Oligomerization Affects the Ability of Human Cyclase-Associated Proteins 1 and 2 to Promote Actin Severing by Cofilins

 , and
, and
Abstract
1. Introduction
2. Results
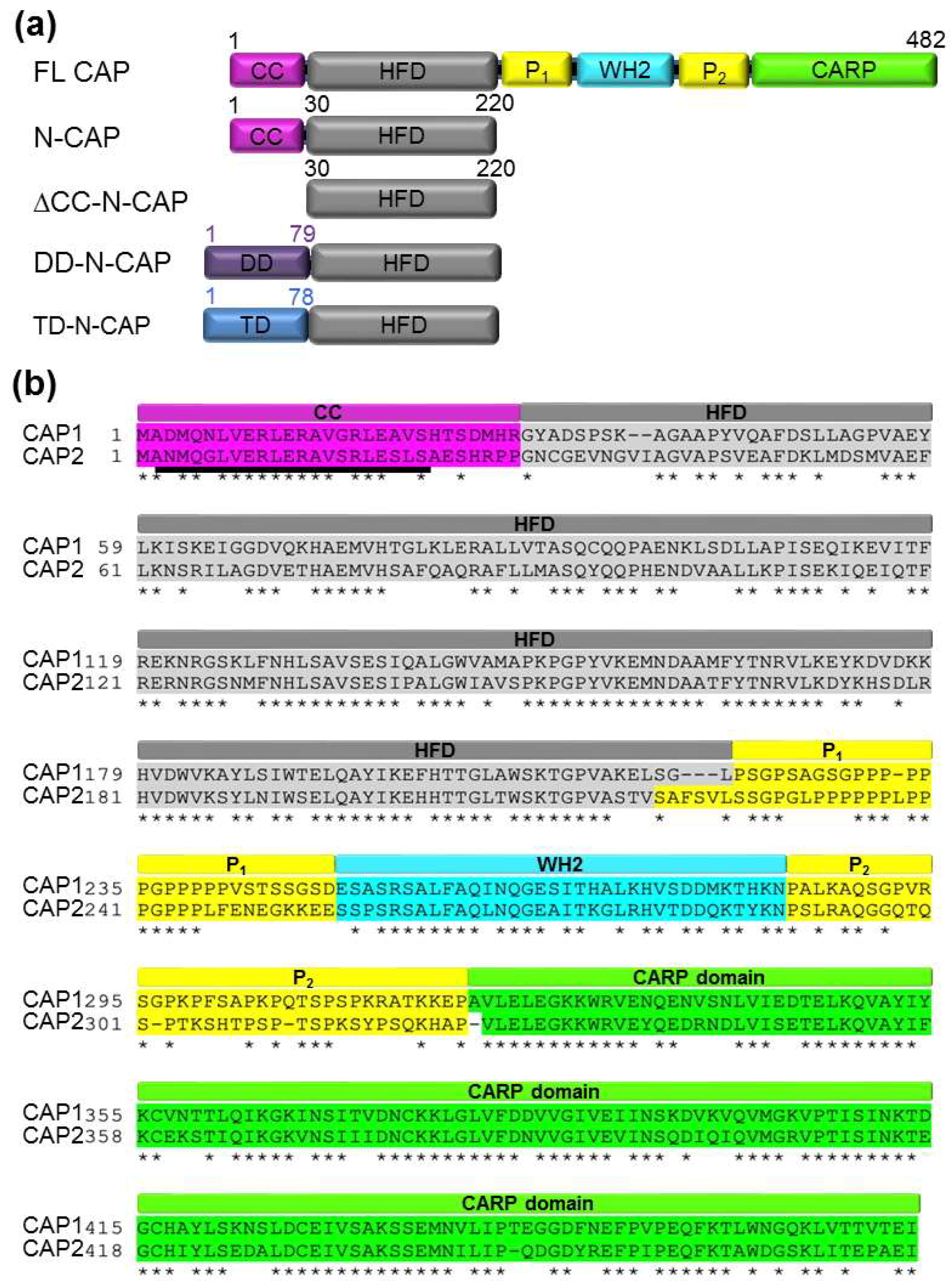
2.1. Analytical Ultracentrifugation and Native Mass Spectrometry Reveaedl that the N-terminal Domains of Recombinant Human CAPs Form Tetramers
2.2. Construction of Monomers, Dimers, and Trimers of Human CAP1 and CAP2 HFD Domains
2.3. Oligomerization through the N-terminal Coiled-Coil Region Affects the Binding Affinity of N-CAPs to F-actin
2.4. Higher Affinities of the N-CAP Constructs to F-actin Correlated with Higher Potentiation of F-actin Depolymerization by Cofilins
2.5. Interaction of CAP1 with CFL1 and CFL2 Isoforms in Cells
3. Discussion
4. Materials and Methods
4.1. Molecular Cloning
4.2. Protein Purification
4.3. Sedimentation Velocity Analytical Ultracentrifugation (SV-AUC)
4.4. F-Actin Binding Cosedimentation Assays
4.5. TIRF Microscopy
4.6. Bulk F-actin Disassembly Assays
4.7. Native Mass Spectrometry (Native MS)
4.8. Cell Culture, Western Blotting, and In Situ Proximity Ligation Assay (PLA)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADF | Actin depolymerizing factor |
| CFL | Cofilin |
| Srv2/CAP | Cyclase-associated protein |
| CC | Coiled-coil |
| HFD | Helical folded domain |
| P1 and P2 | Proline-rich regions 1 and 2 |
| WH2 | Wiskott–Aldrich homology 2 |
| CARP | C-terminal β-sheet domain |
| Aip1 | Actin interacting protein 1 |
| ATP | Adenosine triphosphate |
| ADP | Adenosine diphosphate |
| Pi | Inorganic phosphate |
| SV-AUC | Sedimentation velocity analytical ultracentrifugation |
| c(S) | Sedimentation coefficient distribution |
| TIRF | Total internal reflection fluorescence |
| MS | Mass spectrometry |
| PLA | Proximity ligation assay |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| BUDE | Bristol University Docking Engine |
| COMP | Cartridge oligomeric matrix protein |
| WCL | Whole-cell lysate |
| FITC | Fluorescein isothiocyanate |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| IPTG | Isopropyl-β-D-thiogalactoside |
| PMSF | Phenylmethylsulfonyl fluoride |
| DTT | Dithiothreitol |
| TRIS | Tris(hydroxymethyl)aminomethane |
| PIPES | Piperazine-N,N′-bis(2-ethanesulfonic acid) |
| EDTA | Ethylenediaminetetraacetic acid |
| MOPS | 3-(N-morpholino)propanesulfonic acid |
| EGTA | Ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid |
| PBS | Phosphate-buffered saline |
| GST | Glutathione S-transferase |
| BSA | Bovine serum albumin |
| HRP | Horseradish peroxidase |
References
- Pantaloni, D.; Carlier, M.F.; Korn, E.D. The Interaction between ATP-Actin and ADP-Actin. A Tentative Model for Actin Polymerization. J. Biol. Chem. 1985, 260, 6572–6578. [Google Scholar] [PubMed]
- Pollard, T.D. Rate Constants for the Reactions of ATP- and ADP-Actin with the Ends of Actin Filaments. J. Cell Biol. 1986, 103, 2747–2754. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Berstain, B.; Bamburg, J. Regulating Actin-Filament Dynamics in Vivo. Trends Biochem. Sci. 2000, 25, 19–23. [Google Scholar] [CrossRef]
