Gene Therapy in Retinal Dystrophies

 ,
,
Abstract
1. Introduction
2. Inherited Retinal Dystrophies (IRDs)
3. Gene Delivery Systems
3.1. Viral Vectors
3.1.1. Adenoviruses
3.1.2. Lentiviruses (LVs)
3.1.3. Adeno-Associated Viruses (AAVs)
3.2. Nonviral Gene Transfer
4. Gene Replacement
5. Gene Silencing
5.1. Ribozymes
5.2. siRNA
6. Antisense Oligonucleotides (AONs)
7. Genome Editing: The CRISPR/Cas9 System
8. Retinal Pigment Epithelium Disorders
8.1. Leber Congenital Amaurosis (LCA)
8.2. Choroideremia (CHM)
9. Photoreceptor Disorders
9.1. Retinitis Pigmentosa (RP)
9.2. Usher Syndrome
9.3. Stargardt Maculopathy
9.4. Achromatopsia
10. Inner Retina Disorders
X-Linked Retinoschisis
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tsang, S.H.; Gouras, P. Molecular physiology and pathology of the retina. In Duane’s Clinical Opthalmology; Duane, T.D., Tasman, W., Jaeger, A.E., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1996. [Google Scholar]
- Bringmann, A.; Syrbe, S.; Gorner, K.; Kacza, J.; Francke, M.; Wiedemann, P.; Reichenbach, A. The Primate Fovea: Structure, Function and Development. Prog. Retin. Eye Res. 2018, 66, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Tsui, I.; Song, B.J.; Lin, C.S.; Tsang, S.H. A Practical Approach to Retinal Dystrophies. Adv. Exp. Med. Biol. 2018, 1085, 245–259. [Google Scholar] [PubMed]
- Cunha-Vaz, J.; Bernardes, R.; Lobo, C. Blood-Retinal Barrier. Eur. J. Ophthalmol. 2011, 21, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Grüntzig, J.; Hollmann, F. Lymphatic vessels of the eye-old questions—New insights. Ann. Anat. 2019, 221, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Benhar, I.; London, A.; Schwartz, M. The Privileged Immunity of Immune Privileged Organs: The Case of the Eye. Front. Immunol. 2012, 3, 296. [Google Scholar] [CrossRef] [PubMed]
- Slijkerman, R.W.; Song, F.; Astuti, G.D.; Huynen, M.A.; van Wijk, E.; Stieger, K.; Collin, R.W. The Pros and Cons of Vertebrate Animal Models for Functional and Therapeutic Research on Inherited Retinal Dystrophies. Prog. Retin. Eye Res. 2015, 48, 137–159. [Google Scholar] [CrossRef]
- Moore, N.A.; Morral, N.; Ciulla, T.A.; Bracha, P. Gene Therapy for Inherited Retinal and Optic Nerve Degenerations. Expert Opin. Biol. Ther. 2018, 18, 37–49. [Google Scholar] [CrossRef]
- Ochakovski, G.A.; Bartz-Schmidt, K.U.; Fischer, M.D. Retinal Gene Therapy: Surgical Vector Delivery in the Translation to Clinical Trials. Front. Neurosci. 2017, 11, 174. [Google Scholar] [CrossRef]
- DeAngelis, M.M.; Owen, L.A.; Morrison, M.A.; Morgan, D.J.; Li, M.; Shakoor, A.; Vitale, A.; Iyengar, S.; Stambolian, D.; Kim, I.K. Genetics of Age-Related Macular Degeneration (AMD). Hum. Mol. Genet 2017, 26, R45–R50. [Google Scholar] [CrossRef]
- Thompson, D.A.; Ali, R.R.; Banin, E.; Branham, K.E.; Flannery, J.G.; Gamm, D.M.; Hauswirth, W.W.; Heckenlively, J.R.; Iannaccone, A.; Jayasundera, K.T.; et al. Advancing Therapeutic Strategies for Inherited Retinal Degeneration: Recommendations from the Monaciano Symposium. Investig. Ophthalmol. Vis. Sci. 2015, 56, 918–931. [Google Scholar] [CrossRef]
- Cursiefen, C.; Cordeiro, F.; Cunha-Vaz, J.; Wheeler-Schilling, T.; Scholl, H.P.N. EVI Steering Board. Unmet Needs in Ophthalmology: A European Vision Institute-Consensus Roadmap 2019–2025. Ophthalmic Res. 2019, 62, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, F.S.; Gallenga, C.E.; Bonifazzi, C.; Perri, P. A Challenge to the Striking Genotypic Heterogeneity of Retinitis Pigmentosa: A Better Understanding of the Pathophysiology using the Newest Genetic Strategies. Eye 2016, 30, 1542–1548. [Google Scholar] [CrossRef] [PubMed]
- Broadgate, S.; Yu, J.; Downes, S.M.; Halford, S. Unravelling the Genetics of Inherited Retinal Dystrophies: Past, Present and Future. Prog. Retin. Eye Res. 2017, 59, 53–96. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, J.W.; Tan, M.H.; Ali, R.R. Gene Therapy Progress and Prospects: The Eye. Gene Ther. 2006, 13, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Chaum, E.; Hatton, M.P. Gene Therapy for Genetic and Acquired Retinal Diseases. Surv. Ophthalmol. 2002, 47, 449–469. [Google Scholar] [CrossRef]
- Zeng, Y.; Takada, Y.; Kjellstrom, S.; Hiriyanna, K.; Tanikawa, A.; Wawrousek, E.; Smaoui, N.; Caruso, R.; Bush, R.A.; Sieving, P.A. RS-1 Gene Delivery to an Adult Rs1h Knockout Mouse Model Restores ERG b-Wave with Reversal of the Electronegative Waveform of X-Linked Retinoschisis. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3279–3285. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Conley, S.M.; Naash, M.I. Gene Therapy for Stargardt Disease Associated with ABCA4 Gene. Adv. Exp. Med. Biol. 2014, 801, 719–724. [Google Scholar]
- Cashman, S.M.; Gracias, J.; Adhi, M.; Kumar-Singh, R. Adenovirus-Mediated Delivery of Factor H Attenuates Complement C3 Induced Pathology in the Murine Retina: A Potential Gene Therapy for Age-Related Macular Degeneration. J. Gene Med. 2015, 17, 229–243. [Google Scholar] [CrossRef]
- Matet, A.; Kostic, C.; Bemelmans, A.P.; Moulin, A.; Rosolen, S.G.; Martin, S.; Mavilio, F.; Amirjanians, V.; Stieger, K.; Lorenz, B.; et al. Evaluation of Tolerance to Lentiviral LV-RPE65 Gene Therapy Vector After Subretinal Delivery in Non-Human Primates. Transl. Res. 2017, 188, 40–57. [Google Scholar] [CrossRef]
- Li, T.; Adamian, M.; Roof, D.J.; Berson, E.L.; Dryja, T.P.; Roessler, B.J.; Davidson, B.L. In Vivo Transfer of a Reporter Gene to the Retina Mediated by an Adenoviral Vector. Investig. Ophthalmol. Vis. Sci. 1994, 35, 2543–2549. [Google Scholar]
- Vollrath, D.; Feng, W.; Duncan, J.L.; Yasumura, D.; D’Cruz, P.M.; Chappelow, A.; Matthes, M.T.; Kay, M.A.; LaVail, M.M. Correction of the Retinal Dystrophy Phenotype of the RCS Rat by Viral Gene Transfer of Mertk. Proc. Natl. Acad. Sci. USA 2001, 98, 12584–12589. [Google Scholar] [CrossRef] [PubMed]
