Oncofertility: Pharmacological Protection and Immature Testicular Tissue (ITT)-Based Strategies for Prepubertal and Adolescent Male Cancer Patients

 and
and
Abstract
1. Introduction: The Quest for Fertility Preservation and Restoration
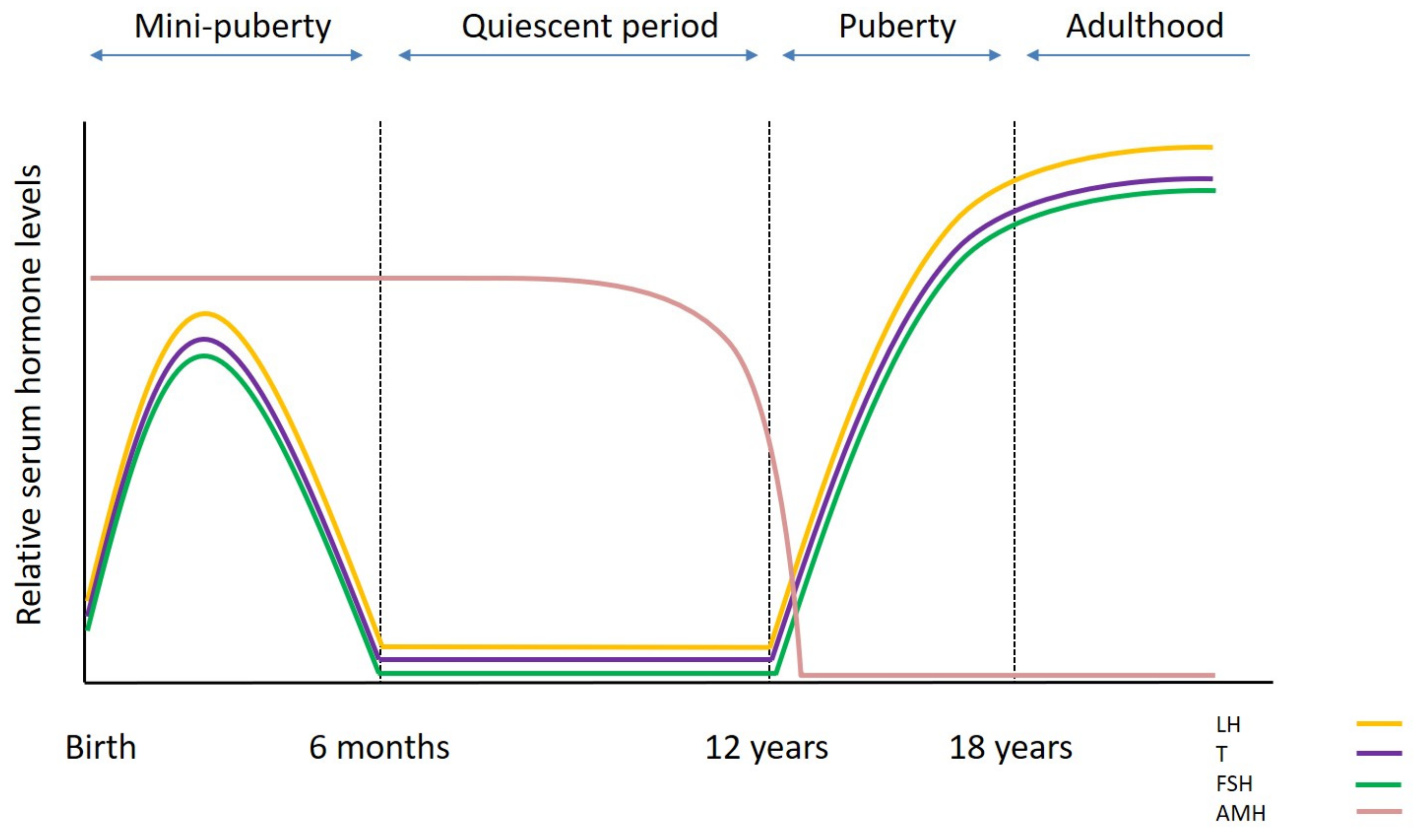
2. Testicular Development and Function
3. Cancer Treatment and Fertility Impairment
4. Strategy Based on Cryopreservation of Sperm
5. Experimental Strategies Based on Pharmacological Protection of the Testis
5.1. Hormonal Protection (GnRHa)
5.2. Granulocyte Colony-Stimulating Factor (G-CSF)
5.3. Antioxidant Treatment
5.4. Future Approaches
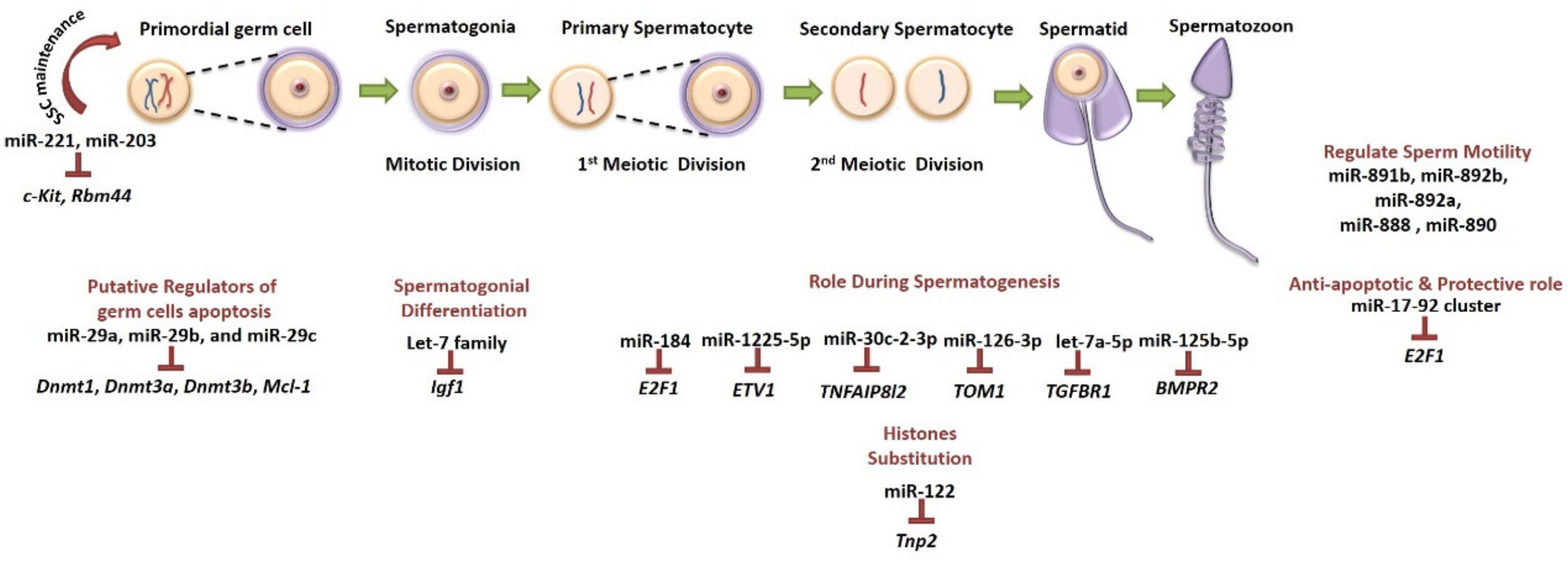
5.4.1. MicroRNAs as Fertility Preservation Tools
5.4.2. Predictive Biomarkers of Chemotherapy-Induced Infertility
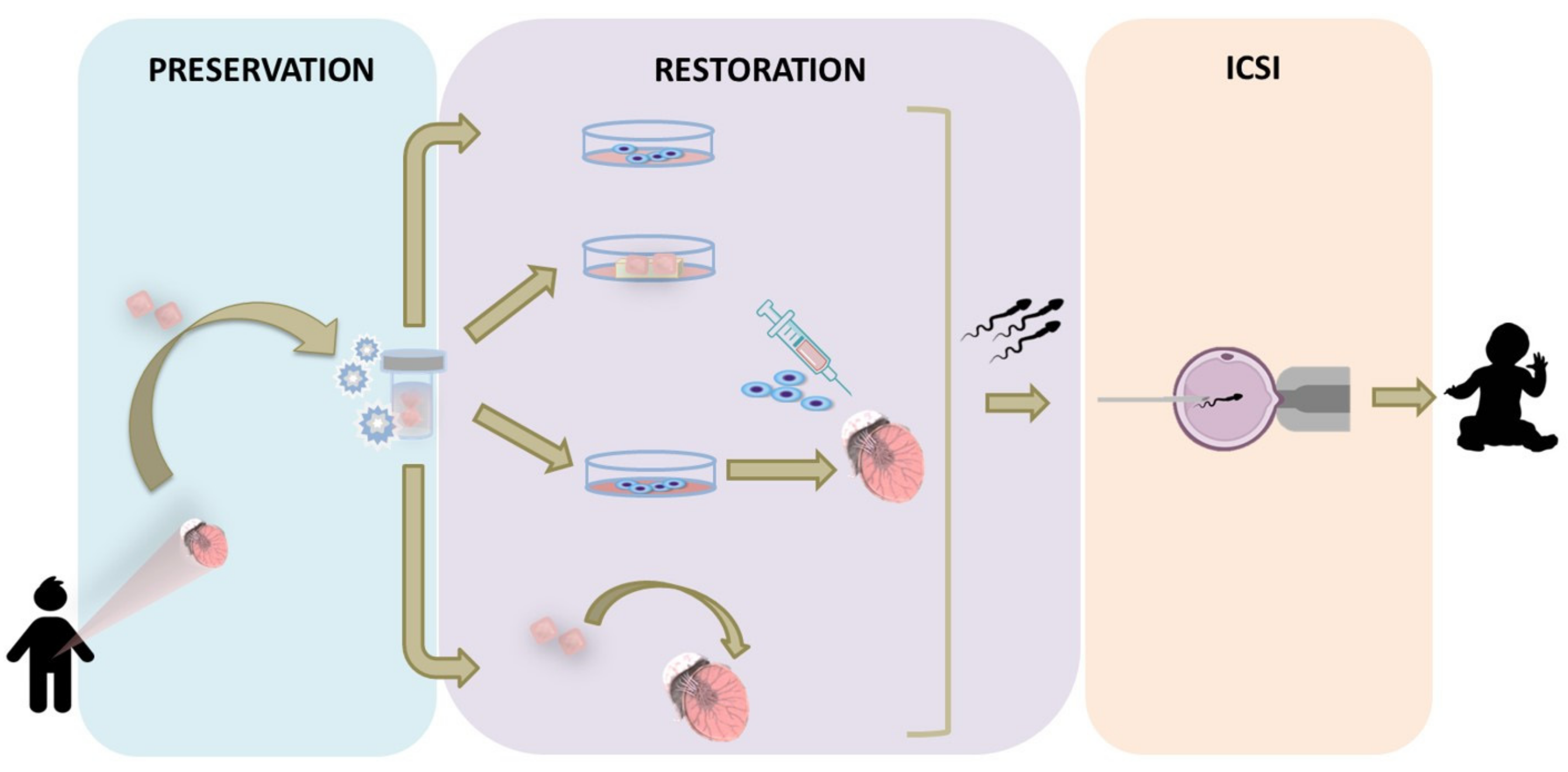
6. Experimental Strategies Based on Cryopreserved Immature Testicular Tissue
6.1. In Vitro Approaches
6.2. In Vivo Approaches
6.2.1. Spermatogonial Stem Cell Transplantation
6.2.2. Testicular Tissue Transplantation
6.2.3. Clinical Challenges and Concerns on ITT Transplantation
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMH | anti-Müllerian hormone |
| AR | androgen receptor |
| FSH | follicle stimulating hormone |
| G-CSF | granulocyte colony-stimulating factor |
| GnRH | gonadotrophin-releasing hormone |
| GnRHa | gonadotrophin-releasing hormone agonist |
| ICSI | intracytoplasmic sperm injection |
| ITT | immature testicular tissue |
| LH | luteinizing hormone |
| MAGE | melanoma-associated antigen |
| miRNA | microRNA |
| ROS | reactive oxygen species |
| SSC | spermatogonial stem cell |
| TTGFb1 | testosteronetransforming growth factor b1 |
References
- Steliarova-Foucher, E.; Colombet, M.; Ries, L.A.G.; Moreno, F.; Dolya, A.; Bray, F.; Hesseling, P.; Shin, H.Y.; Stiller, C.A. IICC-3 contributors. International incidence of childhood cancer, 2001–10: A population-based registry study. Lancet Oncol. 2017, 18, 719–731. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Diller, L.; Chow, E.J.; Gurney, J.G.; Hudson, M.M.; Kadin-Lottick, N.S.; Kawashima, T.I.; Leisenring, W.M.; Meacham, L.R.; Mertens, A.C.; Mulrooney, D.A.; et al. Chronic disease in the Childhood Cancer Survivor Study cohort: A review of published findings. J. Clin. Oncol. 2009, 27, 2339–2355. [Google Scholar] [CrossRef] [PubMed]
- Ward, E.; Desantis, C.; Robbins, A.; Kohler, B.; Jemal, A. Childhood and adolescent cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 83–103. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Green, D.M.; Kawashima, T.; Stovall, M.; Leisenring, W.; Sklar, C.A.; Mertens, A.C.; Donaldson, S.S.; Byrne, J.; Robison, L.L. Fertility of male survivors of childhood cancer: A report from the Childhood Cancer Survivor Study. J. Clin. Oncol. 2010, 28, 332–339. [Google Scholar] [CrossRef]
- Kaatsch, P. Epidemiology of childhood cancer. Cancer Treat. Rev. 2010, 36, 277–285. [Google Scholar] [CrossRef]
- Wasilewski-Masker, K.; Seidel, K.D.; Leisenring, W.; Mertens, A.C.; Shnorhavorian, M.; Ritenour, C.W.; Stovall, M.; Green, D.M.; Sklar, C.A.; Armstrong, G.T.; et al. Male infertility in long-term survivors of pediatric cancer: A report from the childhood cancer survivor study. J. Cancer Surviv. 2014, 8, 437–447. [Google Scholar] [CrossRef]
- Woodruff, T.K. Oncofertility: A grand collaboration between reproductive medicine and oncology. Reproduction 2015, 150, 1–10. [Google Scholar] [CrossRef]
- Picton, H.M.; Wyns, C.; Anderson, R.A.; Goossens, E.; Jahnukainen, K.; Kliesch, S.; Mitchell, R.T.; Pennings, G.; Rives, N.; Tournaye, H.; et al. ESHRE Task force on fertility preservation in severe diseases. A European perspective on testicular tissue cryopreservation for fertility preservation in prepubertal and adolescent boys. Hum Reprod. 2015, 30, 2463–2475. [Google Scholar] [CrossRef]
- Armstrong, G.T.; Chen, Y.; Yasui, Y.; Leisenring, W.; Gibson, T.M.; Mertens, A.C.; Stovall, M.; Oeffinger, K.C.; Bhatia, S.; Krull, K.R.; et al. Reduction in late mortality among 5-year survivors of childhood cancer. N. Engl. J. Med. 2016, 374, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Onofre, J.; Baert, Y.; Faes, K.; Goossens, E. Cryopreservation of testicular tissue or testicular cell suspensions: A pivotal step in fertility preservation. Hum. Reprod. Update 2016, 22, 744–761. [Google Scholar] [CrossRef] [PubMed]
- Rashedi, A.S.; de Roo, S.F.; Ataman, L.M.; Edmonds, M.E.; Silva, A.A.; Scarella, A.; Horbaczewska, A.; Anazodo, A.; Arvas, A.; Ramalho de Carvalho, B.; et al. Survey of fertility preservation options available to patients with cancer around the globe. J. Glob. Oncol. 2018, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Valli-Pulaski, H.; Peters, K.A.; Gassei, K.; Steimer, S.R.; Sukhwani, M.; Hermann, B.P.; Dwomor, L.; David, S.; Fayomi, A.P.; Munyoki, S.K.; et al. Testicular tissue cryopreservation: 8 years of experience from a coordinated network of academic centers. Hum. Reprod. 2019, 34, 966–977. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, D.G. The spermatogonial stem cell niche. Microsc. Res. Tech. 2009, 72, 580–585. [Google Scholar] [CrossRef]
