Transcriptome Analysis of Salt-Sensitive and Tolerant Genotypes Reveals Salt-Tolerance Metabolic Pathways in Sugar Beet


 ,
,
Abstract
1. Introduction
2. Results
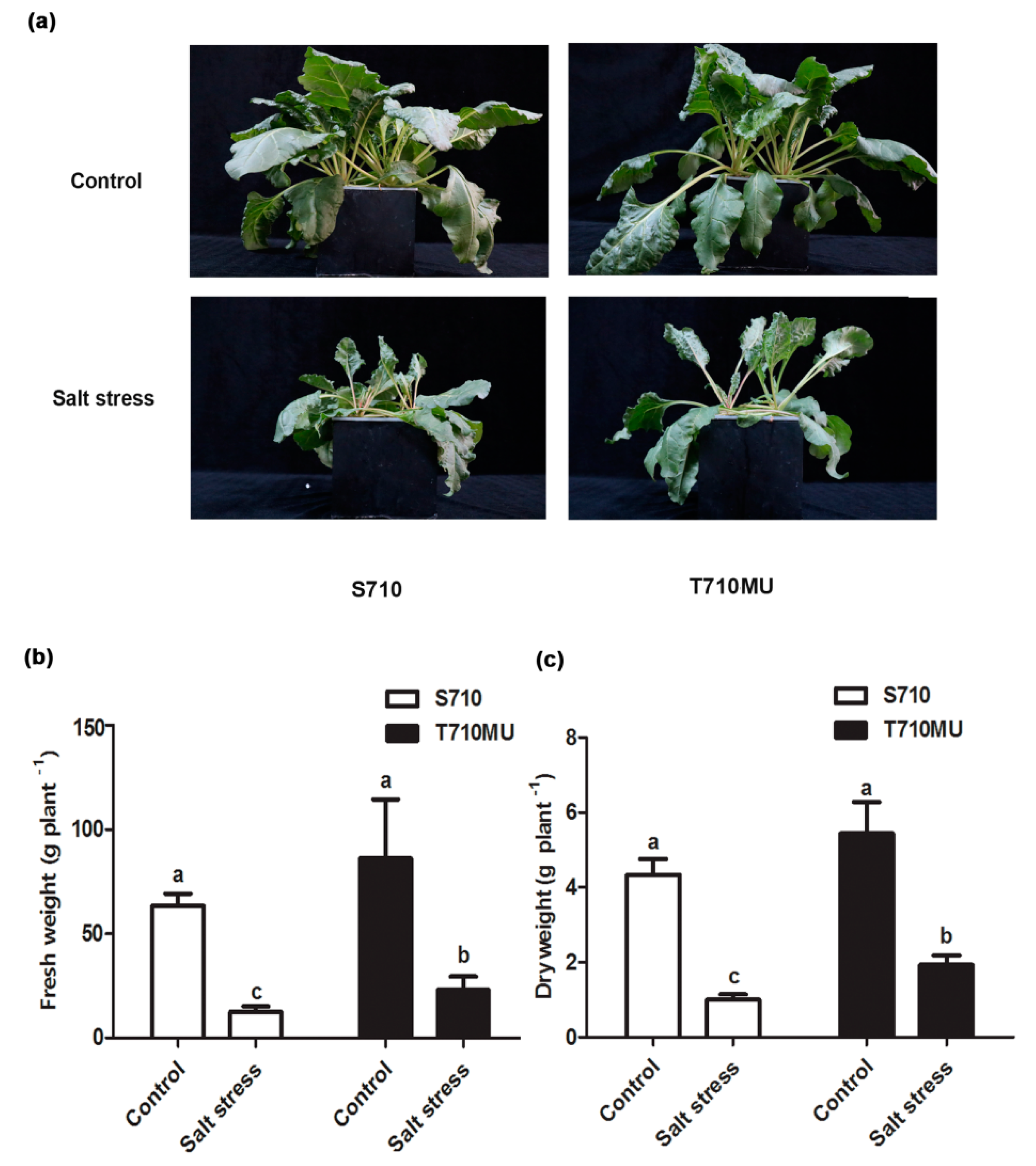
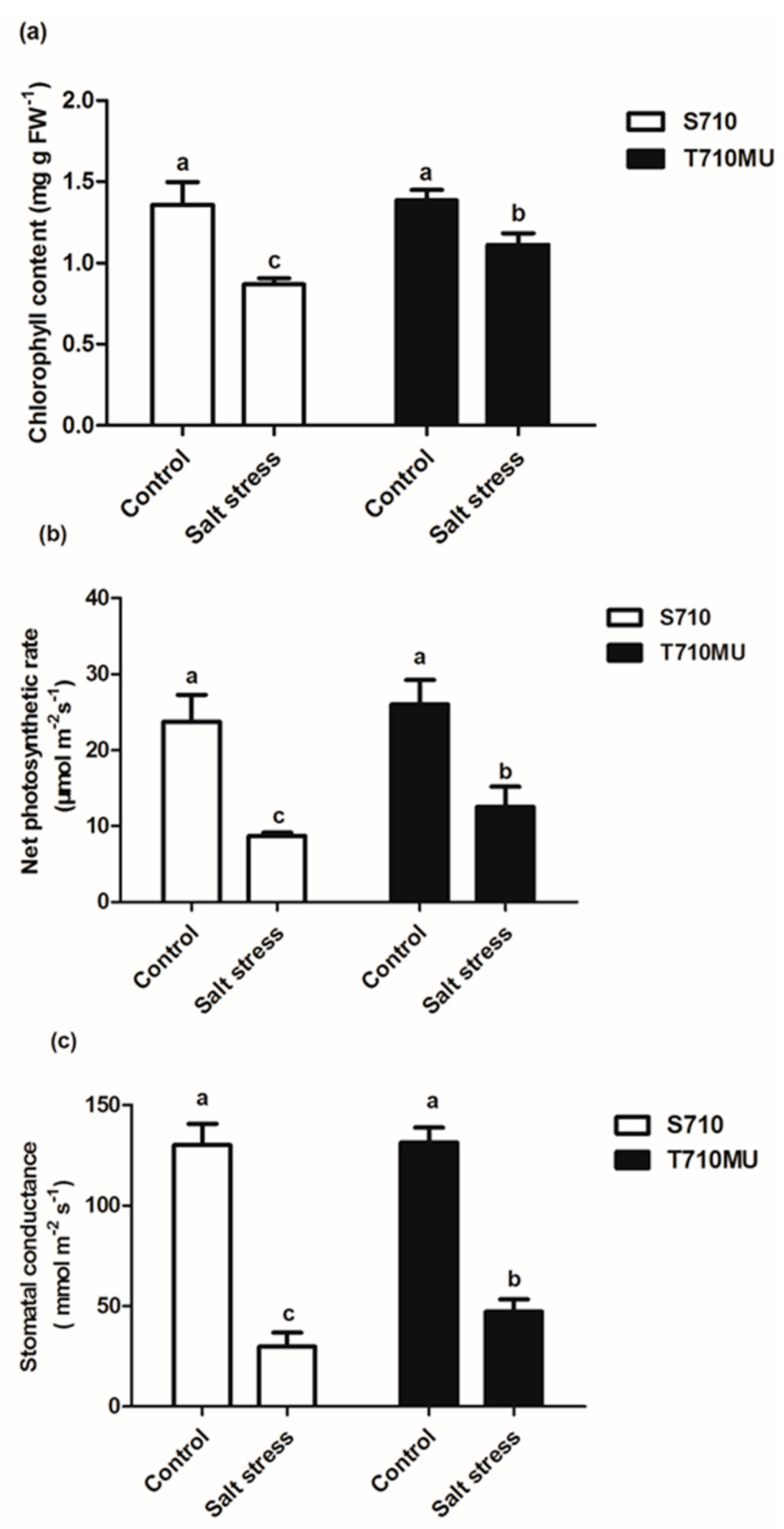
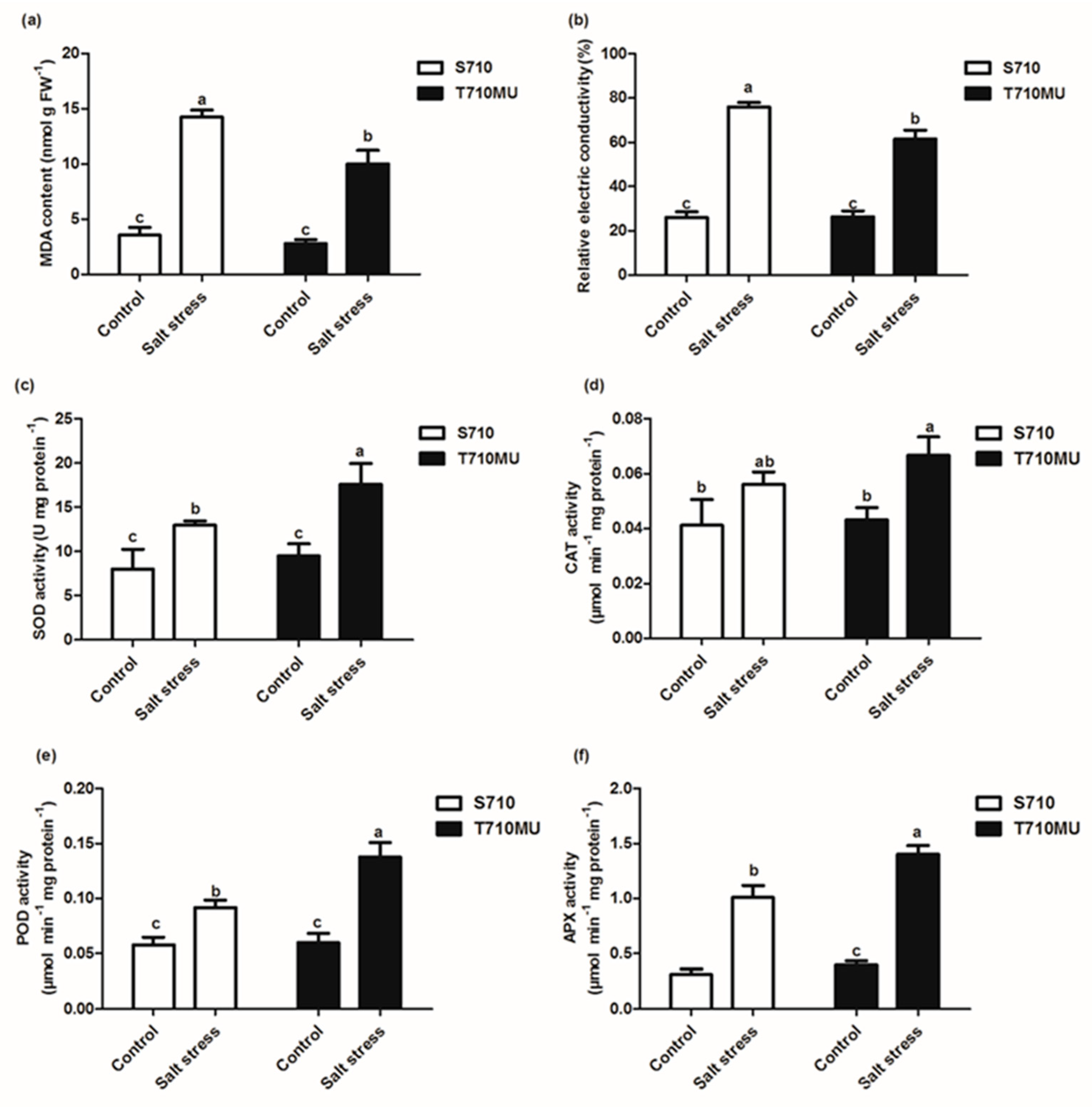
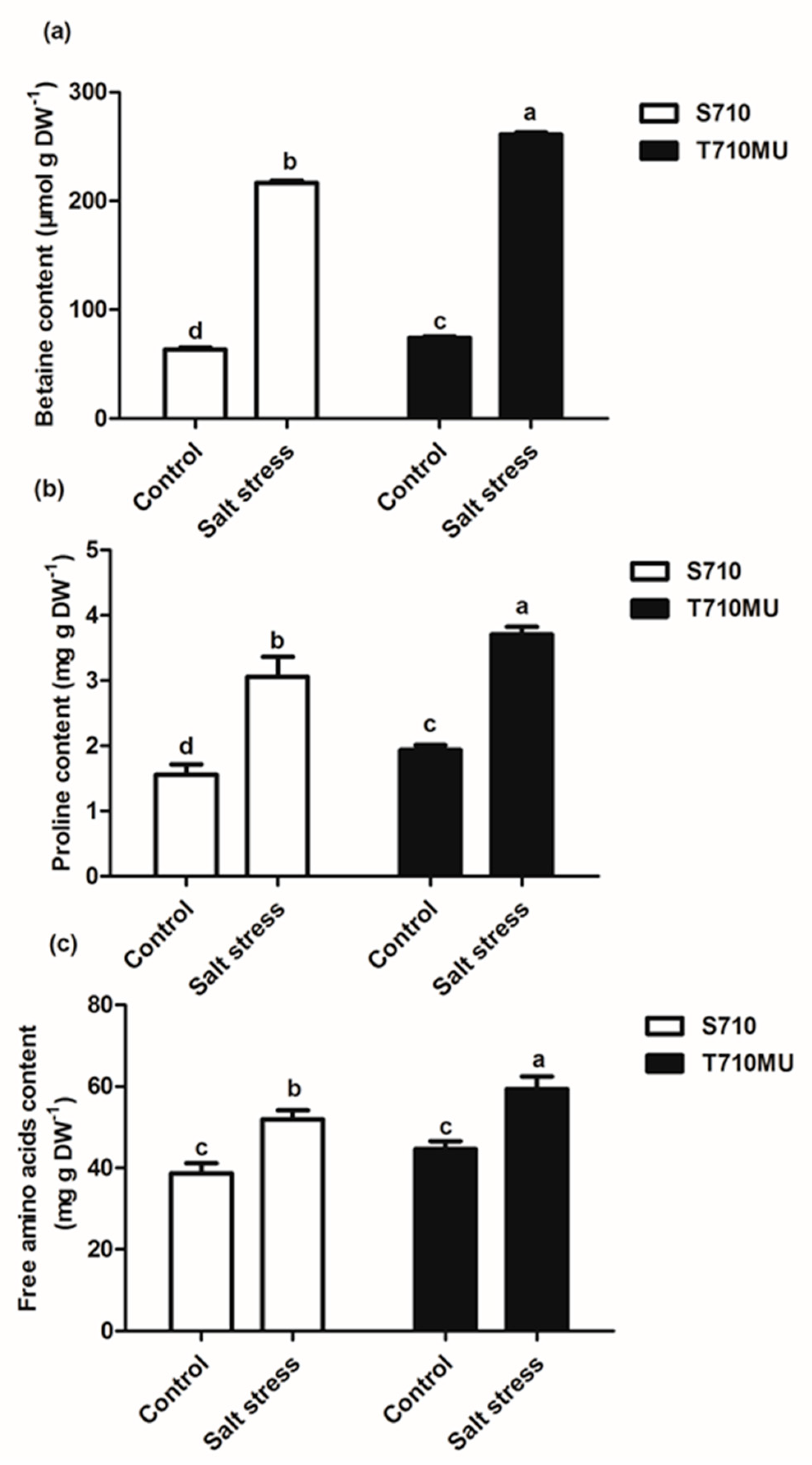
2.1. Physiological Response to Salt Stress in Two suGar Beet Genotypes
2.2. Transcriptome Sequencing and Data Analysis
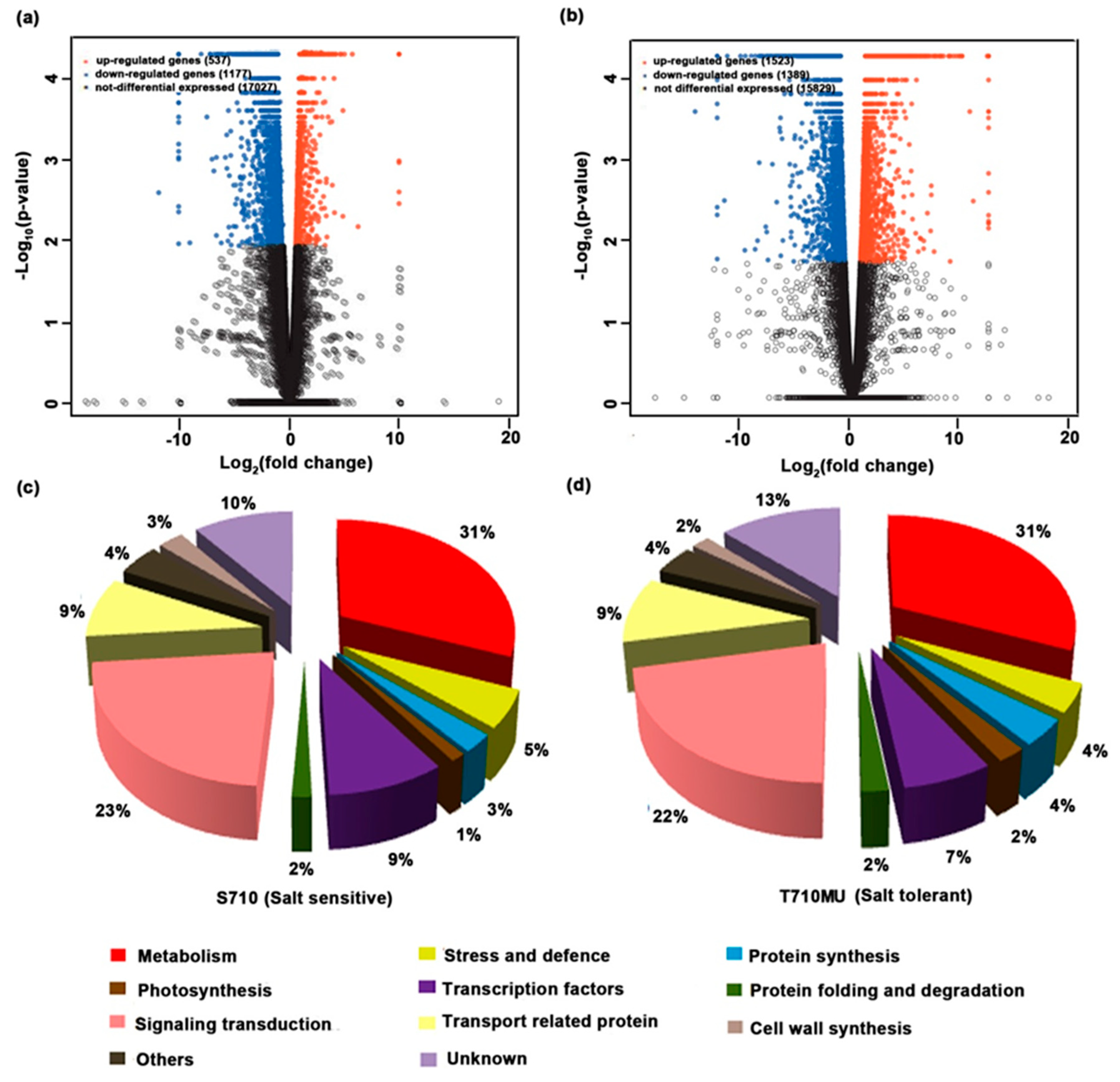
2.3. Identification of Differentially Expressed Genes
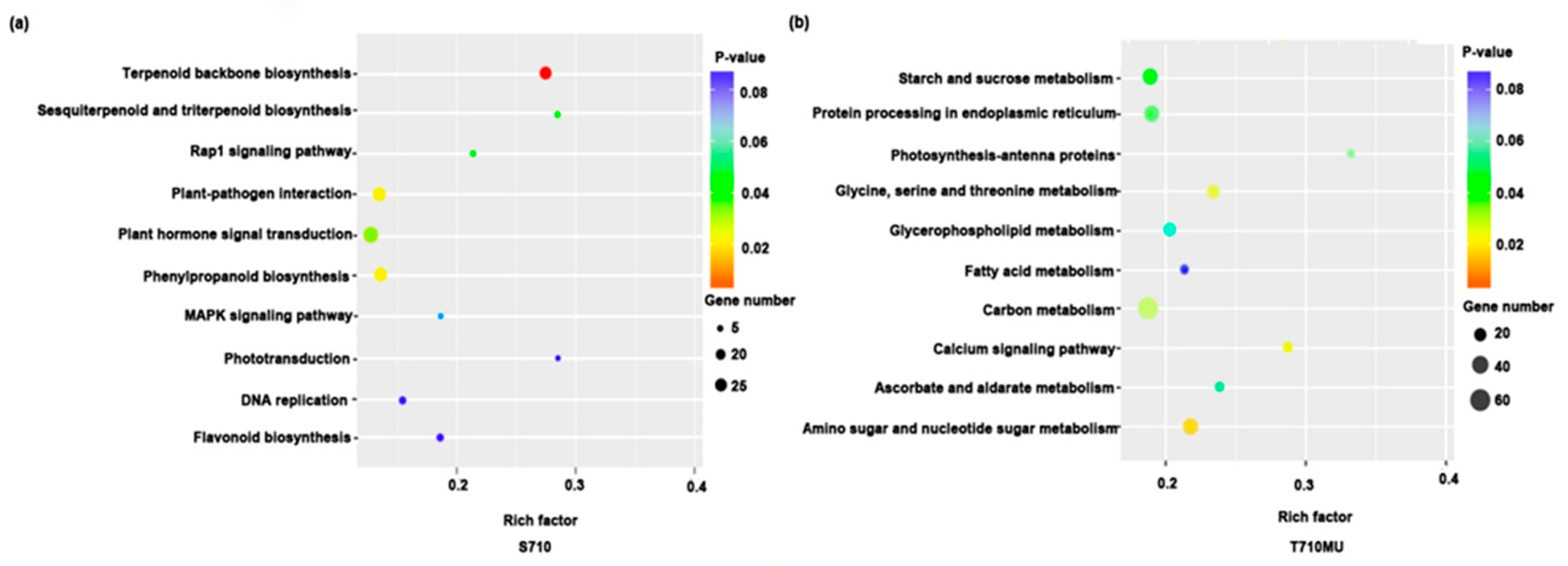
2.4. KEGG Enrichment Analysis of DEGs
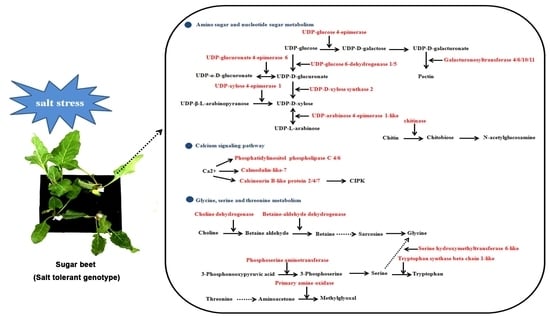
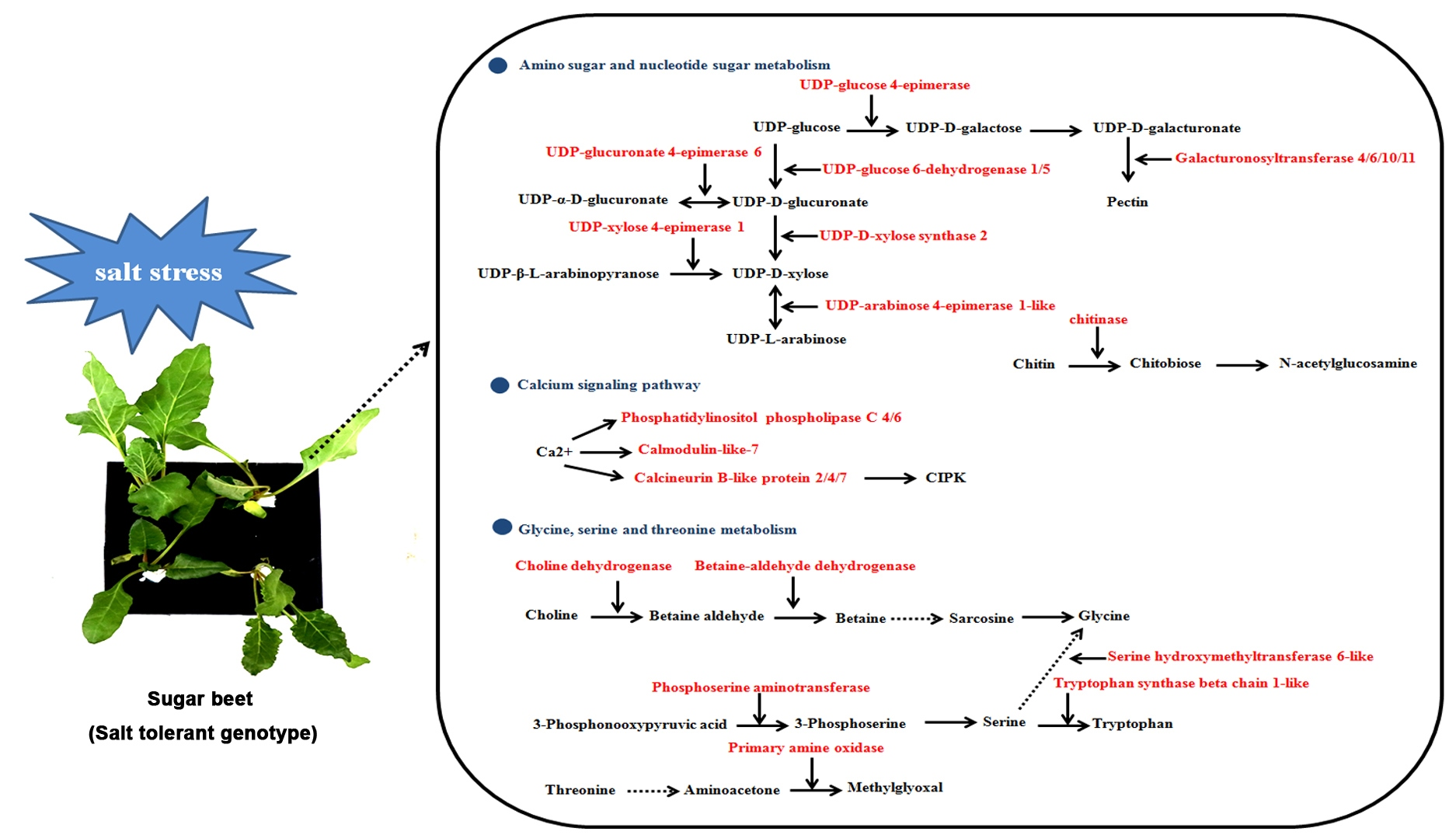
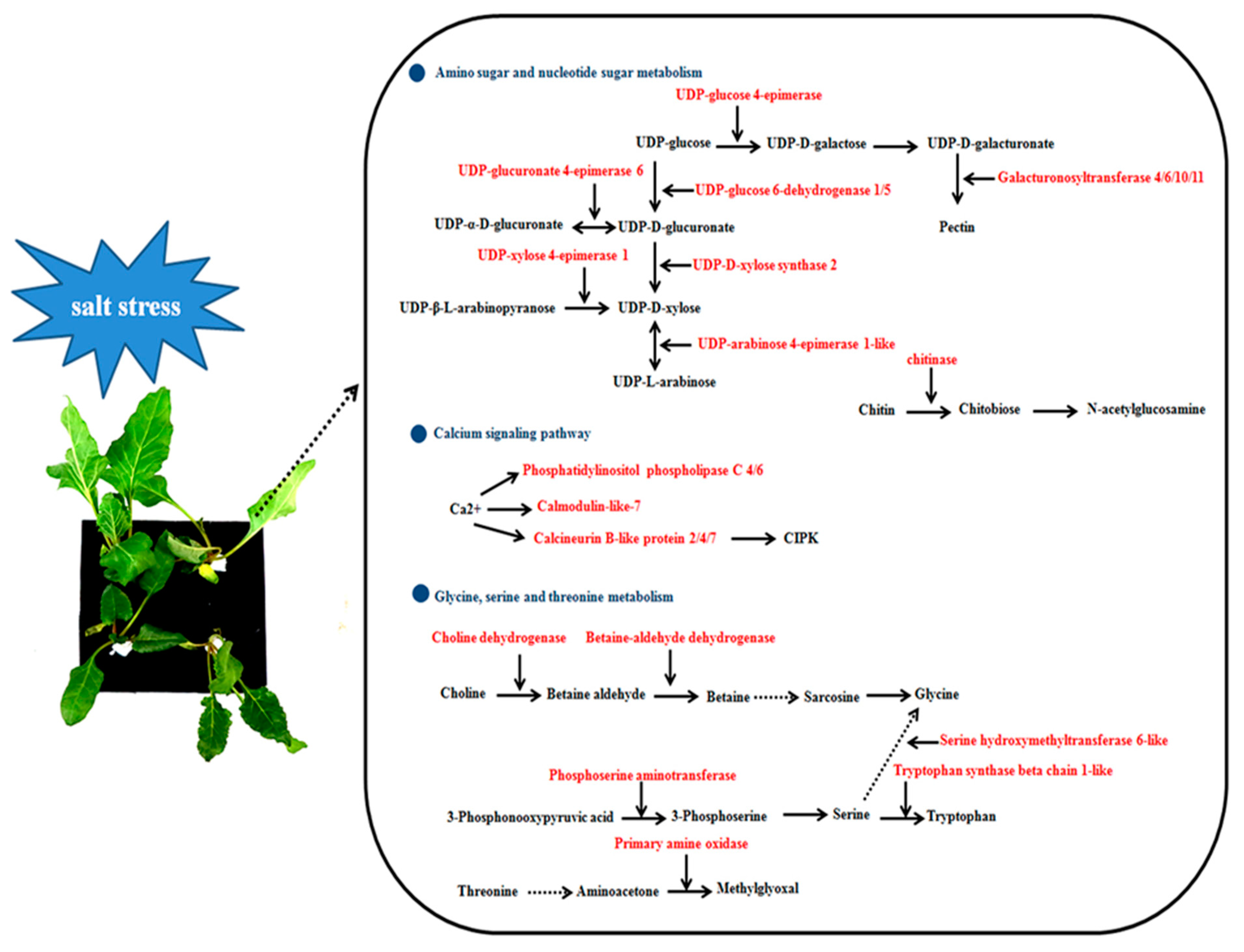
2.5. Amino Sugar and Nucleotide Sugar Metabolism, C-Alcium Signaling Pathway and Glycine, Serine and Threonine Metabolism Are Involved in Sugar Beet Salt Tolerance
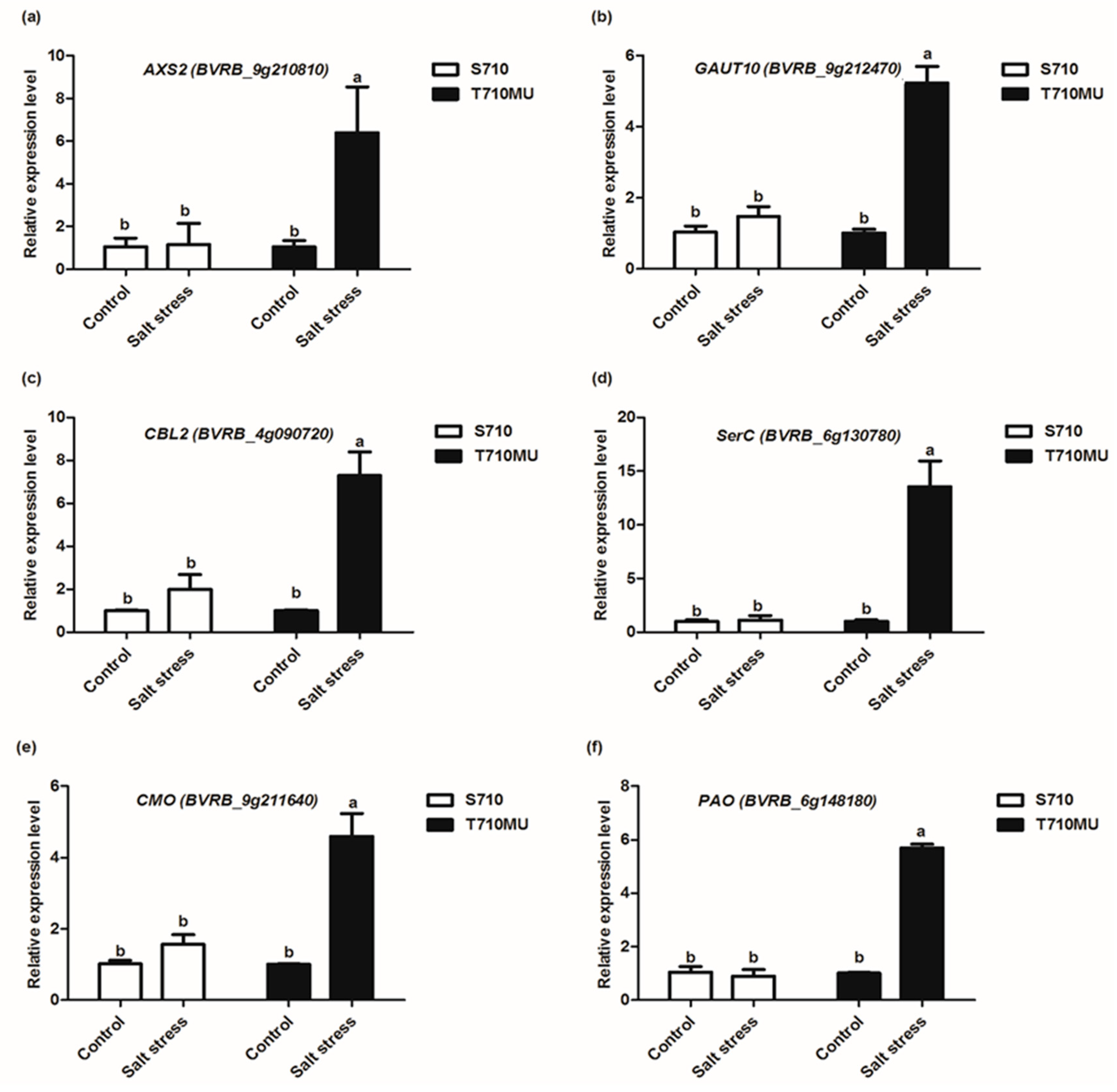
2.6. Validation of Illumina Expression Patterns by qRT-PCR Analysis
3. Discussion
3.1. DEGs Involved in Amino Sugar and Nucleotide Sugar Metabolism
3.2. DEGs Involved in Calcium Signaling Pathway
3.3. DEGs Involved in Glycine, Serine and Threonine Metabolism
3.4. Other DEGs Involved in Salt Stress Tolerance
4. Materials and Methods
4.1. Plant Growth Conditions and Salt Stress Treatments
4.2. Determination of Physiological Indexes
4.3. RNA Extraction, Library Preparation, and Sequencing
4.4. RNA-Seq Data Analysis
4.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abdel Latef, A.; He, C. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salt stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Wang, Y.; Stevanato, P.; Yu, L.; Zhao, H.; Sun, X.; Sun, F.; Li, J.; Geng, G. The physiological and metabolic changes in sugar beet seedlings under different levels of salt stress. J. Plant Res. 2017, 130, 1079–1093. