Genome-Wide Transcriptome Analysis of Rice Seedlings after Seed Dressing with Paenibacillus yonginensis DCY84T and Silicon

 ,
, 
Abstract

1. Introduction
2. Results and Discussion
2.1. Combined DCY84T and Si Treatment Stimulates Root Growth of Rice Plants
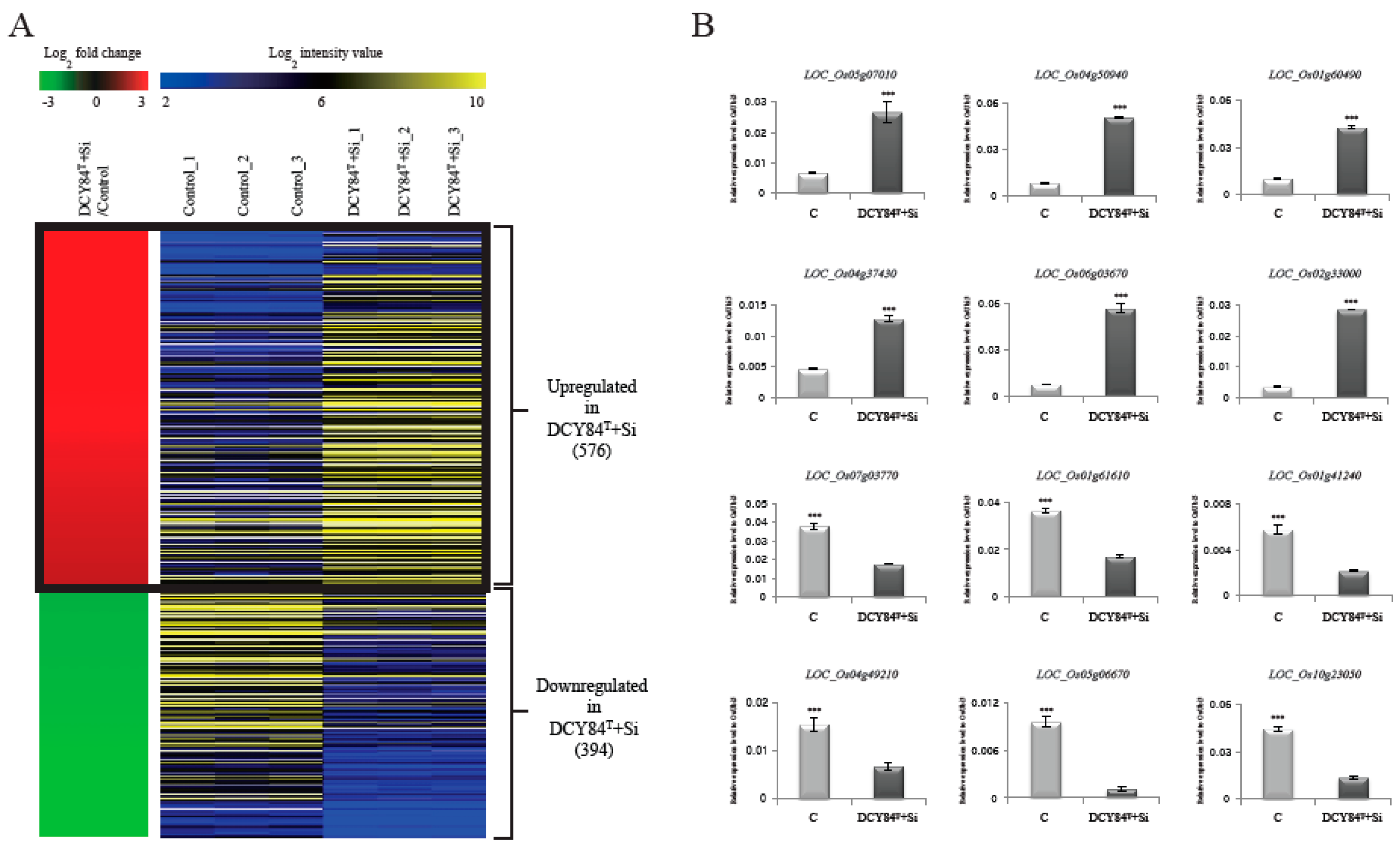
2.2. RNA-Seq Analysis Identified Global Candidate Genes Associated with DCY84T+Si Treatment
2.3. GO Enrichment Analysis Revealed the Significant Biological Processes Associated with DCY84T+Si Treatment
2.4. MapMan Analysis Revealed Involvement of DCY84T+Si Treatment in Phenylpropanoid Metabolism and Ethylene Regulation
2.5. Functions of Candidate Genes Associated with DCY84T+Si Were Evaluated through Literature Searches
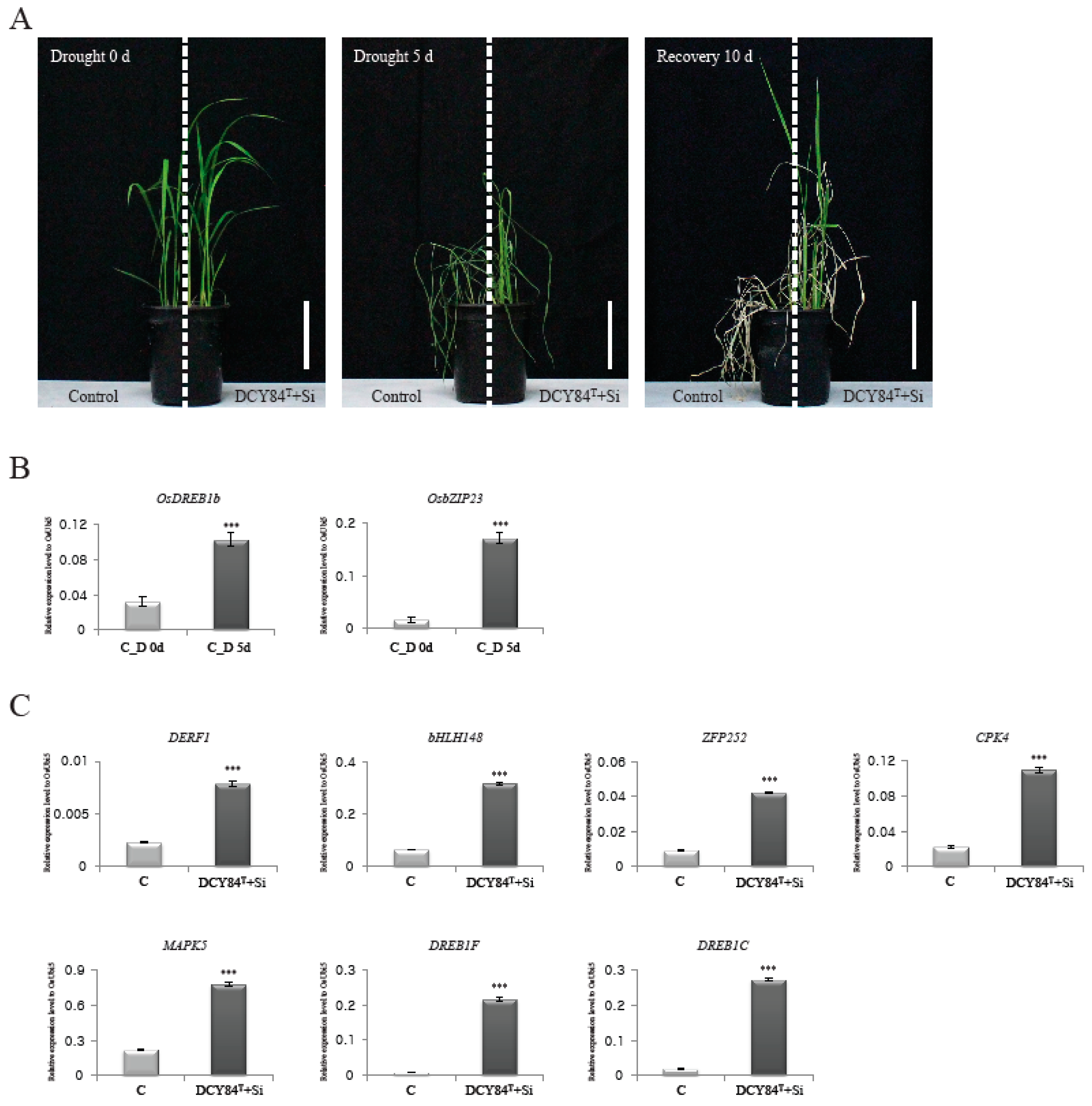
2.6. Treatment with DCY84T+Si Resulted in Increased Rice Drought Tolerance
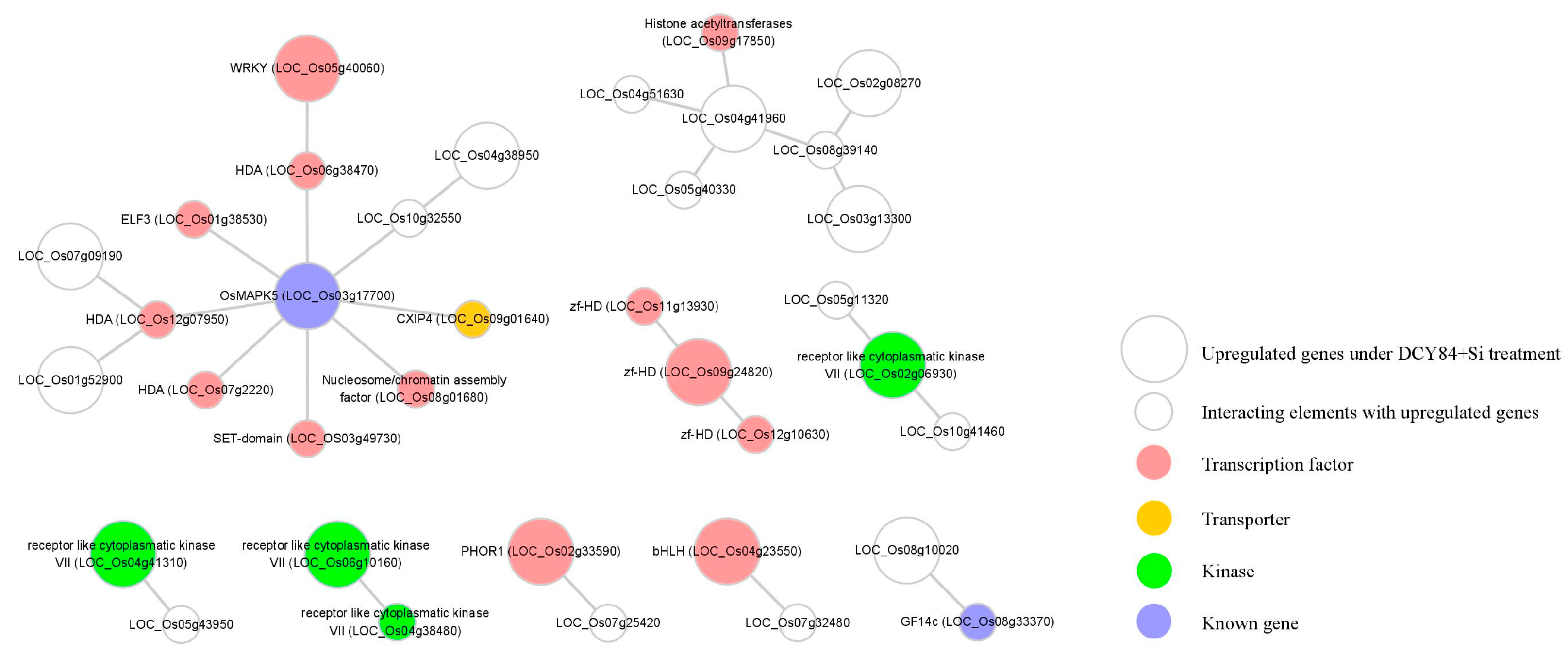
2.7. Analyses of Predicted Protein–Protein Interactions Associated with DCY84T+Si Treatment Suggest a Regulatory Model
3. Materials and Methods
3.1. Plant Materials and Phenotypic Observation
3.2. Statistical Analyses
3.3. RNA-Seq Analysis
3.4. GO Enrichment Analysis
3.5. MapMan Analysis
3.6. Analysis of Rice Genes with Known Functions
3.7. Drought Stress Treatment
3.8. RNA Extraction and Quantitative RT-PCR (qRT-PCR) Anayslis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wartiainen, I.; Eriksson, T.; Zheng, W.; Rasmussen, U. Variation in the active diazotrophic community in rice paddy-nifH PCR-DGGE analysis of rhizosphere and bulk soil. Appl. Soil Ecol. 2008, 39, 65–75. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Plant growth-promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microb. Ecol. 2009, 58, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, V.V.; Kalagudi, G.M.; Gurudatta, B.V. Towards nitrogen autotrophic rice. Curr. Sci. 2001, 81, 451–457. [Google Scholar]
- Glick, B.R. Using soil bacteria to facilitate phytoremediation. Biotechnol. Adv. 2010, 28, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 1–15. [Google Scholar] [CrossRef]
- De Souza, R.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clement, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef]
- Dimkpa, C.; Weinand, T.; Asch, F. Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef]