- Kafri, T.; Morgan, D.; Krahl, T.; Sarvetnick, N.; Sherman, L.; Verma, I. Cellular Immune Response to Adenoviral Vector Infected Cells does Not Require De Novo Viral Gene Expression: Implications for Gene Therapy. Proc. Natl. Acad. Sci. USA 1998, 95, 11377–11382. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, W.K.; Zhou, H.; Morral, N.; Aguilar-Cordova, E.; Pestaner, J.; Langston, C.; Mull, B.; Wang, Y.; Beaudet, A.L.; Lee, B. Toxicological Comparison of E2a-Deleted and First-Generation Adenoviral Vectors Expressing alpha1-Antitrypsin After Systemic Delivery. Hum. Gene Ther. 1998, 9, 1587–1598. [Google Scholar] [CrossRef] [PubMed]
- Verma, I.M.; Weitzman, M.D. Gene Therapy: Twenty-First Century Medicine. Annu. Rev. Biochem. 2005, 74, 711–738. [Google Scholar] [CrossRef] [PubMed]
- Graham, F.L.; Prevec, L. Adenovirus-Based Expression Vectors and Recombinant Vaccines. Biotechnology 1992, 20, 363–390. [Google Scholar] [PubMed]
- Prevec, L.; Christie, B.S.; Laurie, K.E.; Bailey, M.M.; Graham, F.L.; Rosenthal, K.L. Immune Response to HIV-1 Gag Antigens Induced by Recombinant Adenovirus Vectors in Mice and Rhesus Macaque Monkeys. J. Acquir. Immune Defic. Syndr. 1991, 4, 568–576. [Google Scholar]
- Amalfitano, A.; Begy, C.R.; Chamberlain, J.S. Improved Adenovirus Packaging Cell Lines to Support the Growth of Replication-Defective Gene-Delivery Vectors. Proc. Natl. Acad. Sci. USA 1996, 93, 3352–3356. [Google Scholar] [CrossRef]
- Wang, Q.; Finer, M.H. Second-Generation Adenovirus Vectors. Nat. Med. 1996, 2, 714–716. [Google Scholar] [CrossRef]
- Kochanek, S.; Schiedner, G.; Volpers, C. High-Capacity ‘Gutless’ Adenoviral Vectors. Curr. Opin. Mol. Ther. 2001, 3, 454–463. [Google Scholar]
- O’Neal, W.K.; Zhou, H.; Morral, N.; Langston, C.; Parks, R.J.; Graham, F.L.; Kochanek, S.; Beaudet, A.L. Toxicity Associated with Repeated Administration of First-Generation Adenovirus Vectors does Not Occur with a Helper-Dependent Vector. Mol. Med. 2000, 6, 179–195. [Google Scholar] [CrossRef]
- Dudley, R.W.; Lu, Y.; Gilbert, R.; Matecki, S.; Nalbantoglu, J.; Petrof, B.J.; Karpati, G. Sustained Improvement of Muscle Function One Year After Full-Length Dystrophin Gene Transfer into Mdx Mice by a Gutted Helper-Dependent Adenoviral Vector. Hum. Gene Ther. 2004, 15, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Merched, A.; Nour, E.; Dieker, C.; Oka, K.; Chan, L. Low-Density Lipoprotein Receptor Gene Therapy using Helper-Dependent Adenovirus Produces Long-Term Protection Against Atherosclerosis in a Mouse Model of Familial Hypercholesterolemia. Gene Ther. 2004, 11, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Graf, S.; Massey, P.G.; Dichek, D.A. Improved Vascular Gene Transfer with a Helper-Dependent Adenoviral Vector. Circulation 2004, 110, 1484–1491. [Google Scholar] [CrossRef] [PubMed]
- Steinwaerder, D.S.; Carlson, C.A.; Lieber, A. Generation of Adenovirus Vectors Devoid of all Viral Genes by Recombination between Inverted Repeats. J. Virol. 1999, 73, 9303–9313. [Google Scholar] [PubMed]
- Greenberg, K.P.; Lee, E.S.; Schaffer, D.V.; Flannery, J.G. Gene Delivery to the Retina using Lentiviral Vectors. Adv. Exp. Med. Biol. 2006, 572, 255–266. [Google Scholar] [PubMed]
- Lu, J.; Sista, P.; Giguel, F.; Greenberg, M.; Kuritzkes, D.R. Relative Replicative Fitness of Human Immunodeficiency Virus Type 1 Mutants Resistant to Enfuvirtide (T-20). J. Virol. 2004, 78, 4628–4637. [Google Scholar] [CrossRef] [PubMed]
- Saenz, D.T.; Loewen, N.; Peretz, M.; Whitwam, T.; Barraza, R.; Howell, K.G.; Holmes, J.M.; Good, M.; Poeschla, E.M. Unintegrated Lentivirus DNA Persistence and Accessibility to Expression in Nondividing Cells: Analysis with Class I Integrase Mutants. J. Virol. 2004, 78, 2906–2920. [Google Scholar] [CrossRef]
- Vargas, J., Jr.; Gusella, G.L.; Najfeld, V.; Klotman, M.E.; Cara, A. Novel Integrase-Defective Lentiviral Episomal Vectors for Gene Transfer. Hum. Gene Ther. 2004, 15, 361–372. [Google Scholar] [CrossRef]
- Yanez-Munoz, R.J.; Balaggan, K.S.; MacNeil, A.; Howe, S.J.; Schmidt, M.; Smith, A.J.; Buch, P.; MacLaren, R.E.; Anderson, P.N.; Barker, S.E.; et al. Effective Gene Therapy with Nonintegrating Lentiviral Vectors. Nat. Med. 2006, 12, 348–353. [Google Scholar] [CrossRef]
- Cashman, S.M.; Sadowski, S.L.; Morris, D.J.; Frederick, J.; Kumar-Singh, R. Intercellular Trafficking of Adenovirus-Delivered HSV VP22 from the Retinal Pigment Epithelium to the Photoreceptors—Implications for Gene Therapy. Mol. Ther. 2002, 6, 813–823. [Google Scholar] [CrossRef]
- Van Adel, B.A.; Kostic, C.; Deglon, N.; Ball, A.K.; Arsenijevic, Y. Delivery of Ciliary Neurotrophic Factor Via Lentiviral-Mediated Transfer Protects Axotomized Retinal Ganglion Cells for an Extended Period of Time. Hum. Gene Ther. 2003, 14, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Bennicelli, J.; Wright, J.F.; Komaromy, A.; Jacobs, J.B.; Hauck, B.; Zelenaia, O.; Mingozzi, F.; Hui, D.; Chung, D.; Rex, T.S.; et al. Reversal of Blindness in Animal Models of Leber Congenital Amaurosis using Optimized AAV2-Mediated Gene Transfer. Mol. Ther. 2008, 16, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, W.; Dai, X.; Kong, F.; Zheng, Q.; Zhou, X.; Lü, F.; Chang, B.; Rohrer, B.; Hauswirth, W.W.; et al. Gene therapy rescues cone structure and function in the 3-month-old rd12 mouse: a model for midcourse RPE65 leber congenital amaurosis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.J.; Chang, B.; Kumar, A.; Nusinowitz, S.; Noorwez, S.M.; Li, J.; Rani, A.; Foster, T.C.; Chiodo, V.A.; Doyle, T.; et al. Gene therapy restores vision-dependent behavior as well as retinal structure and function in a mouse model of RPE65 Leber congenital amaurosis. Mol. Ther. 2006, 13, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Albright, C. Discovery of EDIT-101 for the Treatment of Leber’s Congenital Amaurosis Type 10. Available online: http://www.editasmedicine.com/data/documents/keystone_-_albright__jan_2018_1518202116.pdf (accessed on 25 September 2019).
