Remote Ischemic Preconditioning and Diazoxide Protect from Hepatic Ischemic Reperfusion Injury by Inhibiting HMGB1-Induced TLR4/MyD88/NF-κB Signaling

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
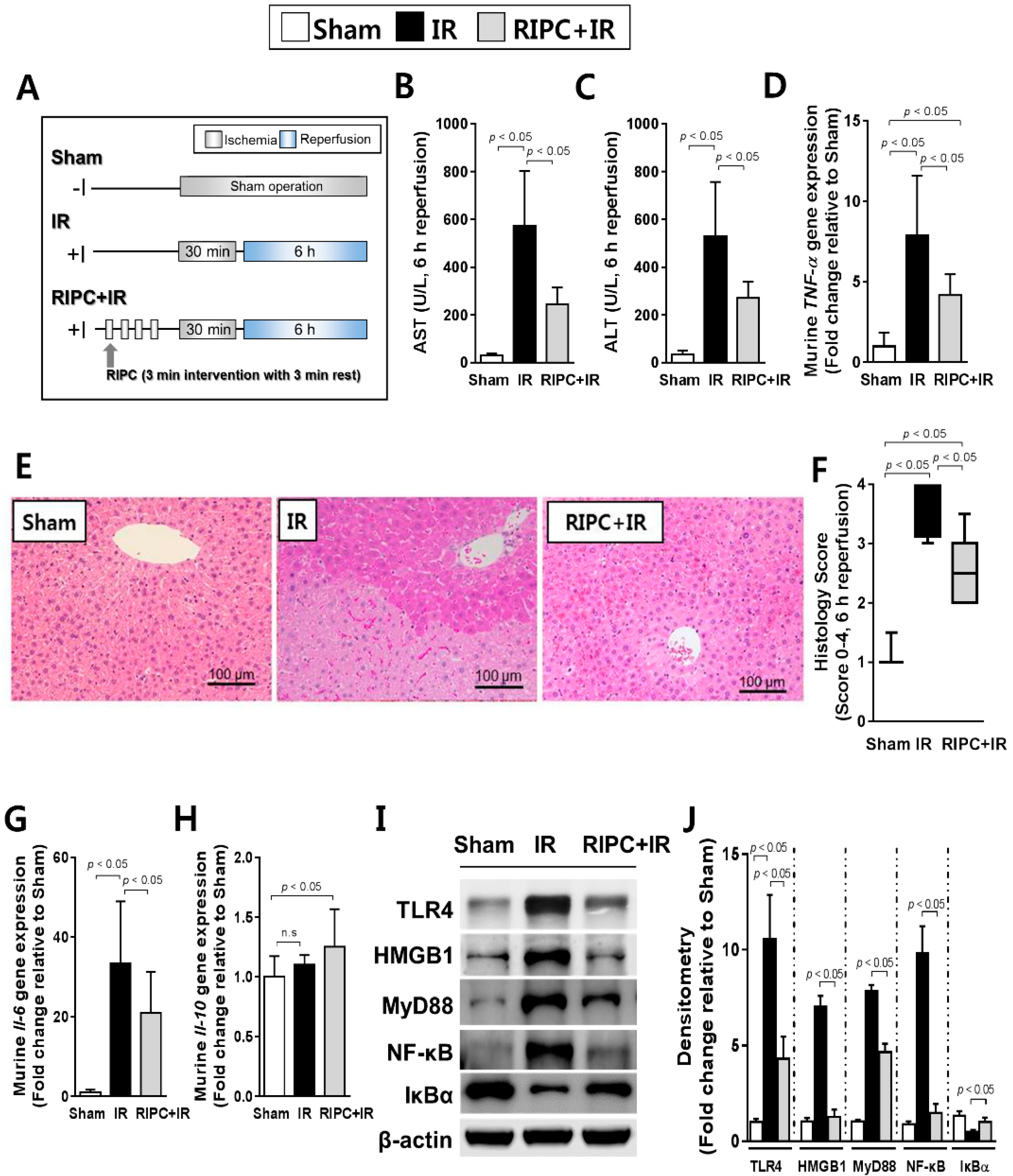
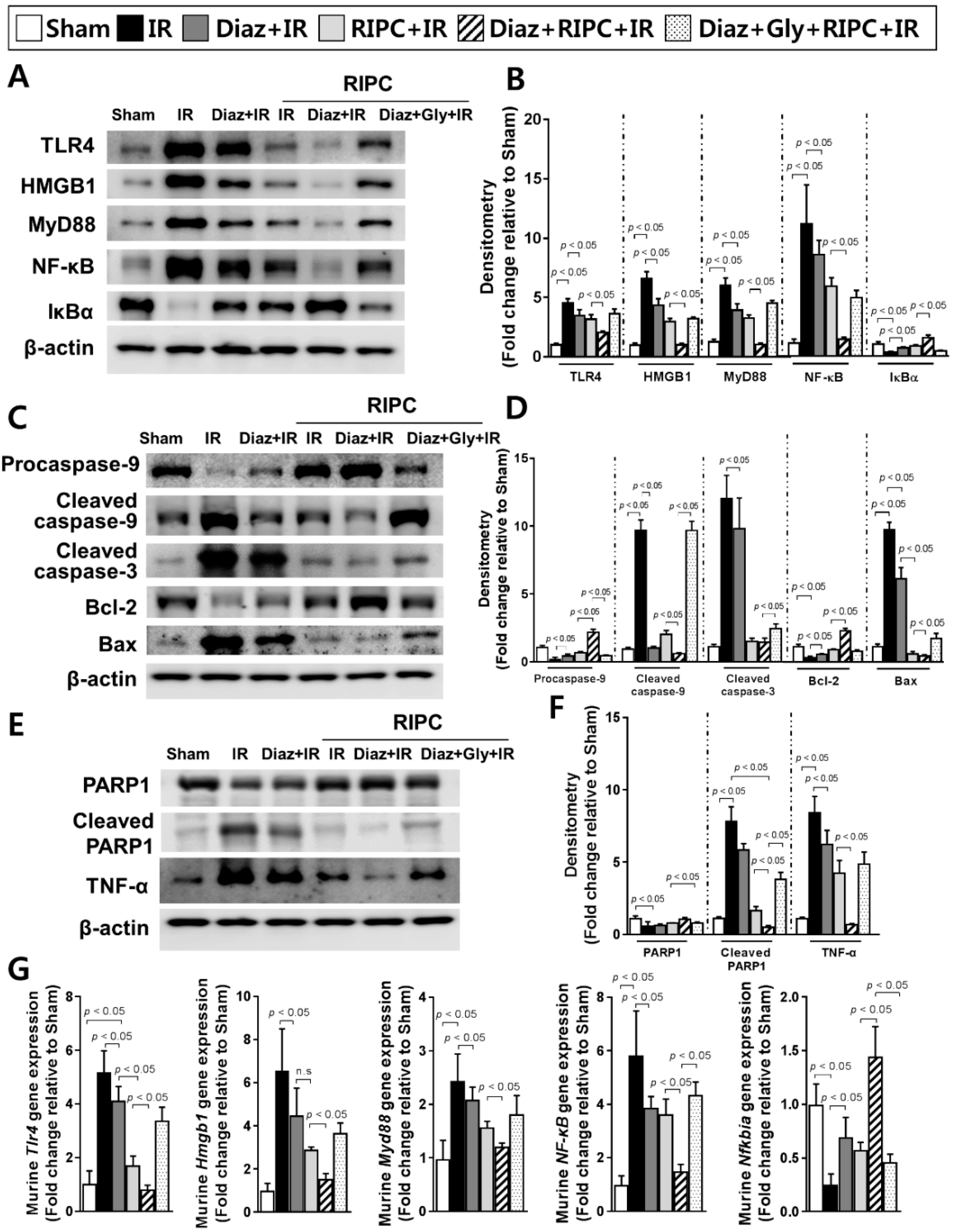
2.1. RIPC Decreases Hepatic IR Injury by Inhibition of the TLR4/MyD88/NF-κB Signaling
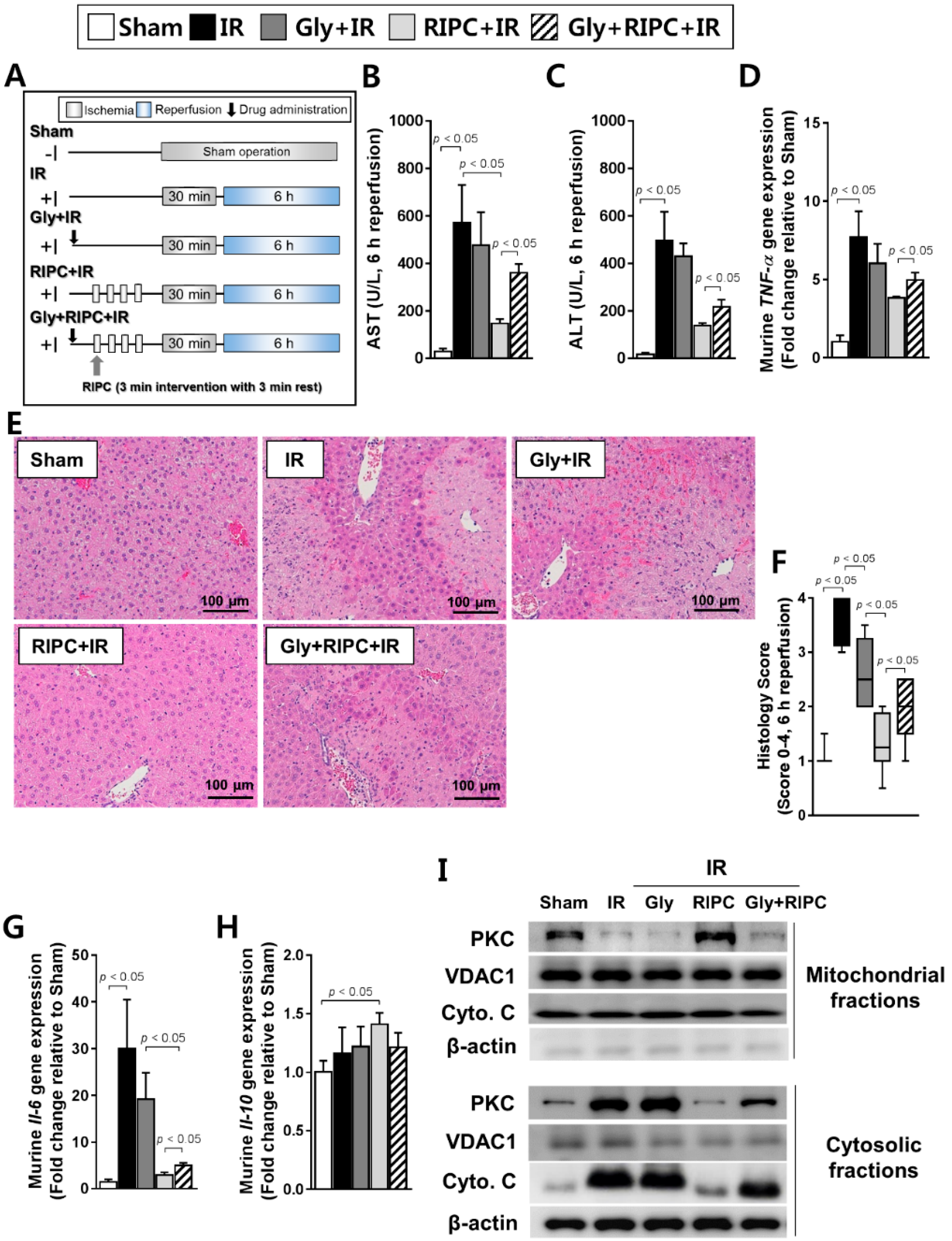
2.2. The Effect of RIPC on Hepatic IR Injury Is Attenuated by KATP Blocker (Glyburide)
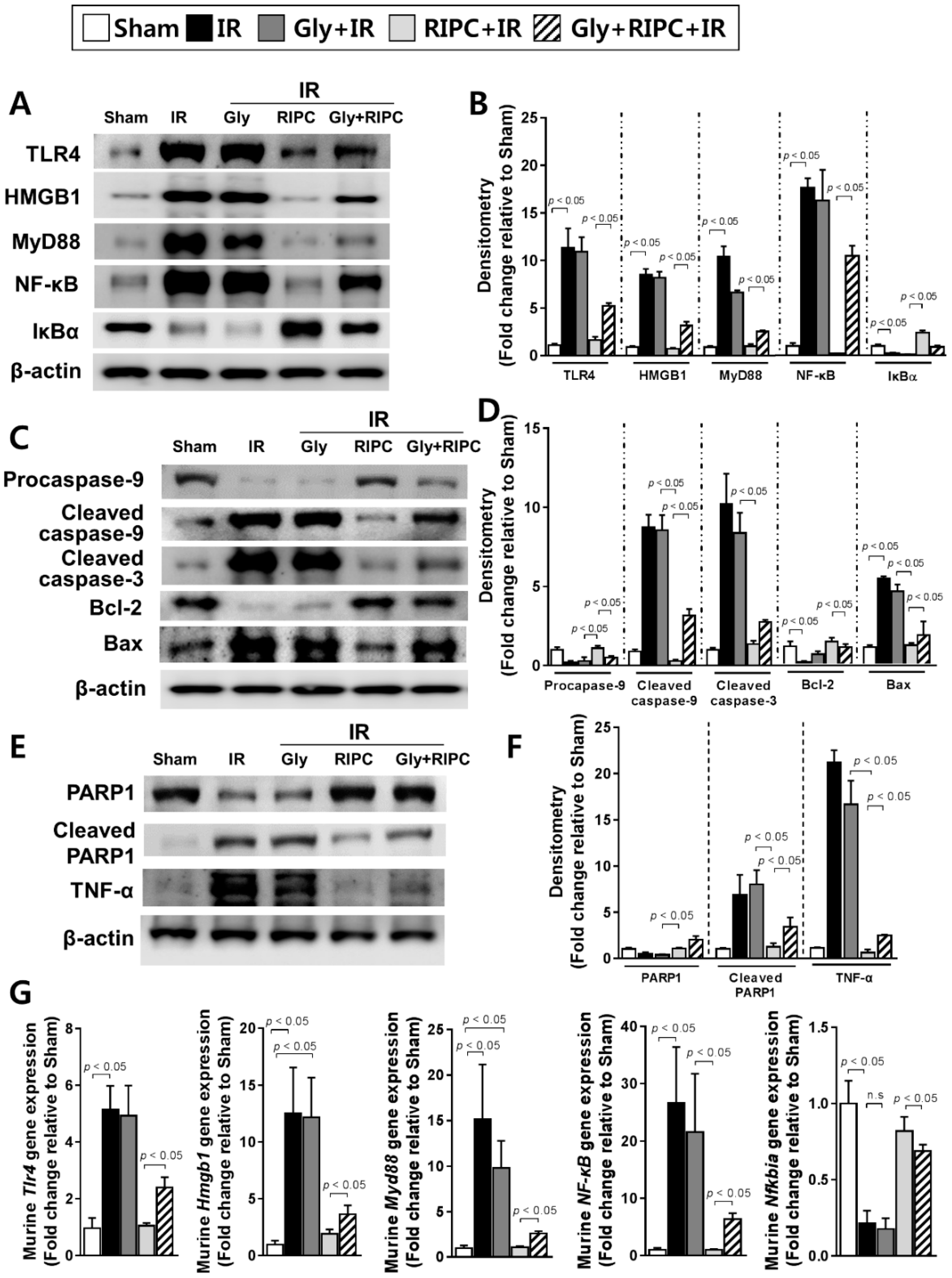
2.3. Glyburide Decreases the Effect of RIPC by Attenuation of the TLR4/MyD88/NF-κB Signaling
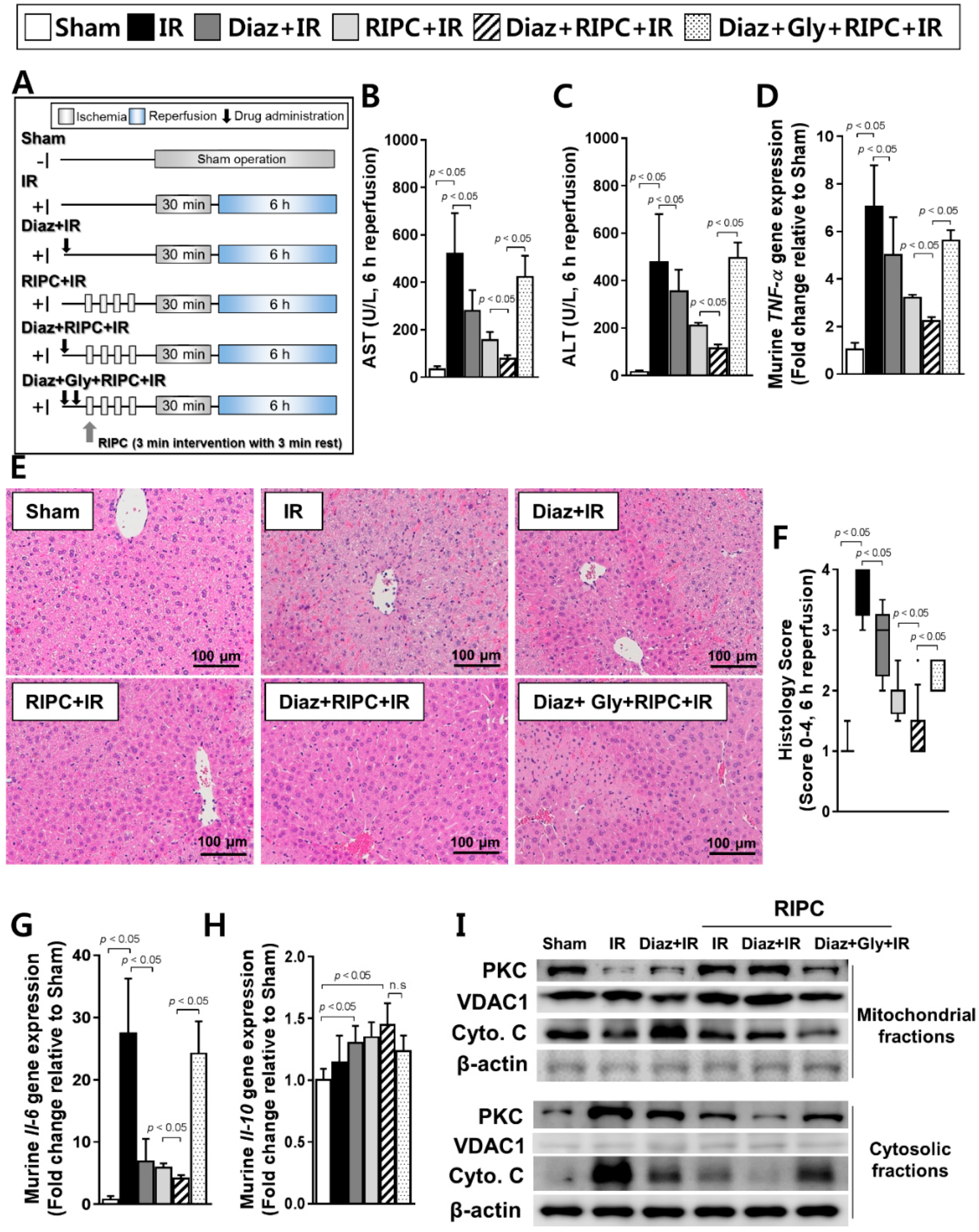
2.4. Additive Effects of KATP Agonist (Diazoxide) and RIPC on Acute Hepatic IR Injury
2.5. Diazoxide Prevents Hepatic Injury by Attenuating the TLR4/MyD88/NF-κB Pathway
3. Discussion
4. Materials and Methods
4.1. Study Group
4.2. Induction of RIPC and Liver IR
4.3. Blood Sampling and Laboratory Tests
4.4. Isolation of mRNA and RT-PCR
4.5. Immunoblotting
4.6. Hepatic Tissue Histology
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Klune, J.R.; Tsung, A. Molecular biology of liver ischemia/reperfusion injury: Established mechanisms and recent advancements. Surg. Clin. N. Am. 2010, 90, 665–677. [Google Scholar] [CrossRef]
- Carden, D.L.; Granger, D.N. Pathophysiology of ischaemia-reperfusion injury. J. Pathol. 2000, 190, 255–266. [Google Scholar] [CrossRef]
- Zhou, H.; Zhu, J.; Yue, S.; Lu, L.; Busuttil, R.W.; Kupiec-Weglinski, J.W.; Wang, X.; Zhai, Y. The dichotomy of endoplasmic reticulum stress response in liver ischemia-reperfusion injury. Transplantation 2016, 100, 365–372. [Google Scholar] [CrossRef]