- Grover, M.; Ali, S.Z.; Sandhya, V.; Rasul, A.; Venkateswarlu, B. Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J. Microbiol. Biotechnol. 2011, 27, 1231–1240. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Yoon, J.H.; Kang, S.J.; Yeo, S.H.; Oh, T.K. Paenibacillus alkaliterrae sp. nov., isolated from an alkaline soil in Korea. Int. J. Syst. Evol. Microbiol. 2005, 55, 2339–2344. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Kim, H.B.; An, D.S.; Yang, H.C.; Oh, S.T.; Chung, H.J.; Yang, D.C. Paenibacillus soli sp. nov., a xylanolytic bacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2007, 57, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.M.; Fritze, D.; Blanco, A.; Spröer, C.; Tindall, B.J.; Schumann, P.; Kroppenstedt, R.M.; Diaz, P.; Pastor, F.I.J. Paenibacillus barcinonensis sp. nov., a xylanase-producing bacterium isolated from a rice field in the Ebro River delta. Int. J. Syst. Evol. Microbiol. 2005, 55, 935–939. [Google Scholar]
- Dsouza, M.; Taylor, M.W.; Ryan, J.; MacKenzie, A.; Lagutin, K.; Anderson, R.F.; Turner, S.J.; Aislabie, J. Paenibacillus darwinianus sp. nov., isolated from gamma-irradiated Antarctic soil. Int. J. Syst. Evol. Microbiol. 2014, 64, 1406–1411. [Google Scholar] [CrossRef] [PubMed]
- Sukweenadhi, J.; Kim, Y.J.; Lee, K.J.; Koh, S.C.; Hoang, V.A.; Nguyen, N.L.; Yang, D.C. Paenibacillus yonginensis sp. nov., a potential plant growth promoting bacterium isolated from humus soil of Yongin forest. Antonie van Leeuwenhoek 2014, 106, 935–945. [Google Scholar] [CrossRef]
- Sukweenadhi, J.; Kim, Y.J.; Choi, E.S.; Koh, S.C.; Lee, S.W.; Kim, Y.J.; Yang, D.C. Paenibacillus yonginensis DCY84T induces changes in Arabidopsis thaliana gene expression against aluminum, drought, and salt stress. Microbiol. Res. 2015, 172, 7–15. [Google Scholar] [CrossRef]
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef]
- Epstein, E.; Bloom, A. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2005; p. 400. [Google Scholar]
- Liang, Y.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef]
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; Datnoff, L.E. Silicon and rice disease management. Fitopatol. Bras. 2005, 30, 457–469. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Savant, N.K.; Snyder, G.H.; Datnoff, L.E. Silicon management and sustainable rice production. Adv. Agron. 1996, 58, 151–199. [Google Scholar]
- Alvarez, J.; Datnoff, L.E. The economic potential of silicon for integrated management and sustainable rice production. Crop Prot. 2001, 20, 43–48. [Google Scholar] [CrossRef]
- Choi, E.S.; Sukweenadhi, J.; Kim, Y.J.; Jung, K.H.; Koh, S.C.; Hoang, V.A.; Yang, D.C. The effects of rice seed dressing with Paenibacillus yonginensis and silicon on crop development on South Korea’s reclaimed tidal land. Field Crop. Res. 2016, 188, 121–132. [Google Scholar] [CrossRef]
- Fleck, A.T.; Nye, T.; Repenning, C.; Stahl, F.; Zahn, M.; Schenk, M.K. Silicon enhances suberization and lignification in roots of rice (Oryza sativa). J. Exp. Bot. 2011, 62, 2001–2011. [Google Scholar] [CrossRef]