- Pollard, T.D.; Cooper, J.A. Actin, a Central Player in Cell Shape and Movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, R. Actin Filament Nucleation and Elongation Factors–Structure–Function Relationships. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Courtemanche, N. Mechanisms of Formin-Mediated Actin Assembly and Dynamics. Biophys. Rev. 2018, 10, 1553–1569. [Google Scholar] [CrossRef] [PubMed]
- Ono, S. Mechanism of Depolymerization and Severing of Actin Filaments and Its Significance in Cytoskeletal Dynamics. Int. Rev. Cytol. 2007, 258, 1–82. [Google Scholar] [PubMed]
- Vartiainen, M.K. The Three Mouse Actin-Depolymerizing Factor/Cofilins Evolved to Fulfill Cell-Type-Specific Requirements for Actin Dynamics. Mol. Biol. Cell 2002, 13, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Kudryashov, D.S.; Grintsevich, E.E.; Rubenstein, P.A.; Reisler, E. A Nucleotide State-Sensing Region on Actin. J. Biol. Chem. 2010, 285, 25591–25601. [Google Scholar] [CrossRef] [PubMed]
- Kudryashov, D.S.; Reisler, E. ATP and ADP Actin States. Biopolymers 2013, 99, 245–256. [Google Scholar] [CrossRef] [PubMed]
- McGough, A.; Pope, B.; Chiu, W.; Weeds, A. Cofilin Changes the Twist of F-Actin: Implications for Actin Filament Dynamics and Cellular Function. J. Cell Biol. 1997, 138, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Galkin, V.E.; Orlova, A.; Kudryashov, D.S.; Solodukhin, A.; Reisler, E.; Schroder, G.F.; Egelman, E.H. Remodeling of Actin Filaments by ADF/Cofilin Proteins. Proc. Natl. Acad. Sci. USA 2011, 108, 20568–20572. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Takeda, S.; Mitsuoka, K.; Oda, T.; Kimura-Sakiyama, C.; Maéda, Y.; Narita, A. Structural Basis for Cofilin Binding and Actin Filament Disassembly. Nat. Commun. 2018, 9, 1860. [Google Scholar] [CrossRef] [PubMed]
- Muhlrad, A.; Kudryashov, D.; Michael Peyser, Y.; Bobkov, A.A.; Almo, S.C.; Reisler, E. Cofilin Induced Conformational Changes in F-Actin Expose Subdomain 2 to Proteolysis. J. Mol. Biol. 2004, 342, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Bobkov, A.A.; Muhlrad, A.; Shvetsov, A.; Benchaar, S.; Scoville, D.; Almo, S.C.; Reisler, E. Cofilin (ADF) Affects Lateral Contacts in F-Actin. J. Mol. Biol. 2004, 337, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Bobkov, A.A.; Muhlrad, A.; Kokabi, K.; Vorobiev, S.; Almo, S.C.; Reisler, E. Structural Effects of Cofilin on Longitudinal Contacts in F-Actin. J. Mol. Biol. 2002, 323, 739–750. [Google Scholar] [CrossRef]
- Kudryashov, D.S.; Galkin, V.E.; Orlova, A.; Phan, M.; Egelman, E.H.; Reisler, E. Cofilin Cross-Bridges Adjacent Actin Protomers and Replaces Part of the Longitudinal F-Actin Interface. J. Mol. Biol. 2006, 358, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Huehn, A.; Cao, W.; Elam, W.A.; Liu, X.; De La Cruz, E.M.; Sindelar, C.V. The Actin Filament Twist Changes Abruptly at Boundaries between Bare and Cofilin-Decorated Segments. J. Biol. Chem. 2018, 293, 5377–5383. [Google Scholar] [CrossRef] [PubMed]