- Maeder, M.L.; Stefanidakis, M.; Wilson, C.J.; Baral, R.; Barrera, L.A.; Bounoutas, G.S.; Bumcrot, D.; Chao, H.; Ciulla, D.M.; DaSilva, J.A.; et al. Development of a Gene-Editing Approach to Restore Vision Loss in Leber Congenital Amaurosis Type 10. Nat. Med. 2019, 25, 229–233. [Google Scholar] [CrossRef]
- Dulla, K.; Aguila, M.; Lane, A.; Jovanovic, K.; Parfitt, D.A.; Schulkens, I.; Chan, H.L.; Schmidt, I.; Beumer, W.; Vorthoren, L.; et al. Splice-Modulating Oligonucleotide QR-110 Restores CEP290 mRNA and Function in Human c. 2991 + 1655A > G LCA10 Models. Mol. Ther. Nucleic Acids 2018, 12, 730–740. [Google Scholar] [CrossRef]
- Vasireddy, V.; Mills, J.A.; Gaddameedi, R.; Basner-Tschakarjan, E.; Kohnke, M.; Black, A.D.; Alexandrov, K.; Zhou, S.; Maguire, A.M.; Chung, D.C.; et al. AAV-Mediated Gene Therapy for Choroideremia: Preclinical Studies in Personalized Models. PLoS ONE 2013, 8, e61396. [Google Scholar] [CrossRef]
- Moosajee, M.; Tracey-White, D.; Smart, M.; Weetall, M.; Torriano, S.; Kalatzis, V.; da Cruz, L.; Coffey, P.; Webster, A.R.; Welch, E. Functional Rescue of REP1 Following Treatment with PTC124 and Novel Derivative PTC-414 in Human Choroideremia Fibroblasts and the Nonsense-Mediated Zebrafish Model. Hum. Mol. Genet. 2016, 25, 3416–3431. [Google Scholar] [CrossRef]
- Moosajee, M.; Tulloch, M.; Baron, R.A.; Gregory-Evans, C.Y.; Pereira-Leal, J.B.; Seabra, M.C. Single Choroideremia Gene in Nonmammalian Vertebrates Explains Early Embryonic Lethality of the Zebrafish Model of Choroideremia. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3009–3016. [Google Scholar] [CrossRef]
- Bitoque, D.B.; Silva, G.A. Molecular Biology Tools for the Study and Therapy of PDE6beta Mutations. J. Biotechnol. 2018, 284, 1–5. [Google Scholar] [CrossRef]
- Beltran, W.A.; Cideciyan, A.V.; Lewin, A.S.; Iwabe, S.; Khanna, H.; Sumaroka, A.; Chiodo, V.A.; Fajardo, D.S.; Roman, A.J.; Deng, W.T.; et al. Gene Therapy Rescues Photoreceptor Blindness in Dogs and Paves the Way for Treating Human X-Linked Retinitis Pigmentosa. Proc. Natl. Acad. Sci. USA 2012, 109, 2132–2137. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.J.; Schlichtenbrede, F.C.; Tschernutter, M.; Bainbridge, J.W.; Thrasher, A.J.; Ali, R.R. AAV-Mediated Gene Transfer Slows Photoreceptor Loss in the RCS Rat Model of Retinitis Pigmentosa. Mol. Ther. 2003, 8, 188–195. [Google Scholar] [CrossRef]
- Deng, W.T.; Dinculescu, A.; Li, Q.; Boye, S.L.; Li, J.; Gorbatyuk, M.S.; Pang, J.; Chiodo, V.A.; Matthes, M.T.; Yasumura, D.; et al. Tyrosine-Mutant AAV8 Delivery of Human MERTK Provides Long-Term Retinal Preservation in RCS Rats. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Zallocchi, M.; Binley, K.; Lad, Y.; Ellis, S.; Widdowson, P.; Iqball, S.; Scripps, V.; Kelleher, M.; Loader, J.; Miskin, J.; et al. EIAV-Based Retinal Gene Therapy in the shaker1 Mouse Model for Usher Syndrome Type 1B: Development of UshStat. PLoS ONE 2014, 9, e94272. [Google Scholar] [CrossRef]
- Kong, J.; Kim, S.R.; Binley, K.; Pata, I.; Doi, K.; Mannik, J.; Zernant-Rajang, J.; Kan, O.; Iqball, S.; Naylor, S.; et al. Correction of the Disease Phenotype in the Mouse Model of Stargardt Disease by Lentiviral Gene Therapy. Gene Ther. 2008, 15, 1311–1320. [Google Scholar] [CrossRef]
- Hassall, M.M.; Barnard, A.R.; MacLaren, R.E. Gene Therapy for Color Blindness. Yale J. Biol. Med. 2017, 90, 543–551. [Google Scholar]
- Bush, R.A.; Zeng, Y.; Colosi, P.; Kjellstrom, S.; Hiriyanna, S.; Vijayasarathy, C.; Santos, M.; Li, J.; Wu, Z.; Sieving, P.A. Preclinical Dose-Escalation Study of Intravitreal AAV-RS1 Gene Therapy in a Mouse Model of X-Linked Retinoschisis: Dose-Dependent Expression and Improved Retinal Structure and Function. Hum. Gene Ther. 2016, 27, 376–389. [Google Scholar] [CrossRef]
- Janssen, A.; Min, S.H.; Molday, L.L.; Tanimoto, N.; Seeliger, M.W.; Hauswirth, W.W.; Molday, R.S.; Weber, B.H. Effect of Late-Stage Therapy on Disease Progression in AAV-Mediated Rescue of Photoreceptor Cells in the Retinoschisin-Deficient Mouse. Mol. Ther. 2008, 16, 1010–1017. [Google Scholar] [CrossRef]
- Marangoni, D.; Bush, R.A.; Zeng, Y.; Wei, L.L.; Ziccardi, L.; Vijayasarathy, C.; Bartoe, J.T.; Palyada, K.; Santos, M.; Hiriyanna, S.; et al. Ocular and Systemic Safety of a Recombinant AAV8 Vector for X-Linked Retinoschisis Gene Therapy: GLP Studies in Rabbits and Rs1-KO Mice. Mol. Ther. Methods Clin. Dev. 2016, 5, 16011. [Google Scholar] [CrossRef]
- Trapani, I. Adeno-Associated Viral Vectors as a Tool for Large Gene Delivery to the Retina. Genes 2019, 10. [Google Scholar] [CrossRef]
- Surace, E.M.; Auricchio, A. Versatility of AAV Vectors for Retinal Gene Transfer. Vis. Res. 2008, 48, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Asokan, A.; Samulski, R.J. Adeno-Associated Virus Serotypes: Vector Toolkit for Human Gene Therapy. Mol. Ther. 2006, 14, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Vandenberghe, L.H.; Alvira, M.R.; Lu, Y.; Calcedo, R.; Zhou, X.; Wilson, J.M. Clades of Adeno-Associated Viruses are Widely Disseminated in Human Tissues. J. Virol. 2004, 78, 6381–6388. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.P.; Alvira, M.R.; Wang, L.; Calcedo, R.; Johnston, J.; Wilson, J.M. Novel Adeno-Associated Viruses from Rhesus Monkeys as Vectors for Human Gene Therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 11854–11859. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; Wang, L.; Takeuchi, T.; Kanda, T. Two Novel Adeno-Associated Viruses from Cynomolgus Monkey: Pseudotyping Characterization of Capsid Protein. Virology 2004, 330, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Grot, E.; Cervenka, P.; Wainer, S.; Buck, C.; Chiorini, J.A. Identification and Characterization of Novel Adeno-Associated Virus Isolates in ATCC Virus Stocks. J. Virol. 2006, 80, 5082–5085. [Google Scholar] [CrossRef]
- Ferrari, F.K.; Samulski, T.; Shenk, T.; Samulski, R.J. Second-Strand Synthesis is a Rate-Limiting Step for Efficient Transduction by Recombinant Adeno-Associated Virus Vectors. J. Virol. 1996, 70, 3227–3234. [Google Scholar]
- Fisher, K.J.; Gao, G.P.; Weitzman, M.D.; DeMatteo, R.; Burda, J.F.; Wilson, J.M. Transduction with Recombinant Adeno-Associated Virus for Gene Therapy is Limited by Leading-Strand Synthesis. J. Virol. 1996, 70, 520–532. [Google Scholar]
- Thomas, C.E.; Storm, T.A.; Huang, Z.; Kay, M.A. Rapid Uncoating of Vector Genomes is the Key to Efficient Liver Transduction with Pseudotyped Adeno-Associated Virus Vectors. J. Virol. 2004, 78, 3110–3122. [Google Scholar] [CrossRef]