- Jung, K.H.; Dardick, C.; Bartley, L.E.; Cao, P.; Phetsom, J.; Canlas, P.; Seo, Y.S.; Shultz, M.; Ouyang, S.; Yuan, Q.; et al. Refinement of light-responsive transcript lists using rice oligonucleotide arrays: Evaluation of gene-redundancy. PLoS ONE 2008, 3, e3337. [Google Scholar] [CrossRef]
- Compant, S.; Reiter, B.; Nowak, J.; Sessitsch, A.; Clément, C.; Barka, E.A. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef]
- Benhamou, N.; Kloepper, J.W.; Quadt-Hallman, A.; Tuzun, S. Induction of defense-related ultrastructural modifications in pea root tissues inoculated with endophytic bacteria. Plant Physiol. 1996, 112, 919–929. [Google Scholar] [CrossRef]
- Benhamou, N.; Kloepper, J.W.; Tuzun, S. Induction of resistance against Fusarium wilt of tomato by combination of chitosan with an endophytic bacterial strain: Ultrastructure and cytochemistry of the host response. Planta 1998, 204, 153–168. [Google Scholar] [CrossRef]
- Galili, G.; Höfgen, R. Metabolic engineering of amino acids and storage proteins in plants. Metab. Eng. 2002, 4, 3–11. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef] [PubMed]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crop. Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Wang, F.; Zhu, H.; Chen, D.; Li, Z.; Peng, R.; Yao, Q. A grape bHLH transcription factor gene, VvbHLH1, increases the accumulation of flavonoids and enhances salt and drought tolerance in transgenic Arabidopsis thaliana. Plant Cell. Tissue Organ Cult. 2016, 125, 387–398. [Google Scholar] [CrossRef]
- Jung, K.H.; An, G. Application of MapMan and RiceNet drives systematic analyses of the early heat stress transcriptome in rice seedlings. J. Plant Biol. 2012, 55, 436–449. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef]
- Glick, B.; Penrose, D.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, J.; Zhang, H.; Zhang, Z.; Quan, R.; Zhou, S.; Huang, R. Transcriptional activation of OsDERF1 in OsERF3 and OsAP2-39 negatively modulates ethylene synthesis and drought tolerance in rice. PLoS ONE 2011, 6, e25216. [Google Scholar] [CrossRef]
- Seo, J.S.; Joo, J.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; Song, S.I.; Cheong, J.J.; Lee, J.S.; Kim, J.K.; Choi, Y.D. OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J. 2011, 65, 907–921. [Google Scholar] [CrossRef]
- Qi, W.; Sun, F.; Wang, Q.; Chen, M.; Huang, Y.; Feng, Y.-Q.; Luo, X.; Yang, J. Rice ethylene-response AP2/ERF factor OsEATB restricts internode elongation by down-regulating a gibberellin biosynthetic gene. Plant Physiol. 2011, 157, 216–228. [Google Scholar] [CrossRef]