- Carlier, M.F.; Laurent, V.; Santolini, J.; Melki, R.; Didry, D.; Xia, G.X.; Hong, Y.; Chua, N.H.; Pantaloni, D. Actin Depolymerizing Factor (ADF/Cofilin) Enhances the Rate of Filament Turnover: Implication in Actin-Based Motility. J. Cell Biol. 1997, 136, 1307–1322. [Google Scholar] [CrossRef] [PubMed]
- Ressad, F.; Didry, D.; Xia, G.; Hong, Y.; Chua, N.; Pantaloni, D.; Carlier, M. Kinetic Analysis of the Interaction of Actin-Depolymerizing Factor (ADF)/Cofilin with G- and F-Actins. J. Biol. Chem. 1998, 273, 20894–20902. [Google Scholar] [CrossRef] [PubMed]
- Ressad, F.; Didry, D.; Egile, C.; Pantaloni, D.; Carlier, M.F. Control of Actin Filament Length and Turnover by Actin Depolymerizing Factor (ADF/Cofilin) in the Presence of Capping Proteins and ARP2/3 Complex. J. Biol. Chem. 1999, 274, 20970–20976. [Google Scholar] [CrossRef] [PubMed]
- Balcer, H.I.; Goodman, A.L.; Rodal, A.A.; Smith, E.; Kugler, J.; Heuser, J.E.; Goode, B.L. Coordinated Regulation of Actin Filament Turnover by a High-Molecular-Weight Srv2/CAP Complex, Cofilin, Profilin, and Aip1. Curr. Biol. 2003, 13, 2159–2169. [Google Scholar] [CrossRef] [PubMed]
- Mattila, P.K.; Quintero-Monzon, O.; Kugler, J.; Moseley, J.B.; Almo, S.C.; Lappalainen, P.; Goode, B.L. A High-Affinity Interaction with ADP-Actin Monomers Underlies the Mechanism and in Vivo Function of Srv2/Cyclase-Associated Protein. Mol. Biol. Cell 2004, 15, 5158–5171. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Ravi, H.; Smith, E.M.; Goode, B.L. Aip1 and Cofilin Promote Rapid Turnover of Yeast Actin Patches and Cables: A Coordinated Mechanism for Severing and Capping Filaments. Mol. Biol. Cell 2006, 17, 2855–2868. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, M.; Achard, V.; Blanchoin, L.; Goode, B.L. Coronin Switches Roles in Actin Disassembly Depending on the Nucleotide State of Actin. Mol. Cell 2009, 34, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; Collins, A.; Chin, S.M.; Ydenberg, C.A.; Gelles, J.; Goode, B.L. Single-Molecule Imaging of a Three-Component Ordered Actin Disassembly Mechanism. Nat. Commun. 2015, 6, 7202. [Google Scholar] [CrossRef] [PubMed]
- Ge, P.; Durer, Z.A.O.; Kudryashov, D.; Zhou, Z.H.; Reisler, E. Cryo-EM Reveals Different Coronin Binding Modes for ADP–and ADP–BeFx Actin Filaments. Nat. Struct. Mol. Biol. 2014, 21, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Kotila, T.; Kogan, K.; Enkavi, G.; Guo, S.; Vattulainen, I.; Goode, B.L.; Lappalainen, P. Structural Basis of Actin Monomer Re-Charging by Cyclase-Associated Protein. Nat. Commun. 2018, 9, 1892. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Monzon, O.; Jonasson, E.M.; Bertling, E.; Talarico, L.; Chaudhry, F.; Sihvo, M.; Lappalainen, P.; Goode, B.L. Reconstitution and Dissection of the 600-KDa Srv2/CAP Complex. J. Biol. Chem. 2009, 284, 10923–10934. [Google Scholar] [CrossRef] [PubMed]
