ANXA1 | Annexin A1 |
| ANXA2 | Annexin A2 |
| ATP5F1B | ATP synthase subunit beta |
| CILP | Cartilage intermediate layer protein |
| COL6A1 | Collagen alpha-1(VI) chain |
| COL6A2 | Collagen alpha-2(VI) chain |
| COL6A3 | Collagen alpha-3(VI) chain |
| ERP44 | Endoplasmic reticulum resident protein 44 |
| HSP90B1 | Endoplasmin |
| ENO1 | Enolase 1 |
| ECHS1 | Enoyl-CoA hydratase |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| HAPLN1 | Hyaluronan and proteoglycan link protein 1 |
| JUP | Junction plakoglobin |
| MFGE8 | Lactadherin |
| LDHB | L-lactate dehydrogenase |
| OAT | Ornithine aminotransferase |
| SUCLG1 | Succinyl-CoA ligase |
| SOD | Superoxide dismutase |
| HADHB | Trifunctional enzyme mitochondrial. |
| TPI1 | Triosephosphate isomerase |
| VIM | Vimentin |
| VDAC2 | Voltage dependent anion channel 2 |
| ECM | Extracellular matrix |
| OC | Osteochondrosis |
| OA | Osteoarthritis |
References
- Ytrehus, B.; Carlson, C.S.; Ekman, S. Etiology and pathogenesis of osteochondrosis. Vet. Pathol. 2007, 44, 429–448. [Google Scholar] [CrossRef]
- Bates, J.T.; Jacobs, J.C.; Shea, K.G.; Oxford, J.T. Emerging genetic basis of osteochondritis dissecans. Clin. Sports Med. 2014, 33, 199–220. [Google Scholar] [CrossRef] [PubMed]
- Naccache, F.; Metzger, J.; Distl, O. Genetic risk factors for osteochondrosis in various horse breeds. Equine Vet. J. 2018, 50, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Van Weeren, P.R.; Jeffcott, L.B. Problems and pointers in osteochondrosis: Twenty years on. Vet. J. 2013, 197, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Van Weeren, R. Fifty years of osteochondrosis. Equine Vet. J. 2018, 50, 554–555. [Google Scholar] [CrossRef]
- Olstad, K.; Ekman, S.; Carlson, C.S. An Update on the Pathogenesis of Osteochondrosis. Vet. Pathol. 2015, 52, 785–802. [Google Scholar] [CrossRef]
- McCoy, A.M.; Toth, F.; Dolvik, N.I.; Ekman, S.; Ellermann, J.; Olstad, K.; Ytrehus, B.; Carlson, C.S. Articular Osteochondrosis: A Comparison of Naturally Occurring Human and Animal Disease. Osteoarthr. Cartil. 2013, 21, 1638–1647. [Google Scholar] [CrossRef]
- Bourebaba, L.; Röcken, M.; Marycz, K. Osteochondritis dissecans (OCD) in Horses—Molecular Background of its Pathogenesis and Perspectives for Progenitor Stem Cell Therapy. Stem Cell Rev. Rep. 2019, 15, 374–390. [Google Scholar] [CrossRef]
- Carballo, C.B.; Nakagawa, Y.; Sekiya, I.; Rodeo, S.A. Basic Science of Articular Cartilage. Clin. Sports Med. 2017, 36, 413–425. [Google Scholar] [CrossRef]
- Ruiz-Romero, C.; Fernández-Puente, P.; Calamia, V.; Blanco, F.J. Lessons from the proteomic study of osteoarthritis. Expert Rev. Proteom. 2015, 12, 433–443. [Google Scholar] [CrossRef]