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Chen, S.; Wang, Y. Advances in understanding the physiological and molecular responses of sugar beet to salt stress. Front. Plant Sci. 2019, 10, 1431. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Moray, C.; Hua, X.; Bromham, L. Salt tolerance is evolutionarily labile in a diverse set of angiosperm families. BMC Evol. Biol. 2015, 15, 90. [Google Scholar] [CrossRef]
- Bendaly, A.; Messedi, D.; Smaoui, A.; Ksouri, R.; Bouchereau, A.; Abdelly, C. Physiological and leaf metabolome changes in the xerohalophyte species Atriplex halimus induced by salinity. Plant Physiol. Biochem. 2016, 103, 208–218. [Google Scholar] [CrossRef]
- Daoud, S.; Harrouni, C.; Huchzermeyer, B.; Koyro, H.W. Comparison of salinity tolerance of two related subspecies of Beta vulgaris: The sea beet (Beta vulgaris ssp. maritima) and the sugar beet (Beta vulgaris ssp. vulgaris). In Biosaline Agric. High Salin. Tolerance; Abdelly, C., Öztürk, M., Ashraf, M., Grignon, C., Eds.; Birkhäuser Basel: Basel, Switzerland, 2008; pp. 115–129. [Google Scholar]
- Daoud, S.; Koyro, H.W.; Harrouni, M.C.; Schmidt, A.; Papenbrock, J. Salinity tolerance of Beta vulgaris ssp. maritima. Part II. Physiological and biochemical regulation. In Cash Crop Halophytes: Recent Studies. Tasks for Vegetation Science; Lieth, H., Mochtchenko, M., Eds.; Springer, Dordrecht: Berlin, Germany, 2003; pp. 51–57. [Google Scholar]
- Yang, L.; Ma, C.; Wang, L.; Chen, S.; Li, H. Salt stress induced proteome and transcriptome changes in sugar beet monosomic addition lineM14. J. Plant Physiol. 2012, 169, 839–850. [Google Scholar] [CrossRef]
- Hiz, M.C.; Canher, B.; Niron, H.; Turet, M. Transcriptome analysis of salt tolerant common bean (Phaseolus vulgaris L.) under saline conditions. PLoS ONE 2014, 9, e92598. [Google Scholar] [CrossRef]
- Long, W.H.; Zou, X.L.; Zhang, X.K. Transcriptome analysis of canola (Brassica napus) under salt stress at the germination stage. PLoS ONE 2015, 10, e0116217. [Google Scholar] [CrossRef]