- Ono, S. The Role of Cyclase-Associated Protein in Regulating Actin Filament Dynamics—More than a Monomer-Sequestration Factor. J. Cell Sci. 2013, 126, 3249–3258. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, F.; Breitsprecher, D.; Little, K.; Sharov, G.; Sokolova, O.; Goode, B.L. Srv2/Cyclase-Associated Protein Forms Hexameric Shurikens That Directly Catalyze Actin Filament Severing by Cofilin. Mol. Biol. Cell 2013, 24, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.B.; Collins, A.; Goode, B.L. High-Speed Depolymerization at Actin Filament Ends Jointly Catalysed by Twinfilin and Srv2/CAP. Nat. Cell Biol. 2015, 17, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Kakurina, G.V.; Kolegova, E.S.; Kondakova, I.V. Adenylyl Cyclase-Associated Protein 1: Structure, Regulation, and Participation in Cellular Processes. Biochemistry 2018, 83, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, F.; Little, K.; Talarico, L.; Quintero-Monzon, O.; Goode, B.L. A Central Role for the WH2 Domain of Srv2/CAP in Recharging Actin Monomers to Drive Actin Turnover in Vitro and in Vivo. Cytoskeleton 2010, 67, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Witke, W. The Role of Profilin Complexes in Cell Motility and Other Cellular Processes. Trends Cell Biol. 2004, 14, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Goode, B.L.; Eck, M.J. Mechanism and Function of Formins in the Control of Actin Assembly. Annu. Rev. Biochem. 2007, 76, 593–627. [Google Scholar] [CrossRef] [PubMed]
- Pantaloni, D.; Carlier, M.F. How Profilin Promotes Actin Filament Assembly in the Presence of Thymosin Β4. Cell 1993, 75, 1007–1014. [Google Scholar] [CrossRef]
- Jansen, S.; Collins, A.; Golden, L.; Sokolova, O.; Goode, B.L. Structure and Mechanism of Mouse Cyclase-Associated Protein (CAP1) in Regulating Actin Dynamics. J. Biol. Chem. 2014, 289, 30732–30742. [Google Scholar] [CrossRef] [PubMed]
- Woolfson, D.N.; Bartlett, G.J.; Bruning, M.; Thomson, A.R. New Currency for Old Rope: From Coiled-Coil Assemblies to α-Helical Barrels. Curr. Opin. Struct. Biol. 2012, 22, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Swiston, J.; Young, D. Comparison of Human CAP and CAP2, Homologs of the Yeast Adenylyl Cyclase-Associated Proteins. J. Cell Sci. 1994, 107, 1671–1678. [Google Scholar] [PubMed]
- Peche, V.; Shekar, S.; Leichter, M.; Korte, H.; Schröder, R.; Schleicher, M.; Holak, T.A.; Clemen, C.S.; Ramanath-Y., B.; Pfitzer, G.; et al. CAP2, Cyclase-Associated Protein 2, Is a Dual Compartment Protein. Cell. Mol. Life Sci. 2007, 64, 2702–2715. [Google Scholar] [CrossRef] [PubMed]
- Peche, V.S.; Holak, T.A.; Burgute, B.D.; Kosmas, K.; Kale, S.P.; Wunderlich, F.T.; Elhamine, F.; Stehle, R.; Pfitzer, G.; Nohroudi, K.; et al. Ablation of Cyclase-Associated Protein 2 (CAP2) Leads to Cardiomyopathy. Cell. Mol. Life Sci. 2013, 70, 527–543. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Song, X.; Zhang, G.; Peng, A.; Li, X.; Li, M.; Liu, Y.; Wang, C. Overexpression of Adenylate Cyclase-Associated Protein 1 Is Associated with Metastasis of Lung Cancer. Oncol. Rep. 2013, 30, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Shen, C.; Tan, M.; Li, M.; Song, X.; Wang, C. Systematic Analysis of Gene Expression Alterations and Clinical Outcomes of Adenylate Cyclase-Associated Protein in Cancer. Oncotarget 2017, 8, 27216–27239. [Google Scholar] [PubMed]