- Lourido, L.; Blanco, F.J.; Ruiz-Romero, C. Defining the proteomic landscape of rheumatoid arthritis: Progress and prospective clinical applications. Expert Rev. Proteom. 2017, 14, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Bay-Jensen, A.-C.; van Spil, W.E.; Larkin, J.; Levesque, M.C. Osteoarthritis Year in Review 2016: Biomarkers (biochemical markers). Osteoarthr. Cartil. 2017, 25, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Trachana, V.; Mourmoura, E.; Papathanasiou, I.; Tsezou, A. Understanding the role of chondrocytes in osteoarthritis: Utilizing proteomics. Expert Rev. Proteom. 2019, 16, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Desjardin, C.; Chat, S.; Gilles, M.; Legendre, R.; Riviere, J.; Mata, X.; Balliau, T.; Esquerré, D.; Cribiu, E.P.; Betch, J.-M.; et al. Involvement of mitochondrial dysfunction and ER-stress in the physiopathology of equine osteochondritis dissecans (OCD). Exp. Mol. Pathol. 2014, 96, 328–338. [Google Scholar] [CrossRef]
- Desjardin, C.; Riviere, J.; Vaiman, A.; Morgenthaler, C.; Diribarne, M.; Zivy, M.; Robert, C.; Le Moyec, L.; Wimel, L.; Lepage, O.; et al. Omics technologies provide new insights into the molecular physiopathology of equine osteochondrosis. BMC Genom. 2014, 15, 947. [Google Scholar] [CrossRef]
- Chiaradia, E.; Pepe, M.; Tartaglia, M.; Scoppetta, F.; D’Ambrosio, C.; Renzone, G.; Avellini, L.; Moriconi, F.; Gaiti, A.; Bertuglia, A.; et al. Gambling on putative biomarkers of osteoarthritis and osteochondrosis by equine synovial fluid proteomics. J. Proteom. 2012, 75, 4478–4493. [Google Scholar] [CrossRef]
- Anderson, J.R.; Smagul, A.; Simpson, D.; Clegg, P.D.; Rubio-Martinez, L.M.; Peffers, M.J. The synovial fluid proteome differentiates between septic and nonseptic articular pathologies. J. Proteom. 2019, 202, 103370. [Google Scholar] [CrossRef]
- Roughley, P.J.; Mort, J.S. The role of aggrecan in normal and osteoarthritic cartilage. J. Exp. Orthop. 2014, 1, 8. [Google Scholar] [CrossRef]
- Zheng, J.; Wu, C.; Ma, W.; Zhang, Y.; Hou, T.; Xu, H.; Wu, S.; Yao, X.; Guo, X. Abnormal expression of chondroitin sulphate N-Acetylgalactosaminyltransferase 1 and Hapln-1 in cartilage with Kashin-Beck disease and primary osteoarthritis. Int. Orthop. 2013, 37, 2051–2059. [Google Scholar] [CrossRef]
- Lorenzo, P.; Bayliss, M.T.; Heinegård, D. Altered patterns and synthesis of extracellular matrix macromolecules in early osteoarthritis. Matrix Biol. 2004, 23, 381–391. [Google Scholar] [CrossRef]
- Tew, S.R.; Clegg, P.D.; Brew, C.J.; Redmond, C.M.; Hardingham, T.E. SOX9 transduction of a human chondrocytic cell line identifies novel genes regulated in primary human chondrocytes and in osteoarthritis. Arthritis Res. Ther. 2007, 9, R107. [Google Scholar] [CrossRef]
- Garvican, E.R.; Vaughan-Thomas, A.; Redmond, C.; Clegg, P.D. Chondrocytes harvested from osteochondritis dissecans cartilage are able to undergo limited in vitro chondrogenesis despite having perturbations of cell phenotype in vivo. J. Orthop. Res. 2008, 26, 1133–1140. [Google Scholar] [CrossRef]
- Henson, F.M.; Davies, M.E.; Jeffcott, L.B. Equine dyschondroplasia (osteochondrosis)—Histological findings and type VI collagen localization. Vet. J. 1997, 154, 53–62. [Google Scholar] [CrossRef]