- Lv, X.; Jin, Y.; Wang, Y. De novo transcriptome assembly and identification of salt-responsive genes in sugar beet M14. Comput. Biol. Chem. 2018, 75, 1–10. [Google Scholar] [CrossRef]
- Ji, M.; Wang, K.; Wang, L.; Chen, S.; Li, H.; Ma, C.; Wang, Y. Overexpression of a S-Adenosylmethionine decarboxylase from sugar beet M14 increased Araidopsis salt tolerance. Int. J. Mol. Sci. 2019, 20, 1990. [Google Scholar] [CrossRef] [PubMed]
- Skorupa, M.; Gołębiewski, M.; Kurnik, K.; Niedojadło, J.; Kęsy, J.; Klamkowski, K.; Wójcik, K.; Treder, W.; Tretyn, A.; Tyburski, J. Salt stress vs. salt shock-the case of sugar beet and its halophytic ancestor. BMC Plant Biol. 2019, 19, 57. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Lu, R.; Dai, Z.; Yan, A.; Tang, Q.; Cheng, C.; Xu, Y.; Yang, W.; Su, J. Salt-stress response mechanisms using de novo transcriptome sequencing of salt-tolerant and sensitive Corchorus spp. Genotypes. Genes 2017, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, D.; Zhou, R.; Wang, X.; Dossa, K.; Wang, L.; Zhang, Y.; Yu, J.; Gong, H.; Zhang, X.; et al. Transcriptome and metabolome analyses of two contrasting sesame genotypes reveal the crucial biological pathways involved in rapid adaptive response to salt stress. BMC Plant Biol. 2019, 19, 66. [Google Scholar] [CrossRef]
- Lee, D.K.; Ahn, S.; Cho, H.Y.; Yun, H.Y.; Park, J.H.; Lim, J. Metabolic response induced by parasitic plant-fungus interactions hinder amino sugar and nucleotide sugar metabolism in the host. Sci. Rep. 2016, 6, 37434. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.W.; Verma, R.; Kim, M.; Lee, J.Y.; Kim, Y.K.; Bang, J.W. Depletion of udp-d-apiose/udp-d-xylose synthases results in rhamnogalacturonan-ii deficiency, cell wall thickening, and cell death in higher plants. J. Biol. Chem. 2006, 281, 13708–13716. [Google Scholar] [CrossRef]
