Wounding and UVB Light Synergistically Induce the Biosynthesis of Phenolic Compounds and Ascorbic Acid in Red Prickly Pears (Opuntia ficus-indica cv. Rojo Vigor)

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
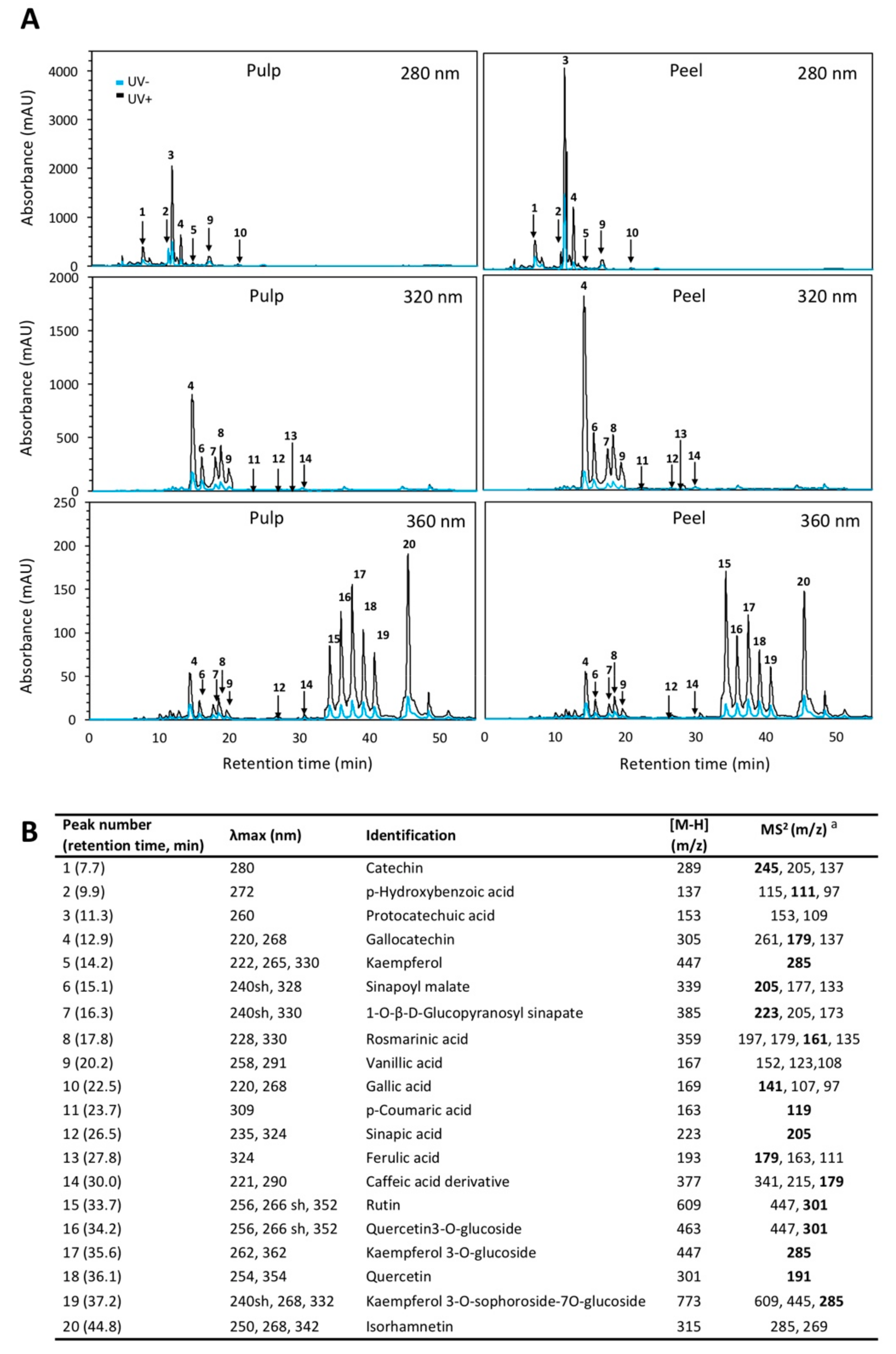
2.1. Phenolic Profile of Red Prickly Pear Pulp and Peel
2.2. Immediate Effect of UVB Radiation on the Phenolic Profile of Whole and Wounded Red Prickly Pear Pulp
2.3. Effect of Storage Time on the Phenolic Profile of Whole and Wounded Red Prickly Pear Pulp Treated with UVB Radiation
2.4. Immediate Effect of UVB Radiation on the Phenolic Profile of Whole and Wounded Red Prickly Pear Peel
2.5. Effect of Storage Time on the Phenolic Profile of Whole and Wounded Red Prickly Pear Peel Treated with UVB Radiation
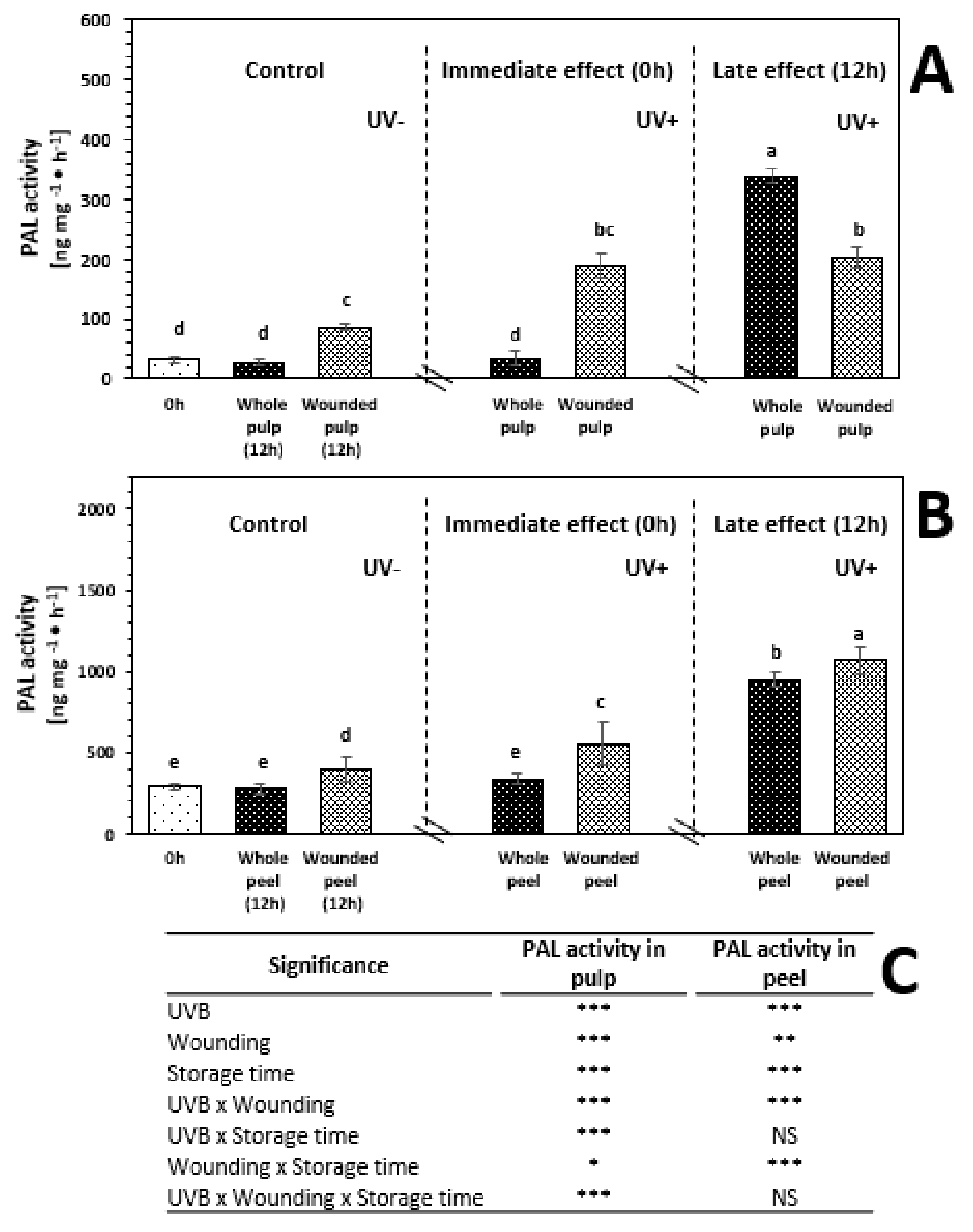
2.6. Phenylalanine Ammonia-Lyase (PAL) Activity of Whole and Wounded Red Prickly Pear Treated with UVB Radiation
2.7. Effect of UVB, Wounding Stress, and Storage Time on Total Ascorbic Acid Content of Red Prickly Pear
2.8. l-Galactono-γ-Lactone Dehydrogenase (GalLDH) Activity
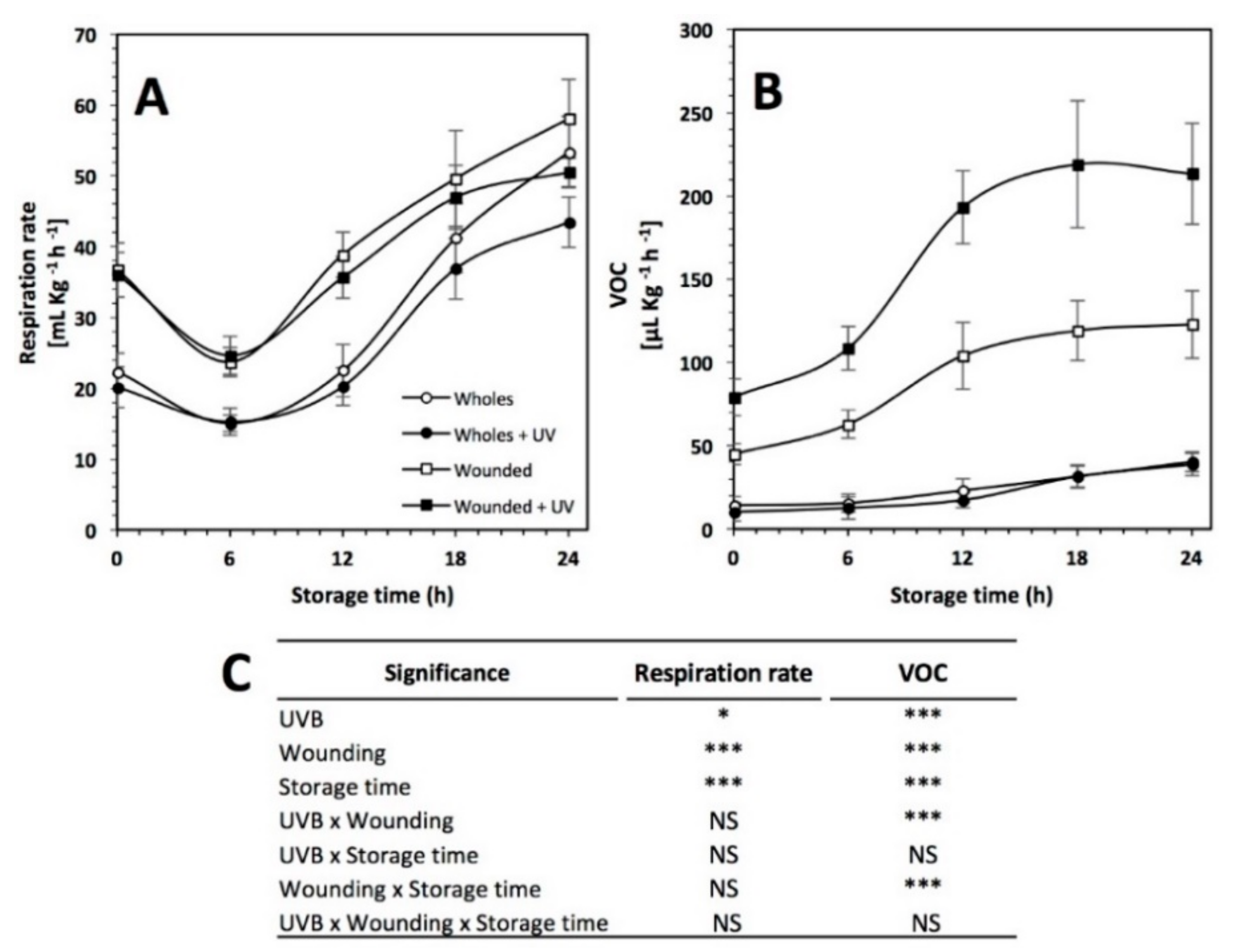
2.9. Respiration Rate and Ethylene Analysis
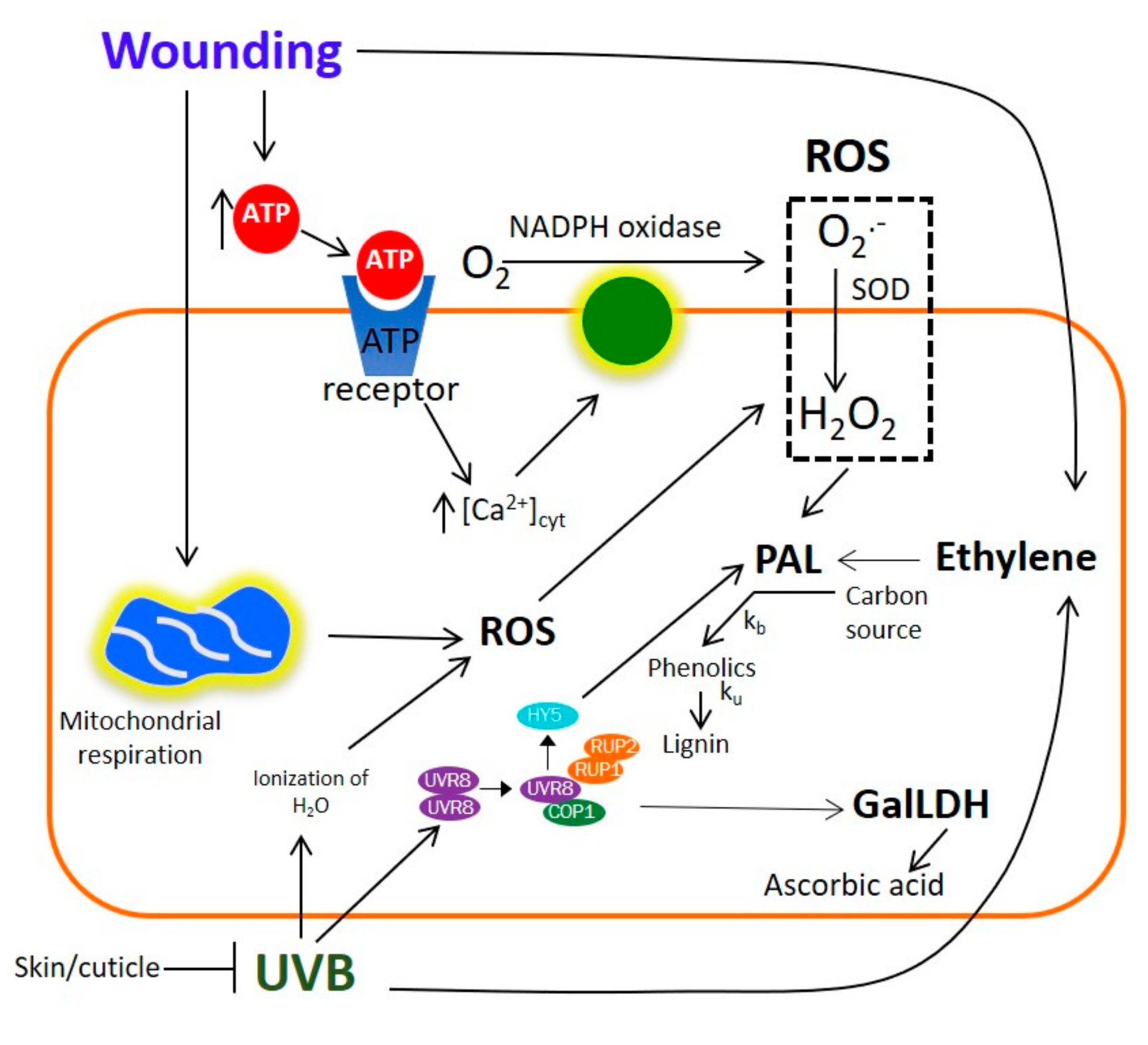
2.10. Potential Mechanism for the UVB and Wound-Induced Accumulation of Phenolic Compounds and Ascorbic Acid in Prickly Pear
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. UVB Treatments and Storage Study
3.4. Phytochemical Analyses
3.4.1. Extraction of Phenolic Compounds
3.4.2. Identification and Quantification of Phenolic Compounds
3.4.3. Determination of Ascorbic Acid
3.5. Enzymatic Analyses
3.5.1. Phenylalanine Ammonia-Lyase (PAL) Assay
3.5.2. l-Galactono-γ-lactone Dehydrogenase (GalLDH) Activity
3.6. Protein Assay
3.7. Respiration Rate and Ethylene Analyses
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Cotazar, V.G.; Nobel, P.S. Biomass and fruit production for the prickly pear cactus Opuntia ficus-indica. J. Am. Soc. Hortic Sci. 1992, 117, 558–562. [Google Scholar] [CrossRef]
- Chauhan, S.P.; Sheth, N.R.; Jivani, N.P.; Rathod, I.S.; Shah, P.I. Biological actions of opuntia species. Syst. Rev. Pharm. 2010, 1, 146–151. [Google Scholar] [CrossRef]
- Correia, R.T.; Borges, K.C.; Medeiros, M.F.; Genovese, M.I. Bioactive compounds and phenolic-linked functionality of powdered tropical fruit residues. Food Sci. Technol. Int. 2012, 18, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in brassica vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef]
- Yeddes, N.; Chérif, J.; Guyot, S.; Sotin, H.; Ayadi, M. Comparative study of antioxidant power, polyphenols, flavonoids and betacyanins of the peel and pulp of three tunisian opuntia forms. Antioxidants 2013, 2, 37–51. [Google Scholar] [CrossRef]
- Kuti, J.O. Growth and compositional changes during the development of prickly pear fruit. J. Am. Soc. Hortic. Sci. 1992, 67, 861–868. [Google Scholar] [CrossRef]
