Human Synovia Contains Trefoil Factor Family (TFF) Peptides 1–3 Although Synovial Membrane Only Produces TFF3: Implications in Osteoarthritis and Rheumatoid Arthritis

 ,
, 
Abstract
1. Introduction
2. Results
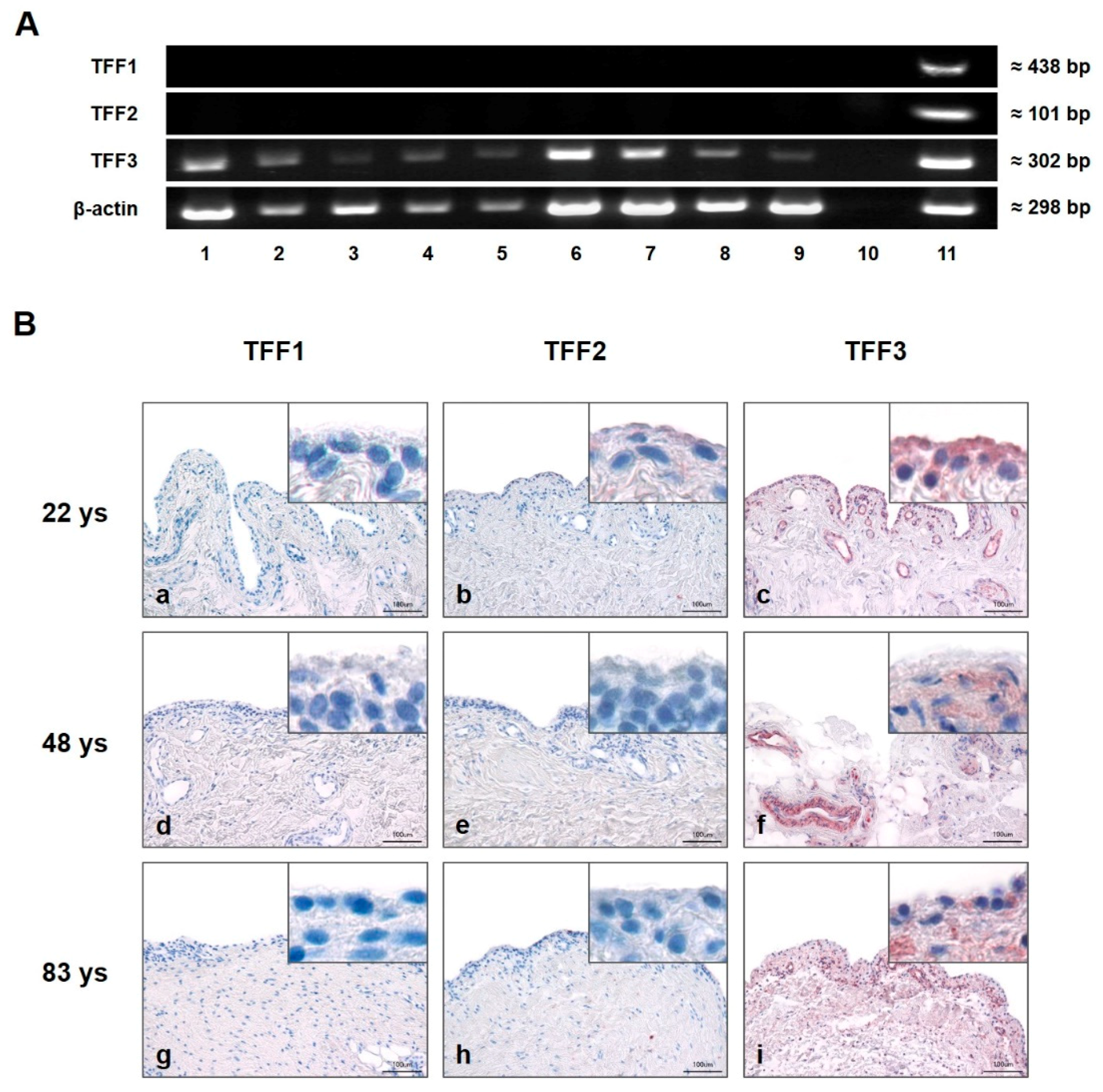
2.1. TFF3, but not TFF1 and TFF2, is Expressed in Human SM of Healthy Donors and SM of Patients Suffering From OA or RA on the Protein and RNA Level
2.1.1. Presence and Localization of TFFs in the Synovial Membrane
2.1.2. Quantification of TFF3 in the Synovial Membrane
2.2. All Three TFF Peptides (TFF1, -2 and -3) Are Detectable in Healthy Synovial Fluid As Well as in SF of Patients Suffering From OA or RA
Western Blot
2.3. TFF2 Protein Concentration Is Significantly Increased in Synovial Fluid of Patients Suffering From RA, TFF3 Protein Concentration Is Reduced in Cases of OA and RA, and TFF1 Shows No Change
ELISA
3. Discussion
4. Materials and Methods
4.1. Human Tissues/Fluids
4.1.1. Synovial Membrane
4.1.2. Synovial Fluid
4.1.3. Human Control Tissue
4.2. RNA Isolation and Complementary DNA (cDNA) Synthesis
4.3. Reverse Transcriptase-Polymerase Chain Reaction (Semi-Quantitative PCR)
4.4. Quantitative Real-Time (qPCR)
4.5. Western Blot Analysis
4.6. Immunohistochemistry
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Statistical Analysis of ELISA and Real-Time PCR Data
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, M.D. The Normal Synovium. Rheumatol. J. 2011, 5 (Suppl 1), 100. [Google Scholar] [CrossRef]
- Culemann, S.; Grüneboom, A.; Nicolás-Ávila, J.Á.; Weidner, D.; Lämmle, K.F.; Rothe, T.; Quintana, J.A.; Kirchner, P.; Krljanac, B.; Eberhardt, M.; et al. Spatiotemporal molecular profiling of synovial macrophates reveals a locally renewing barrier of membrane-forming macrophages shielding the joint. Nature 2019, 66, 289–306. [Google Scholar]
- Iwanaga, T.; Shikichi, M.; Kitamura, H.; Yanase, H.; Nozawa-Inoue, K. Morphology and functional roles of synoviocytes in the joint. Arch. Histol. Cytol. 2000, 63, 17–31. [Google Scholar] [CrossRef]
- Balazs, E.A.; Denlinger, J.L. Sodium hyaluronate and joint function. J. Equine Vet. Sci. 1985, 5, 217–219. [Google Scholar] [CrossRef]
- Jay, G.D.; Britt, D.E.; Cha, C.J. Lubricin is a product of megakaryocyte stimulating factor gene expression by human synovial fibroblasts. J. Rheumatol. 2000, 27, 594–600. [Google Scholar]
- Dobbie, J.W.; Hind, C.; Meijers, P.; Bodart, C.; Tasiaux, N.; Perret, J.; Anderson, J.D. Lamellar body secretion: Ultrastructural analysis of an unexplored function of synoviocytes. Br. J. Rheumatol. 1995, 34, 13–23. [Google Scholar] [CrossRef]
- Hui, A.Y.; McCarty, W.J.; Masuda, K.; Firestein, G.S.; Sah, R.L. A systems biology approach to synovial joint lubrication in health, injury, and disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2012, 4, 15–37. [Google Scholar] [CrossRef]