- Godoy, F.D.; Bermúdez, L.; Lira, B.S.; Souza, A.P.D.; Elbl, P.; Demarco, D. Galacturonosyltransferase 4 silencing alters pectin composition and carbon partitioning in tomato. J. Exp. Bot. 2013, 64, 2449–2466. [Google Scholar] [CrossRef]
- Yin, Y.; Chen, H.; Hahn, M.G.; Mohnen, D.; Xu, Y. Evolution and function of the plant cell wall synthesis-related glycosyltransferase family 8. Plant Physiol. 2010, 153, 1729–1746. [Google Scholar] [CrossRef]
- Kong, W.; Gong, Z.; Zhong, H.; Zhang, Y.; Zhao, G.; Gautam, M.; Deng, X.; Liu, C.; Zhang, C.; Li, Y. Expansion and evolutionary patterns of glycosyltransferase family 8 in gramineae crop genomes and their expression under salt and cold stresses in Oryza sativa ssp. japonica. Biomolecules 2019, 9, 188. [Google Scholar] [CrossRef]
- Magnan, F.; Ranty, B.; Charpenteau, M.; Sotta, B.; Galaud, J.P.; Aldon, D. Mutations in AtCML9, a calmodulin-like protein from Arabidopsis thaliana, alter plant responses to abiotic stress and abscisic acid. Plant J. 2008, 56, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Kudla, J.; Batistic, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Tang, R.J.; Zheng, X.J.; Wang, S.M.; Luan, S. The calcium sensor CBL7 modulates plant responses to low nitrate in Arabidopsis. Biochem. Biophys. Res. Commun. 2015, 468, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Nunes, N.A.; Araújo, W.L.; Braun, H.P. Amino acid catabolism in plants. Mol. Plant. 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, J.; Zhu, T.; Zhao, C.; Li, L.; Chen, M. The role of melatonin in salt stress responses. Int. J. Mol. Sci. 2019, 20, 1735. [Google Scholar] [CrossRef]
- Yamada, N.; Promden, W.; Yamane, K.; Tamagake, H.; Hibino, T.; Tanaka, Y.; Takabe, T. Preferential accumulation of betaine uncoupled to choline monooxygenase in young leaves of sugar beet-importance of long-distance translocation of betaine under normal and salt-stressed conditions. J. Plant Physiol. 2009, 166, 2058–2070. [Google Scholar] [CrossRef]
