The miR396–GRF Regulatory Module Controls the Embryogenic Response in Arabidopsis via an Auxin-Related Pathway


Department of Genetics, University of Silesia, Faculty of Biology and Environmental Protection, 40-032 Katowice, Poland
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(20), 5221; https://doi.org/10.3390/ijms20205221
Submission received: 30 September 2019
/
Accepted: 18 October 2019
/
Published: 21 October 2019
(This article belongs to the Special Issue Transcriptional and Post-transcriptional Gene Regulation in Plants)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:In plants, microRNAs have been indicated to control various developmental processes, including somatic embryogenesis (SE), which is triggered in the in vitro cultured somatic cells of plants. Although a transcriptomic analysis has indicated that numerous MIRNAs are differentially expressed in the SE of different plants, the role of specific miRNAs in the embryogenic reprogramming of the somatic cell transcriptome is still poorly understood. In this study, we focused on performing a functional analysis of miR396 in SE given that the transcripts of MIR396 genes and the mature molecules of miR396 were found to be increased during an SE culture of Arabidopsis. In terms of miR396 in embryogenic induction, we observed the SE-associated expression pattern of MIR396b in explants of the β-glucuronidase (GUS) reporter line. In order to gain insight into the miR396-controlled mechanism that is involved in SE induction, the embryogenic response of mir396 mutants and the 35S:MIR396b overexpressor line to media with different 2,4-Dichlorophenoxyacetic acid (2,4-D) concentrations was evaluated. The results suggested that miR396 might contribute to SE induction by controlling the sensitivity of tissues to auxin treatment. Within the targets of miR396 that are associated with SE induction, we identified genes encoding the GROWTH-REGULATING FACTOR (GRF) transcription factors, including GRF1, GRF4, GRF7, GRF8, and GRF9. Moreover, the study suggested a regulatory relationship between miR396, GRF, and the PLETHORA (PLT1 and PLT2) genes during SE induction. A complex regulatory relationship within the miR396–GRF1/4/8/9–PLT1/2 module that involves the negative and positive control of GRFs and PLT (respectively) by miR396 might be assumed.
1. Introduction
Somatic embryogenesis (SE), a unique plant-specific developmental process, results in the formation of somatic embryos from in vitro cultured somatic cells/tissue. Since its discovery, SE has become widely used for plant regeneration in plant biotechnology and as a model to study the totipotency of plant somatic cells in basic research. The knowledge that is gained from studies on SE and, in particular, from identifying the genetic determinants of embryogenic transition in in vitro cultured plant explants greatly contributes to improving the micropropagation and genetic transformation of plants (reviewed in Reference [1]).
Transcription factors (TFs), which play a central role in regulating gene expression, are believed to have a decisive function in the genetic reprogramming of somatic cells in SE. Accordingly, an extensive modulation of hundreds of TF genes was found during SE induction in Arabidopsis [2,3] and the function of several TFs in SE induction was demonstrated including, BABY BOOM (BBM [4]), LEAFY COTYLEDON2 (LEC2 [5]), WUSCHEL (WUS [6]), AGAMOUS-LIKE15 (AGL15 [7]), MYB118 [8], and EMBRYO-MAKER (EMK [9]).
In concert with TFs, microRNAs, which seem to preferentially target TF genes to control plant development [10,11], have been suggested as regulating SE induction. In line with this assumption, the differential expression of numerous miRNAs has been indicated in an embryogenic culture of Arabidopsis, and the function of the candidate microRNAs was attributed mainly to hormone and stress responses [12]. In addition to Arabidopsis, the differential expression of numerous miRNAs has been reported in the SE of other plants, including Oryza sativa [13], Liriodendron tulipifera L. chinense [14], Larix laptolerix [15], Gossypium hirsutum [16], Zea mays [17], and Manihot esculenta [18]. Several miRNAs have been functionally analyzed during SE, including miR160 and miR166/165 [19], miR167 [20], miR393 [21], and miR156 [22]. In addition, miR396 was postulated to regulate SE induction given the distinctly increased expression of primary (pri-miRNA) and mature-miR396 in an embryogenic culture of Arabidopsis [12]. In vivo, miR396 molecules have been postulated to control leaf development in Arabidopsis by regulating various cellular processes including the proliferation, expansion, and differentiation of cells [23,24]. In root development, the function of miR396 has been associated with regulating the switch between stem cells and transit-amplifying cells [25].
In Arabidopsis, miR396, which is produced from two MIR396 (MIR396a and MIR396b) genes, has been indicated to repress the TF genes that encode the members of the GROWTH-REGULATING FACTOR (GRF) gene family [26]. In Arabidopsis, the GRF gene family comprises nine members, and miR396 recognizes the target site in seven of these genes, including GRF1–4 and GRF7–9 [26]. The miR396-targeted GRFs have been indicated to regulate plant growth and development in vivo, which involves leaf and cotyledon growth [27] and stem, flower, and root development (reviewed in Reference [28]). In the development of the Arabidopsis root, the GRFs act as repressors of PLETHORA1 (PLT1) and PLT2. In turn, the PLTs activate miR396, and as a result, miR396 represses the GRFs [25]. Given that the increased expression of PLT2 induces SE from Arabidopsis seedlings, the involvement of the miR396–GRF–PLT regulatory network in SE induction might be assumed [29].
Importantly for the possible role of miR396 in the stress-related mechanism of SE induction [29], various stress treatments (e.g., drought, salinity, low temperature, and Ultraviolet B (UVB) radiation) have been found to modulate the level of miR396 [30,31].
In order to gain insight into the function of miR396 in SE induction, we studied the embryogenic response that is induced in in vitro cultured Arabidopsis explants in relation to the miR396 expression level and auxin treatment. Some of the evidence implied that miR396 controls efficient SE induction through an auxin biosynthesis-related pathway, and among the targets of miR396, GRFs were suggested, including GRF1/4/7/8/9. In addition, the regulatory relationship between the miR396–GRF module and the PLETHORA genes, including PLT2, was investigated.
2. Results
2.1. miR396 Was Expressed Specifically in the SE-Induced Explant Parts
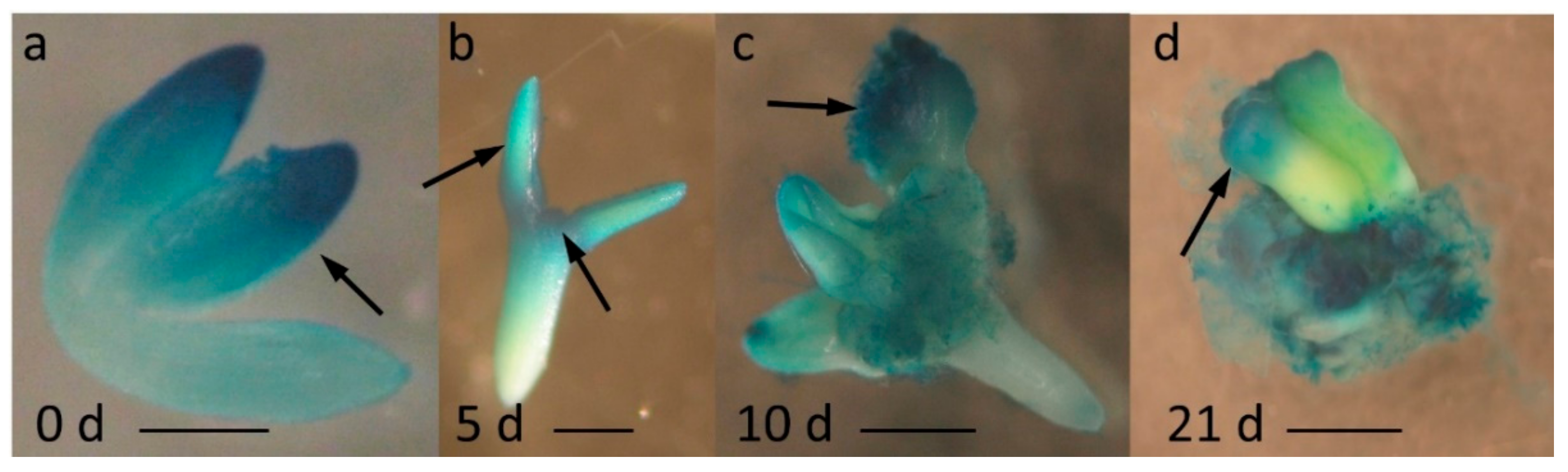
Our previous analysis of the SE transcriptome indicated a significant upregulation of MIR396 genes and their mature product in an embryogenic culture of Arabidopsis, and thus, a role for miR396 in regulating SE was suggested [12]. Consistent with this assumption, the present analysis of the MIR396b:GUS reporter line with promoter:GUS fusions revealed that the explants that were induced toward SE preferentially accumulated the MIR396b transcript in the explant parts that were involved in embryogenic induction, including the cotyledons and the proximity of the shoot apical meristem (SAM) (Figure 1a–c). Moreover, we observed that, similarly to the immature zygotic embryos ((IZEs), 0 d) that were used as the explants for SE induction, somatic embryos that were developed on the SE-induced explants also showed GUS expression in the cotyledons (Figure 1a vs. Figure 1d). This result implied a similarity between the MIR396b expression pattern in the somatic embryos and their zygotic counterparts.
2.2. miR396 Contributed to SE Induction via an Auxin Biosynthesis-Related Pathway
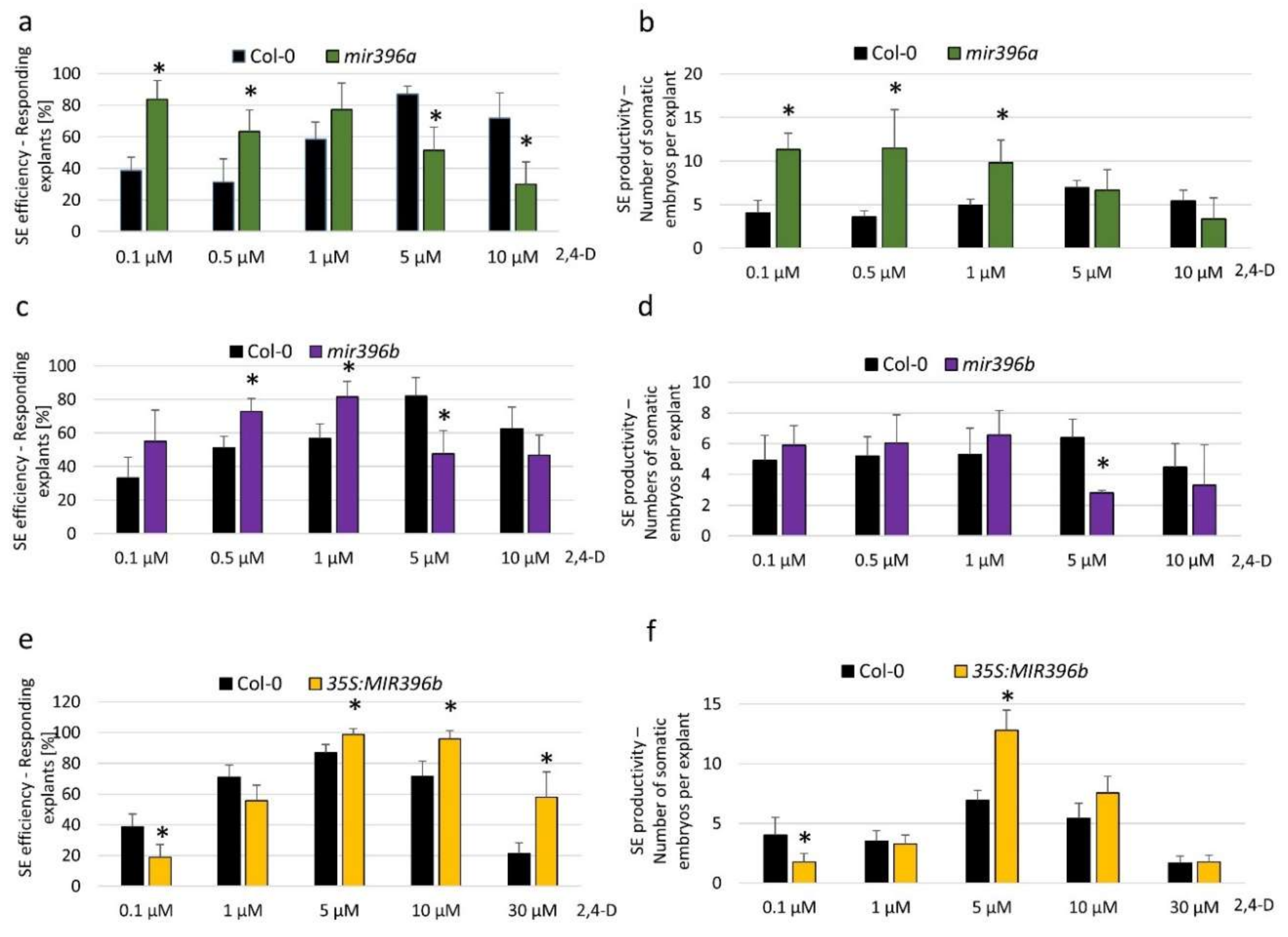
To explore the mechanism that is involved in miR396-mediated SE induction, we analyzed the relationship between the expression level of miR396 and the embryogenic potential of the explants. To this end, explants of miR396 insertional mutants (mir396a and mir396b) and the 35S:MIR396b line with down- and upregulated miR396 expressions, respectively, were analyzed on induction media that had been supplemented with different concentrations (0.1, 0.5, 1.0, 10.0, and 30.0 µM) of 2,4-Dichlorophenoxyacetic acid (2,4-D), which is a synthetic auxin that is commonly used to induce SE. The results showed that on a standard medium with 5 µM 2,4-D, which is highly efficient for SE induction in a wild type (WT; Col-0)) culture, the mutants and the overexpression of the 35S:MIR396b line displayed a decreased and increased SE response, respectively, compared to the WT explants (Figure 2). However, on media that had been supplemented with 2,4-D at concentrations that were suboptimal for SE induction in a WT culture (0.1, 0.5, and 1.0 µM), the embryogenic response of the mir396 mutants was distinctly higher than in the WT explants (Figure 2a–d). In contrast, the explants of the 35S:MIR396b overexpressor line were opposite to the mutants’ phenotypes. Accordingly, the explants overexpressing miR396 showed a distinctly higher embryogenic response after treatment with a 2,4-D concentration that was overoptimal for SE induction in WT culture (Figure 2e,f). Altogether, the results showed a negative relationship between the miR396 expression level and the sensitivity of the explants to exogenous auxin treatment, and thus, we hypothesize that miR396 may contribute to SE induction via an auxin-related pathway.