- Hasan, R.; Zhou, G.L. The Cytoskeletal Protein Cyclase-Associated Protein 1 (CAP1) in Breast Cancer: Context-Dependent Roles in Both the Invasiveness and Proliferation of Cancer Cells and Underlying Cell Signals. Int. J. Mol. Sci. 2019, 20, 2653. [Google Scholar] [CrossRef] [PubMed]
- Hua, M.; Yan, S.; Deng, Y.; Xi, Q.; Liu, R.; Yang, S.; Liu, J.; Tang, C.; Wang, Y.; Zhing, J. CAP1 Is Overexpressed in Human Epithelial Ovarian Cancer and Promotes Cell Proliferation. Int. J. Mol. Med. 2015, 35, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Fu, L.Q.; Wang, H.J.; Wang, Y.Y. CAP2 Is a Valuable Biomarker for Diagnosis and Prognostic in Patients with Gastric Cancer. Pathol. Oncol. Res. 2018, 37, 17784–17792. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Peng, S.; Huang, Q.; Liu, Y.; Jiang, H.; Li, X.; Wang, J. Expression Status of Cyclase-Associated Protein 2 as a Prognostic Marker for Human Breast Cancer. Oncol. Rep. 2016, 36, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Masugi, Y.; Tanese, K.; Emoto, K.; Yamazaki, K.; Effendi, K.; Funakoshi, T.; Mori, M.; Sakamoto, M. Overexpression of Adenylate Cyclase-Associated Protein 2 Is a Novel Prognostic Marker in Malignant Melanoma. Pathol. Int. 2015, 65, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Boyken, S.E.; Chen, Z.; Groves, B.; Langan, R.A.; Oberdorfer, G.; Ford, A.; Gilmore, J.M.; Xu, C.; DiMaio, F.; Pereira, J.H.; et al. De Novo Design of Protein Homo-Oligomers with Modular Hydrogen-Bond Network-Mediated Specificity. Science 2016, 352, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Brautigam, C.A.; Ghirlando, R.; Schuck, P. Overview of Current Methods in Sedimentation Velocity and Sedimentation Equilibrium Analytical Ultracentrifugation. Curr. Protoc. Protein Sci. 2013, 71. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.W.; Woolfson, D.N. CCBuilder 2.0: Powerful and Accessible Coiled-Coil Modeling. Protein Sci. 2018, 27, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Paavilainen, V.O.; Oksanen, E.; Goldman, A.; Lappalainen, P. Structure of the Actin-Depolymerizing Factor Homology Domain in Complex with Actin. J. Cell Biol. 2008, 182, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Chin, S.M.; Jansen, S.; Goode, B.L. TIRF Microscopy Analysis of Human Cof1, Cof2, and ADF Effects on Actin Filament Severing and Turnover. J. Mol. Biol. 2016, 428, 1604–1616. [Google Scholar] [CrossRef] [PubMed]
- Effendi, K.; Yamazaki, K.; Mori, T.; Masugi, Y.; Makino, S.; Sakamoto, M. Involvement of Hepatocellular Carcinoma Biomarker, Cyclase-Associated Protein 2 in Zebrafish Body Development and Cancer Progression. Exp. Cell Res. 2013, 319, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Mu, A.; Fung, T.S.; Kettenbach, A.N.; Chakrabarti, R.; Higgs, H.N. A Complex Containing Lysine-Acetylated Actin Inhibits the Formin INF2. Nat. Cell Biol. 2019, 21, 592–602. [Google Scholar]