- Caires, K.C.; Broady, J.; McLean, D. Maintaining the male germline: Regulation of spermatogonial stem cells. J. Endocrinol. 2010, 205, 133–145. [Google Scholar] [CrossRef]
- Oatley, J.M.; Brinster, R.L. The germline stem cell niche unit in mammalian testes. Physiol. Rev. 2012, 92, 577–595. [Google Scholar] [CrossRef]
- Rey, R.A. Mini-puberty and true puberty: Differences in testicular function. Ann. Endocrinol. 2014, 75, 58–63. [Google Scholar] [CrossRef]
- Hermann, B.P.; Sukhwani, M.; Hansel, M.C.; Orwig, K.E. Spermatogonial stem cells in higher primates: Are there differences from those in rodents? Reproduction 2010, 139, 479–493. [Google Scholar] [CrossRef]
- Mitchell, R.T.; O’Hara, L.; Smith, L.B. Gonadotropin and steroid hormone control of spermatogonial differentiation. In The Biology of Mammalian Spermatogonia, 1st ed.; Oatley, J.M., Griswold, M.D., Eds.; Springer Press: New York, NY, USA, 2017; pp. 4555–4570. [Google Scholar]
- Sharpe, R.M.; McKinnell, C.; Kivlin, C.; Fisher, J.S. Proliferation and functional maturation of Sertoli cells, and their relevance to disorders of testis function in adulthood. Reproduction 2003, 125, 769–784. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Mruk, D.D. A local autocrine axis in the testes that regulates spermatogenesis. Nat. Rev. Endocrinol. 2010, 6, 380–395. [Google Scholar] [CrossRef] [PubMed]
- Rivkees, S.A.; Crawford, J.D. The relationship of gonadal activity and chemotherapy-induced gonadal damage. JAMA 1988, 259, 2123–2125. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, D.G.; Creemers, L.B.; den Ouden, K.; Izadyar, F. Spermatogonial stem cell development. In Testicular Tangrams, 1st ed.; Rommerts, F.F.G., Teerds, K.J., Eds.; Springer Press: Berlin/Heidelberg, Germany, 2002; pp. 121–138. [Google Scholar]
- Jahnukainen, K.; Ehmcke, J.; Hou, M.; Schlatt, S. Testicular function and fertility preservation in male cancer patients. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.A.; Mitchell, R.T.; Kelsey, T.W.; Spears, N.; Telfer, E.E.; Wallace, W.H. Cancer treatment and gonadal function: Experimental and established strategies for fertility preservation in children and young adults. Lancet Diabetes Endocrinol. 2015, 3, 556–567. [Google Scholar] [CrossRef]
- Stukenborg, J.B.; Jahnukainen, K.; Hutka, M.; Mitchell, R.T. Cancer treatment in childhood and testicular function: The importance of the somatic environment. Endocr. Connect. 2018, 7, 69–87. [Google Scholar] [CrossRef]
- Green, D.M.; Liu, W.; Kutteh, W.H.; Ke, R.W.; Shelton, K.C.; Sklar, C.A.; Chemaitilly, W.; Pui, C.H.; Klosky, J.L.; Spunt, S.L.; et al. Cumulative alkylating agent exposure and semen parameters in adult survivors of childhood cancer: A report from the St Jude Lifetime Cohort Study. Lancet Oncol. 2014, 15, 1215–1223. [Google Scholar] [CrossRef]
- Allen, C.M.; Lopes, F.; Mitchell, R.; Spears, N. How does chemotherapy treatment damage the prepubertal testis? Reproduction 2018, 156, 209–233. [Google Scholar] [CrossRef]
- Nurmio, M.; Keros, V.; Lähteenmäki, P.; Salmi, T.; Kallajoki, M.; Jahnukainen, K. Effect of childhood acute lymphoblastic leukemia therapy on spermatogonia populations and future fertility. J. Clin. Endocrinol. Metab. 2009, 94, 2119–2122. [Google Scholar] [CrossRef]
- Poganitsch-Korhonen, M.; Masliukaite, I.; Nurmio, M.; Lähteenmäki, P.; van Wely, M.; van Pelt, A.M.M.; Jahnukainen, K.; Stukenborg, J.B. Decreased spermatogonial quantity in prepubertal boys with leukemia treated with alkylating agents. Leukemia 2017, 31, 1460–1463. [Google Scholar] [CrossRef]
- Stukenborg, J.B.; Alves-Lopes, J.P.; Kurek, M.; Albalushi, H.; Reda, A.; Keros, V.; Töhönen, V.; Bjarnason, R.; Romerius, P.; Sundin, M.; et al. Spermatogonial quantity in human prepubertal testicular tissue collected for fertility preservation prior to potentially sterilizing therapy. Hum. Reprod. 2018, 33, 1677–1683. [Google Scholar] [CrossRef]
- Chow, E.J.; Stratton, K.L.; Leisenring, W.M.; Oeffinger, K.C.; Sklar, C.A.; Donaldson, S.S.; Ginsberg, J.P.; Kenney, L.B.; Levine, J.M.; Robison, L.L.; et al. Pregnancy after chemotherapy in male and female survivors of childhood cancer treated between 1970 and 1999: A report from the Childhood Cancer Survivor Study cohort. Lancet Oncol. 2016, 17, 567–576. [Google Scholar] [CrossRef]
- Sklar, C.A.; Robison, L.L.; Nesbit, M.E.; Sather, H.N.; Meadows, A.T.; Ortega, J.A.; Kim, T.H.; Hammond, G.D. Effects of radiation on testicular function in long-term survivors of childhood acute lymphoblastic leukemia: A report from the children cancer study group. J. Clin. Oncol. 1990, 8, 1981–1987. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.J.; Shalet, S.M. Spermatogenesis after cancer treatment: Damage and recovery. J. Natl. Cancer Inst. Monogr. 2005, 34, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Wyns, C.; Curaba, M.; Vanabelle, B.; van Langendonckt, A.; Donnez, J. Options for fertility preservation in prepubertal boys. Hum. Reprod. Update 2010, 16, 312–328. [Google Scholar] [CrossRef]
- Jahnukainen, K.; Heikkinen, R.; Henriksson, M.; Cooper, T.G.; Puukko-Viertomies, L.R.; Mäkitie, O. Semen quality and fertility in adult long-term survivors of childhood acute lymphoblastic leukemia. Fertil. Steril. 2011, 96, 837–842. [Google Scholar] [CrossRef]