- Schmidt, R.; Mieulet, D.; Hubberten, H.-M.; Obata, T.; Hoefgen, R.; Fernie, A.R.; Fisahn, J.; San Segundo, B.; Guiderdoni, E.; Schippers, J.H.M.; et al. Salt-responsive ERF1 regulates reactive oxygen species-dependent signaling during the initial response to salt stress in rice. Plant Cell 2013, 25, 2115–2131. [Google Scholar] [CrossRef] [PubMed]
- Yokotani, N.; Sato, Y.; Tanabe, S.; Chujo, T.; Shimizu, T.; Okada, K.; Yamane, H.; Shimono, M.; Sugano, S.; Takatsuji, H.; et al. WRKY76 is a rice transcriptional repressor playing opposite roles in blast disease resistance and cold stress tolerance. J. Exp. Bot. 2013, 64, 5085–5097. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.Q.; Huang, J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Zhang, H.S. Overexpression of a TFIIIA-type zinc finger protein gene ZFP252 enhances drought and salt tolerance in rice (Oryza sativa L.). FEBS Lett. 2008, 582, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Baldrich, P.; Messeguer, J.; Lalanne, E.; Coca, M.; San Segundo, B. Overexpression of a calcium-dependent protein kinase confers salt and drought tolerance in rice by preventing membrane lipid peroxidation. Plant Physiol. 2014, 165, 688–704. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.F.; Chao, D.Y.; Shi, M.; Zhu, M.Z.; Gao, J.P.; Lin, H.X. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta 2008, 228, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Yang, Y. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid—Inducible mitogen-activated protein kinase. Plant Cell 2003, 15, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guan, Y.; Wu, Y.; Chen, H.; Chen, F.; Chu, C. Overexpression of a rice OsDREB1F gene increases salt, drought, and low temperature tolerance in both Arabidopsis and rice. Plant Mol. Biol. 2008, 67, 589–602. [Google Scholar] [CrossRef]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol. 2006, 47, 141–153. [Google Scholar] [CrossRef]
- Wang, L.; Ying, Y.; Narsai, R.; Ye, L.; Zheng, L.; Tian, J.; Whelan, J.; Shou, H. Identification of OsbHLH133 as a regulator of iron distribution between roots and shoots in Oryza sativa. Plant Cell Environ. 2013, 36, 224–236. [Google Scholar] [CrossRef]
- Yara, A.; Yaeno, T.; Hasegawa, M.; Seto, H.; Montillet, J.L.; Kusumi, K.; Seo, S.; Iba, K. Disease resistance against Magnaporthe grisea is enhanced in transgenic rice with suppression of ω-3 fatty acid desaturases. Plant Cell Physiol. 2007, 48, 1263–1274. [Google Scholar] [CrossRef]