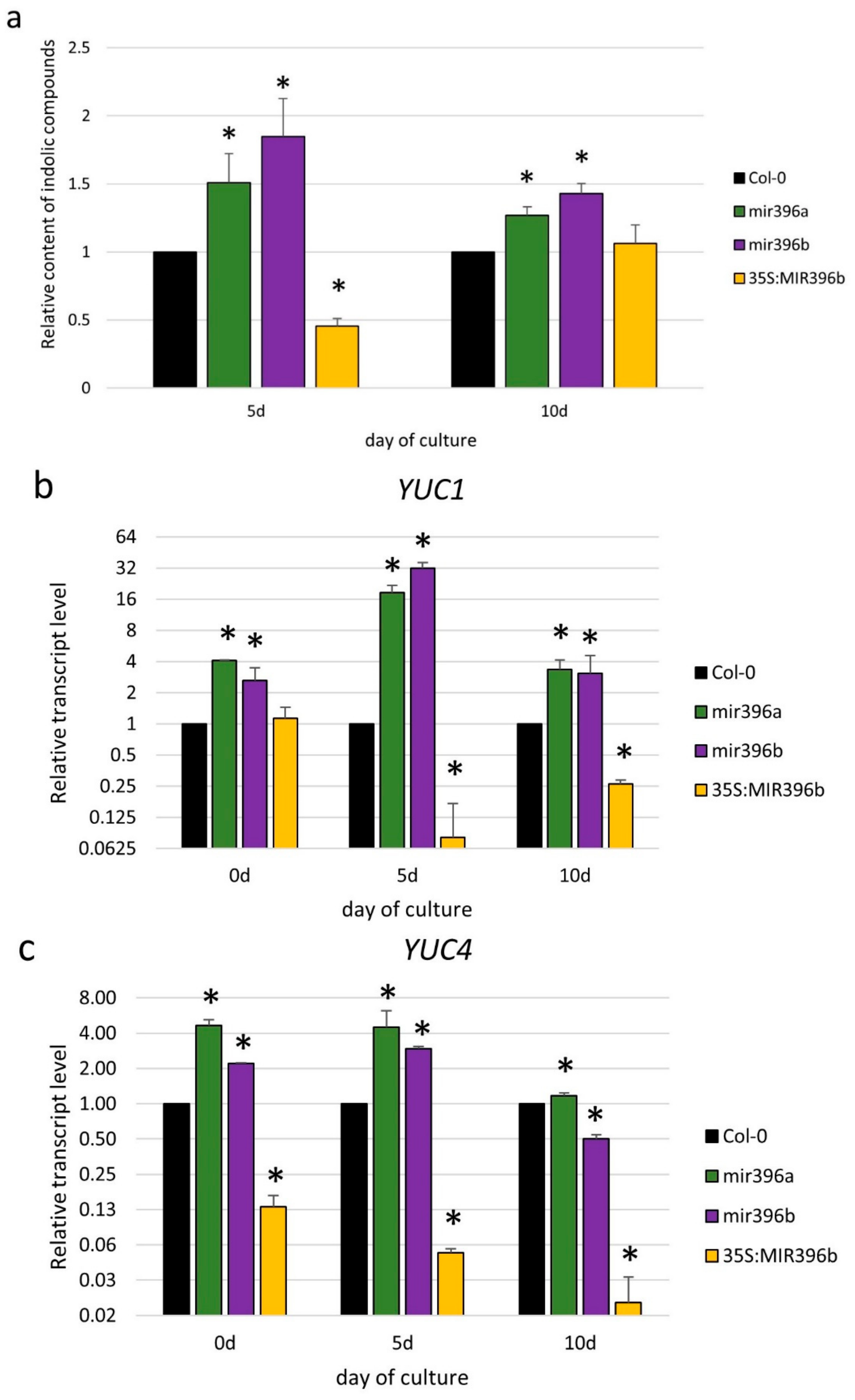
In order to verify this assumption, we evaluated the content of the indolic compounds in relation to the miR396 expression level. An analysis of the mir396a and mir396b mutants showed as much as a 1.85- and 1.50-fold increased level of indolic compounds, respectively (Figure 3a; Table S1). In contrast to the mutant cultures, the level of indolic compounds was decreased by as much as two-fold in the 35S:MIR396b culture. Given that Indolilic-3-acetic acid (IAA) contributed to the pool of indolic compounds, these results suggest a negative impact of the miR396 expression on the IAA accumulation in the SE-induced explants, and thus we assume that miR396 might control the auxin biosynthesis pathway during SE induction.
Thus, we evaluated the expression level of the YUC1, YUC4, and YUC10 genes encoding the key enzymes of the auxin biosynthesis pathway during SE in Arabidopsis [32]. Analyses of the mir396 mutants and the 35S:MIR396b line suggested a relationship between YUC1 and YUC4 and miR396 during SE induction, and accordingly, the gene transcription was substantially up- and downregulated in the cultures of the mir396 mutants and the 35S:MIR396b line, respectively (Figure 3b,c). In contrast to YUC1 and YUC4, the expression of YUC10 was not detected in the cultures with a deregulated miR396 expression.
In conclusion, the results imply that miR396 might contribute to the SE response via negative control of the YUC1 and YUC4 genes of the auxin biosynthesis pathway and to a decrease in the endogenous auxin accumulation.
2.3. miR396 Controlled the GRF Genes during SE Induction
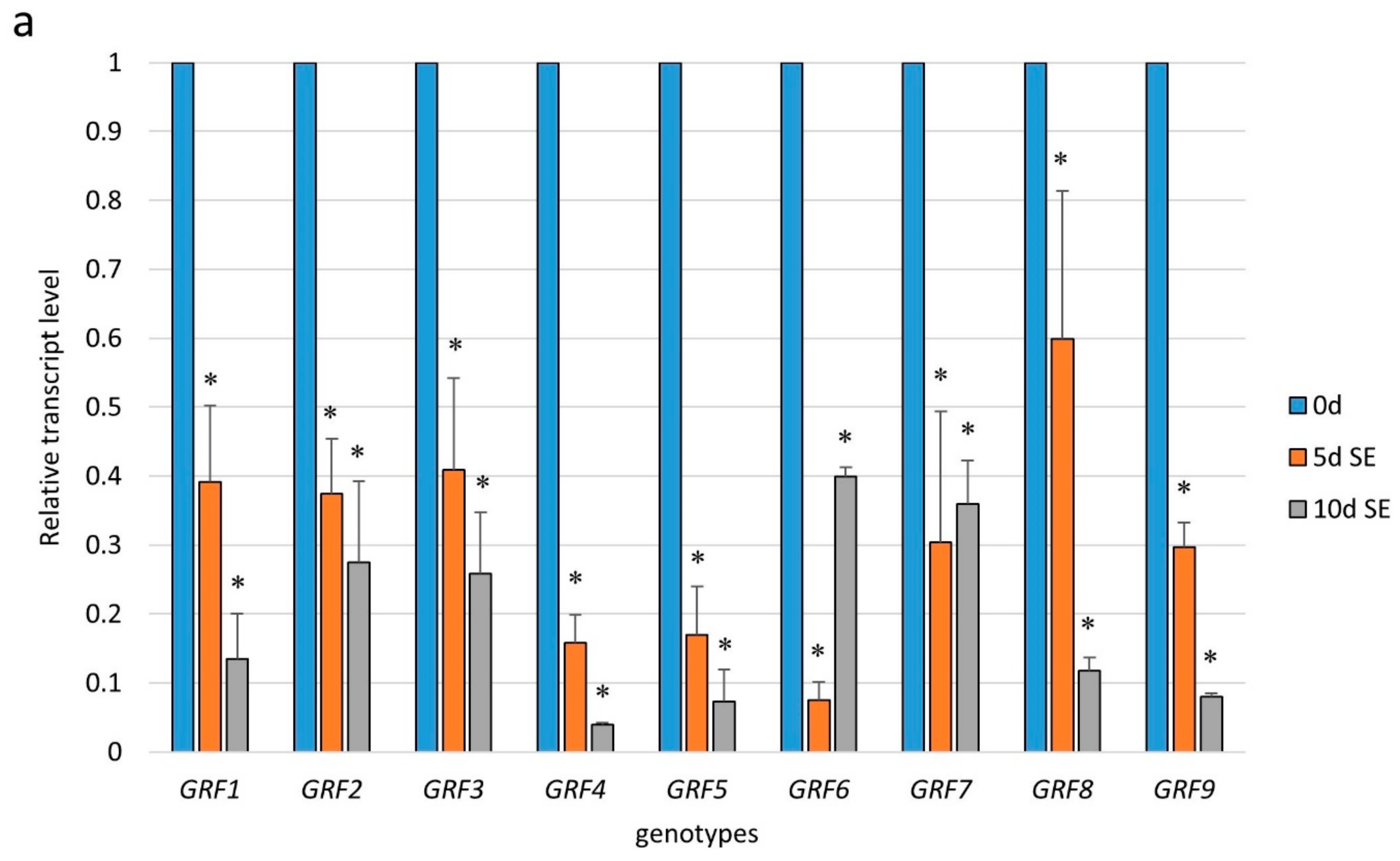
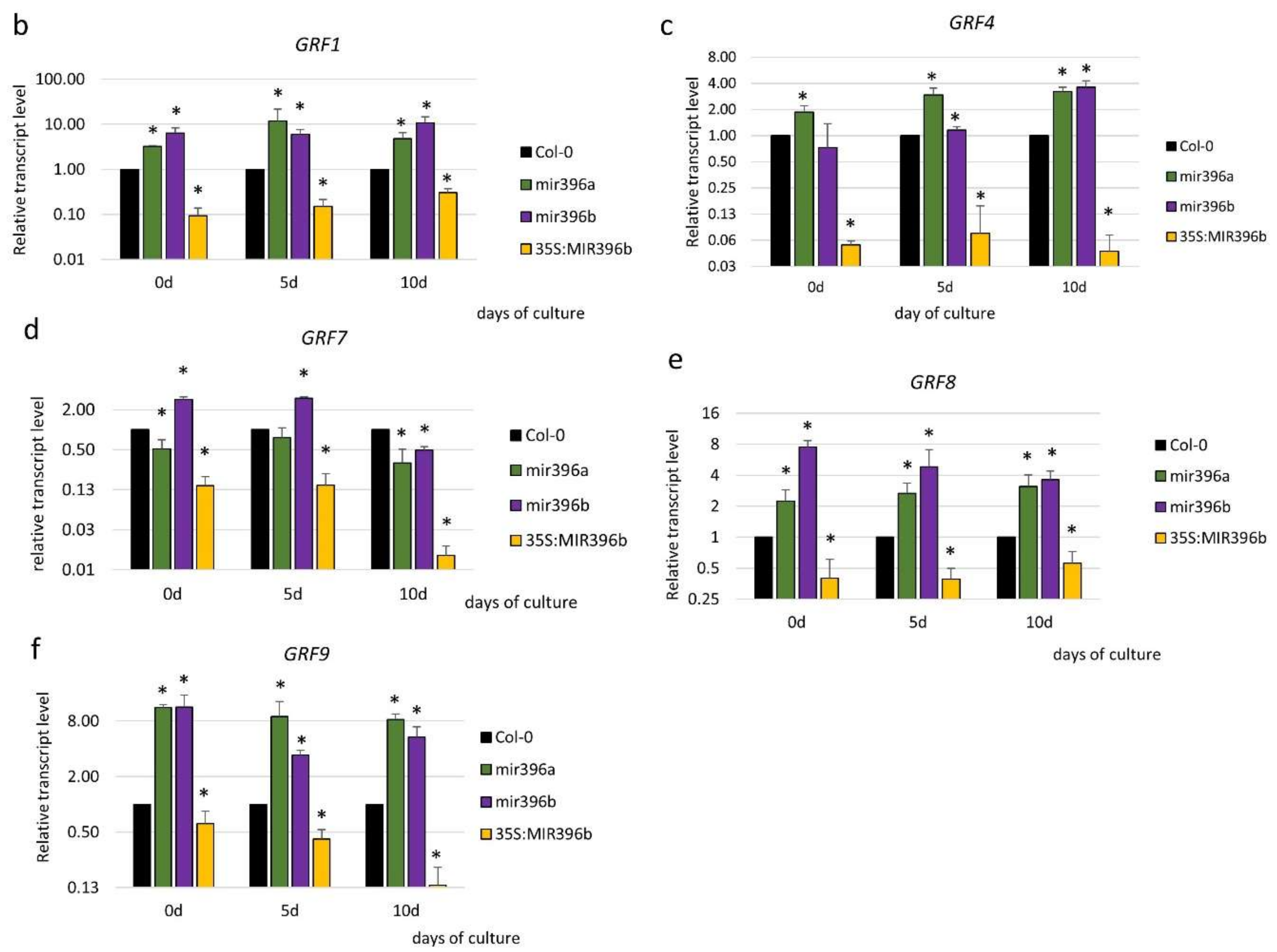
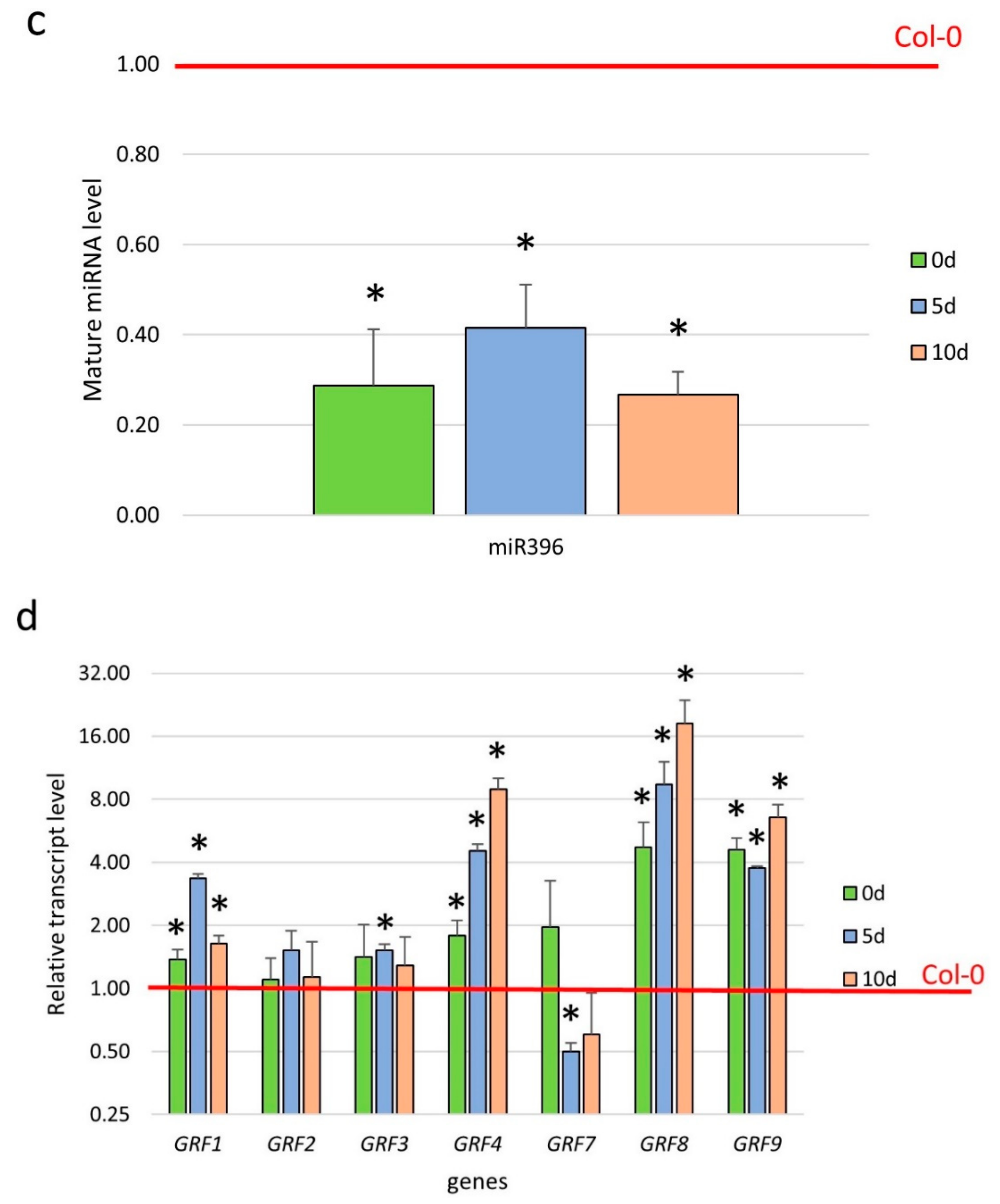
To gain more insight into the function of miR396 in SE induction, we profiled the expression of the GRF genes, which are potential targets of miR396 in plants [29]. The results of an RT-qPCR analysis showed a significant downregulation of all of the analyzed GRFs (GRF1–9) during the early (5 d) and advanced (10 d) stages of the SE culture that had been induced in the WT (Col-0) explants (Figure 4a). A significantly decreased level of the GRF1–9 transcripts in SE was found to be opposite to the distinct accumulation of miR396 during SE (Figure S1), and this observation suggests a regulatory relationship between miR396 and the GRF genes.