- Russel, C.; Felker, P. The prickly pear (Opuntia spp., Cactaceae): A source of human and animal food in semi-arid regions. Econ. Bot. 1987, 41, 433–445. [Google Scholar] [CrossRef]
- Iturriaga, L. Functional Components and Medicinal Properties of Cactus Products. In Functional Properties of Traditional Foods, 1st ed.; Kristbergsson, K., Otles, S., Eds.; Springer: New York, NY, USA, 2016; p. 258. [Google Scholar]
- Cisneros-Zevallos, L. The use of controlled postharvest abiotic stresses as a tool for enhancing the nutraceutical content and adding-value of fresh fruits and vegetables. J. Food Sci. 2003, 68, 1560–1565. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; González-Agüero, M.; Cisneros-Zevallos, L. Cross-talk between signaling pathways: The link between plant secondary metabolite production and wounding. Sci. Rep. 2015, 5, 8608. [Google Scholar] [CrossRef]
- Schreiner, M.; Krumbein, A.; Mewis, I.; Ulrichs, C.; Huyskens-Keil, S. Short-term UV-B radiation effects on secondary metabolism in different organs of Tropaeolum Maj. L. Innov. Food Sci. Emerg. Technol. 2009, 10, 93–96. [Google Scholar] [CrossRef]
- Heijde, M.; Ulm, R. UV-B photoreceptor-mediated signalling in plants. Trends Plant Sci. 2012, 17, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Lavola, A. Accumulation of flavonoids and related compounds in birch induced by UV-B irradiance. Tree Physiol. 1998, 18, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hameed, S.S.; Nagaty, M.A.; Salman, M.S.; Bazaid, S.A. Phytochemicals, nutritionals and antioxidant properties of two prickly pear cactus cultivars (Opuntia ficus-indica Mill.) growing in Taif, KSA. Food Chem. 2014, 160, 31–38. [Google Scholar] [CrossRef]
- Becerra-Moreno, A.; Redondo-Gil, M.; Benavides, J.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Combined effect of water loss and wounding stress on gene activation of metabolic pathways associated with phenolic biosynthesis in carrot. Front. Plant Sci. 2015, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Booij-James, I.S.; Dube, S.K.; Jansen, M.A.; Edelman, M.; Mattoo, A.K. Ultraviolet-B radiation impacts light-mediated turnover of the photosystem II reaction center heterodimer in Arabidopsis mutants altered in phenolic metabolism. Plant Physiol. 2000, 124, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Milkowski, C.; Baumert, A.; Schmidt, D.; Nehlin, L.; Strack, D. Molecular regulation of sinapate ester metabolism in Brassica napus: Expression of genes, properties of the encoded proteins and correlation of enzyme activities with metabolite accumulation. Plant J. 2004, 38, 80–92. [Google Scholar] [CrossRef]
- Moreira-Rodríguez, M.; Nair, V.; Benavides, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. UVA, UVB light, and methyl jasmonate, alone or combined, redirect the biosynthesis of glucosinolates, phenolics, carotenoids, and chlorophylls in broccoli sprouts. Int. J. Mol. Sci. 2017, 18, 2330. [Google Scholar] [CrossRef]