- Guo, Y.; Bozic, D.; Malashkevich, V.N.; Kammerer, R.A.; Schulthess, T.; Engel, J. All-Trans Retinol, Vitamin D and Other Hydrophobic Compounds Bind in the Axial Pore of the Five-Stranded Coiled-Coil Domain of Cartilage Oligomeric Matrix Protein. EMBO J. 1998, 17, 5265–5272. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, A.A.; Orriss, G.; Okun, N.; Meier, M.; Klonisch, T.; Khajehpour, M.; Stetefeld, J. The Pentameric Channel of COMPcc in Complex with Different Fatty Acids. PLoS ONE 2012, 7, e48130. [Google Scholar] [CrossRef] [PubMed]
- Mrazek, J.; Toso, D.; Ryazantsev, S.; Zhang, X.; Zhou, Z.H.; Fernandez, B.C.; Kickhoefer, V.A.; Rome, L.H. Polyribosomes Are Molecular 3D Nanoprinters That Orchestrate the Assembly of Vault Particles. ACS Nano 2014, 8, 11552–11559. [Google Scholar] [CrossRef] [PubMed]
- Dopie, J.; Skarp, K.P.; Rajakylä, E.K.; Tanhuanpää, K.; Vartiainen, M.K. Active Maintenance of Nuclear Actin by Importin 9 Supports Transcription. Proc. Natl. Acad. Sci. USA 2012, 109, E544–E552. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, A.; Pope, B.; Weeds, A.; Koffer, A. Latrunculin B or ATP Depletion Induces Cofilin-Dependent Translocation of Actin into Nuclei of Mast Cells. J. Biol. Chem. 2003, 278, 14394–14400. [Google Scholar] [CrossRef] [PubMed]
- Munsie, L.N.; Desmond, C.R.; Truant, R. Cofilin Nuclear-Cytoplasmic Shuttling Affects Cofilin-Actin Rod Formation during Stress. J. Cell Sci. 2012, 125, 3977–3988. [Google Scholar] [CrossRef] [PubMed]
- Minamide, L.S.; Striegl, A.M.; Boyle, J.A.; Meberg, P.J.; Bamburg, J.R. Neurodegenerative Stimuli Induce Persistent ADF/Cofilin-Actin Rods That Disrupt Distal Neurite Function. Nat. Cell Biol. 2000, 2, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Abe, H.; Nagaoka, R.; Obinata, T. Colocalization of ADF and Cofilin in Intranuclear Actin Rods of Cultured Muscle Cells. J. Muscle Res. Cell Motil. 1993, 14, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Bamburg, J.R.; Bernstein, B.W. Actin Dynamics and Cofilin-Actin Rods in Alzheimer Disease. Cytoskeleton 2016, 73, 477–497. [Google Scholar] [CrossRef] [PubMed]
- Beck, H.; Flynn, K.; Lindenberg, K.S.; Schwarz, H.; Bradke, F.; Di Giovanni, S.; Knöll, B. Serum Response Factor (SRF)-Cofilin-Actin Signaling Axis Modulates Mitochondrial Dynamics. Proc. Natl. Acad. Sci. USA 2012, 109, E2523–E2532. [Google Scholar] [CrossRef] [PubMed]
- Li, G.B.; Zhang, H.W.; Fu, R.Q.; Hu, X.Y.; Liu, L.; Li, Y.N.; Liu, Y.X.; Liu, X.; Hu, J.J.; Deng, Q.; et al. Mitochondrial Fission and Mitophagy Depend on Cofilin-Mediated Actin Depolymerization Activity at the Mitochondrial Fission Site. Oncogene 2018, 37, 1485–1502. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhou, J.; Budhraja, A.; Hu, X.; Chen, Y.; Cheng, Q.; Liu, L.; Zhou, T.; Li, P.; Liu, E.; et al. Mitochondrial Translocation and Interaction of Cofilin and Drp1 Are Required for Erucin-Induced Mitochondrial Fission and Apoptosis. Oncotarget 2015, 6, 1834–1849. [Google Scholar] [CrossRef] [PubMed]