- Zhang, Q.; Song, X.; Bartels, D. Sugar metabolism in the desiccation tolerant grass Oropetium thomaeum in response to environmental stresses. Plant Sci. 2018, 270, 30–36. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, T.; An, X.; Gu, R. Evolution and expression analysis of the β-glucosidase (glu) encoding gene subfamily in maize. Genes Genom. 2012, 34, 179–187. [Google Scholar] [CrossRef]
- Lee, K.H.; Piao, H.L.; Kim, H.Y.; Choi, S.M.; Jiang, F.; Hartung, W.; Hwang, I.; Kwak, J.M.; Lee, I.J.; Hwang, I. Activation of glucosidase via stress-induced polymerization rapidly increases active pools of abscisic acid. Cell 2006, 126, 1109–1120. [Google Scholar] [CrossRef]
- Shafi, A.; Chauhan, R.; Gill, T.; Swarnkar, M.K.; Sreenivasulu, Y.; Kumar, S.; Kumar, N.; Shankar, R.; Ahuja, P.S.; Singh, A.K. Expression of SOD and APX genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol. Biol. 2015, 87, 615–631. [Google Scholar] [CrossRef]
- Survila, M.; Davidsson, P.R.; Pennanen, V.; Kariola, T.; Broberg, M.; Sipari, N.; Heino, P.; Palva, E.T. Peroxidase-generated apoplastic ROS impair cuticle integrity and contribute to DAMP-elicited defenses. Front. Plant Sci. 2016, 23, 1945. [Google Scholar] [CrossRef]
- Nahirñak, V.; Almasia, N.; Hopp, H.E.; Vazquez-Rovere, C. Snakin/GASA proteins: Involvement in hormone crosstalk and redox homeostasis. Plant Signal Behav. 2012, 7, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, R.; Han, T.; Cai, W.; Fu, Z.; Lu, Y. Salt stress reduces root meristem size by nitric oxide-mediated modulation of auxin accumulation and signaling in Arabidopsis. Plant Physiol. 