- Moreira-Rodríguez, M.; Nair, V.; Benavides, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. UVA, UVB light doses and harvesting time differentially tailor glucosinolate and phenolic profiles in broccoli sprouts. Molecules 2017, 22, 1065. [Google Scholar] [CrossRef]
- Heinze, M.; Hanschen, F.S.; Wiesner-Reinhold, M.; Baldermann, S.; Gräfe, J.; Schreiner, M.; Neugart, S. Effects of Developmental Stages and Reduced UVB and Low UV Conditions on Plant Secondary Metabolite Profiles in Pak Choi (Brassica rapa subsp. chinensis). J. Agric. Food Chem. 2018, 66, 1678–1692. [Google Scholar] [CrossRef]
- Silva, N.C.; Macedo, A.; Lage, C.L.; Esquibel, M.A.; Sato, A. Developmental effects of additional ultraviolet a radiation, growth regulators and tyrosine in Alternanthera brasiliana (L.) Kuntze cultured in vitro. Braz. Arch. Biol. Technol. 2005, 48, 779–786. [Google Scholar] [CrossRef]
- Jansen, M.A.K.; Hectors, K.; O’Brien, N.M.; Guisez, Y.; Potters, G. Plant stress and human health: Do human consumer benefit from UV-B acclimated crops? Plant Sci. 2008, 175, 449–458. [Google Scholar] [CrossRef]
- Goetz, M.; Albert, A.; Stich, S.; Heller, W.; Scherb, H.; Krins, A.; Langebartels, C.; Seidlitz, H.K.; Ernst, D. PAR modulation of the UV-dependent levels of flavonoid metabolites in Arabidopsis thaliana (L.) Heynh. Leaf rosettes: Cumulative effects after a whole vegetative growth period. Protoplasma 2010, 243, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, M.M.; Robberecht, R.; Flint, S.D. Internal filters: Prospects for UV-acclimation in higher plants. Physiol. Plant 1983, 58, 445–450. [Google Scholar] [CrossRef]
- Jenkins, G.I. Signal transduction in response to UV-B radiation. Annu Rev. Plant Biol. 2009, 60, 407–431. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Hernández, E.; Welti-Chanes, J.; Jacobo-Velázquez, D.A. Effects of UVB Light, Wounding Stress, and Storage Time on the Accumulation of Betalains, Phenolic Compounds, and Ascorbic Acid in Red Prickly Pear (Opuntia ficus-indica cv. Rojo Vigor). Food Bioprocess Technol. 2018, 11, 2265–2274. [Google Scholar] [CrossRef]
- Giuntini, D.; Lazzeri, V.; Calvenzani, V.; Dall’Asta, C.; Galaverna, G.; Tonelli, C. Flavonoid profiling and biosynthetic gene expression in flesh and peel of two tomato genotypes grown under UV-B-depleted conditions during ripening. J. Agric. Food Chem. 2008, 56, 5905–5915. [Google Scholar] [CrossRef]
- Surjadinata, B.B.; Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. UVA, UVB and UVC light enhances the biosynthesis of phenolic antioxidants in fresh-cut carrot through a synergistic effect with wounding. Molecules 2017, 22, 668. [Google Scholar] [CrossRef]