- Seo, S.; Mitsuhara, I.; Feng, J.; Iwai, T.; Hasegawa, M.; Ohashi, Y. Cyanide, a coproduct of plant hormone ethylene biosynthesis, contributes to the resistance of rice to blast fungus. Plant Physiol. 2011, 155, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Helliwell, E.E.; Wang, Q.; Yang, Y. Transgenic rice with inducible ethylene production exhibits broad-spectrum disease resistance to the fungal pathogens Magnaporthe oryzae and Rhizoctonia solani. Plant Biotechnol. J. 2013, 11, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Dubouzet, J.G.; Maeda, S.; Sugano, S.; Ohtake, M.; Hayashi, N.; Ichikawa, T.; Kondou, Y.; Kuroda, H.; Horii, Y.; Matsui, M.; et al. Screening for resistance against Pseudomonas syringae in rice-FOX Arabidopsis lines identified a putative receptor-like cytoplasmic kinase gene that confers resistance to major bacterial and fungal pathogens in Arabidopsis and rice. Plant Biotechnol. J. 2011, 9, 466–485. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bai, X.; Wang, X.; Chu, C. OsWRKY71, a rice transcription factor, is involved in rice defense response. J. Plant Physiol. 2007, 164, 969–979. [Google Scholar] [CrossRef]
- Peng, Y.; Bartley, L.E.; Canlas, P.; Ronald, P.C. OsWRKY IIa transcription factors modulate rice innate immunity. Rice 2010, 3, 36–42. [Google Scholar] [CrossRef]
- Chujo, T.; Miyamoto, K.; Shimogawa, T.; Shimizu, T.; Otake, Y.; Yokotani, N.; Nishizawa, Y.; Shibuya, N.; Nojiri, H.; Yamane, H.; et al. OsWRKY28, a PAMP-responsive transrepressor, negatively regulates innate immune responses in rice against rice blast fungus. Plant Mol. Biol. 2013, 82, 23–37. [Google Scholar] [CrossRef]
- Koiwai, H.; Tagiri, A.; Katoh, S.; Katoh, E.; Ichikawa, H.; Minami, E.; Nishizawa, Y. RING-H2 type ubiquitin ligase EL5 is involved in root development through the maintenance of cell viability in rice. Plant J. 2007, 51, 92–104. [Google Scholar] [CrossRef]
- Sazuka, T.; Kamiya, N.; Nishimura, T.; Ohmae, K.; Sato, Y.; Imamura, K.; Nagato, Y.; Koshiba, T.; Nagamura, Y.; Ashikari, M.; et al. A rice tryptophan deficient dwarf mutant, tdd1, contains a reduced level of indole acetic acid and develops abnormal flowers and organless embryos. Plant J. 2009, 60, 227–241. [Google Scholar] [CrossRef]
- Kudo, T.; Makita, N.; Kojima, M.; Tokunaga, H.; Sakakibara, H. Cytokinin activity of cis-zeatin and phenotypic alterations induced by overexpression of putative cis-zeatin-O-glucosyltransferase in rice. Plant Physiol. 2012, 160, 319–331. [Google Scholar] [CrossRef]
- Kiribuchi, K.; Sugimori, M.; Takeda, M.; Otani, T.; Okada, K.; Onodera, H.; Ugaki, M.; Tanaka, Y.; Tomiyama-Akimoto, C.; Yamaguchi, T.; et al. RERJ1, a jasmonic acid-responsive gene from rice, encodes a basic helix-loop-helix protein. Biochem. Biophys. Res. Commun. 2004, 325, 857–863. [Google Scholar] [CrossRef]