2015, 168, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Paque, S.; Mouille, G.; Grandont, L.; Alabadi, D.; Gaertner, C.; Goyallon, A.; Muller, P.; Primard-Brisset, C.; Sormani, R.; Blázquez, M.A.; et al. AUXIN BINDING PROTEIN1 links cell wall remodeling, auxin signaling, and cell expansion in Arabidopsis. Plant Cell 2014, 26, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Brini, F.; Amara, I.; Feki, K.; Hanin, M.; Khoudi, H.; Masmoudi, K. Physiological and molecular analyses of seedlings of two Tunisian durum wheat (Triticum turgidum L. subsp Durum [Desf.]) varieties showing contrasting tolerance to salt stress. Acta Physiol. Plant. 2008, 31, 145–154. [Google Scholar] [CrossRef]
- Wang, Y.; Stevanato, P.; Lv, C.; Li, R.; Geng, G. Comparative physiological and proteomic analysis of two sugar beet genotypes with contrasting salt tolerance. J. Agric. Food Chem. 2019, 67, 6056–6073. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhou, Y.; Fan, J.; Fu, Y.; Shen, L.; Yao, Y.; Li, R.; Fu, S.; Duan, R.; Hu, X.; et al. SpBADH of the halophyte Sesuvium portulacastrum strongly confers drought tolerance through ROS scavenging in transgenic Arabidopsis. Plant Physiol. Biochem. 2015, 96, 377–387. [Google Scholar] [CrossRef]
- Ahmada, P.; Johnb, R.; Sarwatc, M.; Umard, S. Responses of proline, lipid peroxidation and antioxidative enzymes in two varieties of Pisum sativum L. under salt stress. Int. J. Plant Prod. 2008, 2, 1735–8043. [Google Scholar]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Wang, Y.; Peng, C.; Zhan, Y.; Yu, L.; Li, M.; Li, J.; Geng, G. Comparative proteomic analysis of two sugar beet cultivars with contrasting drought tolerance. J. Plant Growth Regul. 2017, 36, 537–549. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Annotation | T710MU Log2 (Fold Change) | S710 Log2 (Fold Change) | |
|---|---|---|---|---|
| Amino sugar and nucleotide sugar metabolism | BVRB_9g210810 | UDP-D-xylose synthase 2 | +1.50283 | No change |
| BVRB_9g208380 | Galacturonosyltransferase10 | +1.38083 | No change | |
| BVRB_5g120380 | Galacturonosyltransferase 6 | +1.91287 | No change | |