- Mewis, I.; Schreiner, M.; Nguyen, C.N.; Krumbein, A.; Ulrichs, C.; Lohse, M.; Zrenner, R. UV-B irradiation changes specifically the secondary metabolite profile in broccoli sprouts: Induced signaling overlaps with defense response to biotic stressors. Plant Cell Physiol. 2012, 53, 1546–1560. [Google Scholar] [CrossRef]
- Simontacchi, M.; Galatro, A.; Ramos-Artuso, F.; Santa-María, G. Plant Survival in a Changing Environment: The Role of Nitric Oxide in Plant Responses to Abiotic Stress. Front. Plant Sci. 2015, 9, 977. [Google Scholar] [CrossRef]
- Neugart, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Structurally different flavonol glycosides and hydroxycinnamic acid derivatives respond differently to moderate UV-B radiation exposure. Physiol. Plant 2012, 145, 582–593. [Google Scholar] [CrossRef]
- Lin, J.S.; Huang, X.X.; Li, Q.; Cao, Y.; Bao, Y.; Meng, X.F.; Li, Y.J.; Fu, C.; Hou, B.K. UDP-glycosyltransferase 72B1 catalyzes the glucose conjugation of monolignols and is essential for the normal cell wall lignification in Arabidopsis thaliana. Plant J. 2016, 88, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Jacobo-Velázquez, D.; Martínez-Hernández, G.; Rodríguez, S.C.; Cao, C.M.; Cisneros-Zevallos, L. Plants as biofactories: Physiological role of reactive oxygen species on the accumulation of phenolic antioxidants in carrot tissue under wounding and hyperoxia stress. J. Agric. Food Chem. 2011, 59, 6583–6593. [Google Scholar] [CrossRef] [PubMed]
- Butera, D.; Tesoriere, L.; Di Gaudio, F.; Bongiorno, A.; Allegra, M.; Pintaudi, A.M.; Kohen, R.; Livrea, M.A. Antioxidant Activities of Sicilian Prickly Pear (Opuntia ficus indica) Fruit Extracts and Reducing Properties of Its Betalains: Betanin and Indicaxanthin. J. Agric. Food Chem. 2002, 50, 6895–6901. [Google Scholar] [CrossRef] [PubMed]
- Kovácsa, E.; Keresztesb, Á. Effect of gamma and UV-B/C radiation on plant cells. Micron 2002, 33, 199–210. [Google Scholar] [CrossRef]
- Castagna, A.; Chiavaro, E.; Dall’asta, C.; Rinaldi, M.; Galaverna, G.; Ranieri, A. Effect of postharvest UV-B irradiation on nutraceutical quality and physical properties of tomato fruits. Food Chem. 2012, 137, 151–158. [Google Scholar] [CrossRef]
- Reyes, L.F.; Villarreal, J.E.; Cisneros-Zevallos, L. The increase in antioxidant capacity after wounding depends on the type of fruit and vegetable tissue. Food Chem. 2007, 101, 1254–1262. [Google Scholar] [CrossRef]
- Morales, L.O.; Brosché, M.; Vainonen, J.; Jenkins, G.I.; Wargent, J.J.; Sipari, N.; Srid, A.; Lindfors, A.V.; Tegelberg, R.; Aphalo, P.J. Multiple roles for UV RESISTANCE LOCUS8 in regulating gene expression and metabolite accumulation in Arabidopsis under solar ultraviolet radiation. Plant Physiol. 2013, 161, 744–759. [Google Scholar] [CrossRef]
- Wang, H.; Gui, M.; Tian, X.; Xin, X.; Wang, T.; Li, J. Effects of UV-B on vitamin C, phenolics, flavonoids and their related enzyme activities in mung bean sprouts (Vigna radiata). Food Sci. Technol. 2017, 52, 827–833. [Google Scholar] [CrossRef]