- Sakamoto, T.; Kawabe, A.; Tokida-Segawa, A.; Shimizu, B.I.; Takatsuto, S.; Shimada, Y.; Fujioka, S.; Mizutani, M. Rice CYP734As function as multisubstrate and multifunctional enzymes in brassinosteroid catabolism. Plant J. 2011, 67, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Nakagawa, H.; Tomita, C.; Shimatani, Z.; Ohtake, M.; Nomura, T.; Jiang, C.-J.; Dubouzet, J.G.; Kikuchi, S.; Sekimoto, H.; et al. BRASSINOSTEROID UPREGULATED1, encoding a helix-loop-helix protein, is a novel gene involved in brassinosteroid signaling and controls bending of the lamina joint in rice. Plant Physiol. 2009, 151, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Piao, R.; Shi, J.; Lee, S.I.; Jiang, W.; Kim, B.K.; Lee, J.; Han, L.; Ma, W.; Koh, H.J. Fine mapping and candidate gene analysis of dense and erect panicle 3, dep3, which confers high grain yield in rice (Oryza sativa L.). Appl. Genet. 2011, 122, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Huang, H.-J.; Ren, S.-T.; Li, J.-J.; Sun, Y.; Sun, D.-Y.; Zhang, S.-Q. The rice wall-associated receptor-like kinase gene OsDEES1 plays a role in female gametophyte development. Plant Physiol. 2012, 160, 696–707. [Google Scholar] [CrossRef]
- Du, H.; Wu, N.; Cui, F.; You, L.; Li, X.; Xiong, L. A homolog of ETHYLENE OVERPRODUCER, OsETOL1, differentially modulates drought and submergence tolerance in rice. Plant J. 2014, 78, 834–849. [Google Scholar] [CrossRef]
- Yoo, Y.-H.; Nalini Chandran, A.K.; Park, J.-C.; Gho, Y.-S.; Lee, S.-W.; An, G.; Jung, K.-H. OsPhyB-mediating novel regulatory pathway for drought tolerance in rice root identified by a global RNA-Seq transcriptome analysis of rice genes in response to water deficiencies. Front. Plant Sci. 2017, 8, 580. [Google Scholar] [CrossRef]
- Ho, C.-L.; Wu, Y.; Shen, H.; Provart, N.J.; Geisler, M. A predicted protein interactome for rice. Rice 2012, 5, 15. [Google Scholar] [CrossRef]
- Nautiyal, C.S.; Srivastava, S.; Chauhan, P.S.; Seem, K.; Mishra, A.; Sopory, S.K. Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression profile of leaf and rhizosphere community in rice during salt stress. Plant Physiol. Biochem. 2013, 66, 1–9. [Google Scholar] [CrossRef]
- Singh, M.; Bhatia, P.; Sharma, P.; Khosla, B. Characterization for plant growth promoting rhizobacteria (PGPR) towards rice (Oryza sativa) seedling germination and growth. Ann. Biol. 2014, 30, 567–573. [Google Scholar]
- FastQC, version 0.11.7; Babraham Bioinformatics: Cambridge, UK. Available online: https://www.bioinformatics.babraham.ac.uk /projects/fastqc/ (accessed on 26 April 2010).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Trim Galore, version 0.5.0; Babraham Bioinformatics: Cambridge, UK. Available online: http://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 14 March 2012).