- Riddick, T.L.; Duesterdieck-Zellmer, K.; Semevolos, S.A. Gene and protein expression of cartilage canal and osteochondral junction chondrocytes and full-thickness cartilage in early equine osteochondrosis. Vet. J. 2012, 194, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, R.E.; Sanchez-Adams, J.; Guilak, F. The structure and function of the pericellular matrix of articular cartilage. Matrix Biol. 2014, 39, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Zelenski, N.A.; Leddy, H.A.; Sanchez-Adams, J.; Zhang, J.; Bonaldo, P.; Liedtke, W.; Guilak, F. Type VI collagen regulates pericellular matrix properties, chondrocyte swelling, and mechanotransduction in mouse articular cartilage. Arthritis Rheumatol. 2015, 67, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Guilak, F.; Nims, R.J.; Dicks, A.; Wu, C.-L.; Meulenbelt, I. Osteoarthritis as a disease of the cartilage pericellular matrix. Matrix Biol. 2018, 71–72, 40–50. [Google Scholar] [CrossRef]
- Blain, E.J. Involvement of the cytoskeletal elements in articular cartilage homeostasis and pathology. Int. J. Exp. Pathol. 2009, 90, 1–15. [Google Scholar] [CrossRef]
- Chahine, N.O.; Blanchette, C.; Thomas, C.B.; Lu, J.; Haudenschild, D. Effect of Age and Cytoskeletal Elements on the Indentation-Dependent Mechanical Properties of Chondrocytes. PLoS ONE 2013, 8, 61651. [Google Scholar] [CrossRef]
- Woods, A.; Wang, G.; Beier, F. Regulation of chondrocyte differentiation by the act in cytoskeleton and adhesive interactions. J. Cell. Physiol. 2007, 213, 1–8. [Google Scholar] [CrossRef]
- Bobick, B.E.; Tuan, R.S.; Chen, F.H. The intermediate filament vimentin regulates chondrogenesis of adult human bone marrow-derived multipotent progenitor cells. J. Cell. Biochem. 2010, 109, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yin, L.; Song, X.; Yang, H.; Ren, X.; Gong, X.; Wang, F.; Yang, L. Effects of vimentin disruption on the mechanoresponses of articular chondrocyte. Biochem. Biophys. Res. Commun. 2016, 469, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Ao, X.; Wang, L.; Shao, Y.; Chen, X.; Zhang, J.; Chu, J.; Jiang, T.; Zhang, Z.; Huang, M. Development and Characterization of a Novel Bipedal Standing Mouse Model of Intervertebral Disc and Facet Joint Degeneration. Clin. Orthop. Relat. Res. 2019, 477, 1492–1504. [Google Scholar] [CrossRef] [PubMed]
- Geminiani, M.; Gambassi, S.; Millucci, L.; Lupetti, P.; Collodel, G.; Mazzi, L.; Frediani, B.; Braconi, D.; Marzocchi, B.; Laschi, M.; et al. Cytoskeleton Aberrations in Alkaptonuric Chondrocytes. J. Cell. Physiol. 2017, 232, 1728–1738. [Google Scholar] [CrossRef]
- Duan, W.; Wei, L.; Cao, X.; Guo, H.; Wang, L.; Hao, Y.; Wei, X. Effect of the disruption of three cytoskeleton components on chondrocyte metabolism in rabbit knee cartilage. Chin. Med. J. 2014, 127, 3764–3770. [Google Scholar]
- Al-Amoudi, A.; Castano-Diez, D.; Devos, D.P.; Russell, R.B.; Johnson, G.T.; Frangakis, A.S. The three-dimensional molecular structure of the desmosomal plaque. Proc. Natl. Acad. Sci. USA 2011, 108, 6480–6485. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, S.; Huang, J.; Guo, W.; Chen, J.; Zhang, L.; Zhao, B.; Peng, J.; Wang, A.; Wang, Y.; et al. The ECM-cell interaction of cartilage extracellular matrix on chondrocytes. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Hayes, M.J.; Rescher, U.; Gerke, V.; Moss, S.E. Annexin-Actin interactions. Traffic 2004, 5, 571–576. [Google Scholar] [CrossRef]