In order to verify this assumption, we analyzed the expression of the GRF genes in the SE-induced explants of the mir396 (mir396a and mir396b) mutants and the 35S:MIR396b overexpressor line. The analysis showed that the transcripts of five of the analyzed genes (GRF1, GRF4, GRF7, GRF8 and GRF9) were significantly up- and down-regulated in the mir396 mutants and MIR396b overexpressor line culture, respectively (Figure 4b–f). In contrast, there was no impact of the miR396 expression on the transcript level of GRF2, GRF3, GRF5, and GRF6 (Figure S2). In conclusion, the reverse expression level of GRF1, GRF4, GRF7, GRF8, and GRF9 in the mir396 mutants versus the 35S:MIR396b overexpressor line cultures implies that miR396 might control these GRFs during SE induction.
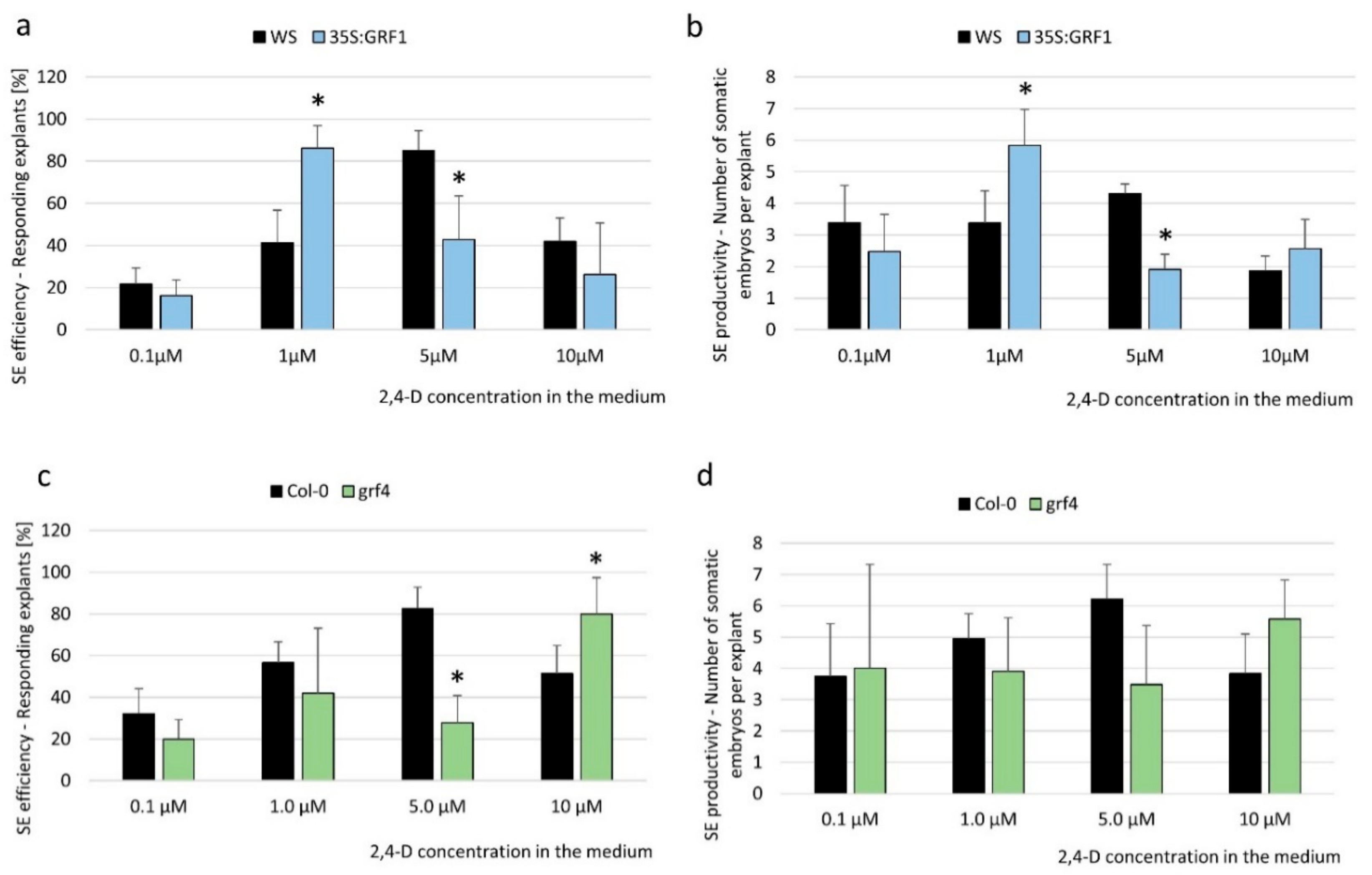
To confirm that the miR396-regulated GRFs contributed to SE induction via an auxin-related mechanism, we evaluated the embryogenic response of the 35S:GRF1 transgenic line and the grf4 mutant in media with different concentrations of 2,4-D. The analyses indicated that the 35S:GRF1 explants that had been treated with an auxin concentration of 1.0 µM, which is suboptimal for SE induction in a WT culture, displayed the highest level of SE efficiency and productivity (Figure 5a,b). This finding implies that similarly to the mir396 mutants with a significantly impaired miR396 accumulation (Figure 2a–d), the overexpression of GRF1 resulted in an increased sensitivity of tissues to the auxin treatment that was effective for SE induction.
Consistent with this result, the grf4 mutant phenotype was found to be opposite to the 35S:GRF1, and the highest embryogenic response of the mutant explants was observed on the medium with 10.0 µM of 2,4-D, i.e., a concentration that was overoptimal for SE induction in a WT culture (Figure 5c,d). Moreover, we observed that the response of the grf4 mutant to auxin treatment was similar to the one that was observed in the culture overexpressing MIR396b, in which the overoptimal 2,4-D concentration (10.0 µM) increased the SE response (Figure 2e,f). In contrast to the SE-affected phenotype of grf4, the embryogenic potential of the grf1, grf8, and grf9 knockdown mutants was not significantly impaired (Figure S3), and that could suggest a functional redundancy between these GRFs in SE.
Taken together, the results suggest that through the repression of GRF1 and GRF4, miR396 might control the embryogenic transition by modulating the sensitivity of tissues to auxin treatment. We also assumed that GRF7, GRF8, and GRF9 might be the targets of miR396 in SE, and further analyses using the relevant mutant lines are required to verify this regulatory relationship.
2.4. The Regulatory Relationship between miR396 and PLT1/PLT2 in SE Induction
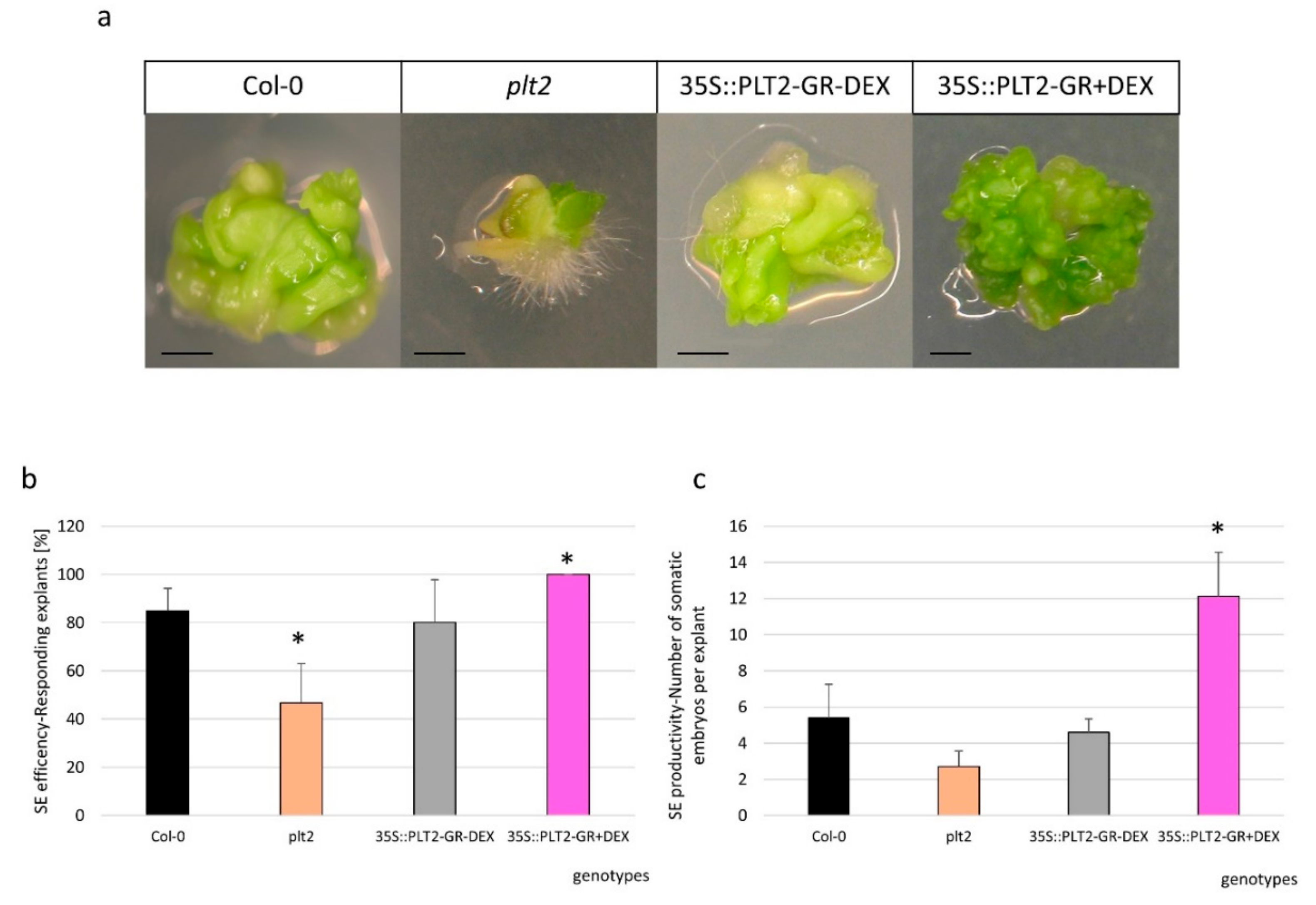
A regulatory interaction between miR396, GRF, and PLTETHORA (PLT) transcription factors in root development [25] motivated us to investigate the relationship between PLT genes and the miR396/GRF module during SE. We found that PLT2 positively regulated SE induction because the cultures of the plt2 mutant and the 35S:PLT2-GR overexpressor line displayed a significantly reduced and enhanced, respectively, SE response (Figure 6a–c).
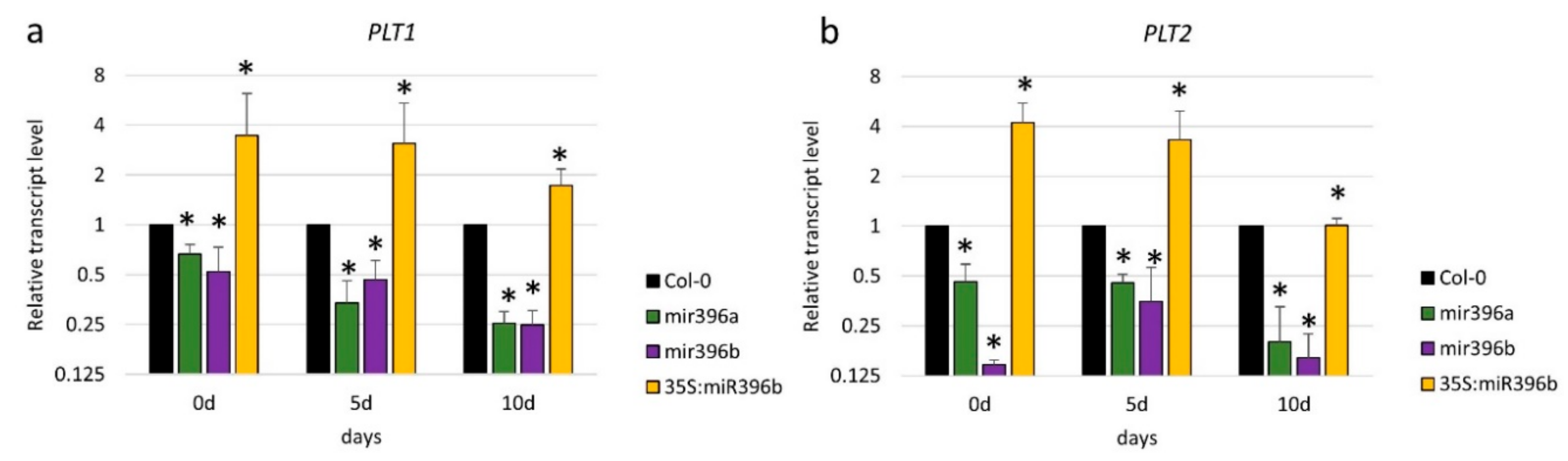
To further explore the relationship between PLT and miR396 in SE induction, we profiled the expression of PLT1 and PLT2 in the Col-0 (WT) explants that had been cultured on a standard auxin (E5) medium. We observed that the PLT1 and PLT2 expression patterns reflected those of miR396 (Figure S4). In addition, we found the PLT1 and PLT2 transcript levels to be down- and upregulated in the cultures of the mir396 mutants and the 35S:MIR396b overexpressing line, respectively (Figure 7a,b). These results suggested a positive relationship between the level of miR396 and the PLT1/PLT2 expression during SE induction. In addition, the plt2 mutant culture displayed a significantly decreased miR396 level (Figure 7c) and the upregulation of GRF1, GRF4, GRF8, and GRF9 transcripts (Figure 7d).