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.; Zhu, W.; Hamilton, J.; Lin, H.; Campbell, M.; Childs, K.; Thibaud-Nissen, F.; Malek, R.L.; Lee, Y.; Zheng, L.; et al. The TIGR rice genome annotation resource: Improvements and new features. Nucleic Acids Res. 2007, 35, D883–D887. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef]
- Hoang, T.V.; Vo, K.T.X.; Hong, W.J.; Jung, K.H.; Jeon, J.S. Defense response to pathogens through Epigenetic regulation in rice. J. Plant Biol. 2018, 61, 1–10. [Google Scholar] [CrossRef]
- Xuan, Y.H.; Kumar, V.; Zhu, X.F.; Je, B.I.; Kim, C.M.; Huang, J.; Cho, J.H.; Yi, G.; Han, C.-d. IDD10 is involved in the interaction between NH4+ and auxin signaling in rice roots. J. Plant Biol. 2018, 61, 72–79. [Google Scholar] [CrossRef]
- Cao, P.; Jung, K.-H.; Choi, D.; Hwang, D.; Zhu, J.; Ronald, P.C. The rice oligonucleotide array database: An atlas of rice gene expression. Rice 2012, 5, 17. [Google Scholar] [CrossRef]
- Urbanczyk-Wochniak, E.; Usadel, B.; Thimm, O.; Nunes-Nesi, A.; Carrari, F.; Davy, M.; Bläsing, O.; Kowalczyk, M.; Weicht, D.; Polinceusz, A.; et al. Conversion of MapMan to allow the analysis of transcript data from Solanaceous species: Effects of genetic and environmental alterations in energy metabolism in the leaf. Plant Mol. Biol. 2006, 60, 773–792. [Google Scholar] [CrossRef]
- Usadel, B.; Nagel, A.; Thimm, O.; Redestig, H.; Blaesing, O.E.; Palacios-Rojas, N.; Selbig, J.; Hannemann, J.; Piques, M.C.; Steinhauser, D.; et al. Extension of the visualization tool MapMan to allow statistical analysis of arrays, display of corresponding genes, and comparison with known responses. Plant Physiol. 2005, 138, 1195–1204. [Google Scholar] [CrossRef]
- Yamamoto, E.; Yonemaru, J.; Yamamoto, T.; Yano, M. OGRO: The overview of functionally characterized genes in rice online database. Rice 2012, 5, 26. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Length (cm) | Root | Leaf | Dry Weight (mg) | Root | Leaf | ||||
|---|---|---|---|---|---|---|---|---|---|
| Chilbo | Chilbo _Py + Si | Chilbo | Chilbo _Py + Si | Chilbo | Chilbo _Py + Si | Chilbo | Chilbo _Py + Si | ||
| One week | 7.16 ± 1.02 | 9.07 ± 1.34 ** | 11.99 ± 0.97 | 13.78 ± 1.31 *** | One week | 2.08 ± 0.38 | 2.56 ± 0.31 ** | 7.82 ± 0.59 | 9.5 ± 0.78 *** |
| Two weeks | 9.39 ± 1.71 | 13.04 ± 2.05 * | 26.48 ± 1.91 | 25.52 ± 1.83 | Two weeks | 7.04 ± 0.95 | 7.89 ± 2.01 | 38.34 ± 3.44 | 37.73 ± 4.78 |
| Three weeks | 15.12 ± 1.24 | 15.05 ± 1.59 | 36.52 ± 1.77 | 36.21 ± 2.84 | Three weeks | 35.50 ± 2.15 | 42.72 ± 5.12 ** | 179.23 ± 15.52 | 176.98 ± 18.67 |
| Four weeks | 16.11 ± 1.03 | 16.32 ± 1.14 | 42.39 ± 2.55 | 43.12 ± 1.93 | Four weeks | 60.56 ± 14.43 | 62.41 ± 15.87 | 255.62 ± 33.85 | 260.23 ± 37.79 |
| Major Category a | Minor Category b | Locus_Id c | Gene Name | Gene Symbol | Method d | DOI References e |
|---|---|---|---|---|---|---|
| RT f | Drought tolerance | LOC_Os08g35240 | drought-responsive ERF 1 | OsDERF1 | KD, OX | 10.1371/journal.pone.0025216 |
| RT | Drought tolerance | LOC_Os03g53020 | basic helix-loop-helix domain148 | OsbHLH148 | OX | 10.1111/j.1365-313X.2010.04477.x |
| RT | Salinity tolerance | LOC_Os09g28440 | ERF protein associated with tillering and panicle branching | OsEATB | OX | 10.1104/pp.111.179945 |
| RT | Salinity tolerance | LOC_Os05g34730 | SALT-RESPONSIVE ERF1 | SERF1 | M | 10.1105/tpc.113.113068 |
| RT | Cold tolerance | LOC_Os09g25060 | OsWRKY76 | OsWRKY76 | OX | 10.1093/jxb/ert298 |
| RT | Drought, salinity tolerance | LOC_Os12g39400 | zinc finger protein252 | ZFP252 | OX | 10.1016/j.febslet.2008.02.052 |
| RT | Drought, salinity tolerance | LOC_Os02g03410 | Ca2+-dependent protein kinase 4 | OsCPK4 | OX | 10.1104/pp.113.230268 |
| RT | Salinity, cold tolerance | LOC_Os02g44230 | trehalose-6-phosphate phosphatase1 | OsTPP1 | OX | 10.1007/s00425-008-0729-x |
| RT | Drought, salinity, cold tolerance | LOC_Os03g17700 | Mitogen-activated protein kinase5 | OsMAPK5 | KD, OX | 10.1105/tpc.008714 |