- Kimura, K.; Shirabe, K.; Yoshizumi, T.; Takeishi, K.; Itoh, S.; Harimoto, N.; Ikegami, T.; Uchiyama, H.; Okano, S.; Maehara, Y. Ischemia-reperfusion injury in fatty liver is mediated by activated NADPH oxidase 2 in rats. Transplantation 2016, 100, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Massip-Salcedo, M.; Rosello-Catafau, J.; Prieto, J.; Avila, M.A.; Peralta, C. The response of the hepatocyte to ischemia. Liver Int. 2007, 27, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Bahde, R.; Spiegel, H.U. Hepatic ischaemia-reperfusion injury from bench to bedside. Br. J. Surg. 2010, 97, 1461–1475. [Google Scholar] [CrossRef] [PubMed]
- Lesurtel, M.; Lehmann, K.; de Rougemont, O.; Clavien, P.A. Clamping techniques and protecting strategies in liver surgery. HPB (Oxford) 2009, 11, 290–295. [Google Scholar] [CrossRef][Green Version]
- Abu-Amara, M.; Yang, S.Y.; Quaglia, A.; Rowley, P.; Tapuria, N.; Seifalian, A.M.; Fuller, B.J.; Davidson, B.R. Effect of remote ischemic preconditioning on liver ischemia/reperfusion injury using a new mouse model. Liver Transplant. 2011, 17, 70–82. [Google Scholar] [CrossRef]
- Kanoria, S.; Jalan, R.; Davies, N.A.; Seifalian, A.M.; Williams, R.; Davidson, B.R. Remote ischaemic preconditioning of the hind limb reduces experimental liver warm ischaemia-reperfusion injury. Br. J. Surg. 2006, 93, 762–768. [Google Scholar] [CrossRef]
- Li, D.Y.; Shi, X.J.; Li, W.; Sun, X.D.; Wang, G.Y. Ischemic preconditioning and remote ischemic preconditioning provide combined protective effect against ischemia/reperfusion injury. Life Sci. 2016, 150, 76–80. [Google Scholar] [CrossRef]
- Oberkofler, C.E.; Limani, P.; Jang, J.H.; Rickenbacher, A.; Lehmann, K.; Raptis, D.A.; Ungethuem, U.; Tian, Y.; Grabliauskaite, K.; Humar, R.; et al. Systemic protection through remote ischemic preconditioning is spread by platelet-dependent signaling in mice. Hepatology 2014, 60, 1409–1417. [Google Scholar] [CrossRef]