- Guo, X.; Liu, R.H.; Fu, X.; Sun, X.; Tang, K. Over-expression of L-galactono-γ-lactone dehydrogenase increases vitamin C, total phenolics and antioxidant activity in lettuce through bio-fortification. Plant Cell Tissue Organ. Cult. 2013, 114, 225–236. [Google Scholar] [CrossRef]
- Abeles, F.B.; Morgan, P.W.; Saltveit, M.E. Ethylene in Plant Biology, 2nd ed.; Academic Press Inc: San Diego, CA, USA, 1992; pp. 56–119. [Google Scholar]
- Corrales-García, J.; Andrade-Rodríguez, J.; Bernabé-Cruz, E. Responses of six cultivars of tuna fruits to cold storage. J. Prof. Assoc. Cactus Dev. 1997, 2, 160–168. [Google Scholar]
- Hajirezaei, M.; Stitt, M. Contrasting roles for pyrophosphate: Fructose 6-phosphate phosphotransferase during aging of tissue slices from potato tubers and carrot storage tissues. Plant Sci. 1991, 77, 177–183. [Google Scholar] [CrossRef]
- Asahi, T. Biogenesis of cell organelles in wounded plant storage tissue cells. In Biochemistry of Wounded Plant Storage Tissues; Kahl, G., Ed.; Walter De Grutyer & Co.: Berlin, Germany, 1978; pp. 391–419. [Google Scholar]
- Laties, G.G. The development and control of respiratory pathways in slices of plant storage organs. In Biochemistry of Wounded Plant Storage Tissues; Kahl, G., Ed.; Walter De Grutyer & Co.: Berlin, Germany, 1978; pp. 421–466. [Google Scholar]
- Germ, M.; Simčič, T.; Gaberščik, A.; Breznik, B.; Hrastel, M. UV-B treated algae exhibiting different responses as a food source for Daphnia magna. J. Plankton Res. 2004, 26, 1219–1228. [Google Scholar] [CrossRef]
- Borisova, T.A.; Bugaje, S.M.; Meshkova, N.V.; Vlasov, P.V. Heat shock increases the tolerance of plants to UV-B radiation: 1. growth, development, and water supply to tissues. Russ. J. Plant Physiol. 2001, 48, 589–595. [Google Scholar]
- Zhalilova, F.K.; Rakitina, T.Y.; Vlasov, P.V.; Kefeli, V.I. Growth and Ethylene Evolution in Three Genetic Lines of Arabidopsis thaliana, as Affected by Ultraviolet Radiation (UV-B). Russ. J. Plant Physiol. 1993, 40, 764–769. [Google Scholar]
- Rushing, J. Cytokinins affect respiration, ethylene production, and chlorophyll retention of packaged broccoli florets. HortScience 1990, 25, 88–90. [Google Scholar] [CrossRef]
- Surjadinata, B.B.; Cisneros-Zevallos, L. Modeling wound-induced respiration of fresh-cut carrots (Daucus carota L.). J. Food Sci. 2006, 68, 2735–2740. [Google Scholar] [CrossRef]
- Singh, S.; Agrawal, S.; Agrawal, M. UVR8 mediated plant protective responses under low UV-B radiation leading to photosynthetic acclimation. J. Photochem. Photobiol. 2014, 137, 67–76. [Google Scholar] [CrossRef]
- Kader, A. Cactus (Prickly) Pear. Recommendations for Maintaining Postharvest Quality. 1999. Available online: http://postharvest.ucdavis.edu/Commodity_Resources/Fact_Sheets/Datastores/Fruit_English/?uid =13&ds=798 (accessed on 19 May 2017).