- Bizzarro, V.; Petrella, A.; Parente, L. Annexin A1: Novel roles in skeletal muscle biology. J. Cell. Physiol. 2012, 227, 3007–3015. [Google Scholar] [CrossRef]
- Lim, L.H.K.; Pervaiz, S. Annexin 1: The new face of an old molecule. FASEB J. 2007, 21, 968–975. [Google Scholar] [CrossRef]
- Kirsch, T. Annexins—Their role in cartilage mineralization. Front. Biosci. 2005, 10, 576–581. [Google Scholar] [CrossRef]
- Sluchanko, N.N.; Gusev, N.B. 14-3-3 Proteins and regulation of cytoskeleton. Biochemistry 2010, 75, 1528–1546. [Google Scholar] [CrossRef]
- Herzog, A.; Genin, O.; Hasdai, A.; Shinder, D.; Pines, M. Hsp90 and angiogenesis in bone disorders-lessons from the avian growth plate. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R140–R147. [Google Scholar] [CrossRef]
- Zhu, G.; Lee, A.S. Role of the unfolded protein response, GRP78 and GRP94 in organ homeostasis. J. Cell. Physiol. 2015, 230, 1413–1420. [Google Scholar] [CrossRef]
- Hughes, A.; Oxford, A.E.; Tawara, K.; Jorcyk, C.L.; Oxford, J.T. Endoplasmic Reticulum Stress and Unfolded Protein Response in Cartilage Pathophysiology; Contributing Factors to Apoptosis and Osteoarthritis. Int. J. Mol. Sci. 2017, 18, 665. [Google Scholar] [CrossRef]
- Reddy, V.S.; Trinath, J.; Reddy, G.B. Implication of homocysteine in protein quality control processes. Biochimie 2019, 165, 19–31. [Google Scholar] [CrossRef]
- Cho, Y.R.; Lee, S.J.; Jeon, H.B.; Park, Z.Y.; Chun, J.-S.; Yoo, Y.J. Under-sulfation by PAPS synthetase inhibition modulates the expression of ECM molecules during chondrogenesis. Biochem. Biophys. Res. Commun. 2004, 323, 769–775. [Google Scholar] [CrossRef]
- Pecora, F.; Gualeni, B.; Forlino, A.; Superti-Furga, A.; Tenni, R.; Cetta, G.; Rossi, A. In vivo contribution of amino acid sulfur to cartilage proteoglycan sulfation. Biochem. J. 2006, 398, 509–514. [Google Scholar] [CrossRef]
- Ursini, F.; Pipicelli, G. Nutritional Supplementation for Osteoarthritis. Altern. Complement. Ther. 2009, 15, 173–177. [Google Scholar] [CrossRef]
- Vijayan, V.; Khandelwal, M.; Manglani, K.; Gupta, S.; Surolia, A. Methionine down-regulates TLR4/MyD88/NF-κB signalling in osteoclast precursors to reduce bone loss during osteoporosis. Br. J. Pharmacol. 2014, 171, 107–121. [Google Scholar] [CrossRef]
- Pajares, M.A.; Pérez-Sala, D. Mammalian Sulfur Amino Acid Metabolism: A Nexus Between Redox Regulation, Nutrition, Epigenetics, and Detoxification. Antioxid. Redox Signal. 2018, 29, 408–452. [Google Scholar] [CrossRef] [PubMed]
- Feigerlova, E.; Demarquet, L.; Guéant, J.-L. One carbon metabolism and bone homeostasis and remodeling: A review of experimental research and population studies. Biochimie 2016, 126, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Hollander, J.M.; Zeng, L. The Emerging Role of Glucose Metabolism in Cartilage Development. Curr. Osteoporos. Rep. 2019, 17, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Semevolos, S.A.; Duesterdieck-Zellmer, K.F.; Larson, M.; Kinsley, M.A. Expression of pro-apoptotic markers is increased along the osteochondral junction in naturally occurring osteochondrosis. Bone Rep. 2018, 9, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Nugent, A.E.; McBurney, D.L.; Horton, W.E. The presence of extracellular matrix alters the chondrocyte response to endoplasmic reticulum stress. J. Cell. Biochem. 2011, 112, 1118–1129. [Google Scholar] [CrossRef]