- Lai, I.R.; Chang, K.J.; Chen, C.F.; Tsai, H.W. Transient limb ischemia induces remote preconditioning in liver among rats: The protective role of heme oxygenase-1. Transplantation 2006, 81, 1311–1317. [Google Scholar] [CrossRef]
- Hu, X.; Yang, Z.; Yang, M.; Qian, J.; Cahoon, J.; Xu, J.; Sun, S.; Tang, W. Remote ischemic preconditioning mitigates myocardial and neurological dysfunction via K(ATP) channel activation in a rat model of hemorrhagic shock. J. Exp. Med. 2014, 42, 228–233. [Google Scholar] [CrossRef]
- McDonald, K.A.; Huang, H.; Tohme, S.; Loughran, P.; Ferrero, K.; Billiar, T.; Tsung, A. Toll-like receptor 4 (TLR4) antagonist eritoran tetrasodium attenuates liver ischemia and reperfusion injury through inhibition of high-mobility group box protein B1 (HMGB1) signaling. Mol. Med. 2015, 20, 639–648. [Google Scholar] [CrossRef]
- Kevelaitis, E.; Oubenaissa, A.; Mouas, C.; Peynet, J.; Menasche, P. Opening of mitochondrial potassium channels: A new target for graft preservation strategies? Transplantation 2000, 70, 576–578. [Google Scholar] [CrossRef]
- Liu, Y.; Sato, T.; Seharaseyon, J.; Szewczyk, A.; O’Rourke, B.; Marban, E. Mitochondrial ATP-dependent potassium channels. Viable candidate effectors of ischemic preconditioning. Ann. N. Y. Acad. Sci. 1999, 874, 27–37. [Google Scholar] [CrossRef]
- Mabanta, L.; Valane, P.; Borne, J.; Frame, M.D. Initiation of remote microvascular preconditioning requires KATP channel activity. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H264–H271. [Google Scholar] [CrossRef][Green Version]
- Konstantinov, I.E.; Li, J.; Cheung, M.M.; Shimizu, M.; Stokoe, J.; Kharbanda, R.K.; Redington, A.N. Remote ischemic preconditioning of the recipient reduces myocardial ischemia-reperfusion injury of the denervated donor heart via a Katp channel-dependent mechanism. Transplantation 2005, 79, 1691–1695. [Google Scholar] [CrossRef]