- Castellanos Santiago, E.; Yahia, E.M. Identification and quantification of betalains from the fruits of 10 mexican prickly pear cultivars by high-performance liquid chromatography and electrospray ionization mass spectrometry. J. Agric. Food Chem. 2008, 56, 5758–5764. [Google Scholar] [CrossRef]
- Gillespie, K.; Ainsworth, E. Measurement of reduced, oxidized and total ascorbate content in plants. Nat. Protoc. 2007, 2, 871–874. [Google Scholar] [CrossRef]
- Ôba, K.; Fukui, M.; Imai, Y.; Iriyama, S.; Nogami, K. L-Galactono-γ-lactone dehydrogenase: Partial characterization, induction of activity and role in the synthesis of ascorbic acid in wounded white potato tuber tissue. Plant Cell Physiol. 1994, 35, 473–478. [Google Scholar]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Meenune, M. Changes in physiochemical quality and browning related enzyme activity of longkong fruit during four different weeks of on-tree maturation. Food Chem. 2012, 131, 1437–1442. [Google Scholar] [CrossRef]
- Cuéllar-Villarreal, M.; Ortega-Hernández, E.; Becerra-Moreno, A.; Welti-Chanes, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D. Effects of ultrasound treatment and storage time on the extractability and biosynthesis of nutraceuticals in carrot (Daucus carota). Postharvest Biol. Technol. 2016, 119, 18–26. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Storage Time | Phenolic Compounds Concentration (mg/100 g) DW 1,2,3,4 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Catechin | p-Hydroxybenzoic acid | Protocatechuic acid | Gallocatechin | Kaempferol | Sinapoyl malate | 1-O-β-d-glucopyranosyl sinapate | |||
| Control | 0 h | 11.54 ± 1.26 a | 42.14 ± 0.95 ab | 197.08 ± 14.51 cd | 15.79 ± 1.08 ab | 23.49 ± 2.00 c | 26.89 ± 1.12 de | 1.17 ± 0.08 d | |
| UV- | Whole | 24 h | 11.99 ± 1.39 a | 44.27 ± 3.61 a | 212.86 ± 14.41 c | 17.22 ± 1.12 a | 20.71 ± 2.09 c | 24.36 ± 0.93 e | 1.20 ± 0.03 d |
| Wounded | 8.44 ± 1.25 c | 38.94 ± 2.17 b | 220.73 ± 15.02 c | 14.13 ± 0.89 b | 21.17 ± 2.14 c | 29.47 ± 1.26 d | 1.36 ± 0.17 d | ||
| UV+ | Whole | 0 h | 10.20 ± 0.23 b | 39.40 ± 0.84 b | 259.90 ± 18.12 b | 13.09 ± 0.15 c | 29.85 ± 2.83 b | 31.55 ± 1.80 d | 1.63 ± 0.03 c |
| Wounded | 8.15 ± 0.21 c | 46.35 ± 0.64 a | 230.85 ± 14.31 b | 11.66 ± 0.78 c | 31.43 ± 2.05 b | 42.83 ± 1.09 c | 1.86 ± 0.07 b | ||
| Whole | 24 h | 12.29 ± 0.12 a | 39.93 ± 0.45 ab | 299.94 ± 17.84 a | 17.82 ± 1.23 a | 34.07 ± 3.79 b | 70.10 ± 2.32 b | 1.90 ± 0.03 b | |
| Wounded | 6.09 ± 0.86 d | 37.49 ± 0.65 b | 215.21 ± 7.67 c | 15.71 ± 1.73 ab | 62.92 ± 4.87 a | 93.56 ± 6.42 a | 7.87 ± 2.39 a | ||
| Significance | |||||||||
| UVB | NS | *** | *** | *** | *** | *** | *** | ||
| Wounding (W) | *** | *** | *** | *** | *** | *** | *** | ||
| Storage time (ST) | *** | *** | *** | *** | *** | *** | *** | ||
| UVB × W | ** | * | *** | *** | *** | *** | *** | ||
| UVB × ST | ** | NS | *** | *** | *** | *** | *** | ||
| W × ST | *** | *** | *** | *** | *** | *** | *** | ||
| UVB × W × ST | ** | NS | ** | *** | *** | *** | *** | ||
| Treatment | Storage Time | Phenolic Compounds Concentration (mg/100 g) DW 1,2,3,4 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Rosmarinic acid | Vanillic acid | Gallic acid | p-Coumaric acid | Sinapic acid | Ferulic acid | Caffeic acid derivative | |||
| Control | 0 h | 5.83 ± 0.21 c | 6.87 ± 0.85 c | 15.74 ± 0.54 ab | 1.53 ± 0.15 c | 0.96 ± 0.04 cd | 1.42 ± 0.07 c | 1.19 ± 0.04 c | |
| UV- | Whole Wounded | 24 h | 4.50 ± 0.27 d | 6.17 ± 0.62 c | 14.85 ± 1.45 b | 1.36 ± 0.37 c | 1.04 ± 0.15 cd | 1.32 ± 0.18 c | 1.29 ± 0.15 c |
| 6.56 ± 0.79 c | 5.69 ± 2.39 d | 13.93 ± 1.62 b | 1.93 ± 0.36 b | 1.48 ± 0.03 b | 1.72 ± 0.28 b | 1.64 ± 0.05 b | |||
| UV+ | Whole | 0 h | 5.58 ± 1.12 c | 6.47 ± 0.08 c | 17.04 ± 0.10 a | 1.69 ± 0.10 bc | 1.06 ± 0.06 cd | 1.31 ± 0.03 cd | 1.27 ± 0.02 c |
| Wounded | 8.65 ± 0.40 b | 7.86 ± 0.16 a | 16.64 ± 2.26 a | 8.25 ± 0.05 a | 2.99 ± 0.06 a | 6.99 ± 0.07 a | 5.65 ± 0.01 a | ||
| Whole | 24 h | 30.80 ± 2.35 a | 7.09 ± 0.27 ab | 13.49 ± 0.31 b | 0.64 ± 0.19 e | 1.28 ± 0.04 b | 1.34 ± 0.02 cd | 1.45 ± 0.01 c | |
| Wounded | 26.87 ± 3.77 a | 5.14 ± 0.03 cd | 12.70 ± 0.06 b | 0.93 ± 0.06 d | 1.16 ± 0.14 c | 1.28 ± 0.11 d | 1.18 ± 0.09 c | ||
| Significance | |||||||||