Altogether, the results suggest a role for the miR396–GRF1/4/8/9–PLT1/2 module in SE induction in Arabidopsis. Complex regulatory relationships within this module that involve negative and positive control over GRFs and PLT (respectively) by miR396 might be assumed.
3. Discussion
3.1. An Accumulation of miR396 Was Associated with SE Induction
Stress responses play a central role in the molecular mechanism of SE induction [29,33]. Given that stresses have been reported to distinctly upregulate miR396 [30,31], the increased accumulation of miR396 in the embryogenic culture of Arabidopsis [12] might reflect the explant’s response to stress that is imposed in vitro by different culture conditions, including treatment with 2,4-D [29]. Accordingly, we found that in response to 2,4-D treatment, the miR396 transcripts were specifically expressed in the SE-involved areas of the explants, i.e., in proximity to cotyledons and the SAM [34]. Thus, to gain insight into the biological function of miR396 in SE induction, we explored the relationship between miR396 and 2,4-D, which is a synthetic auxin that has an SE-promoting activity in Arabidopsis and other plants [35].
3.2. miR396 Controlled the Embryogenic Response by Modulating the Sensitivity of Tissues to Auxin
Numerous hormone-related miRNAs have been suggested to control SE induction in Arabidopsis [12], and among them, miR393, miR160, and miR165/166 have been indicated to impact auxin metabolism and signaling in embryogenic explants [19,21]. A relationship between miR396 and hormones during SE might also be expected given that ethylene has been found to modulate the expression of miR396 in Medicago truncatula roots [36] and that the involvement of miR396 in brassinosteroid (BR) and gibberellin (GA) signaling in rice plants [37] has also been reported.
Here, we provide evidence of the relationship between miR396 and the auxin that accumulated in the SE-induced explants of Arabidopsis [32]. Accordingly, an inverse relationship between the miR396 expression level and the sensitivity of the explants to auxin treatment was demonstrated (Figure 2), and a negative impact of the miR396 expression on the accumulation of the indolic compounds (including IAA in the SE-induced explants) was indicated (Figure 3a). Consistently, two YUC genes (YUC1 and YUC4) of the auxin biosynthesis pathway that contribute to the embryogenic response in Arabidopsis [32] were found to be miR396-repressed during SE induction (Figure 3b,c). In agreement with the assumption that miR396 negatively controls auxin biosynthesis during SE induction in Arabidopsis explants, miR396 was found to repress auxin biosynthesis during the development of Oryza sativa plants [38]. We observed that the level of indolic compounds in the 35S:MIR396b culture was not reduced on day 10 despite a decreased expression level of YUC genes. This may suggest that besides a tryptophan-dependent and YUC-involved auxin biosynthesis pathway, a tryptophan-independent pathway might also contribute to IAA production during an advanced SE stage (10 d), which is related to somatic embryo development. In support of this assumption, an important role of the tryptophan-independent auxin biosynthesis pathway in the establishment of the apical–basal pattern during early zygotic embryogenesis has been indicated in Arabidopsis [39].
3.3. miR396 Regulated SE Induction by Repressing GRFs (GRF1, 4, 7, 8, and 9)
Members of the GRF gene family have been postulated as candidate targets of miR396, and a role for the miR396–GRF module in Arabidopsis development has been reported, including the regulation of cell proliferation in leaves and the transition of stem cells in roots [23,25]. In addition, miR396 has been indicated to control GRFs in other plants, including seed development in rice and barley [39] and the formation of adventitious roots in apple rootstock [40].
Consistent with these findings, our results imply that miR396 targeted the GRFs in order to regulate the embryogenic transition that was induced in in vitro cultured explants of Arabidopsis. Conclusive for this postulate was the negative relationship between the accumulation of miR396 and the expression level of five of the GRF genes (GRF1, 4, 7, 8, and 9) in the embryogenic culture of the mir396 mutants and the 35S:MIR396b line (Figure 4b–f). In support of the postulated engagement of the miR396–GRF module in the embryogenic transition in Arabidopsis, the miR396-controlled repression of GRF1, GRF4, and GRF8 has been reported during SE of Lilium pumilum [41], hybrid yellow poplar [14], and cotton [16]. A report on the GRF4-regulated development of cotyledons and SAM in zygotic embryos that provide SE-responsive tissue in Arabidopsis [34,42] also supported a role for GRFs in embryogenic development. Relevant to the stress and cell reprogramming-associated mechanism of SE induction [33], the miR396–GRF1 module was found to control the specification and differentiation of cells during the stress response in Arabidopsis plants [43].
This study provides evidence that the miR396–GRF module contributes to SE induction via an auxin-related mechanism. In line with this assumption, an analysis of the 35S:GRF1 explants in media with different concentrations of 2,4-D showed that the embryogenic response of these explants phenocopied that of the mir396 mutants but contrasted with the response of the MIR396b overexpression line (Figure 5a,b). Moreover, the explants of the grf4 mutant had a decreased sensitivity to auxin treatment that was also characteristic of the 35S:MIR396b overexpression culture (Figure 5c,d). A lack of the SE-affected phenotype in the other analyzed grf mutants (Figure S3) might suggest a functional redundancy of some GRFs in the control of SE, as was indicated for GRF1–3 in the development of leaves and cotyledons in vivo [27]. In contrast, the distinct SE phenotype of the grf4 mutant (Figure 5b) seemed to confirm an important function of GRF4 in plant embryogenesis given its reported role in the development of cotyledons and the SAM in the zygotic embryogenesis of Arabidopsis [42]. In support of the contribution of GRF4 to SE, the cotyledons and SAM of a zygotic embryo provide highly responsive tissue in SE induction in Arabidopsis [34].
Collectively, these results imply that through the repression of GRFs, including GRF1, 4, 7, 8, and 9, miR396 appeared to regulate the auxin-related mechanism of the embryogenic transition that was induced in Arabidopsis explants.
3.4. miR396 Positively Controlled PLT Genes in SE Induction via GRFs
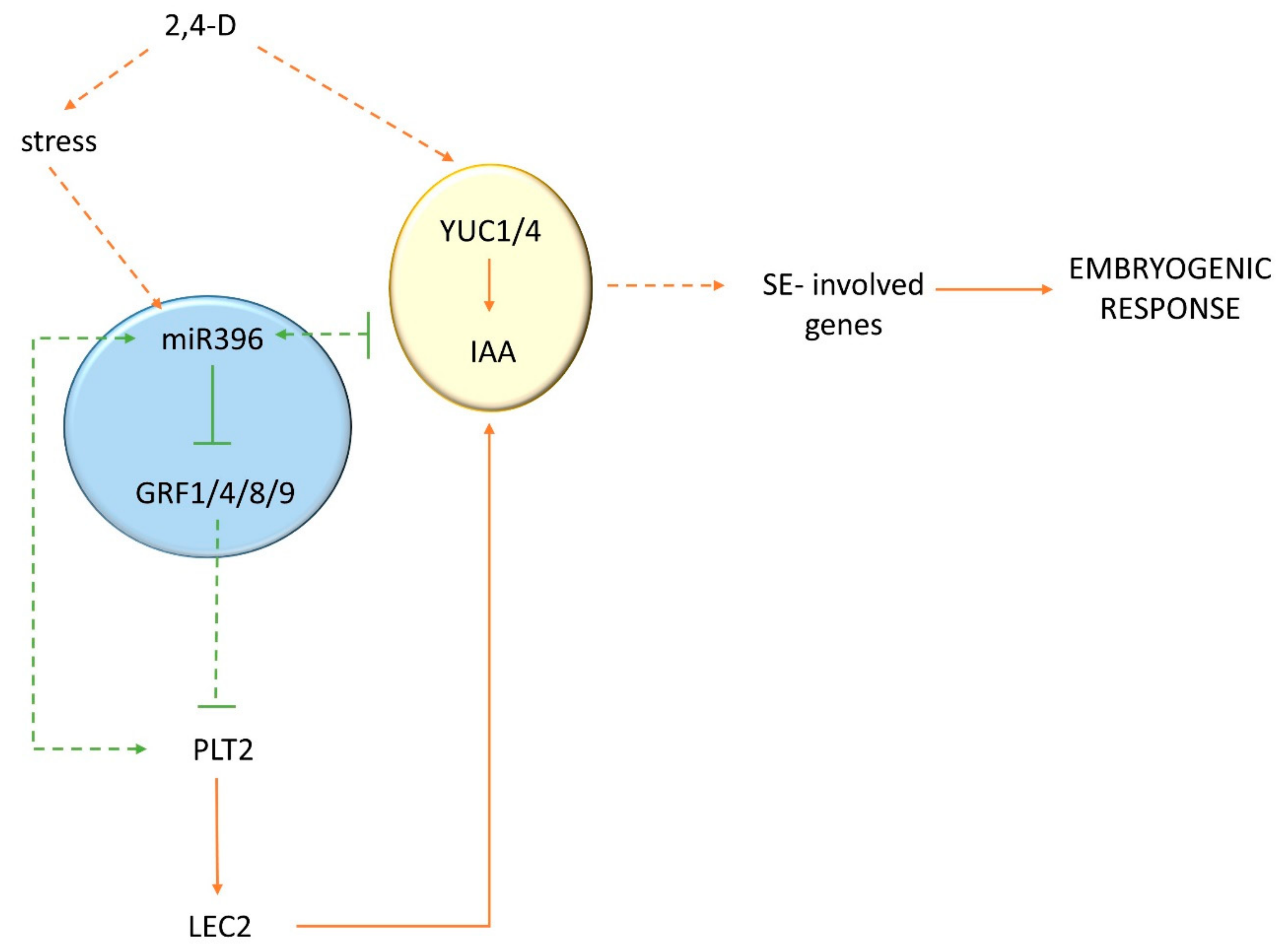
In order to dissect the biological function of the miR396–GRF module in the SE-associated regulatory network of genes, we found it reasonable to gain insight into the relationship of this module with the TFs that have a documented regulatory role in the embryogenic transition. Among the TFs that control embryogenic induction in somatic plant cells, the BABY BOOM (BBM) gene, which is a member of the PLETHORA (PLT) clade of genes that encode AP2/ERF TFs, has been identified [4]. We postulated that miR396 might indirectly control BBM/PLT genes through GRFs during SE induction. The lack of miR396-targeted sequences in the PLT genes [26] rules out a direct miR396–PLT regulatory relationship. A role for the miR396–PLT–GRF node in SE induction was postulated given that the expression pattern of PLT1/PLT2 in the embryogenic culture was found to be similar to miR396 (Figure S4) and that the decreased expression of PLT1/PLT2 in the mir396 mutants contrasted with the accumulation of the PLT1/PLT2 transcripts in the 35S:MIR396b culture (Figure 7a,b). In addition, a decreased expression level of the PLT genes in the GRF2 and GRF3 overexpression lines was reported during root development [25]. GRF1 was reported to have a redundant function to GRF2 and GRF3 [27], and thus a similar regulatory relationship between GRF1 and PLT might be assumed. Further evidence for a positive regulatory impact of miR396 on PLTs included the similar sensitivity to auxin treatment of the miR396 and plt2 mutant explants, which showed enhanced SE induction after treatment with a 2,4-D concentration that was suboptimal for the WT culture (Figure S5).
In order to control the transition of root stem cells into amplifying cells in the root meristem of Arabidopsis, miR396 negatively regulates GRFs, which are repressors of PLTs (PLT1/PLT2/BBM), which in turn are necessary to activate the MIR396 genes [25]. We postulate that a congruous regulatory feedback loop between miR396, GRFs, and PLTs might operate during SE because the embryogenic culture of the plt2 mutant showed a decreased level of miR396 and an increased expression of GRFs (Figure 7c,d).
Taken together, we postulate that through the repression of GRFs (GRF1, GRF4, GRF8, and GRF9), miR396 activates PLTs (PLT1/PLT2/BBM). Given that the PLT genes (BBM and PLT2) have been demonstrated to trigger SE through the direct transcriptional regulation of the LEC1–ABI3–FUS3–LEC2 network that controls embryo identity [44], we also assumed that PLTs would impact auxin accumulation in the SE-induced explants by controlling the LEC2 gene. In support of the positive control of LEC2 by PLT, the plt2 mutant showed a significantly decreased level of LEC2 transcripts (Figure S6).