- Chua, B.T.; Volbracht, C.; Tan, K.O.; Li, R.; Yu, V.C.; Li, P. Mitochondrial Translocation of Cofilin Is an Early Step in Apoptosis Induction. Nat. Cell Biol. 2003, 5, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Rehklau, K.; Gurniak, C.B.; Conrad, M.; Friauf, E.; Ott, M.; Rust, M.B. ADF/Cofilin Proteins Translocate to Mitochondria during Apoptosis but Are Not Generally Required for Cell Death Signaling. Cell Death Differ. 2012, 19, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.; Rust, M.B.; Culmsee, C. Actin(g) on Mitochondria—A Role for Cofilin1 in Neuronal Cell Death Pathways. Biol. Chem. 2019, 400, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Cheng, Q.; Liu, L.; Zhou, T.; Shan, C.; Hu, X.; Zhou, J.; Liu, E.; Li, P.; Gao, N. Mitochondrial Translocation of Cofilin Is Required for Allyl Isothiocyanate-Mediated Cell Death via ROCK1/PTEN/PI3K Signaling Pathway. Cell Commun. Signal. 2013, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fu, R.; Liu, Y.; Li, J.; Zhang, H.; Hu, X.; Chen, Y.; Liu, X.; Li, Y.; Li, P.; et al. Dephosphorylation and Mitochondrial Translocation of Cofilin Sensitizes Human Leukemia Cells to Cerulenin-Induced Apoptosis via the ROCK1/Akt/JNK Signaling Pathway. Oncotarget 2016, 7, 20655–20668. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Serebryannyy, L.A.; Parilla, M.; Annibale, P.; Cruz, C.M.; Laster, K.; Gratton, E.; Kudryashov, D.; Kosak, S.T.; Gottardi, C.J.; de Lanerolle, P. Persistent Nuclear Actin Filaments Inhibit Transcription by RNA Polymerase II. J. Cell Sci. 2016, 129, 3412–3425. [Google Scholar] [CrossRef] [PubMed]
- Spudich, J.A.; Watt, S. The Regulation of Rabbit Skeletal Muscle Contraction. I. Biochemical Studies of the Interaction of the Tropomyosin-Troponin Complex with Actin and the Proteolytic Fragments of Myosin. J. Biol. Chem. 1971, 246, 4866–4871. [Google Scholar] [PubMed]
- Heisler, D.B.; Kudryashova, E.; Grinevich, D.O.; Suarez, C.; Winkelman, J.D.; Birukov, K.G.; Kotha, S.R.; Parinandi, N.L.; Vavylonis, D.; Kovar, D.R.; et al. ACD Toxin–Produced Actin Oligomers Poison Formin-Controlled Actin Polymerization. Science 2015, 349, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Kudryashova, E.; Heisler, D.B.; Williams, B.; Harker, A.J.; Shafer, K.; Quinlan, M.E.; Kovar, D.R.; Vavylonis, D.; Kudryashov, D.S. Actin Cross-Linking Toxin Is a Universal Inhibitor of Tandem-Organized and Oligomeric G-Actin Binding Proteins. Curr. Biol. 2018, 28, 1536–1547. [Google Scholar] [CrossRef] [PubMed]
- Yehl, J.; Kudryashova, E.; Reisler, E.; Kudryashov, D.; Polenova, T. Structural Analysis of Human Cofilin 2/Filamentous Actin Assemblies: Atomic-Resolution Insights from Magic Angle Spinning NMR Spectroscopy. Sci. Rep. 2017, 7, 44506. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Vanaernum, Z.L.; Gilbert, J.D.; Belov, M.E.; Makarov, A.A.; Horning, S.R.; Wysocki, V.H. Surface-Induced Dissociation of Noncovalent Protein Complexes in an Extended Mass Range Orbitrap Mass Spectrometer. Anal. Chem. 2019, 91, 3611–3618. [Google Scholar] [CrossRef] [PubMed]