- Bowles, D.E.; Rabinowitz, J.E.; Samulski, R.J. Marker Rescue of Adeno-Associated Virus (AAV) Capsid Mutants: A Novel Approach for Chimeric AAV Production. J. Virol. 2003, 77, 423–432. [Google Scholar] [CrossRef][Green Version]
- Gigout, L.; Rebollo, P.; Clement, N.; Warrington, K.H., Jr.; Muzyczka, N.; Linden, R.M.; Weber, T. Altering AAV Tropism with Mosaic Viral Capsids. Mol. Ther. 2005, 11, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Hauck, B.; Chen, L.; Xiao, W. Generation and Characterization of Chimeric Recombinant AAV Vectors. Mol. Ther. 2003, 7, 419–425. [Google Scholar] [CrossRef]
- Maheshri, N.; Koerber, J.T.; Kaspar, B.K.; Schaffer, D.V. Directed Evolution of Adeno-Associated Virus Yields Enhanced Gene Delivery Vectors. Nat. Biotechnol. 2006, 24, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Muller, O.J.; Kaul, F.; Weitzman, M.D.; Pasqualini, R.; Arap, W.; Kleinschmidt, J.A.; Trepel, M. Random Peptide Libraries Displayed on Adeno-Associated Virus to Select for Targeted Gene Therapy Vectors. Nat. Biotechnol. 2003, 21, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Muzyczka, N.; Warrington, K.H., Jr. Custom Adeno-Associated Virus Capsids: The Next Generation of Recombinant Vectors with Novel Tropism. Hum. Gene Ther. 2005, 16, 408–416. [Google Scholar] [CrossRef]
- Perabo, L.; Endell, J.; King, S.; Lux, K.; Goldnau, D.; Hallek, M.; Buning, H. Combinatorial Engineering of a Gene Therapy Vector: Directed Evolution of Adeno-Associated Virus. J. Gene Med. 2006, 8, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.E.; Bowles, D.E.; Faust, S.M.; Ledford, J.G.; Cunningham, S.E.; Samulski, R.J. Cross-Dressing the Virion: The Transcapsidation of Adeno-Associated Virus Serotypes Functionally Defines Subgroups. J. Virol. 2004, 78, 4421–4432. [Google Scholar] [CrossRef]
- Ali, R.R.; Sarra, G.M.; Stephens, C.; Alwis, M.D.; Bainbridge, J.W.; Munro, P.M.; Fauser, S.; Reichel, M.B.; Kinnon, C.; Hunt, D.M.; et al. Restoration of Photoreceptor Ultrastructure and Function in Retinal Degeneration Slow Mice by Gene Therapy. Nat. Genet. 2000, 25, 306–310. [Google Scholar] [CrossRef]
- Auricchio, A.; Behling, K.C.; Maguire, A.M.; O’Connor, E.M.; Bennett, J.; Wilson, J.M.; Tolentino, M.J. Inhibition of Retinal Neovascularization by Intraocular Viral-Mediated Delivery of Anti-Angiogenic Agents. Mol. Ther. 2002, 6, 490–494. [Google Scholar] [CrossRef]
- Bainbridge, J.W.; Mistry, A.; De Alwis, M.; Paleolog, E.; Baker, A.; Thrasher, A.J.; Ali, R.R. Inhibition of Retinal Neovascularisation by Gene Transfer of Soluble VEGF Receptor sFlt-1. Gene Ther. 2002, 9, 320–326. [Google Scholar] [CrossRef]
- Bainbridge, J.W.; Smith, A.J.; Barker, S.S.; Robbie, S.; Henderson, R.; Balaggan, K.; Viswanathan, A.; Holder, G.E.; Stockman, A.; Tyler, N.; et al. Effect of Gene Therapy on Visual Function in Leber’s Congenital Amaurosis. N. Engl. J. Med. 2008, 358, 2231–2239. [Google Scholar] [CrossRef] [PubMed]
- Broderick, C.A.; Smith, A.J.; Balaggan, K.S.; Georgiadis, A.; Buch, P.K.; Trittibach, P.C.; Barker, S.E.; Sarra, G.M.; Thrasher, A.J.; Dick, A.D.; et al. Local Administration of an Adeno-Associated Viral Vector Expressing IL-10 Reduces Monocyte Infiltration and Subsequent Photoreceptor Damage during Experimental Autoimmune Uveitis. Mol. Ther. 2005, 12, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.; Lin, K.; Foulks, G.; Ma, L.; Xiao, X.; Chen, K. Highly Efficient Ex Vivo Gene Delivery into Human Corneal Endothelial Cells by Recombinant Adeno-Associated Virus. Curr. Eye Res. 2005, 30, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.J.; Xiao, X.; Wu, J.H. Inhibition of Corneal Neovascularization with Endostatin Delivered by Adeno-Associated Viral (AAV) Vector in a Mouse Corneal Injury Model. J. Biomed. Sci. 2007, 14, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Gehlbach, P.; Yamamoto, S.; Duh, E.; Zack, D.J.; Li, Q.; Berns, K.I.; Raisler, B.J.; Hauswirth, W.W.; Campochiaro, P.A. AAV-Mediated Gene Transfer of Pigment Epithelium-Derived Factor Inhibits Choroidal Neovascularization. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1994–2000. [Google Scholar] [PubMed]
- Pawlyk, B.S.; Smith, A.J.; Buch, P.K.; Adamian, M.; Hong, D.H.; Sandberg, M.A.; Ali, R.R.; Li, T. Gene Replacement Therapy Rescues Photoreceptor Degeneration in a Murine Model of Leber Congenital Amaurosis Lacking RPGRIP. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3039–3045. [Google Scholar] [CrossRef] [PubMed]
- Raisler, B.J.; Berns, K.I.; Grant, M.B.; Beliaev, D.; Hauswirth, W.W. Adeno-Associated Virus Type-2 Expression of Pigmented Epithelium-Derived Factor Or Kringles 1–3 of Angiostatin Reduce Retinal Neovascularization. Proc. Natl. Acad. Sci. USA 2002, 99, 8909–8914. [Google Scholar] [CrossRef]
- Renwick, J.; Narang, M.A.; Coupland, S.G.; Xuan, J.Y.; Baker, A.N.; Brousseau, J.; Petrin, D.; Munger, R.; Leonard, B.C.; Hauswirth, W.W.; et al. XIAP-Mediated Neuroprotection in Retinal Ischemia. Gene Ther. 2006, 13, 339–347. [Google Scholar] [CrossRef]
- Tan, M.H.; Smith, A.J.; Pawlyk, B.; Xu, X.; Liu, X.; Bainbridge, J.B.; Basche, M.; McIntosh, J.; Tran, H.V.; Nathwani, A.; et al. Gene Therapy for Retinitis Pigmentosa and Leber Congenital Amaurosis Caused by Defects in AIPL1: Effective Rescue of Mouse Models of Partial and Complete Aipl1 Deficiency using AAV2/2 and AAV2/8 Vectors. Hum. Mol. Genet. 2009, 18, 2099–2114. [Google Scholar] [CrossRef]
- Zhou, Y.; Pernet, V.; Hauswirth, W.W.; Di Polo, A. Activation of the Extracellular Signal-Regulated Kinase 1/2 Pathway by AAV Gene Transfer Protects Retinal Ganglion Cells in Glaucoma. Mol. Ther. 2005, 12, 402–412. [Google Scholar] [CrossRef]
- Cai, X.; Conley, S.M.; Naash, M.I. RPE65: Role in the Visual Cycle, Human Retinal Disease, and Gene Therapy. Ophthalmic Genet. 2009, 30, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Conley, S.M.; Cai, X.; Naash, M.I. Nonviral Ocular Gene Therapy: Assessment and Future Directions. Curr. Opin. Mol. Ther. 2008, 10, 456–463. [Google Scholar] [PubMed]
- Prud’homme, G.J.; Glinka, Y.; Khan, A.S.; Draghia-Akli, R. Electroporation-Enhanced Nonviral Gene Transfer for the Prevention Or Treatment of Immunological, Endocrine and Neoplastic Diseases. Curr. Gene Ther. 2006, 6, 243–273. [Google Scholar] [CrossRef] [PubMed]
- Dezawa, M.; Takano, M.; Negishi, H.; Mo, X.; Oshitari, T.; Sawada, H. Gene Transfer into Retinal Ganglion Cells by in Vivo Electroporation: A New Approach. Micron 2002, 33, 1–6. [Google Scholar] [CrossRef]