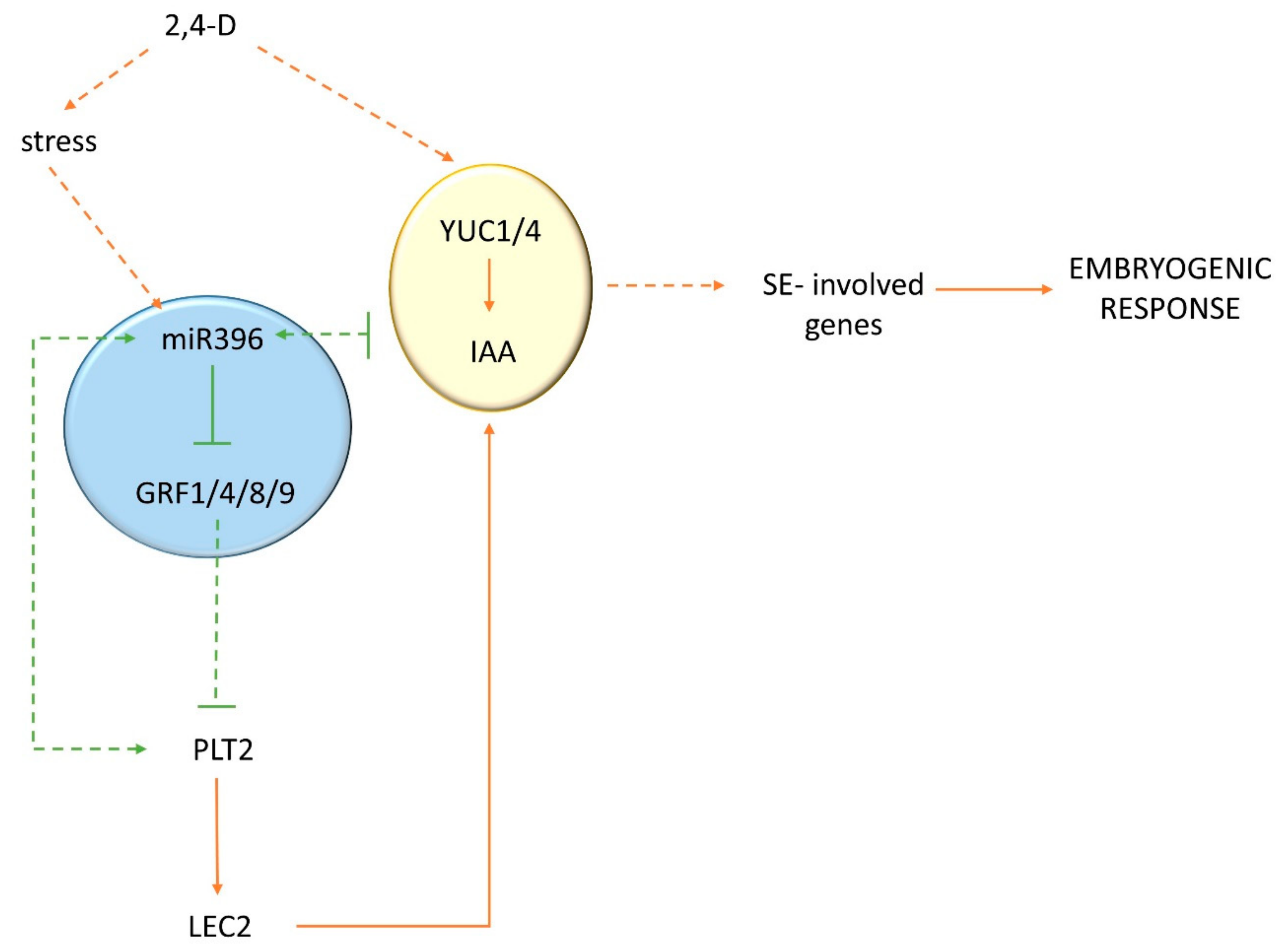
In addition to PLTs, our analysis of the MIR396 promoter sequence [45] revealed that other key regulators of SE induction, including AGL15 [7], might directly regulate miR396 embryogenic induction. AGL15 controls multiple hormone interactions during SE induction, including the direct targeting of LEC2, which is an activator of auxin biosynthesis in SE [32,46]. In support of a relationship between AGL15 and the miR396–GRF module, GRF9 was identified as being a possible direct target of AGL15, and GRF1 was postulated to control PHAVOLUTA (PHV) [47], which positively controls the LEC2-mediated pathway of SE induction in Arabidopsis [19,32]. In summary, complex and versatile regulatory interactions seem to operate between miR396 and the TFs that play a central role in SE induction (BBM, AGL15, and LEC2). In the postulated model, LEC2 seemed to be a key link between the GRF/PLT and auxin-regulated SE induction, and further experiments are required to verify an miR396-controlled network that is involved in auxin-regulated embryogenic transition in Arabidopsis (Figure 8).
4. Materials and Methods
4.1. Plant Material and Growth Conditions
The seeds of different genotypes of Arabidopsis thaliana (L.) Heynh., including the Columbia (Col-0) WT genotype and the insertional mutants mir396a (N447416), mir396b (N412157), grf4 (N657589), and plt2 (N676869), were supplied by NASC (The Nottingham Arabidopsis Stock Centre, Nottingham, UK). The seeds for the 35S:MIR396b and MIR396b/GUS reporter lines were kindly provided by Javier F. Palatnik (Research Council, Institute of Molecular and Cell Biology in Rosario, Argentina). The seeds for the 35S:GRF1 transgenic line, which was derived from the Wassilewskija (WS) genotype, were kindly provided by Jeong Hoe Kim (Department of Biology, Kyungpook National University, Daegu, South Korea). The transgenic line (overexpressing) PLT2 gene (35S:PLT2-GR), was kindly provided by Kim Boutilier (Wageningen Plant Research, Wageningen, Netherlands). Descriptions of the transgenic genotypes that were used in the study are presented in Table S2. The level of mature miR396 was significantly decreased and increased in the analyzed miR396 mutants and 35S:MIR396b line, respectively (Figure S7).
The plants were kept in a growth chamber at 21 ± 1 °C under a 16/8-h photoperiod of 40 μM m−2s−1 white fluorescent light.
4.2. Somatic Embryogenesis Induction
Immature zygotic embryos (IZEs) from different genotypes at the green cotyledonary stage were used as the explants for the in vitro cultures. To induce SE, the standard protocol was used [48]. The IZEs were cultured on an E5 solid medium containing B salts and vitamins [49] that was supplemented with 5 µM 2,4-dichlorophenoxyacetic acid (2,4-D, Sigma-Aldrich, St. Louis, MO, USA), 20 g L−1 sucrose, and 8 g L−1 agar. In some of the experiments, different concentrations of 2,4-D (0.1, 0.5, 1.0, 10, and 30 µM) were used in the induction medium.
At selected time points of the culture (0, 5, and 10 days), the explants of Col-0, mir396a, mir396b, 35S:MIR396b, and plt2 were sampled for transcriptome analysis.
The explant capacity for SE was evaluated on the 21st day of the in vitro culture. Two parameters were calculated: SE efficiency, i.e., the percentage of the explants producing somatic embryos, and SE productivity, i.e., the average number of somatic embryos produced per explant. All of the culture combinations were estimated in three replicates: at least 30 explants (10 explants per Petri dish) were analyzed per 1 replicate.
4.3. Content of Indolic Compounds
To evaluate the content of the indolic compounds, including IAA, the colorimetric technique was used [50]. Explants of the Col-0, mir396a, and mir396b mutants and 35S:MIR396b were induced for 5 and 10 days on an E5 medium. The concentration of IAA was established using a calibration curve of pure IAA as the standard following a linear regression analysis. Each measurement was performed in three replicates.
4.4. RNA Isolation and RT-qPCR Analysis
An Ambion RNAqueous Kit (Thermo Fisher Scientific, Waltham, MA, USA) was used to isolate the total RNA from the IZE explants on days 0, 5, and 10 of the culture on an auxin (E5) medium in three biological repeats. The concentration and purity of the RNA were evaluated using an ND-1000 spectrophotometer (Nano Drop Technologies, LLC, Wilmington, DE, USA). Total RNA that had been treated with RQ1 RNase-free DNase I (Promega Corporation, Madison, WI, USA) was reverse-transcribed using a RevertAid First-Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA, USA) following the manufacturer’s instructions. A LightCycler® 480 SYBR Green I Master (Roche, Basel, Switzerland) was applied for the RT-qPCR reactions. The primers that were used for the expression profiling of the studied genes are listed in Table S3.
4.5. Stem-Loop RT-PCR for Mature miRNA Detection
A mirVana™ miRNA Isolation Kit was used to isolate the small RNAs from the IZE explants that were induced on an auxin (E5) medium on days 0, 5, and 10 of the culture. The concentration and purity of the small RNAs were evaluated using an ND-1000 spectrophotometer (Nano Drop Technologies, LLC, Wilmington, DE, USA). The design of the oligonucleotides, stem-loop reverse transcription, and real-time qPCR were performed according to Speth and Laubinger [51]. The real-time qPCR analysis for the mature miR396 accumulation was performed using a LightCycler 480 (Roche, Basel, Switzerland). The primers that were used to detect miR396 are listed in Table S3.
4.6. Transcript Level Calculation
The relative transcript levels were calculated and normalized to an internal control, the At4g27090 gene-encoded 60S ribosomal protein. The plant tissues for real-time qPCR analysis were produced in three biological repetitions, and two technical replicates of each repetition were performed. The relative expression level was calculated using the 2−∆∆Ct method [52].
4.7. Histochemical Staining of GUS
The histochemical analysis of GUS activity was performed as described by Jefferson et al. (1987). The explants were stained in X-Gluc (5-bromo-4-chloro-3-indolyl β-D-glucuronide-cyclohexylammonium salt) (Sigma Aldrich, St. Louis, MO, USA). The reaction was carried out in the dark at 37 °C for 12 h. The GUS signal was visualized using a Zeiss Stemi 2000-C microscope (Zeiss, Oberkochen, Germany).
4.8. Statistical Analysis
Student’s t-test was used to calculate any significant differences (at p = 0.05) between the compared samples. The figures present the averages with standard deviations.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/1422-0067/20/20/5221/s1.
Author Contributions
M.D.G. conceived of and supervised the study; A.S.-S. conducted the experiments; M.D.G. and A.S.-S. designed the study and prepared the manuscript.
Funding
This work was supported by a research grant from the National Science Center in Poland (OPUS5 2013/09/B/NZ2/ 03233).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| 2,4-D | 2,4-Dichlorophenoxyacetic acid |
| AGL15 | AGAMOUS-LIKE15 |
| BBM | BABY BOOM |
| DEX | Dexamethasone |
| E5 | Embryogenesis induction medium with auxin |
| GRF | GROWTH-REGULATING FACTOR |
| LEC2 | LEAFY COTYLEDON2 |
| MIR | Micro-RNA gene |
| IAA | Indolilic-3-acetic acid |
| IZE | Immature zygotic embryo |
| PLT | PLETHORA |
| SAM | Shoot apical meristem |
| SE | Somatic embryogenesis |
| TF | Transcription factor |
References
- Gordon-Kamm, B.; Sardesai, N.; Arling, M.; Lowe, K.; Hoerster, G.; Betts, S.; Jones, T. Using Morphogenic Genes to Improve Recovery and Regeneration of Transgenic Plants. Plants 2019, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Gliwicka, M.; Nowak, K.; Balazadeh, S.; Mueller-Roeber, B.; Gaj, M.D. Extensive Modulation of the Transcription Factor Transcriptome during Somatic Embryogenesis in Arabidopsis thaliana. Plos One 2013, 8, e69261. [Google Scholar] [CrossRef] [PubMed]
- Wickramasuriya, A.M.; Dunwell, J.M. Global Scale Transcriptome Analysis of Arabidopsis Embryogenesis In Vitro. Bmc Genom. 2015, 16, 301. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, K. Ectopic Expression of BABY BOOM Triggers a Conversion from Vegetative to Embryonic Growth. Plant Cell Online 2002, 14, 1737–1749. [Google Scholar] [CrossRef]
- Gaj, M.D.; Zhang, S.; Harada, J.J.; Lemaux, P.G. Leafy Cotyledon Genes Are Essential for Induction of Somatic Embryogenesis of Arabidopsis. Planta 2005, 222, 977–988. [Google Scholar] [CrossRef]
- Zuo, J.; Niu, Q.-W.; Frugis, G.; Chua, N.-H. The WUSCHEL Gene Promotes Vegetative-to-Embryonic Transition in Arabidopsis. Plant. J. 2002, 30, 349–359. [Google Scholar] [CrossRef]
- Harding, E.W. Expression and Maintenance of Embryogenic Potential Is Enhanced through Constitutive Expression of AGAMOUS-Like 15. Plant. Physiol. 2003, 133, 653–663. [Google Scholar] [CrossRef]
- Wang, X.; Niu, Q.-W.; Teng, C.; Li, C.; Mu, J.; Chua, N.-H.; Zuo, J. Overexpression of PGA37/MYB118 and MYB115 Promotes Vegetative-to-Embryonic Transition in Arabidopsis. Cell Res. 2009, 19, 224–235. [Google Scholar] [CrossRef]
- Tsuwamoto, R.; Yokoi, S.; Takahata, Y. Arabidopsis EMBRYOMAKER Encoding an AP2 Domain Transcription Factor Plays a Key Role in Developmental Change from Vegetative to Embryonic Phase. Plant. Mol. Biol. 2010, 73, 481–492. [Google Scholar] [CrossRef]
- Hobert, O. Common Logic of Transcription Factor and MicroRNA Action. Trends Biochem. Sci. 2004, 29, 462–468. [Google Scholar] [CrossRef]
- Jones-Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and Their Regulatory Roles in Plants. Annu. Rev. Plant. Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef] [PubMed]
- Szyrajew, K.; Bielewicz, D.; Dolata, J.; Wójcik, A.M.; Nowak, K.; Szczygieł-Sommer, A.; Szweykowska-Kulinska, Z.; Jarmolowski, A.; Gaj, M.D. MicroRNAs Are Intensively Regulated during Induction of Somatic Embryogenesis in Arabidopsis. Front. Plant. Sci. 2017, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-J.; Liu, Q.; Zhang, Y.-C.; Qu, L.-H.; Chen, Y.-Q.; Gautheret, D. Genome-Wide Discovery and Analysis of MicroRNAs and Other Small RNAs from Rice Embryogenic Callus. Rna Biol. 2011, 8, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, J.; Qiu, S.; Zhang, Y.; Wang, P.; Yang, L.; Lu, Y.; Shi, J. Deep Sequencing and Microarray Hybridization Identify Conserved and Species-Specific MicroRNAs during Somatic Embryogenesis in Hybrid Yellow Poplar. PLoS ONE 2012, 7, e43451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, S.; Han, S.; Wu, T.; Li, X.; Li, W.; Qi, L. Genome-Wide Identification of MicroRNAs in Larch and Stage-Specific Modulation of 11 Conserved MicroRNAs and Their Targets during Somatic Embryogenesis. Planta 2012, 236, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, L.; Yuan, D.; Lindsey, K.; Zhang, X. Small RNA and Degradome Sequencing Reveal Complex MiRNA Regulation during Cotton Somatic Embryogenesis. J. Exp. Bot. 2013, 64, 1521–1536. [Google Scholar] [CrossRef]