- Peters, H.C.; Otto, T.J.; Enders, J.T.; Jin, W.; Moed, B.R.; Zhang, Z. The protective role of the pericellular matrix in chondrocyte apoptosis. Tissue Eng. Part A 2011, 17, 2017–2024. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; De Pinto, V.; Zweckstetter, M.; Raviv, Z.; Keinan, N.; Arbel, N. VDAC, A multi-functional mitochondrial protein regulating cell life and death. Mol. Asp. Med. 2010, 31, 227–285. [Google Scholar] [CrossRef]
- Maurya, S.R.; Mahalakshmi, R. Mitochondrial VDAC2 and cell homeostasis: Highlighting hidden structural features and unique functionalities. Biol. Rev. Camb. Philos. Soc. 2017, 92, 1843–1858. [Google Scholar] [CrossRef]
- Mancini, F.; Nannarone, S.; Buratta, S.; Ferrara, G.; Stabile, A.M.; Vuerich, M.; Santinelli, I.; Pistilli, A.; Chiaradia, E. Effects of xylazine and dexmedetomidine on equine articular chondrocytes in vitro. Vet. Anaesth. Analg. 2017, 44, 295–308. [Google Scholar] [CrossRef]
- Shevchenko, A.; Tomas, H.; Havlis, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Perkins, D.N.; Pappin, D.J.C.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. ELECTROPHORESIS Int. J. 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Chiaradia, E.; Valiani, A.; Tartaglia, M.; Scoppetta, F.; Renzone, G.; Arena, S.; Avellini, L.; Benda, S.; Gaiti, A.; Scaloni, A. Ovine subclinical mastitis: Proteomic analysis of whey and milk fat globules unveils putative diagnostic biomarkers in milk. J. Proteom. 2013, 83, 144–159. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Bindea, G.; Galon, J.; Mlecnik, B. CluePedia Cytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° Spot | Protein Name | UniProt Accession | Gene Name | pI/Mw Theor(kDa) | pI/Mw Exp(kDa) | Fold Change | p Value |
|---|---|---|---|---|---|---|---|
| 1 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110 | 4.55/105 | 0.44 | 0.035 |
| 2 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110 | 4.68/105 | 0.40 | 0.035 |
| 3 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110 | 4.96/105 | 0.42 | 0.0001 |
| 4 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110 | 5.05/105 | 0.36 | 0.0001 |
| 5 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110 | 5.11/105 | 0.30 | 0.0001 |
| 6 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110 | 5.18/105 | 0.49 | 0.005 |
| 7 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110 | 5.25/106 | 0.41 | 0.045 |
| 8 | Endoplasmin | F6ZUJ2_HORSE | HSP90B1 | 4.76/92.6 | 4.71/83.2 | 0.44 | 0.045 |
| 9 | Collagen alpha-3(VI) chain * | F6QAT0_HORSE | COL6A3 | 6.07/342.2 | 5.27/95 | +++ | |
| 10 | Collagen alpha-3(VI) chain * | F6QAT0_HORSE | COL6A3 | 6.07/342.2 | 5.33/95 | +++ | |
| 11 | Collagen alpha-3(VI) chain * | F6QAT0_HORSE | COL6A3 | 6.07/342.2 | 5.39/95.2 | +++ | |