- Byrne, J.; Fears, T.R.; Mills, J.L.; Zeltzer, L.K.; Sklar, C.A.; Meadows, A.T.; Reaman, G.H.; Robison, L.L. Fertility of long-term male survivors of acute lymphoblastic leukemia diagnosed during childhood. Pediatr. Blood Cancer 2004, 42, 364–372. [Google Scholar] [CrossRef]
- Meistrich, M.L. Effects of chemotherapy and radiotherapy on spermatogenesis in humans. Fertil. Steril. 2013, 100, 1180–1186. [Google Scholar] [CrossRef]
- Meistrich, M.L.; Wilson, G.; Brown, B.W.; de Cunha, M.F.; Lipschultz, L.I. Impact of cyclophosphamide on long-term reduction in sperm count in men treated with combination chemotherapy for ewing and soft tissue sarcomas. Cancer 1992, 70, 2703–2712. [Google Scholar] [CrossRef]
- Chung, K.; Irani, J.; Knee, G.; Efymow, B.; Blasco, L.; Patrizio, P. Sperm cryopreservation for male patients with cancer: An epidemiological analysis at the University of Pennsylvania. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 113, 7–11. [Google Scholar] [CrossRef]
- Kliesch, S.; Behre, H.M.; Jurgens, H.; Nieschlag, E. Cryopreservation of semen from adolescent patients with malignancies. Med. Pediatr. Oncol. 1996, 26, 20–27. [Google Scholar] [CrossRef]
- Muller, J.; Sonksen, J.; Sommer, P.; Schmiegelow, M.; Petersen, P.M.; Heilman, C.; Schmiegelow, K. Cryopreservation of semen from pubertal boys with cancer. Med. Pediatr. Oncol. 2000, 34, 191–194. [Google Scholar] [CrossRef]
- Bahadur, G.; Ling, K.L.; Hart, R.; Ralph, D.; Wafa, R.; Ashraf, A.; Jaman, N.; Mahmud, S.; Oyede, A.W. Semen production in adolescent cancer patients. Hum. Reprod. 2002, 17, 2654–2656. [Google Scholar] [CrossRef] [PubMed]
- Glaser, A.W.; Phelan, L.; Crawshaw, M.; Jagdev, S.; Haleet, J. Fertility preservation in adolescent males with cancer in the United Kingdom: A survey of practice. Arch. Dis. Child. 2004, 89, 736–737. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Velez de la Calle, J.F.; Jégou, B. Protection by steroid contraceptives against procarbazine-induced sterility and genotoxicity in male rats. Cancer Res. 1990, 50, 1308–1315. [Google Scholar] [PubMed]
- Udagawa, K.; Ogawa, T.; Watanabe, T.; Yumura, Y.; Takeda, M.; Hosaka, M. GnRH analog, leuprorelin acetate, promotes regeneration of rat spermatogenesis after severe chemical damage. Int. J. Urol. 2001, 8, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Kangasniemi, M.; Dodge, K.; Pemberton, A.E.; Huhtaniemi, I.; Meistrich, M.L. Suppression of mouse spermatogenesis by a gonadotropin-releasing hormone antagonist and antiandrogen: Failure to protect against radiation-induced gonadal damage. Endocrinology 1996, 137, 949–955. [Google Scholar] [CrossRef]
- Meistrich, M.L.; Shetty, G. Hormonal suppression for fertility preservation in males and females. Reproduction 2008, 136, 691–701. [Google Scholar] [CrossRef]
- Shetty, G.; Uthamanthil, R.K.; Zhou, W.; Shao, S.H.; Weng, C.C.; Tailor, R.C.; Hermann, B.P.; Orwig, K.E.; Meistrich, M.L. Hormone suppression with GnRH antagonist promotes spermatogenic recovery from transplanted spermatogonial stem cells in irradiated cynomolgus monkeys. Andrology 2013, 1, 886–898. [Google Scholar] [CrossRef]
- Shetty, G.; Meistrich, M.L. Hormonal approaches to preservation and restoration of male fertility after cancer treatment. J. Natl. Cancer. Inst. Monogr. 2005, 34, 36–39. [Google Scholar] [CrossRef]
- Boekelheide, K.; Schoenfeld, H.A.; Hall, S.J.; Weng, C.C.; Shetty, G.; Leith, J.; Harper, J.; Sigman, M.; Hess, D.L.; Meistrich, M.L. Gonadotropin-releasing hormone antagonist (cetrorelix) therapy fails to protect nonhuman primates (Macaca arctoides) from radiation-induced spermatogenic failure. J. Androl. 2005, 26, 222–234. [Google Scholar] [CrossRef]
- Kamischke, A.; Kuhlmann, M.; Weinbauer, G.; Luetjens, M.; Yeung, C.; Kronholz, H.; Nieschlag, E. Gonadal protection from radiation by GnRH antagonist or recombinant human FSH: A controlled trial in a male nonhuman primate (Macaca fascicularis). J. Endocrinol. 2003, 179, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.R.; Fraser, H.M. The neonatal period: A critical interval in male primate development. J. Endocrinol. 1996, 149, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Benavides-Garcia, R.; Joachim, R.; Pina, N.A.; Mutoji, K.N.; Reilly, M.A.; Hermann, B.P. Granulocyte colony-stimulating factor prevents loss of spermatogenesis after sterilizing busulfan chemotherapy. Fertil. Steril. 2015, 103, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Kotzur, T.; Benavides-Garcia, R.; Mecklenburg, J.; Sanchez, J.R.; Reilly, M.; Hermann, B.P. Granulocyte colony-stimulating factor (G-CSF) promotes spermatogenic regeneration from surviving spermatogonia after high-dose alkylating chemotherapy. Reprod. Biol. Endocrinol. 2017, 15, 7. [Google Scholar] [CrossRef]
- Rajesh Kumar, T.; Doreswamy, K.; Shrilatha, B.; Muralidhara. Oxidative stress associated DNA damage in testis of mice: Induction of abnormal sperms and effects on fertility. Mutat. Res. Toxicol. Environ. Mutagen. 2002, 513, 103–111. [Google Scholar] [CrossRef]