- Matsuda, T.; Cepko, C.L. Electroporation and RNA Interference in the Rodent Retina in Vivo and in Vitro. Proc. Natl. Acad. Sci. USA 2004, 101, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Foroozandeh, P.; Aziz, A.A. Insight into Cellular Uptake and Intracellular Trafficking of Nanoparticles. Nanoscale Res. Lett. 2018, 13, 339. [Google Scholar] [CrossRef] [PubMed]
- Trigueros, S.; Domenech, E.B.; Toulis, V.; Marfany, G. In Vitro Gene Delivery in Retinal Pigment Epithelium Cells by Plasmid DNA-Wrapped Gold Nanoparticles. Genes 2019, 10. [Google Scholar] [CrossRef]
- Zhang, X. Gold Nanoparticles: Recent Advances in the Biomedical Applications. Cell Biochem. Biophys. 2015, 72, 771–775. [Google Scholar] [CrossRef]
- Azzam, T.; Domb, A.J. Current Developments in Gene Transfection Agents. Curr. Drug Deliv. 2004, 1, 165–193. [Google Scholar] [CrossRef]
- Glover, D.J.; Lipps, H.J.; Jans, D.A. Towards Safe, Non-Viral Therapeutic Gene Expression in Humans. Nat. Rev. Genet. 2005, 6, 299–310. [Google Scholar] [CrossRef]
- Peeters, L.; Sanders, N.N.; Braeckmans, K.; Boussery, K.; Van de Voorde, J.; De Smedt, S.C.; Demeester, J. Vitreous: A Barrier to Nonviral Ocular Gene Therapy. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3553–3561. [Google Scholar] [CrossRef] [PubMed]
- D’Cruz, P.M.; Yasumura, D.; Weir, J.; Matthes, M.T.; Abderrahim, H.; LaVail, M.M.; Vollrath, D. Mutation of the Receptor Tyrosine Kinase Gene Mertk in the Retinal Dystrophic RCS Rat. Hum. Mol. Genet. 2000, 9, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Tschernutter, M.; Schlichtenbrede, F.C.; Howe, S.; Balaggan, K.S.; Munro, P.M.; Bainbridge, J.W.; Thrasher, A.J.; Smith, A.J.; Ali, R.R. Long-Term Preservation of Retinal Function in the RCS Rat Model of Retinitis Pigmentosa Following Lentivirus-Mediated Gene Therapy. Gene Ther. 2005, 12, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Batten, M.L.; Imanishi, Y.; Tu, D.C.; Doan, T.; Zhu, L.; Pang, J.; Glushakova, L.; Moise, A.R.; Baehr, W.; Van Gelder, R.N.; et al. Pharmacological and rAAV Gene Therapy Rescue of Visual Functions in a Blind Mouse Model of Leber Congenital Amaurosis. PLoS Med. 2005, 2, e333. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.M.; Yu, M.J.; Brankov, M.; Barnett, N.L.; Zhou, X.; Redmond, T.M.; Narfstrom, K.; Rakoczy, P.E. Recombinant Adeno-Associated Virus Type 2-Mediated Gene Delivery into the Rpe65 (-/-) Knockout Mouse Eye Results in Limited Rescue. Genet. Vaccines Ther. 2004, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Koenekoop, R.K. The Gene for Stargardt Disease, ABCA4, is a Major Retinal Gene: A Mini-Review. Ophthalmic Genet. 2003, 24, 75–80. [Google Scholar] [CrossRef]
- Allocca, M.; Doria, M.; Petrillo, M.; Colella, P.; Garcia-Hoyos, M.; Gibbs, D.; Kim, S.R.; Maguire, A.; Rex, T.S.; Di Vicino, U.; et al. Serotype-Dependent Packaging of Large Genes in Adeno-Associated Viral Vectors Results in Effective Gene Delivery in Mice. J. Clin. Investig. 2008, 118, 1955–1964. [Google Scholar] [CrossRef]
- Liang, Y.; Fotiadis, D.; Maeda, T.; Maeda, A.; Modzelewska, A.; Filipek, S.; Saperstein, D.A.; Engel, A.; Palczewski, K. Rhodopsin Signaling and Organization in Heterozygote Rhodopsin Knockout Mice. J. Biol. Chem. 2004, 279, 48189–48196. [Google Scholar] [CrossRef]
- Farrar, G.J.; Kenna, P.F.; Humphries, P. On the Genetics of Retinitis Pigmentosa and on Mutation-Independent Approaches to Therapeutic Intervention. EMBO J. 2002, 21, 857–864. [Google Scholar] [CrossRef]
- Birikh, K.R.; Heaton, P.A.; Eckstein, F. The Structure, Function and Application of the Hammerhead Ribozyme. Eur. J. Biochem. 1997, 245, 1–16. [Google Scholar] [CrossRef]
- Lilley, D.M. Folding and Catalysis by the Hairpin Ribozyme. FEBS Lett. 1999, 452, 26–30. [Google Scholar] [CrossRef][Green Version]
- Drenser, K.A.; Timmers, A.M.; Hauswirth, W.W.; Lewin, A.S. Ribozyme-Targeted Destruction of RNA Associated with Autosomal-Dominant Retinitis Pigmentosa. Investig. Ophthalmol. Vis. Sci. 1998, 39, 681–689. [Google Scholar] [PubMed]
- Lewin, A.S.; Drenser, K.A.; Hauswirth, W.W.; Nishikawa, S.; Yasumura, D.; Flannery, J.G.; LaVail, M.M. Ribozyme Rescue of Photoreceptor Cells in a Transgenic Rat Model of Autosomal Dominant Retinitis Pigmentosa. Nat. Med. 1998, 4, 967–971. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, B.; Millington-Ward, S.; O’Reilly, M.; Tuohy, G.; Kiang, A.S.; Kenna, P.F.; Humphries, P.; Farrar, G.J. Ribozyme-Based Therapeutic Approaches for Autosomal Dominant Retinitis Pigmentosa. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2863–2869. [Google Scholar]
- Sullivan, J.M.; Pietras, K.M.; Shin, B.J.; Misasi, J.N. Hammerhead Ribozymes Designed to Cleave all Human Rod Opsin mRNAs which Cause Autosomal Dominant Retinitis Pigmentosa. Mol. Vis. 2002, 8, 102–113. [Google Scholar]
- Gorbatyuk, M.S.; Pang, J.J.; Thomas, J., Jr.; Hauswirth, W.W.; Lewin, A.S. Knockdown of Wild-Type Mouse Rhodopsin using an AAV Vectored Ribozyme as Part of an RNA Replacement Approach. Mol. Vis. 2005, 11, 648–656. [Google Scholar]
- Gorbatyuk, M.; Justilien, V.; Liu, J.; Hauswirth, W.W.; Lewin, A.S. Preservation of Photoreceptor Morphology and Function in P23H Rats using an Allele Independent Ribozyme. Exp. Eye Res. 2007, 84, 44–52. [Google Scholar] [CrossRef]
- O’Reilly, M.; Palfi, A.; Chadderton, N.; Millington-Ward, S.; Ader, M.; Cronin, T.; Tuohy, T.; Auricchio, A.; Hildinger, M.; Tivnan, A.; et al. RNA Interference-Mediated Suppression and Replacement of Human Rhodopsin in Vivo. Am. J. Hum. Genet. 2007, 81, 127–135. [Google Scholar] [CrossRef]
- Ketting, R.F.; Fischer, S.E.; Bernstein, E.; Sijen, T.; Hannon, G.J.; Plasterk, R.H. Dicer Functions in RNA Interference and in Synthesis of Small RNA Involved in Developmental Timing in C. Elegans. Genes Dev. 2001, 15, 2654–2659. [Google Scholar] [CrossRef]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-Nucleotide RNAs Mediate RNA Interference in Cultured Mammalian Cells. Nature 2001, 411, 494–498. [Google Scholar] [CrossRef]
- Elbashir, S.M.; Lendeckel, W.; Tuschl, T. RNA Interference is Mediated by 21- and 22-Nucleotide RNAs. Genes Dev. 2001, 15, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Tolentino, M.J.; Brucker, A.J.; Fosnot, J.; Ying, G.S.; Wu, I.H.; Malik, G.; Wan, S.; Reich, S.J. Intravitreal Injection of Vascular Endothelial Growth Factor Small Interfering RNA Inhibits Growth and Leakage in a Nonhuman Primate, Laser-Induced Model of Choroidal Neovascularization. Retina 2004, 24, 660. [Google Scholar] [CrossRef] [PubMed]