| UVB | *** | ** | *** | NS | *** | NS | NS | ||
| Wounding (W) | *** | * | *** | *** | *** | *** | *** | ||
| Storage time (ST) | *** | NS | *** | NS | *** | NS | * | ||
| UVB × W | *** | * | ** | ** | *** | *** | *** | ||
| UVB × ST | *** | * | ** | *** | *** | ** | *** | ||
| W × ST | *** | NS | NS | NS | *** | NS | *** | ||
| UVB × W × ST | *** | * | * | *** | *** | *** | *** | ||
| Treatment | Storage Time | Phenolic Compounds Concentration (mg/100 g) DW 1,2,3,4 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Rutin | Quercetin 3-O-glucoside | Kaempferol 3-O-glucoside | Quercetin | Kaempferol 3-O-sophoroside-7O-glucoside | Isorhamnetin | Total | |||
| Control | 0 h | 1.08 ± 0.06 b | 0.91 ± 0.04 d | 0.38 ± 0.02 d | 33.18 ± 2.30 c | 0.72 ± 0.11 c | 1.80 ± 0.03 ab | 389.70 ± 32.15 e | |
| UV- | Whole | 24 h | 1.25 ± 0.11 a | 0.97 ± 0.06 d | 0.35 ± 0.00 d | 36.20 ± 1.03 c | 0.63 ± 0.06 d | 1.66 ± 0.15 ab | 404.18 ± 27.09 e |
| Wounded | 1.07 ± 0.01 b | 2.23 ± 0.00 c | 0.49 ± 0.04 c | 36.28 ± 1.12 c | 0.69 ± 0.05 c | 1.71 ± 0.13 ab | 409.67 ± 16.56 e | ||
| UV+ | Whole | 0 h | 1.24 ± 0.00 a | 0.81 ± 0.02 d | 0.60 ± 0.02 b | 33.28 ± 2.01 c | 1.05 ± 0.07 b | 1.51 ± 0.04 ab | 458.52 ± 32.42 de |
| Wounded | 1.13 ± 0.00 ab | 4.23 ± 0.00 b | 1.05 ± 0.00 b | 47.53 ± 1.30 a | 1.35 ± 0.00 a | 1.69 ± 0.03 ab | 487.25 ± 28.78 c | ||
| Whole | 24 h | 1.24 ± 0.01 a | 1.24 ± 0.00 c | 0.05 ± 0.00 e | 44.14 ± 3.00 b | 0.99 ± 0.03 b | 1.54 ± 0.00 ab | 581.34 ± 43.36 a | |
| Wounded | 0.97 ± 0.00 bc | 7.79 ± 0.38 a | 6.82 ± 0.45 a | 52.13 ± 3.94 a | 1.49 ± 0.18 a | 1.90 ± 0.04 a | 559.21 ± 37.85 b | ||
| Significance | |||||||||
| UVB | *** | *** | *** | *** | *** | *** | *** | ||
| Wounding (W) | *** | *** | *** | *** | * | *** | *** | ||
| Storage time (ST) | *** | *** | *** | *** | ** | *** | *** | ||
| UVB × W | *** | *** | *** | *** | ** | *** | *** | ||
| UVB × ST | *** | *** | *** | *** | *** | *** | *** | ||
| W × ST | *** | *** | *** | *** | *** | *** | *** | ||
| UVB × W × ST | *** | *** | *** | *** | *** | *** | *** | ||
| Treatment | Storage Time | Phenolic Compounds Concentration (mg/100 g) DW 1,2,3,4 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Catechin | p-Hydroxybenzoic acid | Protocatechuic acid | Gallocatechin | Kaempferol | Sinapoyl malate | 1-O-β-d-Glucopyranosyl sinapate | |||
| Control | 0 h | 46.55 ± 3.60 b | 13.67 ± 0.95 a | 453.05 ± 31.56 a | 29.91 ± 1.93 a | 6.01 ± 1.39 e | 55.48 ± 2.28 f | 10.00 ± 0.79 e | |
| UV- | Whole | 24 h | 42.72 ± 2.55 b | 11.21 ± 0.36 b | 390.78 ± 24.24 b | 26.52 ± 1.03 ab | 5.60± 4.38 f | 75.04 ± 4.63 e | 25.36 ± 0.88 d |
| Wounded | 51.20 ± 3.67 ab | 12.98 ± 1.03 a | 468.71 ± 41.41 a | 33.47 ± 2.72 a | 6.20 ± 2.41 e | 58.30 ± 2.49 f | 8.13 ± 1.67 e | ||
| UV+ | Whole | 0 h | 43.20 ± 2.45 b | 7.33 ± 0.14 c | 216.37 ± 19.12 d | 24.53 ± 2.51 b | 38.14 ± 2.17 a | 86.92 ± 2.37 d | 38.10 ± 1.51 cd |
| Wounded | 37.78 ± 2.32 c | 6.47 ± 0.78 cd | 389.04 ± 28.30 b | 21.67 ± 1.82 c | 28.66 ± 1.29 c | 128.99 ± 4.95 c | 43.47 ± 1.55 bc | ||
| Whole | 24 h | 45.12 ± 2.86 b | 4.00 ± 0.78 d | 412.00 ± 37.55 ab | 18.84 ± 2.35 d | 32.60 ± 1.44 b | 158.83 ± 12.65 b | 49.21 ± 0.82 b | |
| Wounded | 32.25 ± 2.82 d | 2.63 ± 0.89 e | 330.77 ± 29.04 c | 12.04 ± 1.78 e | 23.31 ± 2.41 d | 202.26 ± 18.81 a | 55.46 ± 5.20 a | ||
| Significance | |||||||||
| UVB | *** | NS | *** | *** | *** | *** | *** | ||
| Wounding (W) | NS | *** | *** | *** | *** | *** | *** | ||
| Storage time (ST) | NS | *** | *** | NS | NS | *** | *** | ||
| UVB × W | NS | *** | * | *** | *** | NS | NS | ||
| UVB × ST | *** | *** | NS | *** | *** | NS | NS | ||
| W × ST | *** | *** | NS | *** | *** | * | *** | ||
| UVB × W × ST | *** | *** | * | *** | *** | NS | NS | ||
| Treatment | Storage Time | Phenolic Compounds Concentration (mg/100 g) DW 1,2,3,4 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Rosmarinic acid | Vanillic acid | Gallic acid | p-Coumaric acid | Sinapic acid | Ferulic acid | Caffeic acid derivative | |||
| Control | 0 h | 38.73 ± 1.75 e | 8.15 ± 0.69 a | 7.27 ± 0.88 ab | 1.32 ± 0.15 d | 2.33 ± 0.09 d | 1.55 ± 0.14 d | 0.74 ± 0.06 c | |
| UV- | Whole | 24 h | 54.56 ± 2.57 d | 7.13 ± 1.00 ab | 8.13 ± 0.15 a | 1.82 ± 0.04 c | 4.47 ± 0.58 c | 2.92 ± 0.35 c | 2.70 ± 0.09 a |