- Chávez-Hernández, E.C.; Alejandri-Ramírez, N.D.; Juárez-González, V.T.; Dinkova, T.D. Maize MiRNA and Target Regulation in Response to Hormone Depletion and Light Exposure during Somatic Embryogenesis. Front. Plant. Sci. 2015, 6, 555. [Google Scholar] [CrossRef]
- Mongomake, K.; Doungous, O.; Khatabi, B.; Fondong, V.N. Somatic Embryogenesis and Plant Regeneration of Cassava (Manihot esculenta Crantz) Landraces from Cameroon. SpringerPlus 2015, 4, 477. [Google Scholar] [CrossRef]
- Wójcik, A.M.; Nodine, M.D.; Gaj, M.D. miR160 and miR166/165 Contribute to the LEC2-Mediated Auxin Response Involved in the Somatic Embryogenesis Induction in Arabidopsis. Front. Plant. Sci. 2017, 8, 2024. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.H.; Liu, Y.B.; Zhou, C.; Li, X.M.; Zhang, X.S. The microRNA167 Controls Somatic Embryogenesis in Arabidopsis Through Regulating Its Target Genes ARF6 and ARF8. Plant. Cell Tiss. Organ. Cult. 2016, 124, 405–417. [Google Scholar] [CrossRef]
- Wójcik, A.M.; Gaj, M.D. miR393 Contributes to the Embryogenic Transition Induced In Vitro in Arabidopsis via the Modification of the Tissue Sensitivity to Auxin Treatment. Planta 2016, 244, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Long, J.-M.; Liu, C.-Y.; Feng, M.-Q.; Liu, Y.; Wu, X.-M.; Guo, W.-W. miR156-SPL Modules Regulate Induction of Somatic Embryogenesis in Citrus Callus. J. Exp. Bot. 2018, 69, 2979–2993. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.E.; Mecchia, M.A.; Debernardi, J.M.; Schommer, C.; Weigel, D.; Palatnik, J.F. Control of Cell Proliferation in Arabidopsis thaliana by microRNA miR396. Development 2010, 137, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.E.; Schommer, C.; Palatnik, J.F. Control of Cell Proliferation by microRNAs in Plants. Curr. Opin. Plant. Biol. 2016, 34, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.E.; Ercoli, M.F.; Debernardi, J.M.; Breakfield, N.W.; Mecchia, M.A.; Sabatini, M.; Cools, T.; De Veylder, L.; Benfey, P.N.; Palatnik, J.F. MicroRNA miR396 Regulates the Switch between Stem Cells and Transit-Amplifying Cells in Arabidopsis Roots. Plant. Cell 2015, 27, 3354–3366. [Google Scholar] [CrossRef] [PubMed]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational Identification of Plant MicroRNAs and Their Targets, Including a Stress-Induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, D.; Kende, H. The AtGRF Family of Putative Transcription Factors is Involved in Leaf and Cotyledon Growth in Arabidopsis. Plant. J. 2003, 36, 94–104. [Google Scholar] [CrossRef]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-Regulating Factors (GRFs): A Small Transcription Factor Family with Important Functions in Plant Biology. Mol. Plant. 2015, 8, 998–1010. [Google Scholar] [CrossRef] [Green Version]
- Horstman, A.; Li, M.; Heidmann, I.; Weemen, M.; Chen, B.; Muiño, J.M.; Angenent, G.C.; Boutilier, K. The BABY BOOM Transcription Factor Activates the LEC1-ABI3-FUS3-LEC2 Network to Induce Somatic Embryogenesis. Plant. Physiol. 2017, 175, 848–857. [Google Scholar] [CrossRef]
- Fehér, A. Somatic embryogenesis — Stress-Induced Remodeling of Plant Cell Fate. Biochim. Et Biophys. Acta (Bba) – Gene. Regul. Mech. 2015, 1849, 358–402. [Google Scholar] [CrossRef]
- Casadevall, R.; Rodriguez, R.E.; Debernardi, J.M.; Palatnik, J.F.; Casati, P. Repression of Growth Regulating Factors by the MicroRNA396 Inhibits Cell Proliferation by UV-B Radiation in Arabidopsis Leaves. Plant. Cell 2013, 25, 3570–3583. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-H.; Tian, X.; Li, Y.-J.; Wu, C.-A.; Zheng, C.-C. Microarray-Based Analysis of Stress-Regulated microRNAs in Arabidopsis thaliana. RNA 2008, 14, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Wójcikowska, B.; Jaskóła, K.; Gąsiorek, P.; Meus, M.; Nowak, K.; Gaj, M.D. LEAFY COTYLEDON2 (LEC2) Promotes Embryogenic Induction in Somatic Tissues of Arabidopsis, via YUCCA-Mediated Auxin Biosynthesis. Planta 2013, 238, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Zavattieri, M.A.; Frederico, A.M.; Lima, M.; Sabino, R.; Arnholdt-Schmitt, B. Induction of Somatic Embryogenesis as an Example of Stress-Related Plant Reactions. Electro. J. Biotech. 2010, 13, 1. [Google Scholar] [CrossRef]
- Kurczyńska, E.U.; Gaj, M.D.; Ujczak, A.; Mazur, E. Histological Analysis of Direct Somatic Embryogenesis in Arabidopsis thaliana (L.) Heynh. Planta 2007, 226, 619–628. [Google Scholar] [CrossRef]
- Gaj, M.D. Factors Influencing Somatic Embryogenesis Induction and Plant Regeneration with Particular Reference to Arabidopsis thaliana (L.) Heynh. Plant. Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Chen, L.; Wang, T.; Zhao, M.; Zhang, W. Ethylene-Responsive miRNAs in Roots of Medicago truncatula Identified by High-Throughput Sequencing at Whole Genome Level. Plant. Sci. 2012, 184, 14–19. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, H.; Guo, S.; Wang, B.; Li, Z.; Chong, K.; Xu, Y. OsmiR396d Affects Gibberellin and Brassinosteroid Signaling to Regulate Plant Architecture in Rice. Plant. Physiol. 2018, 176, 946–959. [Google Scholar] [CrossRef]
- Gao, F.; Wang, K.; Liu, Y.; Chen, Y.; Chen, P.; Shi, Z.; Luo, J.; Jiang, D.; Fan, F.; Zhu, Y.; et al. Blocking miR396 Increases Rice Yield by Shaping Inflorescence Architecture. Nat. Plants 2016, 2, 15196. [Google Scholar] [CrossRef]
- Wang, B.; Chu, J.; Yu, T.; Xu, Q.; Sun, X.; Yuan, J.; Xiong, G.; Wang, G.; Wang, Y.; Li, J. Tryptophan-Independent Auxin Biosynthesis Contributes to Early Embryogenesis in Arabidopsis. Proc. Natl. Acad. Sci. Usa 2015, 112, 4821–4826. [Google Scholar] [CrossRef]
- Bai, B.; Shi, B.; Hou, N.; Cao, Y.; Meng, Y.; Bian, H.; Zhu, M.; Han, N. MicroRNAs Participate in Gene Expression Regulation and Phytohormone Cross-Talk in Barley Embryo during Seed Development and Germination. Bmc Plant. Biol. 2017, 17, 150. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, Z.; Xing, L.; Wei, Y.; Mao, J.; Meng, Y.; Bao, L.; Han, M.; Zhao, C.; Zhang, D. MiRNAs Associated with Auxin Signaling, Stress Response, and Cellular Activities Mediate Adventitious Root Formation in Apple Rootstocks. Plant. Physiol. Biochem. 2019, 139, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xue, B.; Gai, M.; Song, S.; Jia, N.; Sun, H. Small RNA and Transcriptome Sequencing Reveal a Potential miRNA-Mediated Interaction Network That Functions during Somatic Embryogenesis in Lilium pumilum DC. Fisch. Front. Plant. Sci. 2017, 8, 566. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, B.H. GROWTH-REGULATING FACTOR4 of Arabidopsis thaliana Is Required for Development of Leaves, Cotyledons, and Shoot Apical Meristem. J. Plant. Biol. 2006, 49, 463–468. [Google Scholar] [CrossRef]
- Hewezi, T.; Maier, T.R.; Nettleton, D.; Baum, T.J. The Arabidopsis MicroRNA396-GRF1/GRF3 Regulatory Module Acts as a Developmental Regulator in the Reprogramming of Root Cells during Cyst Nematode Infection. Plant. Physiol. 2012, 159, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.-H.; Chiang-Hsieh, Y.-F.; Chen, Y.-A.; Chow, C.-N.; Wu, N.-Y.; Hou, P.-F.; Chang, W.-C. AtmiRNET: A Web-Based Resource for Reconstructing Regulatory Networks of Arabidopsis microRNAs. Database 2015. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ren, N.; Wang, H.; Stromberg, A.J.; Perry, S.E. Global Identification of Targets of the Arabidopsis MADS Domain Protein AGAMOUS-Like15. Plant. Cell Online 2009, 21, 2563–2577. [Google Scholar] [CrossRef]
- Yilmaz, A.; Mejia-Guerra, M.K.; Kurz, K.; Liang, X.; Welch, L.; Grotewold, E. AGRIS: The Arabidopsis Gene Regulatory Information Server, an update. Nucleic Acids Res. 2011, 39, D1118–D1122. [Google Scholar] [CrossRef]
- Stone, S.L.; Kwong, L.W.; Yee, K.M.; Pelletier, J.; Lepiniec, L.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON2 Encodes a B3 Domain Transcription Factor that Induces Embryo Development. Proc. Natl. Acad. Sci. 2001, 98, 11806–11811. [Google Scholar] [CrossRef]
- Gaj, M.D. Direct Somatic Embryogenesis as a Rapid and Efficient System for In Vitro Regeneration of Arabidopsis thaliana. Plant. Cell Tissue Organ. Cult. 2001, 64, 39–46. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient Requirements of Suspension Cultures of Soybean Root Cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Bric, J.M.; Bostock, R.M.; SILVERSTONEt, S.E. Rapid In Situ Assay for Indoleacetic Acid Production by Bacteria Immobilized on a Nitrocellulose Membrane. Appl. Env. Microbiol. 1991, 57, 535–538. [Google Scholar] [Green Version]
Figure 1.
The spatiotemporal expression pattern of MIR396b in immature zygotic embryo (IZE) explants that were cultured for (a) 0, (b) 5, (c) 10, and (d) 21 days on a standard somatic embryogenesis (SE) induction medium (E5). Cotyledons and the shoot apical meristem (SAM) area of the IZE, i.e., the explant parts that were involved in SE induction, showed a β-glucuronidase (GUS) signal (a–c). Similarly to the IZEs (a), the somatic embryos (d) had a GUS signal in the cotyledons. Arrows point to the GUS signal. Scale bar: 0.2 mm (a,b), 1 mm (c,d).
Figure 1.
The spatiotemporal expression pattern of MIR396b in immature zygotic embryo (IZE) explants that were cultured for (a) 0, (b) 5, (c) 10, and (d) 21 days on a standard somatic embryogenesis (SE) induction medium (E5). Cotyledons and the shoot apical meristem (SAM) area of the IZE, i.e., the explant parts that were involved in SE induction, showed a β-glucuronidase (GUS) signal (a–c). Similarly to the IZEs (a), the somatic embryos (d) had a GUS signal in the cotyledons. Arrows point to the GUS signal. Scale bar: 0.2 mm (a,b), 1 mm (c,d).
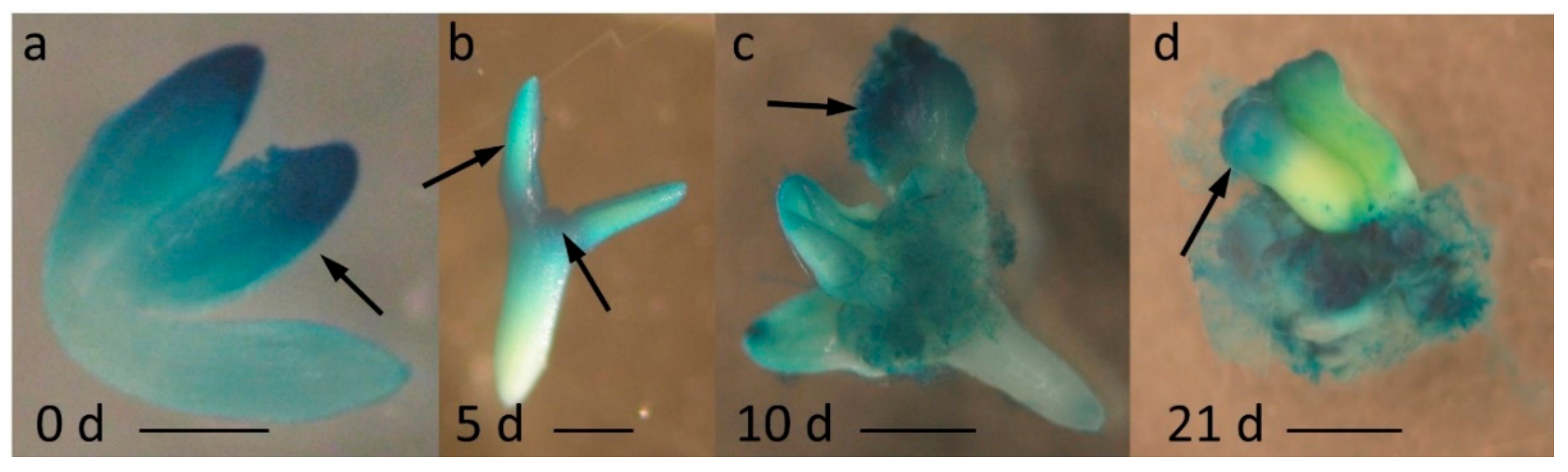
Figure 2.
The embryogenic potential of the explants in relation to the miR396 expression level and auxin treatment. Explants of Col-0 (wild type - WT): (a,b) mir396a, (c,d) mir396b, and (e,f) 35S:MIR396b were cultured on an SE induction medium that had been supplemented with different concentrations of 2,4-Dichlorophenoxyacetic acid (2,4-D). The efficiency (a,c,e) and productivity (b,d,f) of the SE were evaluated. Values that were significantly different from those of the WT culture are indicated with asterisks (*) (p < 0.05; n = 3 ± SD).
Figure 2.
The embryogenic potential of the explants in relation to the miR396 expression level and auxin treatment. Explants of Col-0 (wild type - WT): (a,b) mir396a, (c,d) mir396b, and (e,f) 35S:MIR396b were cultured on an SE induction medium that had been supplemented with different concentrations of 2,4-Dichlorophenoxyacetic acid (2,4-D). The efficiency (a,c,e) and productivity (b,d,f) of the SE were evaluated. Values that were significantly different from those of the WT culture are indicated with asterisks (*) (p < 0.05; n = 3 ± SD).