- Taupin, D.R.; Kinoshita, K.; Podolsky, D.K. Intestinal trefoil factor confers colonic epithelial resistance to apoptosis. Proc. Natl. Acad. Sci. USA 2000, 97, 799–804. [Google Scholar] [CrossRef]
- Hoffmann, W.; Jagla, W.; Wiede, A. Molecular medicine of TFF-peptides: From gut to brain. Histol. Histopathol. 2001, 16, 319–334. [Google Scholar]
- Wong, W.M.; Poulsom, R.; Wright, N.A. Trefoil peptides. Gut 1999, 44, 890–895. [Google Scholar] [CrossRef]
- Madsen, J.; Nielsen, O.; Tornøe, I.; Thim, L.; Holmskov, U. Tissue localization of human trefoil factors 1, 2, and 3. J. Histochem. Cytochem. 2007, 55, 505–513. [Google Scholar] [CrossRef]
- Kjellev, S. The trefoil factor family–small peptides with multiple functionalities. Cell. Mol. Life Sci. 2009, 66, 1350–1369. [Google Scholar] [CrossRef]
- Thim, L.; Madsen, F.; Poulsen, S.S. Effect of trefoil factors on the viscoelastic properties of mucus gels. Eur. J. Clin. Investig. 2002, 32, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, W. Trefoil factor family (TFF) peptides: Regulators of mucosal regeneration and repair, and more. Peptides 2004, 25, 727–730. [Google Scholar] [CrossRef]
- Krüger, K.; Schmid, S.; Paulsen, F.; Ignatius, A.; Klinger, P.; Hotfiel, T.; Swoboda, B.; Gelse, K. Trefoil Factor Peptide 3 (TFF3) is involved in cell migration for skeletal repair. Int. J. Mol. Sci. 2019, 20, 4277. [Google Scholar] [CrossRef]
- Cook, G.A.; Familari, M.; Thim, L.; Giraud, A.S. The trefoil peptides TFF2 and TFF3 are expressed in rat lymphoid tissues and participate in the immune response. FEBS Lett. 1999, 456, 155–159. [Google Scholar] [CrossRef]
- Taupin, D.; Podolsky, D.K. Trefoil factors: Initiators of mucosal healing. Nat. Rev. Mol. Cell Biol. 2003, 4, 721–732. [Google Scholar] [CrossRef]
- Hoffmann, W. Trefoil factor family (TFF) peptides and chemokine receptors: A promising relationship. J. Med. Chem. 2009, 52, 6505–6510. [Google Scholar] [CrossRef]
- Rösler, S.; Haase, T.; Claassen, H.; Schulze, U.; Schicht, M.; Riemann, D.; Brandt, J.; Wohlrab, D.; Müller-Hilke, B.; Goldring, M.B.; et al. Trefoil factor 3 is induced during degenerative and inflammatory joint disease, activates matrix metalloproteinases, and enhances apoptosis of articular cartilage chondrocytes. Arthritis Rheumatol. 2010, 62, 815–825. [Google Scholar] [CrossRef]
- Woolf, A.D.; Pfleger, B. Burden of major musculoskeletal conditions. Bull. World Health Organ. 2003, 81, 646–656. [Google Scholar]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.D.; Triantafillou, S.; Parker, A.; Youssef, P.P.; Coleman, M. Synovial membrane inflammation and cytokine production in patients with early osteoarthritis. J. Rheumatol. 1997, 24, 365–371. [Google Scholar] [PubMed]
- Pelletier, J.P.; Martel-Pelletier, J.; Abramson, S.B. Osteoarthritis, an inflammatory disease: Potential implication for the selection of new therapeutic targets. Arthritis Rheumatol. 2001, 44, 1237–1247. [Google Scholar] [CrossRef]