- Garlid, K.D.; Costa, A.D.; Quinlan, C.L.; Pierre, S.V.; Dos Santos, P. Cardioprotective signaling to mitochondria. J. Mol. Cell. Cardiol. 2009, 46, 858–866. [Google Scholar] [CrossRef]
- Garlid, K.D.; Paucek, P.; Yarov-Yarovoy, V.; Murray, H.N.; Darbenzio, R.B.; D’Alonzo, A.J.; Lodge, N.J.; Smith, M.A.; Grover, G.J. Cardioprotective effect of diazoxide and its interaction with mitochondrial ATP-sensitive K+ channels. Possible mechanism of cardioprotection. Circ. Res. 1997, 81, 1072–1082. [Google Scholar] [CrossRef]
- Wu, Q.; Tang, C.; Zhang, Y.J.; Jiang, Y.; Li, X.W.; Wang, S.G.; Bie, P. Diazoxide suppresses hepatic ischemia/reperfusion injury after mouse liver transplantation by a BCL-2-dependent mechanism. J. Surg. Res. 2011, 169, e155–e166. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Yoshioka, M.; Abe, Y.; Uchinami, H.; Ohba, T.; Ono, K.; Yamamoto, Y. Enhancement of liver regeneration by adenosine triphosphate-sensitive K+ channel opener (diazoxide) after partial hepatectomy. Transplantation 2012, 93, 1094–1100. [Google Scholar] [CrossRef]
- Grossini, E.; Pollesello, P.; Bellofatto, K.; Sigaudo, L.; Farruggio, S.; Origlia, V.; Mombello, C.; Mary, D.A.; Valente, G.; Vacca, G. Protective effects elicited by levosimendan against liver ischemia/reperfusion injury in anesthetized rats. Liver Transplant. 2014, 20, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Huang, H.F.; He, F.; Wu, L.X.; Lin, J.; Chen, M.Q. Diazoxide attenuates ischemia/reperfusion injury via upregulation of heme oxygenase-1 after liver transplantation in rats. World J. Gastroenterol. 2012, 18, 1765–1772. [Google Scholar] [CrossRef]