- Ilbey, Y.O.; Ozbek, E.; Simsek, A.; Otunctemur, A.; Cekmen, M.; Somay, A. Potential chemoprotective effect of melatonin in cyclophosphamide- and cisplatin-induced testicular damage in rats. Fertil. Steril. 2009, 92, 1124–1132. [Google Scholar] [CrossRef]
- Probin, V.; Wang, Y.; Zhou, D. Busulfan-induced senescence is dependent on ROS production upstream of the MAPK pathway. Free Radic. Biol. Med. 2007, 42, 1858–1865. [Google Scholar] [CrossRef]
- Goldman, J.M. Chronic Myeloid leukemia: A historical perspective. Semin. Hematol. 2010, 47, 302–311. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, Q.; Wei, R.; Song, H.; Mi, J.; Lin, Z.; Yang, Y.; Sun, Z.; Zou, K. Melatonin protects spermatogonia from the stress of chemotherapy and oxidation via eliminating reactive oxidative species. Free Radic. Biol. Med. 2019, 137, 74–86. [Google Scholar] [CrossRef]
- Büyükavci, M.; Ozdemir, O.; Buck, S.; Ravindranath, Y.; Savaşan, S. Effect of melatonin on the cytotoxicity of chemotherapeutic drugs in human leukemia cells. In Vivo 2011, 25, 405–409. [Google Scholar]
- Rodriguez, C.; Martín, V.; Herrera, F.; García-Santos, G.; Rodriguez-Blanco, J.; Casado-Zapico, S.; Sánchez-Sánchez, A.M.; Suárez, S.; Puente-Moncada, N.; Anítua, M.J.; et al. 1. Mechanisms involved in the pro-apoptotic effect of melatonin in cancer cells. Int. J. Mol. Sci. 2013, 14, 6597–6613. [Google Scholar] [CrossRef] [PubMed]
- Carmely, A.; Meirow, D.; Peretz, A.; Albeck, M.; Bartoov, B.; Sredni, B. Protective effect of the immunomodulator AS101 against cyclophosphamide-induced testicular damage in mice. Hum. Reprod. 2009, 24, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.Y.; Cheng, C.C.; Chiang, Y.S.; Cheng, W.T.K.; Liu, I.H.; Wu, S.C. Exosomal miR-10a derived from amniotic fluid stem cells preserves ovarian follicles after chemotherapy. Sci. Rep. 2016, 6, 23120. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; He, Y.; Wang, X.; Peng, D.; Chen, X.; Li, X.; Wang, Q. Overexpression of miR-21 in stem cells improves ovarian structure and function in rats with chemotherapy-induced ovarian damage by targeting PDCD4 and PTEN to inhibit granulosa cell apoptosis. Stem Cell Res. Ther. 2017, 8, 187. [Google Scholar] [CrossRef]
- Alexandri, C.; Stamatopoulos, B.; Rothé, F.; Bareche, Y.; Devos, M.; Demeestere, I. MicroRNA profiling and identification of let-7a as a target to prevent chemotherapy-induced primordial follicles apoptosis in mouse ovaries. Sci. Rep. 2019, 9, 9636. [Google Scholar] [CrossRef]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Biophys. Acta 2010, 1803, 1231–1243. [Google Scholar] [CrossRef]
- Reza, A.M.M.T.; Choi, Y.J.; Han, S.G.; Song, H.; Park, C.; Hong, K.; Kim, J.H. Roles of microRNAs in mammalian reproduction: From the commitment of germ cells to peri-implantation embryos. Biol. Rev. 2019, 94, 415–438. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA–gene interactions. Nucleic Acids Res. 2018, 46, 239–245. [Google Scholar] [CrossRef]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, 296–302. [Google Scholar] [CrossRef]
- Akinjo, O.O.; Gant, T.W.; Marczylo, E.L. Perturbation of microRNA signalling by doxorubicin in spermatogonial, Leydig and Sertoli cell lines in vitro. Toxicol. Res. 2018, 7, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, F.H.; Li, Y.; Wang, Z.; Kong, D.; Ali, S. Implication of microRNAs in drug resistance for designing novel cancer therapy. Drug Resist. Update 2010, 13, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Lian, J.; Zhang, H.; Tian, H.; Liang, M.; Yin, M.; Sun, F. MicroRNA-302a sensitizes testicular embryonal carcinoma cells to cisplatin-induced cell death. J. Cell Physiol. 2013, 228, 2294–2304. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Liu, Y.; Chen, L.; Ellis, M.; Li, M.; Wang, J.; Zhang, Y.; Fu, P.; Wang, K.; Li, X.; et al. microRNA profiling in three main stages during porcine spermatogenesis. J. Assist. Reprod. Genet. 2015, 32, 451–460. [Google Scholar] [CrossRef]
- Novotny, G.W.; Sonne, S.B.; Nielsen, J.E.; Jonstrup, S.P.; Hansen, M.A.; Skakkebaek, N.E.; Rajpert-De Meyts, E.; Kjems, J.; Leffers, H. Translational repression of E2F1 mRNA in carcinoma in situ and normal testis correlates with expression of the miR-17-92 cluster. Cell Death Differ. 2007, 14, 879–882. [Google Scholar] [CrossRef]
- Xie, R.; Lin, X.; Du, T.; Xu, K.; Shen, H.; Wei, F.; Hao, W.; Lin, T.; Lin, X.; Qin, Y.; et al. Targeted disruption of miR-17-92 impairs mouse spermatogenesis by activating mTOR signaling pathway. Medicine 2016, 95, 2713. [Google Scholar] [CrossRef]
- Meunier, L.; Siddeek, B.; Vega, A.; Lakhdari, N.; Inoubli, L.; Bellon, R.P.; Lemaire, G.; Mauduit, C.; Benahmed, M. Perinatal programming of adult rat germ cell death after exposure to xenoestrogens: Role of microRNA miR-29 family in the down-regulation of DNA methyltransferases and Mcl-1. Endocrinology 2012, 153, 1936–1947. [Google Scholar] [CrossRef]
- Chen, R.; Deng, L.; Yu, X.; Wang, X.; Zhu, L.; Yu, T.; Zhang, Y.; Zhou, B.; Xu, W.; Chen, L.; et al. MiR-122 partly mediates the ochratoxin A-induced GC-2 cell apoptosis. Toxicol. Vitr. 2015, 30, 264–273. [Google Scholar] [CrossRef]