- VanAernum, Z.; Buch, F.; Jones, B.J.; Jia, M.; Chen, Z.; Boyken, S.E.; Sahasrabuddhe, A.; Baker, D.; Wysocki, V. Rapid Online Buffer Exchange: A Method for Screening of Proteins, Protein Complexes, and Cell Lysates by Native Mass Spectrometry. ChemRxiv 2019. [Google Scholar] [CrossRef]
- Marty, M.T.; Baldwin, A.J.; Marklund, E.G.; Hochberg, G.K.A.; Benesch, J.L.P.; Robinson, C.V. Bayesian Deconvolution of Mass and Ion Mobility Spectra: From Binary Interactions to Polydisperse Ensembles. Anal. Chem. 2015, 87, 4370–4376. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AUC (ƒ) | AUC (S) | AUC MW (kDa) | MS MW (kDa) | Theoretical MW (kDa) | |
|---|---|---|---|---|---|
| ΔCC-N-CAP1 | 1.97 | 1.36 | 25.30 | 23.25 | 23.48 (monomer) |
| DD-N-CAP1 | 2.02 | 2.53 | 60.40 | 66.03 | 66.10 (dimer) |
| TD-N-CAP1 | 1.97 | 3.33 | 92.3 | 97.60 | 97.46 (trimer) |
| N-CAP1 | 2.17 | 3.43 | 92.1 | 101.46 | 101.46 (tetramer) |
| ΔCC-N-CAP2 | 2.07 | 1.62 | 23.50 | 23.69 | 23.69 (monomer) |
| DD-N-CAP2 | 1.44 | 3.78 | 66.8 | 66.63 | 66.52 (dimer) |
| TD-N-CAP2 | 1.41 | 4.84 | 92.3 | 98.36 | 98.03 (trimer) |
| N-CAP2 | 2.17 | 3.54 | 102.00 | 102.34 | 102.34 (tetramer) |
| Oligomeric State | BUDE Energy | |
|---|---|---|
| N-terminal CC of CAP1 | Tetramer | −398.7 ± 13.7 |
| Pentamer | −671.4 ± 10.2 | |
| Hexamer | −857.1 ± 73.0 | |
| Heptamer | −916.4 ± 42.5 | |
| N-terminal CC of CAP2 | Tetramer | −388.5 ± 15.4 |
| Pentamer | −601.2 ± 15.4 | |
| Hexamer | −831.6 ± 65.1 | |
| Heptamer | −878.6 ± 23.2 |
| No CAPs | CAP1 | CAP2 | |
|---|---|---|---|
| Actin alone | 12.9 ± 2.8 | ||
| CFL1 | 44.0 ± 4.0 | ||
| CFL1 + ΔCC-N-CAP | 44 ± 4.0 | 48 ± 6.9 | |
| CFL1 + DD-N-CAP | 52 ± 4.0 | 52 ± 4.0 | |
| CFL1 + TD-N-CAP | 60 ± 12 | 64 ± 4.0 | |
| CFL1 + N-CAP | 72 ± 3.4 | 68 ± 14 | |
| CFL2 | 15.8 ± 6.5 | ||
| CFL2 + ΔCC-N-CAP | 24 ± 4.0 | 12 ± 1.4 | |
| CFL2 + DD-N-CAP | 30 ± 0.8 | 48 ± 3.5 | |
| CFL2 + TD-N-CAP | 42 ± 6.0 | 44 ± 4.0 | |
| CFL2 + N-CAP | 39 ± 4.0 | 48 ± 2.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purde, V.; Busch, F.; Kudryashova, E.; Wysocki, V.H.; Kudryashov, D.S. Oligomerization Affects the Ability of Human Cyclase-Associated Proteins 1 and 2 to Promote Actin Severing by Cofilins. Int. J. Mol. Sci. 2019, 20, 5647. https://doi.org/10.3390/ijms20225647
Purde V, Busch F, Kudryashova E, Wysocki VH, Kudryashov DS. Oligomerization Affects the Ability of Human Cyclase-Associated Proteins 1 and 2 to Promote Actin Severing by Cofilins. International Journal of Molecular Sciences. 2019; 20(22):5647. https://doi.org/10.3390/ijms20225647
Chicago/Turabian StylePurde, Vedud, Florian Busch, Elena Kudryashova, Vicki H. Wysocki, and Dmitri S. Kudryashov. 2019. "Oligomerization Affects the Ability of Human Cyclase-Associated Proteins 1 and 2 to Promote Actin Severing by Cofilins" International Journal of Molecular Sciences 20, no. 22: 5647. https://doi.org/10.3390/ijms20225647
APA StylePurde, V., Busch, F., Kudryashova, E., Wysocki, V. H., & Kudryashov, D. S. (2019). Oligomerization Affects the Ability of Human Cyclase-Associated Proteins 1 and 2 to Promote Actin Severing by Cofilins. International Journal of Molecular Sciences, 20(22), 5647. https://doi.org/10.3390/ijms20225647