- Sellam, J.; Berenbaum, F. The role of synovitis in pathophysiology and clinical symptoms of osteoarthritis. Nat. Rev. Rheumatol. 2010, 6, 625–635. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. The Pathogenesis of Rheumatoid Arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef]
- Pap, T.; Ladner, U.M.; Gay, R.E.; Gay, S. Fibroblast biology. Role of synovial fibroblasts in the pathogenesis of rheumatoid arthritis. Arthritis Res. 2000, 2, 361–367. [Google Scholar] [CrossRef][Green Version]
- Bijelic, N.; Belovari, T.; Baus Loncar, M. Trefoil factor family protein 3 (TFF3) is present in cartilage during endochondral ossification in the developing mouse fetus. Acta Histochem. 2013, 115, 204–208. [Google Scholar] [CrossRef]
- Brandt, K.D.; Radin, E.L.; Dieppe, P.A.; van de Putte, L. Yet more evidence that osteoarthritis is not a cartilage disease. Ann. Rheum. Dis. 2006, 65, 1261–1264. [Google Scholar] [CrossRef]
- Ropes, M.W.; Rossmeisl, E.C.; Bauer, W. The Origin And Nature Of Normal Human Synovial Fluid. J. Clin. Investig. 1940, 19, 795–799. [Google Scholar] [CrossRef]
- Levick, J.R. Microvascular architecture and exchange in synovial joints. Microcirculation 1995, 2, 217–233. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, X.; Lu, H.; Wu, L.; Wang, D.; Zhang, Q.; Ding, H. Serum trefoil factor 3 is a promising non-invasive biomarker for gastric cancer screening: A monocentric cohort study in China. BMC Gastroenterol. 2014, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- Chaiyarit, P.; Utrawichian, A.; Leelayuwat, C.; Vatanasapt, P.; Chanchareonsook, N.; Samson, M.H.; Giraud, A.S. Investigation of trefoil factor expression in saliva and oral mucosal tissues of patients with oral squamous cell carcinoma. Clin. Oral Investig. 2012, 16, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.J.; Bjerner, J.; Paus, E.; Børmer, O.P.; Nustad, K. Use of an in vivo biotinylated single-chain antibody as capture reagent in an immunometric assay to decrease the incidence of interference from heterophilic antibodies. Clin. Chem. 2005, 51, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Hampel, U.; Sesselmann, S.; Iserovich, P.; Sel, S.; Paulsen, F.; Sack, R. Chemokine and cytokine levels in osteoarthritis and rheumatoid arthritis synovial fluid. J. Immunol. Methods 2013, 396, 134–139. [Google Scholar] [CrossRef]
- Samson, M.H. Quantitative measurements of trefoil factor family peptides: Possibilities and pitfalls. Scand. J. Clin. Lab. Investig. 2013, 73, 193–202. [Google Scholar] [CrossRef]
- Chadwick, M.P.; Westley, B.R.; May, F.E. Homodimerization and hetero-oligomerization of the single-domain trefoil protein pNR-2/pS2 through cysteine 58. Biochem. J. 1997, 327, 117–123. [Google Scholar] [CrossRef]
- Chinery, R.; Bates, P.A.; De, A.; Freemont, P.S. Characterisation of the single copy trefoil peptides intestinal trefoil factor and pS2 and their ability to form covalent dimers. FEBS Lett. 1995, 357, 50–54. [Google Scholar] [CrossRef]