- Guan, Y.; Liang, G.; Hawken, P.A.R.; Malecki, I.A.; Cozens, G.; Vercoe, P.E.; Martin, G.B.; Guan, L.L. Roles of small RNAs in the effects of nutrition on apoptosis and spermatogenesis in the adult testis. Sci. Rep. 2015, 5, 10372. [Google Scholar] [CrossRef]
- Bao, J.; Li, D.; Wang, L.; Wu, J.; Hu, Y.; Wang, Z.; Chen, Y.; Cao, X.; Jiang, C.; Yan, W.; et al. MicroRNA-449 and MicroRNA-34b/c function redundantly in murine testes by targeting E2F transcription factor-retinoblastoma protein (E2F-pRb) pathway. J. Biol. Chem. 2012, 287, 21686–21698. [Google Scholar] [CrossRef]
- Smorag, L.; Zheng, Y.; Nolte, J.; Zechner, U.; Engel, W.; Pantakani, D.V.K. MicroRNA signature in various cell types of mouse spermatogenesis: Evidence for stage-specifically expressed miRNA-221, -203 and -34b-5p mediated spermatogenesis regulation. Biol. Cell 2012, 104, 677–692. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.; Wu, R.; Liu, B.; Dong, W.; Tu, Z.; Yang, J.; Xu, Z.; Pan, T. Upstream and downstream mechanisms for the promoting effects of IGF-1 on differentiation of spermatogonia to primary spermatocytes. Life Sci. 2014, 101, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Niu, M.; Yao, C.; Hai, Y.; Yuan, Q.; Liu, Y.; Guo, Y.; Li, Z.; He, Z. Fractionation of human spermatogenic cells using STA-PUT gravity sedimentation and their miRNA profiling. Sci. Rep. 2015, 5, 8084. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Raabe, T.; Hecht, N.B. MicroRNA Mirn122a reduces expression of the post-transcriptionally regulated germ cell transition protein 2 (Tnp2) messenger RNA (mRNA) by mRNA cleavage1. Biol. Reprod. 2005, 73, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Qing, X.; Shi, J.; Dong, T.; Wu, C.; Hu, L.; Li, H. Dysregulation of an X-linked primate-specific epididymal microRNA cluster in unexplained asthenozoospermia. Oncotarget 2017, 8, 56839–56849. [Google Scholar] [CrossRef]
- Fon Tacer, K.; Montoya, M.C.; Oatley, M.J.; Lord, T.; Oatley, J.M.; Klein, J.; Ravichandran, R.; Tillman, H.; Kim, M.; Connelly, J.P.; et al. MAGE cancer-testis antigens protect the mammalian germline under environmental stress. Sci. Adv. 2019, 5, 4832. [Google Scholar] [CrossRef] [PubMed]
- Cremades, N.; Bernabeu, R.; Barros, A.; Sousa, M. In-vitro maturation of round spermatids using co-culture on Vero cells. Hum. Reprod. 1999, 14, 1287–1293. [Google Scholar] [CrossRef]
- Cremades, N.; Sousa, M.; Bernabeu, R.; Barros, A. Developmental potential of elongating and elongated spermatids obtained after in-vitro maturation of isolated round spermatids. Hum. Reprod. 2001, 16, 1938–1944. [Google Scholar] [CrossRef]
- Sousa, M.; Cremades, N.; Alves, C.; Silva, J.; Barros, A. Developmental potential of human spermatogenic cells co-cultured with Sertoli cells. Hum. Reprod. 2002, 17, 161–172. [Google Scholar] [CrossRef]
- Riboldi, M.; Rubio, C.; Pellicer, A.; Gil-Salom, M.; Simon, C. In vitro production of haploid cells after coculture of CD49f+ with Sertoli cells from testicular sperm extraction in nonobstructive azoospermic patients. Fertil. Steril. 2012, 98, 580–590. [Google Scholar] [CrossRef]
- Yang, S.; Ping, P.; Ma, M.; Li, P.; Tian, R.; Yang, H.; Liu, Y.; Gong, Y.; Zhang, Z.; Li, Z.; et al. Generation of haploid spermatids with fertilization and development capacity from human spermatogonial stem cells of cryptorchid patients. Stem Cell Rep. 2014, 3, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.R.; Kim, K.S.; Yang, Y.H.; Oh, H.S.; Lee, S.H.; Chung, T.G.; Cho, J.H.; Kim, H.J.; Yoon, T.K.; Cha, K.Y. Isolation of male germ stem cell-like cells from testicular tissue of non-obstructive azoospermic patients and differentiation into haploid male germ cells in vitro. Hum. Reprod. 2006, 21, 471–476. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stukenborg, J.B.; Wistuba, J.; Luetjens, C.M.; Elhija, M.A.; Huleihel, M.; Lunenfeld, E.; Gromoll, J.; Nieschlag, E.; Schlatt, S. Coculture of spermatogonia with somatic cells in a novel three-dimensional soft-agar-culture-system. J. Androl. 2008, 29, 312–329. [Google Scholar] [CrossRef] [PubMed]
- Stukenborg, J.B.; Schlatt, S.; Simoni, M.; Yeung, C.H.; Elhija, M.A.; Luetjens, C.M.; Huleihel, M.; Wistuba, J. New horizons for in vitro spermatogenesis? An update on novel three-dimensional culture systems as tools for meiotic and post-meiotic differentiation of testicular germ cells. Mol. Hum. Reprod. 2009, 15, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Baert, Y.; De Kock, J.; Alves-Lopes, J.P.; Söder, O.; Stukenborg, J.B.; Goossens, E. Primary human testicular cells self-organize into organoids with testicular properties. Stem Cell Rep. 2017, 10, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Alves-Lopes, J.P.; Soder, O.; Stukenborg, J.B. Testicular organoid generation by a novel in vitro three-layer gradient system. Biomaterials 2017, 130, 76–89. [Google Scholar] [CrossRef]
- Pendergraft, S.S.; Sadri-Ardekani, H.; Atala, A.; Bishop, C.E. Three-dimensional testicular organoid: A novel tool for the study of human spermatogenesis and gonadotoxicity in vitro. Biol. Reprod. 2017, 96, 720–732. [Google Scholar] [CrossRef]