- Nogueira, M.A.; Coelho, A.M.; Sampietre, S.N.; Patzina, R.A.; Pinheiro da Silva, F.; D’Albuquerque, L.A.; Machado, M.C. Beneficial effects of adenosine triphosphate-sensitive K+ channel opener on liver ischemia/reperfusion injury. World J. Gastroenterol. 2014, 20, 15319–15326. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Abdel-Gaber, S.A.; Amin, E.F.; Ibrahim, S.A.; Mohammed, R.K.; Abdelrahman, A.M. Molecular mechanisms contributing to the protective effect of levosimendan in liver ischemia-reperfusion injury. Eur. J. Pharmacol. 2014, 741, 64–73. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Yu, P.; Chen, M.; Peng, Q.; Wang, Z.; Dong, N. Remote Ischaemic Preconditioning and Sevoflurane Postconditioning Synergistically Protect Rats from Myocardial Injury Induced by Ischemia and Reperfusion Partly via Inhibition TLR4/MyD88/NF-kappaB Signaling Pathway. Cell. Physiol. Biochem. 2017, 41, 21–32. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Li, J.H.; Zhao, H.W.; Zhang, F.Z. Morphine alleviates myocardial ischemia/reperfusion injury in rats by inhibiting TLR4/NF-kappaB signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8616–8624. [Google Scholar] [CrossRef]
- Colletti, L.M.; Cortis, A.; Lukacs, N.; Kunkel, S.L.; Green, M.; Strieter, R.M. Tumor necrosis factor up-regulates intercellular adhesion molecule 1, which is important in the neutrophil-dependent lung and liver injury associated with hepatic ischemia and reperfusion in the rat. Shock 1998, 10, 182–191. [Google Scholar] [CrossRef]
- Tang, Z.Y.; Loss, G.; Carmody, I.; Cohen, A.J. TIMP-3 ameliorates hepatic ischemia/reperfusion injury through inhibition of tumor necrosis factor-alpha-converting enzyme activity in rats. Transplantation 2006, 82, 1518–1523. [Google Scholar] [CrossRef]
- Jo, H.Y.; Kim, S.Y.; Lee, S.; Jeong, S.; Kim, S.J.; Kang, T.M.; Lee, K.Y. Kir3.1 channel is functionally involved in TLR4-mediated signaling. Biochem. Biophys. Res. Commun. 