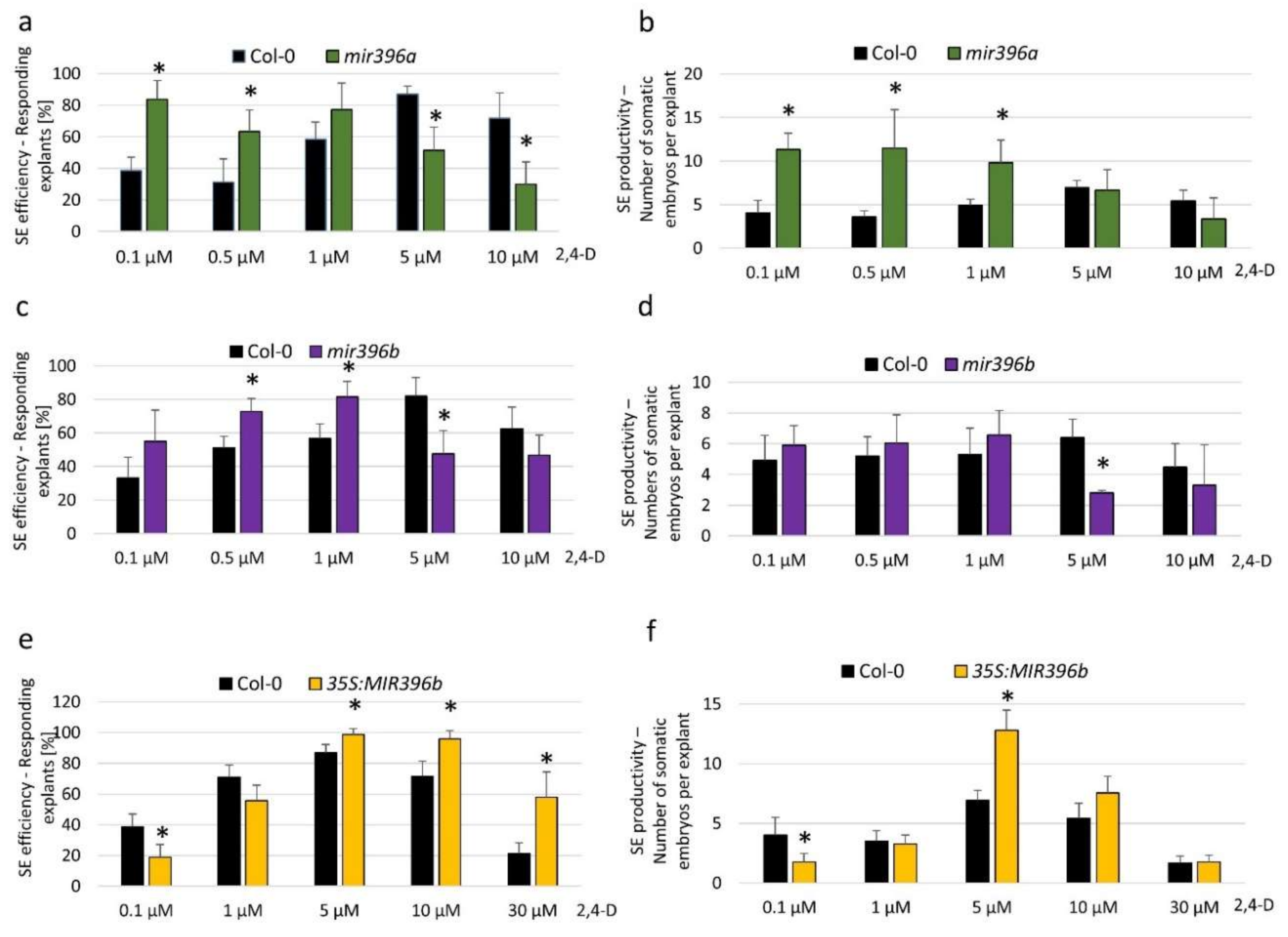
Figure 3.
The auxin-related function of miR396 in an SE culture. Explants of mir396a, mir396b, 35S:MIR396b, and Col-0 (WT) were cultured on a standard SE induction medium (E5). (a) The relative content of the indolic compounds and the expression level of the auxin biosynthesis of the (b) YUC1 and (c) YUC4 genes were evaluated in the SE-induced explants. The relative transcript level was normalized to the internal control (At4g27090) and calibrated to a Col-0 culture of the same age. The bars represent the standard deviation. Values that were significantly different from those of a WT culture of the same age are indicated with asterisks (*) (p < 0.05; n = 3 ± SD).
Figure 3.
The auxin-related function of miR396 in an SE culture. Explants of mir396a, mir396b, 35S:MIR396b, and Col-0 (WT) were cultured on a standard SE induction medium (E5). (a) The relative content of the indolic compounds and the expression level of the auxin biosynthesis of the (b) YUC1 and (c) YUC4 genes were evaluated in the SE-induced explants. The relative transcript level was normalized to the internal control (At4g27090) and calibrated to a Col-0 culture of the same age. The bars represent the standard deviation. Values that were significantly different from those of a WT culture of the same age are indicated with asterisks (*) (p < 0.05; n = 3 ± SD).
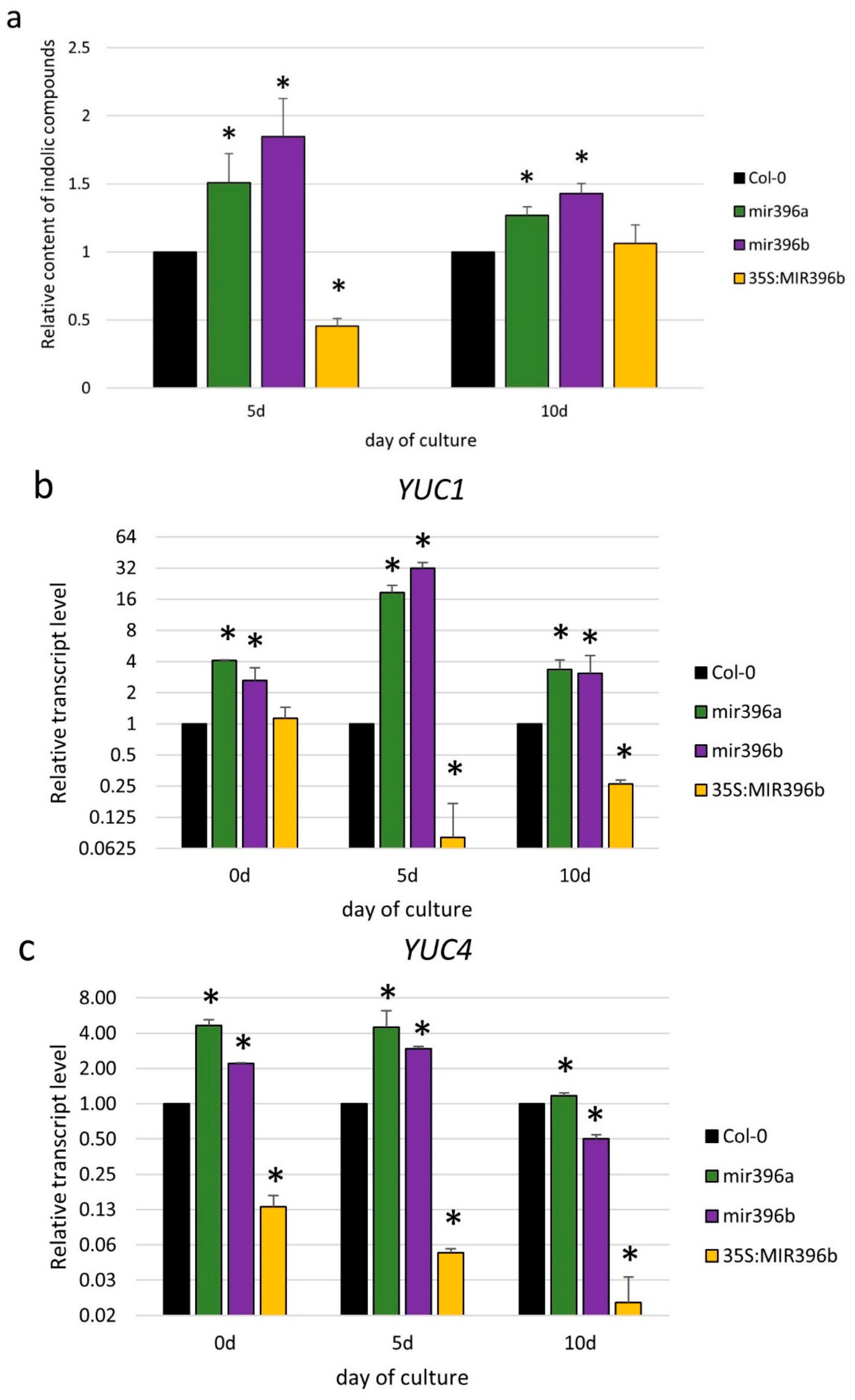
Figure 4.
The regulatory relationship between miR396 and the GRF genes during SE. The expression of (a) the GRF genes in the SE culture of Col-0 and (b) GRF1, (c) GRF4, (d) GRF7, (e) GRF8, and (f) GRF9 in the SE cultures of the mir396a, the mir396b mutants, and the 35S:MIR396b line. The explants were induced on a standard SE induction medium (E5). The relative transcript level was normalized to the internal control (At4g27090) and calibrated to a WT culture of the same age. Values that were significantly different from those of a WT culture of the same age are indicated with asterisks (*) (p < 0.05; n = 3 ± SD).
Figure 4.
The regulatory relationship between miR396 and the GRF genes during SE. The expression of (a) the GRF genes in the SE culture of Col-0 and (b) GRF1, (c) GRF4, (d) GRF7, (e) GRF8, and (f) GRF9 in the SE cultures of the mir396a, the mir396b mutants, and the 35S:MIR396b line. The explants were induced on a standard SE induction medium (E5). The relative transcript level was normalized to the internal control (At4g27090) and calibrated to a WT culture of the same age. Values that were significantly different from those of a WT culture of the same age are indicated with asterisks (*) (p < 0.05; n = 3 ± SD).
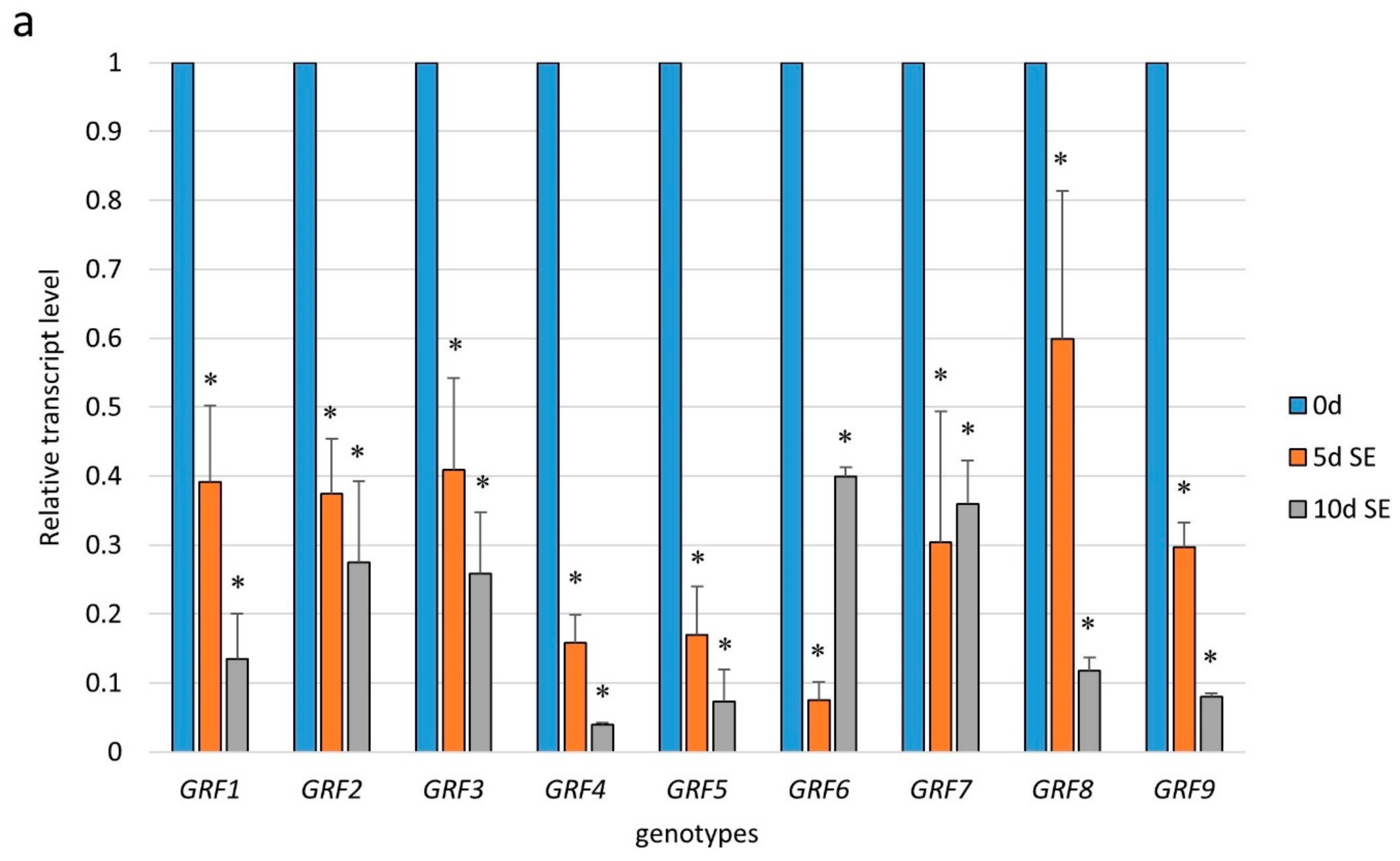
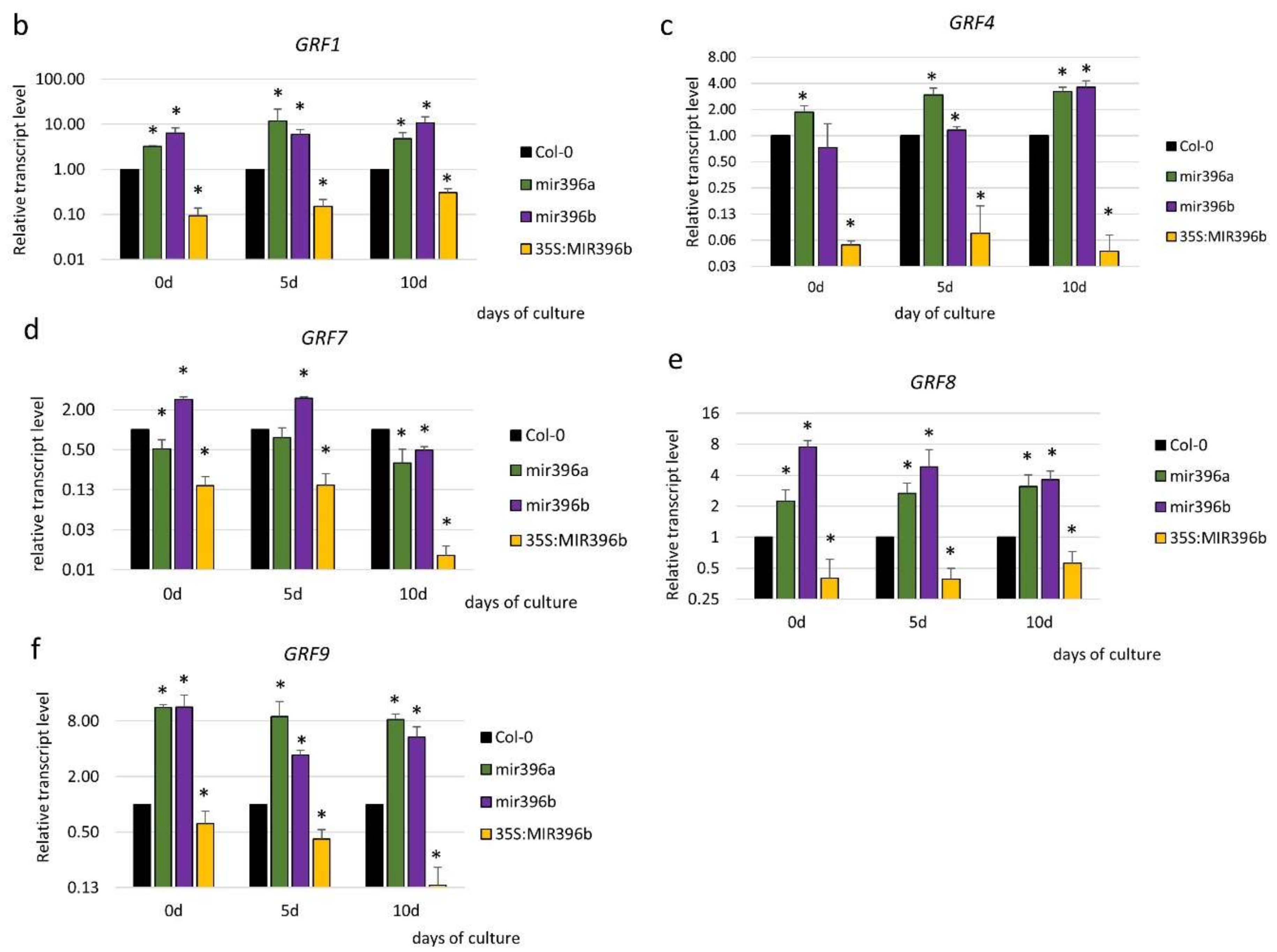
Figure 5.