- Otto, W.R.; Thim, L. Trefoil factor family-interacting proteins. Cell. Mol. Life Sci. 2005, 62, 2939–2946. [Google Scholar] [CrossRef]
- Hoffmann, W.; Jagla, W. Cell type specific expression of secretory TFF peptides: Colocalization with mucins and synthesis in the brain. In International Review of Cytology; Kwang, W.J., Ed.; Academic Press: Cambridge, Ma, USA, 2002; pp. 147–181. [Google Scholar]
- Ruchaud-Sparagano, M.H.; Westley, B.R.; May, F.E. The trefoil protein TFF1 is bound to MUC5AC in human gastric mucosa. Cell. Mol. Life Sci. 2004, 61, 1946–1954. [Google Scholar] [CrossRef]
- Volin, M.V.; Shahrara, S.; Haines, G.K., 3rd; Woods, J.M.; Koch, A.E. Expression of mucin 3 and mucin 5AC in arthritic synovial tissue. Arthritis Rheum. 2008, 58, 46–52. [Google Scholar] [CrossRef][Green Version]
- Hamaguchi, M.; Kawahito, Y.; Ishino, H.; Takeuchi, N.; Tokunaga, D.; Hojo, T.; Yamamoto, A.; Kadoya, M.; Seno, T.; Kohno, M.; et al. Mucin from rheumatoid arthritis synovial fluid enhances interleukin-6 production by human peripheral blood mononuclear cells. Hum. Immunol. 2011, 72, 241–248. [Google Scholar] [CrossRef]
- Benito, M.J.; Veale, D.J.; FitzGerald, O.; van den Berg, W.B.; Bresnihan, B. Synovial tissue inflammation in early and late osteoarthritis. Ann. Rheum. Dis. 2005, 64, 1263–1267. [Google Scholar] [CrossRef]
- Chaiyarit, P.; Chayasadom, A.; Wara-Aswapati, N.; Hormdee, D.; Sittisomwong, S.; Nakaresisoon, S.; Samson, M.H.; Pitiphat, W.; Giraud, A.S. Trefoil factors in saliva and gingival tissues of patients with chronic periodontitis. J. Periodontol. 2012, 83, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Siber-Hoogeboom, R.; Schicht, M.; Hoogeboom, S.; Paulsen, F.; Traxdorf, M. Obstructive sleep apnea and rhonchopathy are associated with downregulation of trefoil factor family peptide 3 (TFF3)-Implications of changes in oral mucus composition. PLoS ONE 2017, 12, e0185200. [Google Scholar] [CrossRef]
- De Lange-Brokaar, B.J.E.; Ioan-Facsinay, A.; van Osch, G.J.; Zuurmond, A.M.; Schoones, J.; Toes, R.E.; Huizinga, T.W.; Kloppenburg, M. Synovial inflammation, immune cells and their cytokines in osteoarthritis: A review. Osteoarthr. Cartil. 2012, 20, 1484–1499. [Google Scholar] [CrossRef]
- Goldring, M.B. Anticytokine therapy for osteoarthritis. Expert Opin. Biol. Ther. 2001, 1, 817–829. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2014, 2014, 19. [Google Scholar] [CrossRef]
- Barrera Roa, G.J.; Sanchez Tortolero, G.; Gonzalez, J.E. Trefoil factor 3 (TFF3) expression is regulated by insulin and glucose. J. Health Sci. 2013, 2013, 3. [Google Scholar] [CrossRef]
- Vestergaard, E.M.; Brynskov, J.; Ejskjaer, K.; Clausen, J.T.; Thim, L.; Nexø, E.; Poulsen, S.S. Immunoassays of human trefoil factors 1 and 2: Measured on serum from patients with inflammatory bowel disease. Scand. J. Clin. Lab. Investig. 2004, 64, 146–156. [Google Scholar] [CrossRef]