| BVRB_8g184210 | Galacturonosyltransferase 4 | +1.31858 | No change | |
| BVRB_5g113520 | Galacturonosyltransferase 11 | +1.15123 | No change | |
| BVRB_9g212470 | Galacturonosyltransferase 3 | +1.01747 | No change | |
| BVRB_5g113830 | UDP-glucose 6-dehydrogenase 1 | +2.96586 | No change | |
| BVRB_9g204050 | UDP-glucose 6-dehydrogenase 1 | +1.69573 | No change | |
| BVRB_7g169190 | UDP-glucose 6-dehydrogenase 5-like | +1.69210 | No change | |
| BVRB_3g051020 | Acidic endochitinase SE2 | +1.06524 | No change | |
| BVRB_3g058210 | Acidic endochitinase SP2 | +4.00091 | No change | |
| BVRB_5g109600 | UDP-arabinose 4-epimerase 1 | +1.34773 | No change | |
| BVRB_5g100870 | UDP-xylose 4-epimerase 1 | +1.24388 | No change | |
| BVRB_3g065540 | UDP-glucuronate 4-epimerase 6 | +1.06591 | No change | |
| BVRB_8g189980 | Chitinase 6-like | +3.43721 | No change | |
| Calcium signaling pathway | BVRB_9g207120 | Calmodulin-like-7 | +1.07209 | No change |
| BVRB_4g090720 | Calcineurin B-like protein 2 | +1.45304 | No change | |
| BVRB_3g051930 | Calcineurin B-like protein 7 | +2.06100 | No change | |
| BVRB_1g003040 | Calcineurin B-like protein 4 | +1.71420 | No change | |
| BVRB_7g172070 | Phosphoinositide phospholipase C 4 | +2.26537 | No change | |
| BVRB_2g023760 | Phosphoinositide phospholipase C 6 | +2.05940 | No change | |
| Glycine, serine and threonine metabolism | BVRB_2g026140 | Tryptophan synthase beta chain 1-like | +2.3822 | No change |
| BVRB_6g130780 | Phosphoserine aminotransferase | +1.36011 | No change | |
| BVRB_6g143730 | Serine hydroxymethyltransferase 6-like | +1.0856 | No change | |
| BVRB_7g169880 | Betaine-aldehyde dehydrogenase 7 | +1.2431 | No change | |
| BVRB_9g211640 | Choline monooxygenase | +1.01747 | No change | |
| BVRB_5g106360 | Glycine cleavage system H protein | +1.62749 | No change | |
| BVRB_6g148180 | Primary amine oxidase | +1.23031 | No change |