- Beverly, M.; Hartsough, K.; Machemer, L.; Pavco, P.; Lockridge, J. Liquid Chromatography Electrospray Ionization Mass Spectrometry Analysis of the Ocular Metabolites from a Short Interfering RNA Duplex. J. Chromatogr. B. 2006, 835, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Kurz, D.; Ciulla, T.A. Novel Approaches for Retinal Drug Delivery. Ophthalmol. Clin. North. Am. 2002, 15, 405–410. [Google Scholar] [CrossRef]
- Pitkanen, L.; Ruponen, M.; Nieminen, J.; Urtti, A. Vitreous is a Barrier in Nonviral Gene Transfer by Cationic Lipids and Polymers. Pharm. Res. 2003, 20, 576–583. [Google Scholar] [CrossRef]
- Peeters, L.; Sanders, N.N.; Demeester, J.; De Smedt, S.C. Challenges in Non-Viral Ocular Gene Transfer. Biochem. Soc. Trans. 2007, 35, 47–49. [Google Scholar] [CrossRef]
- Deshpande, M.C.; Davies, M.C.; Garnett, M.C.; Williams, P.M.; Armitage, D.; Bailey, L.; Vamvakaki, M.; Armes, S.P.; Stolnik, S. The Effect of Poly (Ethylene Glycol) Molecular Architecture on Cellular Interaction and Uptake of DNA Complexes. J. Control. Release 2004, 97, 143–156. [Google Scholar] [CrossRef]
- Shi, F.; Wasungu, L.; Nomden, A.; Stuart, M.C.; Polushkin, E.; Engberts, J.B.; Hoekstra, D. Interference of Poly(Ethylene Glycol)-Lipid Analogues with Cationic-Lipid-Mediated Delivery of Oligonucleotides; Role of Lipid Exchangeability and Non-Lamellar Transitions. Biochem. J. 2002, 366, 333–341. [Google Scholar] [CrossRef]
- Song, L.Y.; Ahkong, Q.F.; Rong, Q.; Wang, Z.; Ansell, S.; Hope, M.J.; Mui, B. Characterization of the Inhibitory Effect of PEG-Lipid Conjugates on the Intracellular Delivery of Plasmid and Antisense DNA Mediated by Cationic Lipid Liposomes. Biochim. Biophys. Acta 2002, 1558, 1–13. [Google Scholar] [CrossRef]
- Turchinovich, A.; Zoidl, G.; Dermietzel, R. Non-Viral siRNA Delivery into the Mouse Retina in Vivo. BMC Ophthalmol. 2010, 10, 25. [Google Scholar] [CrossRef]
- Hernan, I.; Gamundi, M.J.; Planas, E.; Borras, E.; Maseras, M.; Carballo, M. Cellular Expression and siRNA-Mediated Interference of Rhodopsin Cis-Acting Splicing Mutants Associated with Autosomal Dominant Retinitis Pigmentosa. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3723–3729. [Google Scholar] [CrossRef] [PubMed]
- Tessitore, A.; Parisi, F.; Denti, M.A.; Allocca, M.; Di Vicino, U.; Domenici, L.; Bozzoni, I.; Auricchio, A. Preferential Silencing of a Common Dominant Rhodopsin Mutation does Not Inhibit Retinal Degeneration in a Transgenic Model. Mol. Ther. 2006, 14, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Cashman, S.M.; Binkley, E.A.; Kumar-Singh, R. Towards Mutation-Independent Silencing of Genes Involved in Retinal Degeneration by RNA Interference. Gene Ther. 2005, 12, 1223–1228. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kiang, A.S.; Palfi, A.; Ader, M.; Kenna, P.F.; Millington-Ward, S.; Clark, G.; Kennan, A.; O’reilly, M.; Tam, L.C.; Aherne, A.; et al. Toward a Gene Therapy for Dominant Disease: Validation of an RNA Interference-Based Mutation-Independent Approach. Mol. Ther. 2005, 12, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Gorbatyuk, M.S.; Rossmiller, B.; Hauswirth, W.W.; Lewin, A.S. Long-Term Rescue of Retinal Structure and Function by Rhodopsin RNA Replacement with a Single Adeno-Associated Viral Vector in P23H RHO Transgenic Mice. Hum. Gene Ther. 2012, 23, 356–366. [Google Scholar] [CrossRef]
- Palfi, A.; Ader, M.; Kiang, A.S.; Millington-Ward, S.; Clark, G.; O’Reilly, M.; McMahon, H.P.; Kenna, P.F.; Humphries, P.; Farrar, G.J. RNAi-Based Suppression and Replacement of Rds-Peripherin in Retinal Organotypic Culture. Hum. Mutat. 2006, 27, 260–268. [Google Scholar] [CrossRef]
- Petrs-Silva, H.; Yasumura, D.; Matthes, M.T.; LaVail, M.M.; Lewin, A.S.; Hauswirth, W.W. Suppression of Rds Expression by siRNA and Gene Replacement Strategies for Gene Therapy using rAAV Vector. Adv. Exp. Med. Biol. 2012, 723, 215–223. [Google Scholar]
- Rowe-Rendleman, C.L.; Durazo, S.A.; Kompella, U.B.; Rittenhouse, K.D.; Di Polo, A.; Weiner, A.L.; Grossniklaus, H.E.; Naash, M.I.; Lewin, A.S.; Horsager, A.; et al. Drug and Gene Delivery to the Back of the Eye: From Bench to Bedside. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2714–2730. [Google Scholar] [CrossRef]
- Gerard, X.; Perrault, I.; Munnich, A.; Kaplan, J.; Rozet, J.M. Intravitreal Injection of Splice-Switching Oligonucleotides to Manipulate Splicing in Retinal Cells. Mol. Ther. Nucleic Acids 2015, 4, e250. [Google Scholar] [CrossRef]
- Murray, S.F.; Jazayeri, A.; Matthes, M.T.; Yasumura, D.; Yang, H.; Peralta, R.; Watt, A.; Freier, S.; Hung, G.; Adamson, P.S.; et al. Allele-Specific Inhibition of Rhodopsin with an Antisense Oligonucleotide Slows Photoreceptor Cell Degeneration. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6362–6375. [Google Scholar] [CrossRef]
- Garanto, A.; Chung, D.C.; Duijkers, L.; Corral-Serrano, J.C.; Messchaert, M.; Xiao, R.; Bennett, J.; Vandenberghe, L.H.; Collin, R.W. In Vitro and in Vivo Rescue of Aberrant Splicing in CEP290-Associated LCA by Antisense Oligonucleotide Delivery. Hum. Mol. Genet. 2016, 25, 2552–2563. [Google Scholar] [PubMed]
- Ran, F.A.; Hsu, P.D.; Lin, C.Y.; Gootenberg, J.S.; Konermann, S.; Trevino, A.E.; Scott, D.A.; Inoue, A.; Matoba, S.; Zhang, Y.; et al. Double Nicking by RNA-Guided CRISPR Cas9 for Enhanced Genome Editing Specificity. Cell 2013, 154, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Komor, A.C.; Badran, A.H.; Liu, D.R. CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes. Cell 2017, 169, 559. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.E.; Hakim, C.H.; Ousterout, D.G.; Thakore, P.I.; Moreb, E.A.; Castellanos Rivera, R.M.; Madhavan, S.; Pan, X.; Ran, F.A.; Yan, W.X.; et al. In Vivo Genome Editing Improves Muscle Function in a Mouse Model of Duchenne Muscular Dystrophy. Science 2016, 351, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Xue, W.; Chen, S.; Bogorad, R.L.; Benedetti, E.; Grompe, M.; Koteliansky, V.; Sharp, P.A.; Jacks, T.; Anderson, D.G. Genome Editing with Cas9 in Adult Mice Corrects a Disease Mutation and Phenotype. Nat. Biotechnol. 2014, 32, 551–553. [Google Scholar] [CrossRef]
- Wang, H.; Yang, H.; Shivalila, C.S.; Dawlaty, M.M.; Cheng, A.W.; Zhang, F.; Jaenisch, R. One-Step Generation of Mice Carrying Mutations in Multiple Genes by CRISPR/Cas-Mediated Genome Engineering. Cell 2013, 153, 910–918. [Google Scholar] [CrossRef]
- Ousterout, D.G.; Perez-Pinera, P.; Thakore, P.I.; Kabadi, A.M.; Brown, M.T.; Qin, X.; Fedrigo, O.; Mouly, V.; Tremblay, J.P.; Gersbach, C.A. Reading Frame Correction by Targeted Genome Editing Restores Dystrophin Expression in Cells from Duchenne Muscular Dystrophy Patients. Mol. Ther. 2013, 21, 1718–1726. [Google Scholar] [CrossRef]