- Sakib, S.; Uchida, A.; Valenzuela-Leon, P.; Yu, Y.; Valli-Pulaski, H.; Orwig, K.; Ungrin, M.; Dobrinski, I. Formation of organotypic testicular organoids in microwell culture. Biol. Reprod. 2019, 100, 1648–1660. [Google Scholar] [CrossRef]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y.; Ogawa, T. In vitro production of functional sperm in cultured neonatal mouse testes. Nature 2011, 24, 504–507. [Google Scholar] [CrossRef]
- Sato, T.; Katagiri, K.; Kubota, Y.; Ogawa, T. In vitro sperm production from mouse spermatogonial stem cell lines using an organ culture method. Nat. Protoc. 2013, 8, 2098–2104. [Google Scholar] [CrossRef]
- Yokonishi, T.; Sato, T.; Komeya, M.; Katagiri, K.; Kubota, Y.; Nakabayashi, K.; Hata, K.; Inoue, K.; Ogonuki, N.; Ogura, A.; et al. Offspring production with sperm grown in vitro from cryopreserved testis tissues. Nat. Commun. 2014, 5, 4320. [Google Scholar] [CrossRef] [PubMed]
- De Michele, F.; Poels, J.; Weerens, L.; Petit, C.; Evrard, Z.; Ambroise, J.; Gruson, D.; Wyns, C. Preserved seminiferous tubule integrity with spermatogonial survival and induction of Sertoli and Leydig cell maturation after long-term organotypic culture of prepubertal human testicular tissue. Hum. Reprod. 2017, 32, 32–45. [Google Scholar] [CrossRef] [PubMed]
- De Michele, F.; Poels, J.; Vermeulen, M.; Ambroise, J.; Gruson, D.; Guiot, Y.; Wyns, C. Haploid germ cells generated in organotypic culture of testicular tissue from prepubertal boys. Front Physiol. 2018, 9, 1413. [Google Scholar] [CrossRef] [PubMed]
- Baert, Y.; Dvorakova-Hortova, K.; Margaryan, H.; Goossens, E. Mouse in vitro spermatogenesis on alginate-based 3D bioprinted scaffolds. Biofabrication 2019, 11, 035011. [Google Scholar] [CrossRef]
- Laronda, M.M.; Rutz, A.L.; Xiao, S.; Whelan, K.A.; Duncan, F.E.; Roth, E.W.; Woodruff, T.K.; Shah, R.N. A bioprosthetic ovary created using 3D printed microporous scaffolds restores ovarian function in sterilized mice. Nat. Commun. 2017, 8, 15261. [Google Scholar] [CrossRef]
- Brinster, R.L.; Avarbock, M.R. Germline transmission of donor haplotype following spermatogonial transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11303–11307. [Google Scholar] [CrossRef]
- Brinster, R.L.; Zimmermann, J.W. Spermatogenesis following male germ-cell transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11298–11302. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Inoue, K.; Ogura, A.; Toyokuni, S.; Honjo, T.; Shinohara, T. Allogeneic offspring produced by male germ line stem cell transplantation into infertile mouse testis. Biol. Reprod. 2003, 68, 167–173. [Google Scholar] [CrossRef]
- Goossens, E.; Frederickx, V.; de Block, G.; van Steirteghem, A.; Tournaye, H. Evaluation of in vivo conception after testicular stem cell transplantation in a mouse model shows altered post-implantation development. Hum. Reprod. 2006, 21, 2057–2060. [Google Scholar] [CrossRef]
- Wu, X.; Goodyear, S.M.; Abramowitz, L.K.; Bartolomei, M.S.; Tobias, J.W.; Avarbock, M.R.; Brinster, R.L. Fertile offspring derived from mouse spermatogonial stem cells cryopreserved for more than 14 years. Hum. Reprod. 2012, 27, 1249–1259. [Google Scholar] [CrossRef]
- Valli, H.; Phillips, B.T.; Shetty, G.; Byrne, J.A.; Clark, A.T.; Meistrich, M.L.; Orwig, K.E. Germline stem cells: Toward the regeneration of spermatogenesis. Fertil. Steril. 2014, 101, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Hermann, B.P.; Sukhwani, M.; Winkler, F.; Pascarella, J.N.; Peters, K.A.; Sheng, Y.; Valli, H.; Rodriguez, M.; Ezzelarab, M.; Dargo, G.; et al. Spermatogonial stem cell transplantation into Rhesus testes regenerates spermatogenesis producing functional sperm. Cell Stem Cell 2012, 11, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Radford, J. Restoration of fertility after treatment for cancer. Horm. Res. Paediatr. 2003, 59, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Tegelenbosch, R.A.; de Rooij, D.G. A quantitative study of spermatogonial multiplication and stem cell renewal in the C3H/101 F1 hybrid mouse. Mutat. Res. 1993, 290, 193–200. [Google Scholar] [CrossRef]
- Hermann, B.P.; Sukhwani, M.; Simorangkir, D.R.; Chu, T.; Plant, T.M.; Orwig, K.E. Molecular dissection of the male germ cell lineage identifies putative spermatogonial stem cells in rhesus macaques. Hum. Reprod. 2009, 24, 1704–1716. [Google Scholar] [CrossRef]
- Nagano, M.; Avarbock, M.R.; Brinster, R.L. Pattern and kinetics of mouse donor spermatogonial stem cell colonization in recipient testes. Biol. Reprod. 1999, 60, 1429–1436. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Inoue, K.; Miki, H.; Ogura, A.; Toyokuni, S.; Shinohara, T. Long-term proliferation in culture and germline transmission of mouse male germline stem cells. Biol. Reprod. 2003, 69, 612–616. [Google Scholar] [CrossRef]
- Sadri-Ardekani, H.; Mizrak, S.C.; van Daalen, S.K.M.; Korver, C.M.; Roepers-Gajadien, H.L.; Koruji, M.; Hovingh, S.; de Reijke, T.M.; de la Rosette, J.J.; van der Veen, F.; et al. Propagation of human spermatogonial stem cells in vitro. Jama 2009, 302, 2127–2134. [Google Scholar] [CrossRef]
- Sadri-Ardekani, H.; Akhondi, M.A.; van der Veen, F.; Repping, S.; van Pelt, A.M. In vitro propagation of human prepubertal spermatogonial stem cells. Jama 2011, 305, 2416–2418. [Google Scholar] [CrossRef]