| Gene ID | Annotation | T710MU Log2 (Fold Change) | S710 Log2 (Fold Change) | |
|---|---|---|---|---|
| BVRB_4g087470 | Beta-glucosidase 18-like | +2.87219 | No change | |
| BVRB_9g214730 | Snakin-2-like | +5.05443 | No change | |
| BVRB_5g119620 | Vicilin-like antimicrobial peptides 2–3 | +3.99004 | No change | |
| BVRB_2g029800 | Peroxidase P7-like | +2.99716 | No change | |
| BVRB_9g206190 | Superoxide dismutase [Cu-Zn] | +1.15195 | No change | |
| BVRB_3g058070 | Auxin-binding protein ABP19a-like | +1.34462 | No change | |
| BVRB_7g160890 | Auxin-binding protein ABP19a | +13.1000 | −1.49 | |
| BVRB_5g111820 | Auxin transporter-like protein 4 | +4.38131 | No change |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, G.; Lv, C.; Stevanato, P.; Li, R.; Liu, H.; Yu, L.; Wang, Y. Transcriptome Analysis of Salt-Sensitive and Tolerant Genotypes Reveals Salt-Tolerance Metabolic Pathways in Sugar Beet. Int. J. Mol. Sci. 2019, 20, 5910. https://doi.org/10.3390/ijms20235910
Geng G, Lv C, Stevanato P, Li R, Liu H, Yu L, Wang Y. Transcriptome Analysis of Salt-Sensitive and Tolerant Genotypes Reveals Salt-Tolerance Metabolic Pathways in Sugar Beet. International Journal of Molecular Sciences. 2019; 20(23):5910. https://doi.org/10.3390/ijms20235910
Chicago/Turabian StyleGeng, Gui, Chunhua Lv, Piergiorgio Stevanato, Renren Li, Hui Liu, Lihua Yu, and Yuguang Wang. 2019. "Transcriptome Analysis of Salt-Sensitive and Tolerant Genotypes Reveals Salt-Tolerance Metabolic Pathways in Sugar Beet" International Journal of Molecular Sciences 20, no. 23: 5910. https://doi.org/10.3390/ijms20235910
APA StyleGeng, G., Lv, C., Stevanato, P., Li, R., Liu, H., Yu, L., & Wang, Y. (2019). Transcriptome Analysis of Salt-Sensitive and Tolerant Genotypes Reveals Salt-Tolerance Metabolic Pathways in Sugar Beet. International Journal of Molecular Sciences, 20(23), 5910. https://doi.org/10.3390/ijms20235910