| 12 | Collagen alpha-3(VI) chain * | F6QAT0_HORSE | COL6A3 | 6.07/342.2 | 5.45/95.2 | +++ | |
| 13 | Collagen alpha-3(VI) chain * | F6QAT0_HORSE | COL6A3 | 6.07/342.2 | 5.68/82.4 | 3.01 | 0.011 |
| 14 | Collagen alpha-3(VI) chain * | F6QAT0_HORSE | COL6A3 | 6.07/342.2 | 5.78/81.8 | 4.02 | 0.017 |
| 15 | Collagen alpha-3(VI) chain * | F6QAT0_HORSE | COL6A3 | 6.07/342.2 | 5.87/81.8 | 3.64 | 0.002 |
| 16 | Collagen alpha-3(VI) chain * | F6QAT0_HORSE | COL6A3 | 6.07/342.2 | 6.00/81.7 | 2.73 | 0.031 |
| 17 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 6.41/81.4 | 0.48 | 0.009 |
| 18 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 6.65/80.8 | 0.46 | 0.0001 |
| 19 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 6.79/80.6 | 0.47 | 0.002 |
| 20 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 7.11/80.4 | 0.28 | 0.0001 |
| 21 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 7.33/80.1 | 0.36 | 0.001 |
| 22 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 7.39/77.1 | 0.15 | 0.0001 |
| 23 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 7.66/76.6 | 0.25 | 0.0001 |
| 24 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 7.94/75.6 | 0.20 | 0.0001 |
| 25 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 8.20/75.3 | 0.42 | 0.0001 |
| 26 | Vimentin | F7B5C4_HORSE | VIM | 4.67/42.2 | 5.18/74.3 | 0.28 | 0.005 |
| 27 | Vimentin | F7B5C4_HORSE | VIM | 4.67/42.2 | 5.29/73.9 | 0.40 | 0.003 |
| 28 | Lactadherin | F7B0S3_HORSE | MFGE8 | 6.22/46.4 | 6.06/65.2 | 2.76 | 0.014 |
| 29 | Lactadherin | F7B0S3_HORSE | MFGE8 | 6.22/46.4 | 6.70/64.2 | 2.10 | 0.0002 |
| 30 | ATP synthase subunit beta | F6U187_HORSE | ATP5F1B | 5.15/56.2 | 5.24/55.5 | 3.15 | 0.017 |
| 31 | ATP synthase subunit beta | F6U187_HORSE | ATP5F1B | 5.15/56.2 | 5.16/54.5 | 5.90 | 0.038 |
| 32 | Hyaluronan and proteoglycan link protein 1 | HPLN1_HORSE | HAPLN1 | 8.19/40. 7 | 5.13/50.1 | 6.59 | 0.001 |
| 33 | Endoplasmic reticulum resident protein 44 | F6WY40_HORSE | ERP44 | 5.00/47.44 | 5.25/50.9 | 2.27 | 0.005 |
| 34 | Trifunctional enzyme mitochondrial. | F6TCZ6_HORSE | HADHB | 9.49/51.4 | 5.36/50.9 | 7.27 | 0.0001 |
| 35 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110 | 5.45/50.8 | 3.60 | 0.006 |
| 36 | Junction plakoglobin * | F6V1T9_HORSE | JUP | 5.73/82.5 | 5.54/50.9 | 6.64 | 0.004 |
| 37 | Enolase 1 | F6V7C1_HORSE | ENO1 | 6.37/47.5 | 5.63/51.0 | 2.76 | 0.007 |
| 38 | Ornithine aminotransferase | F7C9Y5_HORSE | OAT | 6.84/48.7 | 7.45/50.4 | 0.29 | 0.0001 |
| 39 | Adenosylhomocysteinase | F6VIG9_HORSE | AHCY | 5.86/48.1 | 6.90/48.7 | 0.21 | 0.009 |
| 40 | Adenosylhomocysteinase | F6VIG9_HORSE | AHCY | 5.86/48.1 | 7.15/48.4 | 0.15 | 0.001 |
| 41 | Cartilage intermediate layer protein | F7C2J3_HORSE | CILP | 8.85/135 | 7.37/48.3 | 0.37 | 0.0001 |
| 42 | Actin, cytoplasmic 1 | F7AAK7_HORSE | ACTG1 | 5.31/42.1 | 5.51/43.2 | 11.9 | 0.0001 |
| 43 | Actin, cytoplasmic 1 | F7AAK7_HORSE | ACTG1 | 5.31/42.1 | 5.60/43.3 | 4.22 | 0.008 |
| 44 | Hyaluronan and proteoglycan link protein 1 | HPLN1_HORSE | HAPLN1 | 8.19/40.6 | 5.66/42.7 | 8.64 | 0.0005 |