- Viby, N.E.; Pedersen, L.; Lund, T.K.; Kissow, H.; Backer, V.; Nexø, E.; Thim, L.; Poulsen, S.S. Trefoil factor peptides in serum and sputum from subjects with asthma and COPD. Clin. Respir. J. 2014, 9, 322–329. [Google Scholar] [CrossRef]
- Du, T.-Y.; Luo, H.M.; Qin, H.C.; Wang, F.; Wang, Q.; Xiang, Y.; Zhang, Y. Circulating Serum Trefoil Factor 3 (TFF3) Is Dramatically Increased in Chronic Kidney Disease. PLoS ONE 2013, 8, e80271. [Google Scholar] [CrossRef]
- Lebherz-Eichinger, D.; Tudor, B.; Ankersmit, H.J.; Reiter, T.; Haas, M.; Einwallner, E.; Roth-Walter, F.; Krenn, C.G.; Roth, G.A. Increased trefoil factor 2 levels in patients with chronic kidney disease. PLoS ONE 2017, 12, e0174551. [Google Scholar] [CrossRef] [PubMed]
- Koitabashi, A.; Shimada, T.; Fujii, Y.; Hashimoto, T.; Hosaka, K.; Tabei, K.; Namatame, T.; Yoneda, M.; Hiraishi, H.; Terano, A. Indometacin up-regulates TFF2 expression in gastric epithelial cells. Aliment. Pharmacol. Ther. 2004, 20 (Suppl. 1), 171–176. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheumatol. 2010, 62, 2569–2581. [Google Scholar] [CrossRef]
- Schicht, M.; Rausch, F.; Beron, M.; Jacobi, C.; Garreis, F.; Hartjen, N.; Beileke, S.; Kruse, F.; Bräuer, L.; Paulsen, F. Palate Lung Nasal Clone (PLUNC), a Novel Protein of the Tear Film: Three-Dimensional Structure, Immune Activation, and Involvement in Dry Eye Disease (DED). Investig. Ophthalmol. Vis. Sci. 2015, 56, 7312–7323. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Relative Protein Concentration of TFF3 in Human Synovial Membrane (SM) | ||
|---|---|---|
| Samples | Mean (pg/mg) | Range (pg/mg) |
| healthy (n = 10) | 298.4 | 46.5–663.0 |
| OA (n = 10) | 473.7 | 141.1–1125.3 |
| RA (n = 10) | 299.3 | 155.9–701.8 |
| Relative Protein Concentration of TFF1 in Human Synovial Membrane (SM) | ||
|---|---|---|
| Samples | Mean (pg/mg) | Range (pg/mg) |
| healthy (n = 13) | 50.9 | 0.8–258.1 |
| OA (n = 20) | 31.2 | 9.5–117.3 |
| RA (n = 20) | 33.0 | 18.8–67.7 |
| Relative Protein Concentration of TFF2 in Human Synovial Membrane (SM) | ||
|---|---|---|
| Samples | Mean (pg/mg) | Range (pg/mg) |
| healthy (n = 13) | 13.7 | 2.4–74.3 |
| OA (n = 20) | 9.0 | 0.4–45.9 |
| RA (n = 20) | 203.5 | 0.3–835.4 |
| Relative Protein Concentration of TFF3 in Human Synovial Membrane (SM) | ||
|---|---|---|
| Samples | Mean (pg/mg) | Range (pg/mg) |
| healthy (n = 13) | 7807.0 | 2214.6–29279.2 |
| OA (n = 20) | 2833.9 | 1730.8–4828.8 |
| RA (n = 20) | 4083.9 | 2051.8–8413.0 |
| Primers Used for Semi Quantitative PCR | ||||
|---|---|---|---|---|
| Template | Sense 5´-3´ | Antisense 5′-3′ | Product | Annealing Temperature |
| TFF1 | TTTGGAGCAGAGAGGAGG | TTGAGTAGTCAAAGTCAGAGCAG | 438 bp | 60 °C |
| TFF2 | GTGTTTTGACAATGGATGCTG | CCTCCATGACGCACTGATC | 110 bp | 59 °C |