- Suzuki, K.; Tsunekawa, Y.; Hernandez-Benitez, R.; Wu, J.; Zhu, J.; Kim, E.J.; Hatanaka, F.; Yamamoto, M.; Araoka, T.; Li, Z.; et al. In Vivo Genome Editing Via CRISPR/Cas9 Mediated Homology-Independent Targeted Integration. Nature 2016, 540, 144–149. [Google Scholar] [CrossRef]
- Wu, W.H.; Tsai, Y.T.; Justus, S.; Lee, T.T.; Zhang, L.; Lin, C.S.; Bassuk, A.G.; Mahajan, V.B.; Tsang, S.H. CRISPR Repair Reveals Causative Mutation in a Preclinical Model of Retinitis Pigmentosa. Mol. Ther. 2016, 24, 1388–1394. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Shivalila, C.S.; Cheng, A.W.; Shi, L.; Jaenisch, R. One-Step Generation of Mice Carrying Reporter and Conditional Alleles by CRISPR/Cas-Mediated Genome Engineering. Cell 2013, 154, 1370–1379. [Google Scholar] [CrossRef]
- Wu, Y.; Liang, D.; Wang, Y.; Bai, M.; Tang, W.; Bao, S.; Yan, Z.; Li, D.; Li, J. Correction of a Genetic Disease in Mouse Via use of CRISPR-Cas9. Cell Stem Cell 2013, 13, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Cheng, T.; Yao, Y.; Li, X.; Ma, Y.; Li, L.; Zhao, H.; Bao, J.; Zhang, M.; Qiu, Z.; et al. In Vivo Genome Editing Rescues Photoreceptor Degeneration Via a Cas9/RecA-Mediated Homology-Directed Repair Pathway. Sci. Adv. 2019, 5, eaav3335. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Lin, V.G.; Guo, N.; Yang, Y. Obligate Ligation-Gated Recombination (ObLiGaRe): Custom-Designed Nuclease-Mediated Targeted Integration through Nonhomologous End Joining. Genome Res. 2013, 23, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Redmond, T.M.; Yu, S.; Lee, E.; Bok, D.; Hamasaki, D.; Chen, N.; Goletz, P.; Ma, J.X.; Crouch, R.K.; Pfeifer, K. Rpe65 is Necessary for Production of 11-Cis-Vitamin A in the Retinal Visual Cycle. Nat. Genet. 1998, 20, 344–351. [Google Scholar] [CrossRef]
- Nilsson, S.E.; Wrigstad, A.; Narfstrom, K. Changes in the DC Electroretinogram in Briard Dogs with Hereditary Congenital Night Blindness and Partial Day Blindness. Exp. Eye Res. 1992, 54, 291–296. [Google Scholar] [CrossRef]
- Pang, J.J.; Chang, B.; Hawes, N.L.; Hurd, R.E.; Davisson, M.T.; Li, J.; Noorwez, S.M.; Malhotra, R.; McDowell, J.H.; Kaushal, S.; et al. Retinal Degeneration 12 (rd12): A New, Spontaneously Arising Mouse Model for Human Leber Congenital Amaurosis (LCA). Mol. Vis. 2005, 11, 152–162. [Google Scholar]
- Rohrer, B.; Goletz, P.; Znoiko, S.; Ablonczy, Z.; Ma, J.X.; Redmond, T.M.; Crouch, R.K. Correlation of Regenerable Opsin with Rod ERG Signal in Rpe65−/− Mice during Development and Aging. Investig. Ophthalmol. Vis. Sci. 2003, 44, 310–315. [Google Scholar] [CrossRef][Green Version]
- Hauswirth, W.W.; Aleman, T.S.; Kaushal, S.; Cideciyan, A.V.; Schwartz, S.B.; Wang, L.; Conlon, T.J.; Boye, S.L.; Flotte, T.R.; Byrne, B.J.; et al. Treatment of Leber Congenital Amaurosis due to RPE65 Mutations by Ocular Subretinal Injection of Adeno-Associated Virus Gene Vector: Short-Term Results of a Phase I Trial. Hum. Gene Ther. 2008, 19, 979–990. [Google Scholar] [CrossRef]
- Maguire, A.M.; Simonelli, F.; Pierce, E.A.; Pugh, E.N., Jr.; Mingozzi, F.; Bennicelli, J.; Banfi, S.; Marshall, K.A.; Testa, F.; Surace, E.M.; et al. Safety and Efficacy of Gene Transfer for Leber’s Congenital Amaurosis. N. Engl. J. Med. 2008, 358, 2240–2248. [Google Scholar] [CrossRef]
- Patel, U.; Boucher, M.; de Leseleuc, L.; Visintini, S. Voretigene Neparvovec: An Emerging Gene Therapy for the Treatment of Inherited Blindness. In CADTH Issues in Emerging Health Technologies; CADTH: Ottawa ON, Canada, 2016; pp. 1–11. [Google Scholar]
- Testa, F.; Maguire, A.M.; Rossi, S.; Pierce, E.A.; Melillo, P.; Marshall, K.; Banfi, S.; Surace, E.M.; Sun, J.; Acerra, C.; et al. Three-Year Follow-Up After Unilateral Subretinal Delivery of Adeno-Associated Virus in Patients with Leber Congenital Amaurosis Type 2. Ophthalmology 2013, 120, 1283–1291. [Google Scholar] [CrossRef]
- Bainbridge, J.W.; Mehat, M.S.; Sundaram, V.; Robbie, S.J.; Barker, S.E.; Ripamonti, C.; Georgiadis, A.; Mowat, F.M.; Beattie, S.G.; Gardner, P.J.; et al. Long-Term Effect of Gene Therapy on Leber’s Congenital Amaurosis. N. Engl. J. Med. 2015, 372, 1887–1897. [Google Scholar] [CrossRef] [PubMed]
- Maeder, M.L.; Gersbach, C.A. Genome-Editing Technologies for Gene and Cell Therapy. Mol. Ther. 2016, 24, 430–446. [Google Scholar] [CrossRef] [PubMed]
- Cideciyan, A.V.; Jacobson, S.G.; Drack, A.V.; Ho, A.C.; Charng, J.; Garafalo, A.V.; Roman, A.J.; Sumaroka, A.; Han, I.C.; Hochstedler, M.D.; et al. Effect of an Intravitreal Antisense Oligonucleotide on Vision in Leber Congenital Amaurosis due to a Photoreceptor Cilium Defect. Nat. Med. 2019, 25, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Sankila, E.M.; Tolvanen, R.; van den Hurk, J.A.; Cremers, F.P.; de la Chapelle, A. Aberrant Splicing of the CHM Gene is a Significant Cause of Choroideremia. Nat. Genet. 1992, 1, 109–113. [Google Scholar] [CrossRef]
- Khan, K.N.; Islam, F.; Moore, A.T.; Michaelides, M. Clinical and Genetic Features of Choroideremia in Childhood. Ophthalmology 2016, 123, 2158–2165. [Google Scholar] [CrossRef]
- MacDonald, I.M.; Sereda, C.; McTaggart, K.; Mah, D. Choroideremia Gene Testing. Expert Rev. Mol. Diagn. 2004, 4, 478–484. [Google Scholar] [CrossRef]
- MacLaren, R.E.; Groppe, M.; Barnard, A.R.; Cottriall, C.L.; Tolmachova, T.; Seymour, L.; Clark, K.R.; During, M.J.; Cremers, F.P.; Black, G.C.; et al. Retinal Gene Therapy in Patients with Choroideremia: Initial Findings from a Phase 1/2 Clinical Trial. Lancet 2014, 383, 1129–1137. [Google Scholar] [CrossRef]
- Edwards, T.L.; Jolly, J.K.; Groppe, M.; Barnard, A.R.; Cottriall, C.L.; Tolmachova, T.; Black, G.C.; Webster, A.R.; Lotery, A.J.; Holder, G.E.; et al. Visual Acuity After Retinal Gene Therapy for Choroideremia. N. Engl. J. Med. 2016, 374, 1996–1998. [Google Scholar] [CrossRef]
- Simunovic, M.P.; Jolly, J.K.; Xue, K.; Edwards, T.L.; Groppe, M.; Downes, S.M.; MacLaren, R.E. The Spectrum of CHM Gene Mutations in Choroideremia and their Relationship to Clinical Phenotype. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6033–6039. [Google Scholar]
- Hamel, C. Retinitis Pigmentosa. Orphanet J. Rare Dis. 2006, 1, 40. [Google Scholar] [CrossRef]
- Mackay, D.S.; Henderson, R.H.; Sergouniotis, P.I.; Li, Z.; Moradi, P.; Holder, G.E.; Waseem, N.; Bhattacharya, S.S.; Aldahmesh, M.A.; Alkuraya, F.S.; et al. Novel Mutations in MERTK Associated with Childhood Onset Rod-Cone Dystrophy. Mol. Vis. 2010, 16, 369–377. [Google Scholar] [PubMed]
- Ksantini, M.; Lafont, E.; Bocquet, B.; Meunier, I.; Hamel, C.P. Homozygous Mutation in MERTK Causes Severe Autosomal Recessive Retinitis Pigmentosa. Eur. J. Ophthalmol. 2012, 22, 647–653. [Google Scholar] [CrossRef] [PubMed]