| 45 | Hyaluronan and proteoglycan link protein 1 | HPLN1_HORSE | HAPLN1 | 8.19/40.6 | 5.26/40.1 | 0.09 | 0.004 |
| 46 | Hyaluronan and proteoglycan link protein 1 | HPLN1_HORSE | HAPLN1 | 8.19/40.6 | 5.45/40.0 | 0.23 | 0.0006 |
| 47 | Collagen alpha-2(VI) chain | F7CGV8_HORSE | COL6A2 | 5.93/105.6 | 4.94/37.9 | 0.42 | 0.012 |
| 48 | Annexin A2 | F6ZI51_HORSE | ANXA2 | 6.92/38.8 | 7.21/37.9 | 2.00 | 0.0023 |
| 49 | Annexin A1 | ANXA1_HORSE | ANXA1 | 6.57/39 | 7.42/38.0 | 2.02 | 0.0002 |
| 50 | Glyceraldehyde-3-phosphate dehydrogenase | F6YV40_HORSE | GAPDH | 8.22/36.0 | 7.75/38.2 | 3.11 | 0.0001 |
| 51 | L-lactate dehydrogenase | C6L1J5_HORSE | LDHB | 5.85/36.8 | 6.58/33.9 | 2.93 | 0.004 |
| 52 | Collagen alpha-1(VI) chain | F6UW03_HORSE | COL6A1 | 5.24/110. | 7.01/33.7 | 2.8 | 0.0004 |
| 53 | Succinyl-CoA ligase | F6QTC9_HORSE | SUCLG1 | 9.27/36.6 | 7.28/33.2 | 2.22 | 0.0003 |
| 54 | Voltage dependent anion channel 2 | F6TLU0_HORSE | VDAC2 | 7.60/32.3 | 7.62/33.7 | 4.01 | 0.0001 |
| 55 | 14-3-3 protein theta | F6SP02_HORSE | YWHAQ | 4.68/28.0 | 4.72/30.4 | 0.39 | 0.015 |
| 56 | Triosephosphate isomerase | F6TZS9_HORSE | TPI1 | 6.96/26.9 | 8.10/28.2 | 2.98 | 0.0001 |
| 57 | Enoyl-CoA hydratase | F6QV70_HORSE | ECHS1 | 7.55/28.2 | 8.64/27.1 | 3.49 | 0.0001 |
| 58 | Superoxide dismutase | F6U991_HORSE | HADHB | 8.44/24.4 | 7.24/26.4 | 4.39 | 0.0001 |
| 59 | Superoxide dismutase | F6U991_HORSE | HADHB | 8.44/24.4 | 7.75/25.5 | 0.27 | 0.0001 |
| 60 | Superoxide dismutase | F6U991_HORSE | HADHB | 8.44/24.4 | 8.11/25.4 | 0.23 | 0.0001 |
| 61 | Collagen alpha-1(VI) chain * | F6UW03_HORSE | COL6A1 | 5.24/110 | 7.82/23.1 | +++ | |
| 62 | Collagen alpha-1(VI) chain * | F6UW03_HORSE | COL6A1 | 5.24/110 | 7.81/22.4 | +++ | |
| 63 | Collagen alpha-1(VI) chain * | F6UW03_HORSE | COL6A1 | 5.24/110 | 8.11/22.7 | +++ | |
| 64 | Collagen alpha-1(VI) chain * | F6UW03_HORSE | COL6A1 | 5.24/110 | 8.11/22.2 | +++ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiaradia, E.; Pepe, M.; Orvietani, P.L.; Renzone, G.; Magini, A.; Sforna, M.; Emiliani, C.; Di Meo, A.; Scaloni, A. Proteome Alterations in Equine Osteochondrotic Chondrocytes. Int. J. Mol. Sci. 2019, 20, 6179. https://doi.org/10.3390/ijms20246179
Chiaradia E, Pepe M, Orvietani PL, Renzone G, Magini A, Sforna M, Emiliani C, Di Meo A, Scaloni A. Proteome Alterations in Equine Osteochondrotic Chondrocytes. International Journal of Molecular Sciences. 2019; 20(24):6179. https://doi.org/10.3390/ijms20246179
Chicago/Turabian StyleChiaradia, Elisabetta, Marco Pepe, Pier Luigi Orvietani, Giovanni Renzone, Alessandro Magini, Monica Sforna, Carla Emiliani, Antonio Di Meo, and Andrea Scaloni. 2019. "Proteome Alterations in Equine Osteochondrotic Chondrocytes" International Journal of Molecular Sciences 20, no. 24: 6179. https://doi.org/10.3390/ijms20246179
APA StyleChiaradia, E., Pepe, M., Orvietani, P. L., Renzone, G., Magini, A., Sforna, M., Emiliani, C., Di Meo, A., & Scaloni, A. (2019). Proteome Alterations in Equine Osteochondrotic Chondrocytes. International Journal of Molecular Sciences, 20(24), 6179. https://doi.org/10.3390/ijms20246179