| RT | Drought, salinity, cold tolerance | LOC_Os01g73770 | dehydration-responsive element-binding transcription factor 1F | OsDREB1F | OX | 10.1007/s11103-008-9340-6 |
| RT | Drought, salinity, cold tolerance | LOC_Os06g03670 | dehydration-responsive element-binding transcription factor 1C | OsDREB1C | OX | 10.1093/pcp/pci230 |
| RT | Other soil stress tolerance | LOC_Os12g32400 | basic helix loop helix 133 | OsbHLH133 | M | 10.1111/j.1365-3040.2012.02569.x |
| RT | Blast resistance | LOC_Os03g18070 | fatty acid desaturase7 | OsFAD7 | KD | 10.1093/pcp/pcm107 |
| RT | Blast resistance | LOC_Os04g48850 | 1-aminocyclopropane-1-carboxylic acid synthase 2 | OsACS2 | KD, OX | 10.1104/pp.110.16241210.1111/pbi.12004 |
| RT | Blast resistance | LOC_Os03g17700 | Mitogen-activated protein kinase5 | OsMAPK5 | KD, OX | 10.1105/tpc.008714 |
| RT | Blast resistance | LOC_Os09g36320 | BROAD-SPECTRUM RESISTANCE 1 | BSR1 | OX | 10.1111/j.1467-7652.2010.00568.x |
| RT | Bacterial blight resistance | LOC_Os02g08440 | OsWRKY71 | OsWRKY71 | OX | 10.1016/j.jplph.2006.07.006 |
| RT | Blast, bacterial blight resistance | LOC_Os06g44010 | OsWRKY28 | OsWRKY28 | OX | 10.1007-s11103-013-0032-5 10.1007/s12284-010-9039-6 |
| RT | Blast, bacterial blight resistance | LOC_Os09g25060 | OsWRKY76 | OsWRKY76 | OX | 10.1093/jxb/ert29810.1007/s12284-010-9039-6 |
| MT g | Root | LOC_Os02g35347 | Elicitor 5 | EL5 | Others | 10.1111/j.1365-313X.2007.03120.x |
| MT | Root | LOC_Os04g38950 | tryptophan deficient dwarf 1 | tdd1 | M | 10.1111/j.1365-313X.2009.03952.x |
| MT | Root | LOC_Os04g46990 | cZ-O-glucosyltransferase 2 | cZOGT2 | OX | 10.1104/pp.112.196733 |
| MT | Dwarf | LOC_Os04g23550 | response to exogenous JA 1 | RERJ1 | KD, OX | 10.1016/j.bbrc.2004.10.126 |
| MT | Dwarf | LOC_Os06g39880 | cytochrome P450 monooxygenase 734A4 | CYP734A4 | OX | 10.1111/j.1365-313X.2011.04567.x |
| MT | Dwarf | LOC_Os02g03410 | Ca2+-dependent protein kinase 4 | OsCPK4 | KD | 10.1104/pp.113.230268 |
| MT | Leaf, Seed | LOC_Os06g12210 | BRASSINOSTEROID UPREGULATED1 | bu1 | OX | 10.1104/pp.109.140806 |
| MT | Panicle flower | LOC_Os06g46350 | dense and erect panicle 3 | dep3 | M | 10.1007/s00122-011-1543-6 |
| MT | Panicle flower | LOC_Os09g28440 | ERF protein associated with tillering and panicle branching | OsEATB | OX | 10.1104/pp.111.179945 |
| PT h | Sterility | LOC_Os09g38850 | DEFECT IN EARLY EMBRYO SAC1 | OsDEES1 | KD | 10.1104/pp.112.203943 |
| PT | Spikelet fertility | LOC_Os04g48850 | 1-amino-cyclopropane-1-carboxylate (ACC) synthase 2 | OsACS2 | KD | 10.1111/tpj.12508 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, Y.-H.; Kim, M.; Chandran, A.K.N.; Hong, W.-J.; Ahn, H.R.; Lee, G.T.; Kang, S.; Suh, D.; Kim, J.-O.; Kim, Y.-J.; et al. Genome-Wide Transcriptome Analysis of Rice Seedlings after Seed Dressing with Paenibacillus yonginensis DCY84T and Silicon. Int. J. Mol. Sci. 2019, 20, 5883. https://doi.org/10.3390/ijms20235883
Yoo Y-H, Kim M, Chandran AKN, Hong W-J, Ahn HR, Lee GT, Kang S, Suh D, Kim J-O, Kim Y-J, et al. Genome-Wide Transcriptome Analysis of Rice Seedlings after Seed Dressing with Paenibacillus yonginensis DCY84T and Silicon. International Journal of Molecular Sciences. 2019; 20(23):5883. https://doi.org/10.3390/ijms20235883
Chicago/Turabian StyleYoo, Yo-Han, Minjae Kim, Anil Kumar Nalini Chandran, Woo-Jong Hong, Hye Ryun Ahn, Gang Taik Lee, Sungju Kang, Dabin Suh, Jin-O Kim, Yeon-Ju Kim, and et al. 2019. "Genome-Wide Transcriptome Analysis of Rice Seedlings after Seed Dressing with Paenibacillus yonginensis DCY84T and Silicon" International Journal of Molecular Sciences 20, no. 23: 5883. https://doi.org/10.3390/ijms20235883
APA StyleYoo, Y.-H., Kim, M., Chandran, A. K. N., Hong, W.-J., Ahn, H. R., Lee, G. T., Kang, S., Suh, D., Kim, J.-O., Kim, Y.-J., & Jung, K.-H. (2019). Genome-Wide Transcriptome Analysis of Rice Seedlings after Seed Dressing with Paenibacillus yonginensis DCY84T and Silicon. International Journal of Molecular Sciences, 20(23), 5883. https://doi.org/10.3390/ijms20235883