- Kadam, P.; Ntemou, E.; Baert, Y.; van Laere, S.; van Saen, D.; Goossens, E. Co-transplantation of mesenchymal stem cells improves spermatogonial stem cell transplantation efficiency in mice. Stem Cell Res. Ther. 2018, 9, 317. [Google Scholar] [CrossRef]
- Goossens, E.; de Vos, P.; Tournaye, H. Array comparative genomic hybridization analysis does not show genetic alterations in spermatozoa and offspring generated after spermatogonial stem cell transplantation in the mouse. Hum. Reprod. 2010, 25, 1836–1842. [Google Scholar] [CrossRef] [PubMed]
- Kadam, P.; Ntemou, E.; Onofre, J.; van Saen, D.; Goossens, E. Does co-transplantation of mesenchymal and spermatogonial stem cells improve reproductive efficiency and safety in mice? Stem Cell Res. Ther. in press.
- Mulder, C.L.; Catsburg, L.A.E.; Zheng, Y.; de Winter-Korver, C.M.; van Daalen, S.K.M.; van Wely, M.; Pals, S.; Repping, S.; van Pelt, A.M.M. Long-term health in recipients of transplanted in vitro propagated spermatogonial stem cells. Hum. Reprod. 2018, 33, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Jahnukainen, K.; Hou, M.; Petersen, C.; Setchell, B.; Söder, O. Intratesticular transplantation of testicular cells from leukemic rats causes transmission of leukemia. Cancer Res. 2001, 61, 706–710. [Google Scholar]
- Geens, M.; van de Velde, H.; De Block, G.; Goossens, E.; van Steirteghem, A.; Tournaye, H. The efficiency of magnetic-activated cell sorting and fluorescence-activated cell sorting in the decontamination of testicular cell suspensions in cancer patients. Hum. Reprod. 2007, 22, 733–742. [Google Scholar] [CrossRef]
- Geens, M.; Goossens, E.; Tournaye, H. Cell selection by selective matrix adhesion is not sufficiently efficient for complete malignant cell depletion from contaminated human testicular cell suspensions. Fertil. Steril. 2011, 95, 787–791. [Google Scholar] [CrossRef]
- Dovey, S.L.; Valli, H.; Hermann, B.P.; Sukhwani, M.; Donohue, J.; Castro, C.A.; Chu, T.; Sanfilippo, J.S.; Orwig, K.E. Eliminating malignant contamination from therapeutic human spermatogonial stem cells. J. Clin. Invest. 2013, 123, 1833–1843. [Google Scholar] [CrossRef]
- Sadri-Ardekani, H.; Homburg, C.H.; van Capel, T.M.M.; van den Berg, H.; van der Veen, F.; van der Schoot, C.E.; van Pelt, A.M.; Repping, S. Eliminating acute lymphoblastic leukemia cells from human testicular cell cultures: A pilot study. Fertil. Steril. 2014, 101, 1072–1078. [Google Scholar] [CrossRef]
- Demeestere, I.; Simon, P.; Dedeken, L.; Moffa, F.; Tsépélidis, S.; Brachet, C.; Delbaere, A.; Devreker, F.; Ferster, A. Live birth after autograft of ovarian tissue cryopreserved during childhood. Hum. Reprod. 2015, 30, 2107–2109. [Google Scholar] [CrossRef]
- Gellert, S.E.; Pors, S.E.; Kristensen, S.G.; Bay-Bjørn, A.M.; Ernst, E.; Yding Andersen, C. Transplantation of frozen-thawed ovarian tissue: An update on worldwide activity published in peer-reviewed papers and on the Danish cohort. J. Assist. Reprod. Genet. 2018, 35, 561–570. [Google Scholar] [CrossRef]
- Honaramooz, A.; Snedaker, A.; Boiani, M.; Schöler, H.; Dobrinski, I.; Schlatt, S. Sperm from neonatal mammalian testes grafted in mice. Nature 2002, 418, 778–781. [Google Scholar] [CrossRef] [PubMed]
- Schlatt, S.; Honaramooz, A.; Boiani, M.; Scholer, H.R.; Dobrinski, I. Progeny from sperm obtained after ectopic grafting of neonatal mouse testes. Biol. Reprod. 2003, 68, 2331–2335. [Google Scholar] [CrossRef] [PubMed]
- Arregui, L.; Dobrinski, I. Xenografting of testicular tissue pieces: Twelve years of an in vivo spermatogenesis system. Reproduction 2014, 148, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Luetjens, C.M.; Stukenborg, J.B.; Nieschlag, E.; Simoni, M.; Wistuba, J. Complete spermatogenesis in orthotopic but not in ectopic transplants of autologously grafted marmoset testicular tissue. Endocrinology 2008, 149, 1736–1747. [Google Scholar] [CrossRef]
- Jahnukainen, K.; Ehmcke, J.; Nurmio, M.; Schlatt, S. Autologous ectopic grafting of cryopreserved testicular tissue preserves the fertility of prepubescent monkeys that receive sterilizing cytotoxic therapy. Cancer Res. 2012, 72, 5174–5178. [Google Scholar] [CrossRef]
- Fayomi, A.P.; Peters, K.; Sukhwani, M.; Valli-Pulaski, H.; Shetty, G.; Meistrich, M.L.; Houser, L.; Robertson, N.; Roberts, V.; Ramsey, C.; et al. Autologous grafting of cryopreserved prepubertal rhesus testis produces sperm and offspring. Science 2019, 363, 1314–1319. [Google Scholar] [CrossRef]
- Marcial-Rojas, A.; Castro, J.R. Irradiation injury to elastic arteries in the course of treatment for neoplastic disease. Ann. Otol. Rhinol. Laryngol. 1962, 71, 945–958. [Google Scholar] [CrossRef]
- Poels, J.; Abou-Ghannam, G.; Decamps, A.; Leyman, M.; Des Rieux, A.; Wyns, C. Transplantation of testicular tissue in alginate hydrogel loaded with VEGF nanoparticles improves spermatogonial recovery. J. Control. Release 2016, 234, 79–89. [Google Scholar] [CrossRef]