| TFF3 | GTGCAAGCCAAGGACAG | CGTTAAGACATCAGGCTCCAG | 302 bp | 56 °C |
| β-actin | GATCCTCACCGAGCGCGGCTACA | GCGGATGTCCACGTCACACTTCA | 298 bp | 60 °C |
| Primers used for real-time PCR | ||||
| TFF3 | TCCAGCTCTGCTGAGGAGTA | CAGTCCACCCTGTCCTTG | self-provided | 62 °C |
| Antibodies | Method | Specifity | Source |
|---|---|---|---|
| rabbit anti-TFF1 | IHC | monoclonal | AJ1765a, Abgent |
| rabbit anti-TFF1 | WB | polyclonal | orb100742, Biorbyt |
| goat anti-TFF2 | IHC, WB | polyclonal | SP (P-19): sc-23558, Santa Cruz |
| mouse anti-TFF2 | WB | monoclonal | H00007032-M01, Abnova |
| rabbit anti-TFF3 | IHC | polyclonal | ABIN 669786, Bioss Inc. |
| rabbit anti-h-TFF3 | WB | polyclonal | provided by Prof. W. Hoffmann, Magdeburg |
| rabbit anti-actin | WB | polyclonal | Actin (H-300): sc-10731, Santa Cruz |
| rabbit anti-alpha-1-antitrypsin | WB | polyclonal | A 0012, Dako |
| goat anti-rabbit | IHC | polyclonal | BA-1000, Vector Laboratories Inc. |
| rabbit anti-goat | IHC | polyclonal | E 0466, Dako |
| goat anti-rabbit | WB | polyclonal | P 0448, Dako |
| goat anti-mouse | WB | polyclonal | IgG-HRP: sc-2005, Santa Cruz |
| donkey anti-goat | WB | polyclonal | IgG-HRP: sc-2020, Santa Cruz |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popp, J.; Schicht, M.; Garreis, F.; Klinger, P.; Gelse, K.; Sesselmann, S.; Tsokos, M.; Etzold, S.; Stiller, D.; Claassen, H.; et al. Human Synovia Contains Trefoil Factor Family (TFF) Peptides 1–3 Although Synovial Membrane Only Produces TFF3: Implications in Osteoarthritis and Rheumatoid Arthritis. Int. J. Mol. Sci. 2019, 20, 6105. https://doi.org/10.3390/ijms20236105
Popp J, Schicht M, Garreis F, Klinger P, Gelse K, Sesselmann S, Tsokos M, Etzold S, Stiller D, Claassen H, et al. Human Synovia Contains Trefoil Factor Family (TFF) Peptides 1–3 Although Synovial Membrane Only Produces TFF3: Implications in Osteoarthritis and Rheumatoid Arthritis. International Journal of Molecular Sciences. 2019; 20(23):6105. https://doi.org/10.3390/ijms20236105
Chicago/Turabian StylePopp, Judith, Martin Schicht, Fabian Garreis, Patricia Klinger, Kolja Gelse, Stefan Sesselmann, Michael Tsokos, Saskia Etzold, Dankwart Stiller, Horst Claassen, and et al. 2019. "Human Synovia Contains Trefoil Factor Family (TFF) Peptides 1–3 Although Synovial Membrane Only Produces TFF3: Implications in Osteoarthritis and Rheumatoid Arthritis" International Journal of Molecular Sciences 20, no. 23: 6105. https://doi.org/10.3390/ijms20236105
APA StylePopp, J., Schicht, M., Garreis, F., Klinger, P., Gelse, K., Sesselmann, S., Tsokos, M., Etzold, S., Stiller, D., Claassen, H., & Paulsen, F. (2019). Human Synovia Contains Trefoil Factor Family (TFF) Peptides 1–3 Although Synovial Membrane Only Produces TFF3: Implications in Osteoarthritis and Rheumatoid Arthritis. International Journal of Molecular Sciences, 20(23), 6105. https://doi.org/10.3390/ijms20236105