2011, 407, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Chen, M.; Zheng, D.; Li, J.; Song, M.; Zhang, W.; Feng, J.; Lan, J. The opening of ATP-sensitive K+ channels protects H9c2 cardiac cells against the high glucose-induced injury and inflammation by inhibiting the ROS-TLR4-necroptosis pathway. Cell. Physiol. Biochem. 2017, 41, 1020–1034. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Gamboni-Robertson, F.; He, Q.; Svetkauskaite, D.; Kim, J.Y.; Strassheim, D.; Sohn, J.W.; Yamada, S.; Maruyama, I.; Banerjee, A.; et al. High mobility group box 1 protein interacts with multiple Toll-like receptors. Am. J. Physiol. Cell Physiol. 2006, 290, C917–C924. [Google Scholar] [CrossRef] [PubMed]
- Hreggvidsdottir, H.S.; Lundberg, A.M.; Aveberger, A.C.; Klevenvall, L.; Andersson, U.; Harris, H.E. High mobility group box protein 1 (HMGB1)-partner molecule complexes enhance cytokine production by signaling through the partner molecule receptor. Mol. Med. 2012, 18, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.; Arcaroli, J.; Carmody, A.; Wang, H.; Tracey, K.J. HMG-1 as a mediator of acute lung inflammation. J. Immunol. 2000, 165, 2950–2954. [Google Scholar] [CrossRef]
- Andrassy, M.; Volz, H.C.; Igwe, J.C.; Funke, B.; Eichberger, S.N.; Kaya, Z.; Buss, S.; Autschbach, F.; Pleger, S.T.; Lukic, I.K.; et al. High-mobility group box-1 in ischemia-reperfusion injury of the heart. Circulation 2008, 117, 3216–3226. [Google Scholar] [CrossRef]
- Tsung, A.; Sahai, R.; Tanaka, H.; Nakao, A.; Fink, M.P.; Lotze, M.T.; Yang, H.; Li, J.; Tracey, K.J.; Geller, D.A.; et al. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. J. Exp. Med. 2005, 201, 1135–1143. [Google Scholar] [CrossRef]
- Yamamoto, T.; Tajima, Y. HMGB1 is a promising therapeutic target for acute liver failure. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 673–682. [Google Scholar] [CrossRef]