| Wounded | 42.02 ± 5.79 de | 8.53 ± 0.51 a | 6.51 ± 0.03 b | 1.23 ± 0.45 d | 2.11 ± 0.18 d | 1.39 ± 0.61 d | 0.61 ± 0.29 d | ||
| UV+ | Whole | 0 h | 87.11 ± 6.86 c | 7.10 ± 1.38 b | 7.13 ± 0.10 ab | 1.91 ± 0.12 c | 5.76 ± 0.08 c | 1.71 ± 0.27 d | 0.69 ± 0.02 d |
| Wounded | 141.97 ± 6.06 b | 6.95 ± 0.04 b | 7.95 ± 0.03 a | 8.37 ± 1.12 a | 14.36 ± 0.93 b | 21.15 ± 1.01 a | 1.87 ± 0.05 b | ||
| Whole | 24 h | 209.11 ± 0.38 a | 3.94 ± 0.26 d | 7.45 ± 0.08 ab | 1.90 ± 0.13 c | 15.63 ± 1.21 b | 5.36 ± 1.99 b | 0.81 ± 0.02 c | |
| Wounded | 155.71 ± 8.55 b | 5.59 ± 2.05 c | 7.77 ± 0.72 a | 2.83 ± 0.69 b | 24.89 ± 0.77 a | 5.5± 0.57 b | 2.99 ± 0.07 a | ||
| Significance | |||||||||
| UVB | NS | * | *** | *** | *** | NS | *** | ||
| Wounding (W) | *** | NS | * | *** | *** | NS | *** | ||
| Storage time (ST) | NS | NS | *** | *** | ** | NS | *** | ||
| UVB × W | NS | * | NS | *** | *** | NS | NS | ||
| UVB × ST | NS | NS | *** | ** | *** | NS | *** | ||
| W × ST | *** | NS | NS | *** | * | NS | ** | ||
| UVB × W × ST | *** | * | NS | *** | *** | NS | ** | ||
| Treatment | Storage Time | Phenolic Compounds Concentration (mg/100 g) DW 1,2,3,4 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Rutin | Quercetin 3-O-glucoside | Kaempferol 3-O-glucoside | Quercetin | Kaempferol 3-O-sophoroside-7O-glucoside | Isorhamnetin | Total | |||
| Control | 0 h | 9.09 ± 0.32 a | 24.59 ± 0.27 d | 13.98 ± 0.45 c | 70.74 ± 3.39 e | 1.28 ± 0.14 e | 17.29 ± 0.86 a | 811.74 ± 48.64 de | |
| UV- | Whole | 24 h | 7.34 ± 0.64 bc | 21.26 ± 6.59 d | 10.22 ± 0.41 cd | 63.29 ± 4.38 ef | 1.08 ± 0.24 e | 14.14 ± 1.22 ab | 776.29 ± 260.14 e |
| Wounded | 10.34 ± 0.14 a | 22.90 ± 1.05 d | 12.86 ± 1.48 c | 78.32 ± 2.41 e | 1.92 ± 0.31 d | 15.63 ± 0.73 ab | 843.37 ± 275.24 de | ||
| UV+ | Whole | 0 h | 8.55 ± 0.19 ab | 59.90 ± 5.74 a | 31.25 ± 1.81 a | 252.74 ± 24.03 d | 4.71 ± 0.17 b | 16.41 ± 1.95 a | 939.61 ± 89.50 d |
| Wounded | 8.20 ± 1.42 ab | 43.86 ± 3.59 c | 19.34 ± 3.48 b | 312.16 ± 21.29 c | 2.33 ± 0.21 c | 14.44 ± 0.26 ab | 1259.38 ± 96.94 c | ||
| Whole | 24 h | 9.45 ± 0.32 a | 53.54 ± 2.03 b | 33.55 ± 0.17 a | 572.60 ± 40.33 a | 5.05 ± 0.00 a | 15.11 ± 0.58 ab | 1654.06 ± 118.02 a | |
| Wounded | 6.57 ± 5.70 c | 42.46 ± 6.30 c | 22.91 ± 1.96 b | 428.65 ± 30.41 b | 1.49 ± 0.37 d | 10.05 ± 0.76 b | 1376.19 ± 134.56 b | ||
| Significance | |||||||||
| UVB | *** | NS | *** | NS | NS | NS | *** | ||
| Wounding (W) | *** | NS | NS | ** | *** | *** | *** | ||
| Storage time (ST) | NS | NS | *** | ** | ** | NS | *** | ||
| UVB × W | *** | NS | *** | NS | *** | NS | * | ||
| UVB × ST | *** | ** | *** | *** | *** | *** | NS | ||
| W × ST | ** | NS | *** | NS | NS | * | NS | ||
| UVB × W × ST | *** | NS | ** | * | NS | *** | * | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortega-Hernández, E.; Nair, V.; Welti-Chanes, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Wounding and UVB Light Synergistically Induce the Biosynthesis of Phenolic Compounds and Ascorbic Acid in Red Prickly Pears (Opuntia ficus-indica cv. Rojo Vigor). Int. J. Mol. Sci. 2019, 20, 5327. https://doi.org/10.3390/ijms20215327
Ortega-Hernández E, Nair V, Welti-Chanes J, Cisneros-Zevallos L, Jacobo-Velázquez DA. Wounding and UVB Light Synergistically Induce the Biosynthesis of Phenolic Compounds and Ascorbic Acid in Red Prickly Pears (Opuntia ficus-indica cv. Rojo Vigor). International Journal of Molecular Sciences. 2019; 20(21):5327. https://doi.org/10.3390/ijms20215327
Chicago/Turabian StyleOrtega-Hernández, Erika, Vimal Nair, Jorge Welti-Chanes, Luis Cisneros-Zevallos, and Daniel A. Jacobo-Velázquez. 2019. "Wounding and UVB Light Synergistically Induce the Biosynthesis of Phenolic Compounds and Ascorbic Acid in Red Prickly Pears (Opuntia ficus-indica cv. Rojo Vigor)" International Journal of Molecular Sciences 20, no. 21: 5327. https://doi.org/10.3390/ijms20215327
APA StyleOrtega-Hernández, E., Nair, V., Welti-Chanes, J., Cisneros-Zevallos, L., & Jacobo-Velázquez, D. A. (2019). Wounding and UVB Light Synergistically Induce the Biosynthesis of Phenolic Compounds and Ascorbic Acid in Red Prickly Pears (Opuntia ficus-indica cv. Rojo Vigor). International Journal of Molecular Sciences, 20(21), 5327. https://doi.org/10.3390/ijms20215327