- LaVail, M.M.; Yasumura, D.; Matthes, M.T.; Yang, H.; Hauswirth, W.W.; Deng, W.T.; Vollrath, D. Gene Therapy for MERTK-Associated Retinal Degenerations. Adv. Exp. Med. Biol. 2016, 854, 487–493. [Google Scholar] [PubMed]
- Lentz, J.; Keats, B.J.B. Usher Syndrome Type I. In GeneReviews®; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Stephens, K., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1999. [Google Scholar]
- Sengillo, J.D.; Justus, S.; Cabral, T.; Tsang, S.H. Correction of Monogenic and Common Retinal Disorders with Gene Therapy. Genes 2017, 8. [Google Scholar] [CrossRef]
- Allikmets, R.; Singh, N.; Sun, H.; Shroyer, N.F.; Hutchinson, A.; Chidambaram, A.; Gerrard, B.; Baird, L.; Stauffer, D.; Peiffer, A.; et al. A Photoreceptor Cell-Specific ATP-Binding Transporter Gene (ABCR) is Mutated in Recessive Stargardt Macular Dystrophy. Nat. Genet. 1997, 15, 236–246. [Google Scholar] [CrossRef]
- Tanna, P.; Strauss, R.W.; Fujinami, K.; Michaelides, M. Stargardt Disease: Clinical Features, Molecular Genetics, Animal Models and Therapeutic Options. Br. J. Ophthalmol. 2017, 101, 25–30. [Google Scholar] [CrossRef]
- Davis, J.L. The Blunt End: Surgical Challenges of Gene Therapy for Inherited Retinal Diseases. Am. J. Ophthalmol. 2018, 196, xxv–xxix. [Google Scholar] [CrossRef]
- Schwartz, S.D.; Regillo, C.D.; Lam, B.L.; Eliott, D.; Rosenfeld, P.J.; Gregori, N.Z.; Hubschman, J.P.; Davis, J.L.; Heilwell, G.; Spirn, M.; et al. Human Embryonic Stem Cell-Derived Retinal Pigment Epithelium in Patients with Age-Related Macular Degeneration and Stargardt’s Macular Dystrophy: Follow-Up of Two Open-Label Phase 1/2 Studies. Lancet 2015, 385, 509–516. [Google Scholar] [CrossRef]
- Remmer, M.H.; Rastogi, N.; Ranka, M.P.; Ceisler, E.J. Achromatopsia: A Review. Curr. Opin. Ophthalmol. 2015, 26, 333–340. [Google Scholar] [CrossRef]
- Grayson, C.; Reid, S.N.; Ellis, J.A.; Rutherford, A.; Sowden, J.C.; Yates, J.R.; Farber, D.B.; Trump, D. Retinoschisin, the X-Linked Retinoschisis Protein, is a Secreted Photoreceptor Protein, and is Expressed and Released by Weri-Rb1 Cells. Hum. Mol. Genet. 2000, 9, 1873–1879. [Google Scholar] [CrossRef]
- Molday, R.S.; Kellner, U.; Weber, B.H. X-Linked Juvenile Retinoschisis: Clinical Diagnosis, Genetic Analysis, and Molecular Mechanisms. Prog. Retin. Eye Res. 2012, 31, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Min, S.H.; Molday, L.L.; Seeliger, M.W.; Dinculescu, A.; Timmers, A.M.; Janssen, A.; Tonagel, F.; Tanimoto, N.; Weber, B.H.; Molday, R.S.; et al. Prolonged Recovery of Retinal structure/function After Gene Therapy in an Rs1h-Deficient Mouse Model of x-Linked Juvenile Retinoschisis. Mol. Ther. 2005, 12, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Cukras, C.; Wiley, H.E.; Jeffrey, B.G.; Sen, H.N.; Turriff, A.; Zeng, Y.; Vijayasarathy, C.; Marangoni, D.; Ziccardi, L.; Kjellstrom, S.; et al. Retinal AAV8-RS1 Gene Therapy for X-Linked Retinoschisis: Initial Findings from a Phase I/IIa Trial by Intravitreal Delivery. Mol. Ther. 2018, 26, 2282–2294. [Google Scholar] [CrossRef] [PubMed]


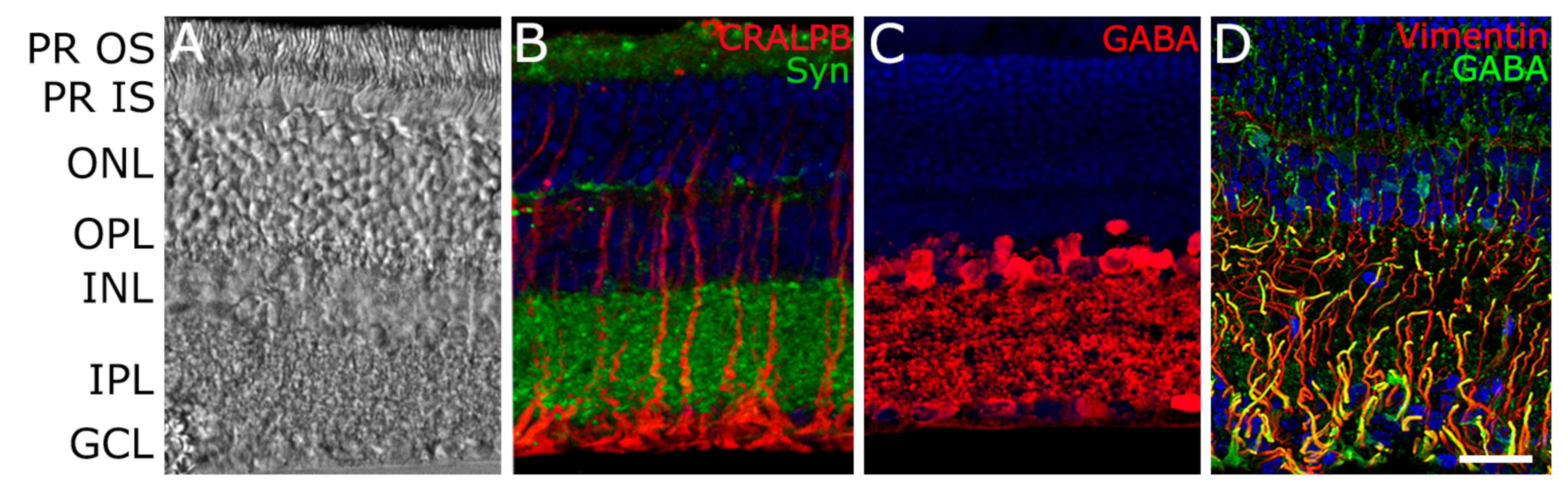{kind=link}
| Viral Mediated Therapy (Adenovirus, Adeno-Associated Virus, Lentivirus) | ||||
|---|---|---|---|---|
| Gene Replacement | Gene Silencing | Therapeutic Oligo Nucleotides | Genome editing the CRISPR-Cas9 system | |
| Integration Into Target Cell Genome | No Yes (LVV) a | No Yes (LVV) a | No Yes (LVV) a | No Yes (LVV) a |
| Lifetime Transgene Expression | Transient | - | - | Prolonged |
| Immunogenic Response | Yes | Yes | Yes | Yes |
| Mis-Insertion Risk | Yes (LVV) a | Yes (LVV) a | No | No |
| Development Costs | High | High | High | High |
| LoF c Mutation | Yes | No | Yes (AON) b | Yes |
| GoF d Mutation | No | Yes | Yes | Yes |
| Nanoparticles Mediated Therapy (Naked-DNA, Liposome-DNA, Metal-DNA) | ||||
| Gene Replacement | Gene Silencing | Therapeutic Oligonucleotides | Genome editing the CRISPR-Cas9 system | |
| Integration Into Target Cell Genome | No | No | No | No |
| Immunogenic Response | None | None | None | None |
| Lifetime Transgene Expression | Transient | - | - | Prolonged |
| Mis-Insertion Risk | No | No | No | No |
| Development Costs | Low | Low | Low | Low |
| LoF c Mutation | Yes | No | Yes (AON) b | Yes |
| GoF d Mutation | No | Yes | Yes | Yes |
| Retinal Dystrophy | Gene | Animal Model | Construct |
|---|---|---|---|
| Leber congenital amaurosis | RPE65 | rd12 mouse, Rpe65−/− mouse rd12 mouse rd12 mouse | AAV2.RPE65 [43] scAAV5-smCBA-hRPE65 [44] rAAV5-CBA-hRPE65 [45] |
| CEP290 | mice containing the human ISV26 Wild-type mouse and rabbit, non-human primates HuCEP290 knock-in mice | gRNAs and SaCas9 packaged into AAV5 vectors [46,47] QR-110 [48] Edit-101 [47] | |
| Choriodermia | CHM | normal-sighted mice chmru848 zebrafish chmru848 zebrafish | rAAV-CHM virus [49] PTC124 and PTC-414 [50] PTC124 and PTC-414 [51] |
| Retinitis pigmentosa | PDE6B | rd10 mouse | pEPito-hCMVeGFP [52] |
| RPGR | Canine models XLPRA1/2 | AAV2/5-hIRBP-hRPGR, AAV2/5-hGRK1-hRPGR [53] | |
| MERTK | Royal College of Surgeons rat Royal College of Surgeons rat | rAAV-MERTK [54] AAV8-Y733F-MERTK [55] | |
| Usher Syndrome | MYO7A | shaker1 mouse | EIAV-CMV-MYO7A [56] |
| Stargardt maculopathy | ABCA4 | Abca4−/− mouse | EIAV-CMV-ABCA4 [57] |
| Achromatopsia | CNGA3 | Cnga3–/– mouse | rAAV.CBA.CNGA3 [58] |
| X-linked retinoschisis | RS1 | Rs1-KO mouse New Zealand white rabbits, Rs1-KO mouse | AAV8-scRS/IRBPhRS [59] AAV5-mOP-RS1 [60] AAV8-scRS/IRBPhRS [61] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziccardi, L.; Cordeddu, V.; Gaddini, L.; Matteucci, A.; Parravano, M.; Malchiodi-Albedi, F.; Varano, M. Gene Therapy in Retinal Dystrophies. Int. J. Mol. Sci. 2019, 20, 5722. https://doi.org/10.3390/ijms20225722
Ziccardi L, Cordeddu V, Gaddini L, Matteucci A, Parravano M, Malchiodi-Albedi F, Varano M. Gene Therapy in Retinal Dystrophies. International Journal of Molecular Sciences. 2019; 20(22):5722. https://doi.org/10.3390/ijms20225722
Chicago/Turabian StyleZiccardi, Lucia, Viviana Cordeddu, Lucia Gaddini, Andrea Matteucci, Mariacristina Parravano, Fiorella Malchiodi-Albedi, and Monica Varano. 2019. "Gene Therapy in Retinal Dystrophies" International Journal of Molecular Sciences 20, no. 22: 5722. https://doi.org/10.3390/ijms20225722
APA StyleZiccardi, L., Cordeddu, V., Gaddini, L., Matteucci, A., Parravano, M., Malchiodi-Albedi, F., & Varano, M. (2019). Gene Therapy in Retinal Dystrophies. International Journal of Molecular Sciences, 20(22), 5722. https://doi.org/10.3390/ijms20225722