- Yokoi, T.; Yokoyama, Y.; Kokuryo, T.; Yamaguchi, J.; Nagino, M. Inhibition of Toll-like receptor 4 ameliorates experimental postischemic injury in the cholestatic liver through inhibition of high-mobility group box protein b1 (HMGB1) signaling. Surgery 2017, 163, 270–276. [Google Scholar] [CrossRef]
- Sugihara, M.; Sadamori, H.; Nishibori, M.; Sato, Y.; Tazawa, H.; Shinoura, S.; Umeda, Y.; Yoshida, R.; Nobuoka, D.; Utsumi, M.; et al. Anti-high mobility group box 1 monoclonal antibody improves ischemia/reperfusion injury and mode of liver regeneration after partial hepatectomy. Am. J. Surg. 2016, 211, 179–188. [Google Scholar] [CrossRef]
- Zhao, G.; Fu, C.; Wang, L.; Zhu, L.; Yan, Y.; Xiang, Y.; Zheng, F.; Gong, F.; Chen, S.; Chen, G. Down-regulation of nuclear HMGB1 reduces ischemia-induced HMGB1 translocation and release and protects against liver ischemia-reperfusion injury. Sci. Rep. 2017, 7, 46272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, M.; Xie, H.Y.; Zhou, L.; Meng, X.Q.; Shi, J.; Zheng, S. Role of reactive oxygen species in mediating hepatic ischemia-reperfusion injury and its therapeutic applications in liver transplantation. Transplant. Proc. 2007, 39, 1332–1337. [Google Scholar] [CrossRef] [PubMed]
- Tejima, K.; Arai, M.; Ikeda, H.; Tomiya, T.; Yanase, M.; Inoue, Y.; Nagashima, K.; Nishikawa, T.; Watanabe, N.; Omata, M.; et al. Ischemic preconditioning protects hepatocytes via reactive oxygen species derived from Kupffer cells in rats. Gastroenterology 2004, 127, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Tsung, A.; Klune, J.R.; Zhang, X.; Jeyabalan, G.; Cao, Z.; Peng, X.; Stolz, D.B.; Geller, D.A.; Rosengart, M.R.; Billiar, T.R. HMGB1 release induced by liver ischemia involves Toll-like receptor 4 dependent reactive oxygen species production and calcium-mediated signaling. J. Exp. Med. 2007, 204, 2913–2923. [Google Scholar] [CrossRef] [PubMed]