- Ntemou, E.; Kadam, P.; van Laere, S.; van Saen, D.; Vicini, E.; Goossens, E. Effect of recombinant human vascular endothelial growth factor on testis tissue xenotransplants from prepubertal boys: A three-case study. Reprod. Biomed. Online 2019, 39, 119–133. [Google Scholar] [CrossRef]
- Wyns, C.; van Langendonckt, A.; Wese, F.H.; Donnez, J.; Curaba, M. Long-term spermatogonial survival in cryopreserved and xenografted immature human testicular tissue. Hum. Reprod. 2008, 23, 2402–2414. [Google Scholar] [CrossRef]
- Sato, Y.; Nozawa, S.; Yoshiike, M.; Arai, M.; Sasaki, C.; Iwamoto, T. Xenografting of testicular tissue from an infant human donor results in accelerated testicular maturation. Hum. Reprod. 2010, 25, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Van Saen, D.; Goossens, E.; Bourgain, C.; Ferster, A.; Tournaye, H. Meiotic activity in orthotopic xenografts derived from human postpubertal testicular tissue. Hum. Reprod. 2011, 26, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Van Saen, D.; Goossens, E.; Haentjens, P.; Baert, Y.; Tournaye, H. Exogenous administration of recombinant human FSH does not improve germ cell survival in human prepubertal xenografts. Reprod. Biomed. Online 2013, 26, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Poels, J.; van Langendonckt, A.; Many, M.C.; Wese, F.X.; Wyns, C. Vitrification preserves proliferation capacity in human spermatogonia. Hum. Reprod. 2013, 28, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Ntemou, E.; Kadam, P.; van Saen, D.; Wistuba, J.; Mitchell, R.T.; Schlatt, S.; Goossens, E. Complete spermatogenesis in intratesticular testis tissue xenotransplants from immature non-human primate. Hum. Reprod. 2019, 34, 403–413. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, R.; O’Donnell, L.; Meachem, S.J.; Stanton, P.G.; de Kretser, D.M.; Pratis, K.; Robertson, D.M. Hormonal regulation of spermatogenesis in primates and man: Insights for development of the male hormonal contraceptive. J. Androl. 2002, 23, 149–162. [Google Scholar]
- Smith, L.B.; Walker, W.H. The regulation of spermatogenesis by androgens. Semin. Cell Dev. Biol. 2014, 30, 2–13. [Google Scholar] [CrossRef]
- Amann, R.P. The cycle of the seminiferous epithelium in humans: A need to revisit? J. Androl. 2008, 29, 469–487. [Google Scholar] [CrossRef]
- Honaramooz, A.; Li, M.W.; Penedo, M.C.; Meyers, S.; Dobrinski, I. Accelerated maturation of primate testis by xenografting into mice. Biol. Reprod. 2004, 70, 1500–1503. [Google Scholar] [CrossRef]
- Liu, Z.; Nie, Y.H.; Zhang, C.C.; Cai, Y.J.; Wang, Y.; Lu, H.P.; Li, Y.Z.; Cheng, C.; Qiu, Z.L.; Sun, Q. Generation of macaques with sperm derived from juvenile monkey testicular xenografts. Cell Res. 2016, 26, 139–142. [Google Scholar] [CrossRef]
- Goossens, E.; Bilgec, T.; van Saen, D.; Tournaye, H. Mouse germ cells go through typical epigenetic modifications after intratesticular tissue grafting. Hum. Reprod. 2011, 26, 3388–3400. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNAs in Male Germline Apoptosis Regulation | ||||
|---|---|---|---|---|
| miRNA | Targeted Genes | Function | Species | Reference |
| miR-16 | Ccnd1 | Apoptosis induction | pig | [76] |
| miR-17-92 | c-MYC, E2F1 | Downregulation leads to testicular atrophy, apoptosis induction and germ cell free seminiferous tubules | mouse | [77,78] |
| miR-29 | Dnmt, Mcl-1 | Extensive germ cells apoptosis | rat | [79] |
| miR-122 | Unknown | Inhibition minimizes ochratoxin-A-toxicity | spermatocyte-like cell line | [80] |
| miR-144 | FASL, CAS3, TP53, BCL2L1 | Regulates apoptosis-related genes, apoptosis induction | sheep | [81] |
| miR-449, miR-34b/c | E2F-pRb pathway | Acts redundantly to suppress E2F-pRb pathway during the meiotic phase of spermatogenesis | mouse | [82] |
| miR-34 | Genes involved in cell cycle, apoptosis and growth factor signaling | Regulates cell cycle spermatogenesis progression and sperm senescence | pig | [76] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntemou, E.; Alexandri, C.; Lybaert, P.; Goossens, E.; Demeestere, I. Oncofertility: Pharmacological Protection and Immature Testicular Tissue (ITT)-Based Strategies for Prepubertal and Adolescent Male Cancer Patients. Int. J. Mol. Sci. 2019, 20, 5223. https://doi.org/10.3390/ijms20205223
Ntemou E, Alexandri C, Lybaert P, Goossens E, Demeestere I. Oncofertility: Pharmacological Protection and Immature Testicular Tissue (ITT)-Based Strategies for Prepubertal and Adolescent Male Cancer Patients. International Journal of Molecular Sciences. 2019; 20(20):5223. https://doi.org/10.3390/ijms20205223
Chicago/Turabian StyleNtemou, Elissavet, Chrysanthi Alexandri, Pascale Lybaert, Ellen Goossens, and Isabelle Demeestere. 2019. "Oncofertility: Pharmacological Protection and Immature Testicular Tissue (ITT)-Based Strategies for Prepubertal and Adolescent Male Cancer Patients" International Journal of Molecular Sciences 20, no. 20: 5223. https://doi.org/10.3390/ijms20205223
APA StyleNtemou, E., Alexandri, C., Lybaert, P., Goossens, E., & Demeestere, I. (2019). Oncofertility: Pharmacological Protection and Immature Testicular Tissue (ITT)-Based Strategies for Prepubertal and Adolescent Male Cancer Patients. International Journal of Molecular Sciences, 20(20), 5223. https://doi.org/10.3390/ijms20205223