- Eltzschig, H.K.; Ibla, J.C.; Furuta, G.T.; Leonard, M.O.; Jacobson, K.A.; Enjyoji, K.; Robson, S.C.; Colgan, S.P. Coordinated adenine nucleotide phosphohydrolysis and nucleoside signaling in posthypoxic endothelium: Role of ectonucleotidases and adenosine A2B receptors. J. Exp. Med. 2003, 198, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.L.; Much, C.; Kohler, D.; Schittenhelm, J.; Gorzolla, I.C.; Stahl, G.L.; Eltzschig, H.K. Use of a hanging-weight system for liver ischemic preconditioning in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1431–G1440. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koh, W.U.; Kim, J.; Lee, J.; Song, G.-W.; Hwang, G.S.; Tak, E.; Song, J.-G. Remote Ischemic Preconditioning and Diazoxide Protect from Hepatic Ischemic Reperfusion Injury by Inhibiting HMGB1-Induced TLR4/MyD88/NF-κB Signaling. Int. J. Mol. Sci. 2019, 20, 5899. https://doi.org/10.3390/ijms20235899
Koh WU, Kim J, Lee J, Song G-W, Hwang GS, Tak E, Song J-G. Remote Ischemic Preconditioning and Diazoxide Protect from Hepatic Ischemic Reperfusion Injury by Inhibiting HMGB1-Induced TLR4/MyD88/NF-κB Signaling. International Journal of Molecular Sciences. 2019; 20(23):5899. https://doi.org/10.3390/ijms20235899
Chicago/Turabian StyleKoh, Won Uk, Jiye Kim, Jooyoung Lee, Gi-Won Song, Gyu Sam Hwang, Eunyoung Tak, and Jun-Gol Song. 2019. "Remote Ischemic Preconditioning and Diazoxide Protect from Hepatic Ischemic Reperfusion Injury by Inhibiting HMGB1-Induced TLR4/MyD88/NF-κB Signaling" International Journal of Molecular Sciences 20, no. 23: 5899. https://doi.org/10.3390/ijms20235899
APA StyleKoh, W. U., Kim, J., Lee, J., Song, G.-W., Hwang, G. S., Tak, E., & Song, J.-G. (2019). Remote Ischemic Preconditioning and Diazoxide Protect from Hepatic Ischemic Reperfusion Injury by Inhibiting HMGB1-Induced TLR4/MyD88/NF-κB Signaling. International Journal of Molecular Sciences, 20(23), 5899. https://doi.org/10.3390/ijms20235899