GRF1 and GRF4 controlled the embryogenic response of the explants that had been cultured on the auxin media. Explants of (a,b) the 35S:GRF1 line, (c,d) the grf4 mutant, and their WT genotypes, (Wassilewskija (WS) and Col-0, relevantly) were cultured on an SE induction medium that had been supplemented with different concentrations of 2,4-D. (a,c) The efficiency and (b,d) productivity of SE were evaluated in a 21-day-old culture. Values that were significantly different from those of the WT culture are indicated with asterisks (*) p < 0.05; n = 3 ± SD).
Figure 5.
GRF1 and GRF4 controlled the embryogenic response of the explants that had been cultured on the auxin media. Explants of (a,b) the 35S:GRF1 line, (c,d) the grf4 mutant, and their WT genotypes, (Wassilewskija (WS) and Col-0, relevantly) were cultured on an SE induction medium that had been supplemented with different concentrations of 2,4-D. (a,c) The efficiency and (b,d) productivity of SE were evaluated in a 21-day-old culture. Values that were significantly different from those of the WT culture are indicated with asterisks (*) p < 0.05; n = 3 ± SD).
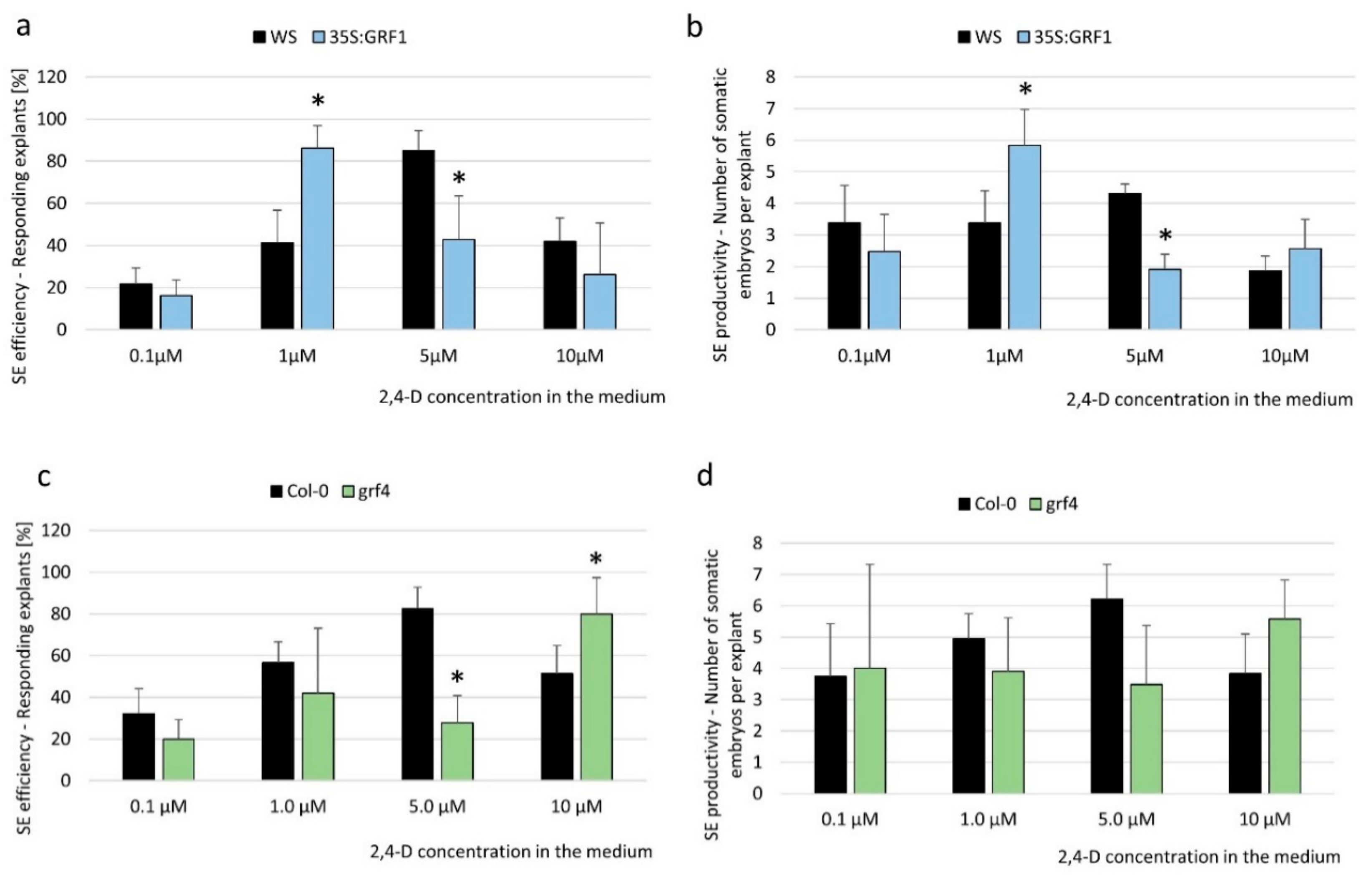
Figure 6.
PLT2 controlled the embryogenic response of the explants that were cultured on the auxin medium. (a) The explants of Col-0 (WT), the plt2 mutant, and the 35S:PLT2-GR line were cultured for 21 days on a standard E5 medium, after which the levels of (b) efficiency and (c) productivity of SE were evaluated. Dexamethasone (DEX) treatment (+DEX) was used to induce PLT2 overexpression in the 35S:PLT2-GR explants (35S:PLT2-GR + DEX). The bars represent the standard deviation. Values that were significantly different from those for the WT culture are indicated with asterisks (*) (p < 0.05; n = 3 ± SD). Scale bar: 1 mm.
Figure 6.
PLT2 controlled the embryogenic response of the explants that were cultured on the auxin medium. (a) The explants of Col-0 (WT), the plt2 mutant, and the 35S:PLT2-GR line were cultured for 21 days on a standard E5 medium, after which the levels of (b) efficiency and (c) productivity of SE were evaluated. Dexamethasone (DEX) treatment (+DEX) was used to induce PLT2 overexpression in the 35S:PLT2-GR explants (35S:PLT2-GR + DEX). The bars represent the standard deviation. Values that were significantly different from those for the WT culture are indicated with asterisks (*) (p < 0.05; n = 3 ± SD). Scale bar: 1 mm.
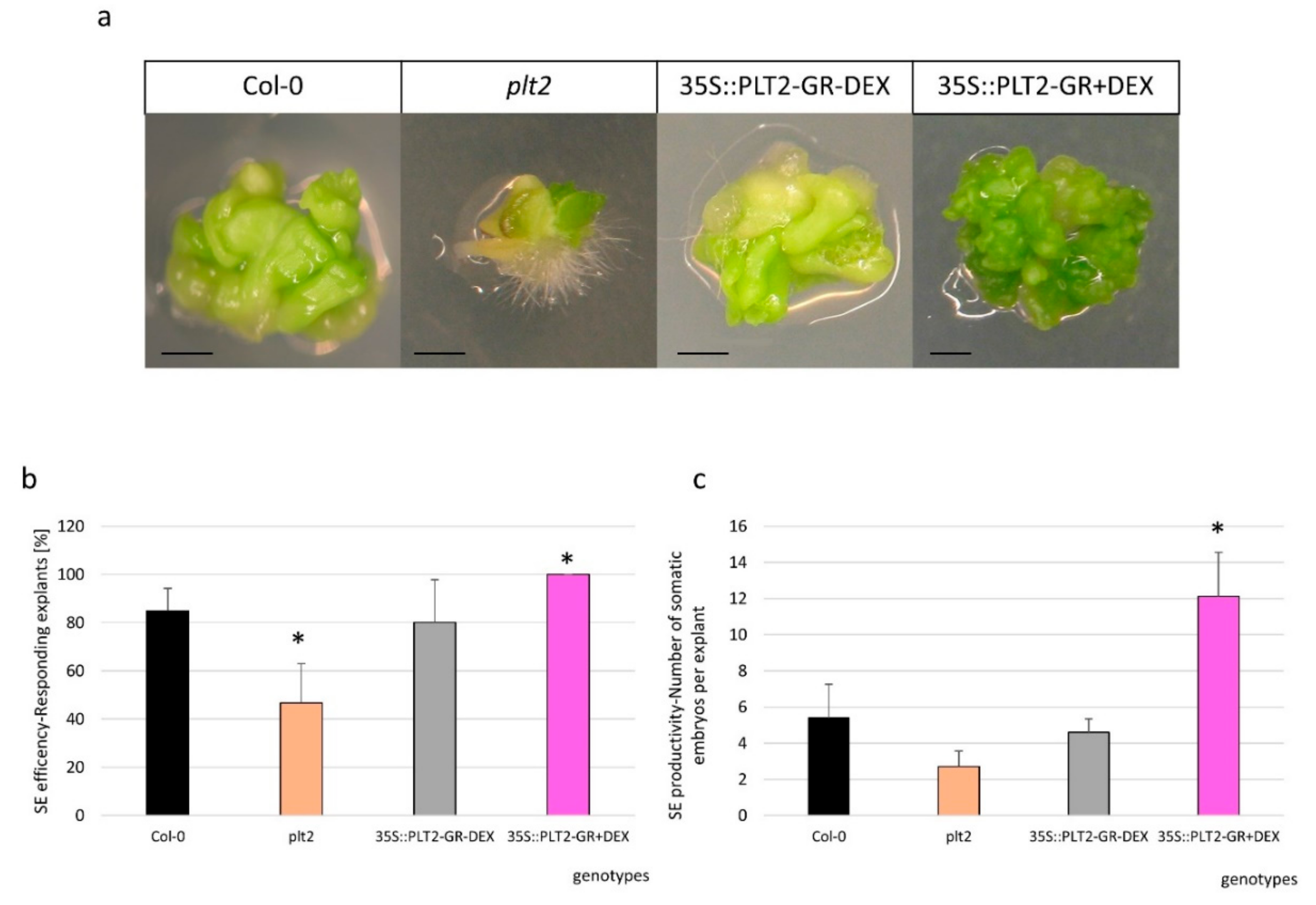
Figure 7.
The regulatory relationship within the miR396–GROWTH REGULATING FACTOR (GRF)–PLETHORA (PLT) module. (a) The expression level of the PLT1 and (b) PLT2 genes in the cultures of the mir396 mutants and the 35S:MIR396b line; (c) the decreased accumulation of miR396 in the culture of the plt2 mutant; (d) the increased expression level of GRFs (GRF1, GRF2, GRF3, GRF4, GRF7, GRF8, and GRF9) in the culture of the plt2 mutant. The explants were cultured on a standard E5 medium. The relative transcript level was normalized to the internal control (At4g27090) and calibrated to a WT (Col-0) culture of the same age. The bars represent the standard deviation. Values that were significantly different from those of the WT culture are indicated with asterisks (*) (p < 0.05; n = 3 ± SD).
Figure 7.
The regulatory relationship within the miR396–GROWTH REGULATING FACTOR (GRF)–PLETHORA (PLT) module. (a) The expression level of the PLT1 and (b) PLT2 genes in the cultures of the mir396 mutants and the 35S:MIR396b line; (c) the decreased accumulation of miR396 in the culture of the plt2 mutant; (d) the increased expression level of GRFs (GRF1, GRF2, GRF3, GRF4, GRF7, GRF8, and GRF9) in the culture of the plt2 mutant. The explants were cultured on a standard E5 medium. The relative transcript level was normalized to the internal control (At4g27090) and calibrated to a WT (Col-0) culture of the same age. The bars represent the standard deviation. Values that were significantly different from those of the WT culture are indicated with asterisks (*) (p < 0.05; n = 3 ± SD).
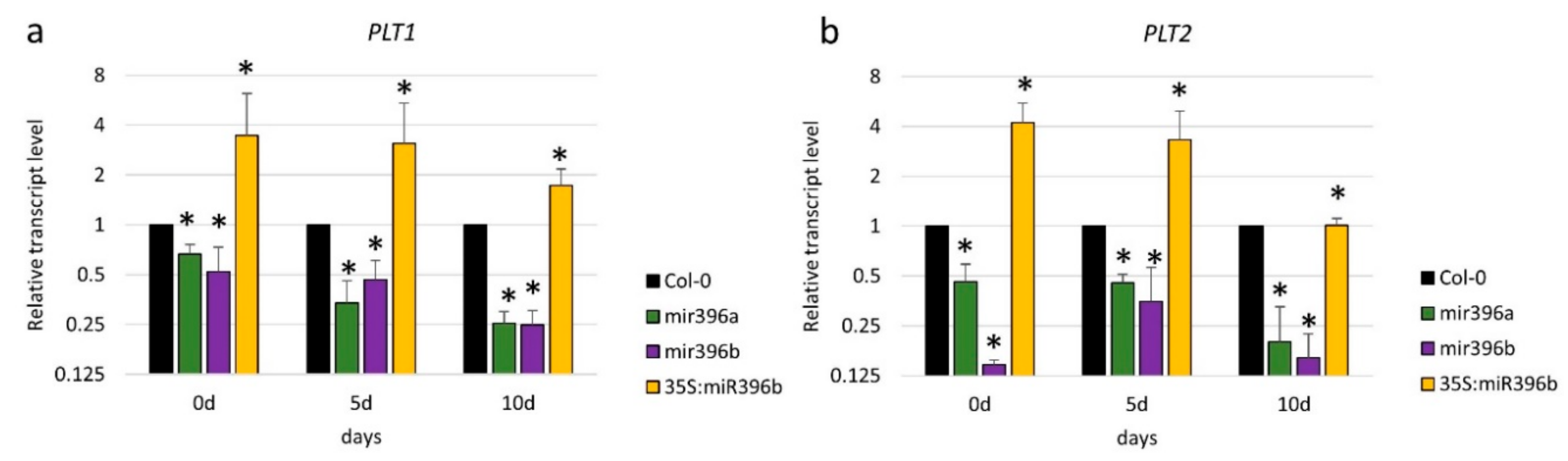
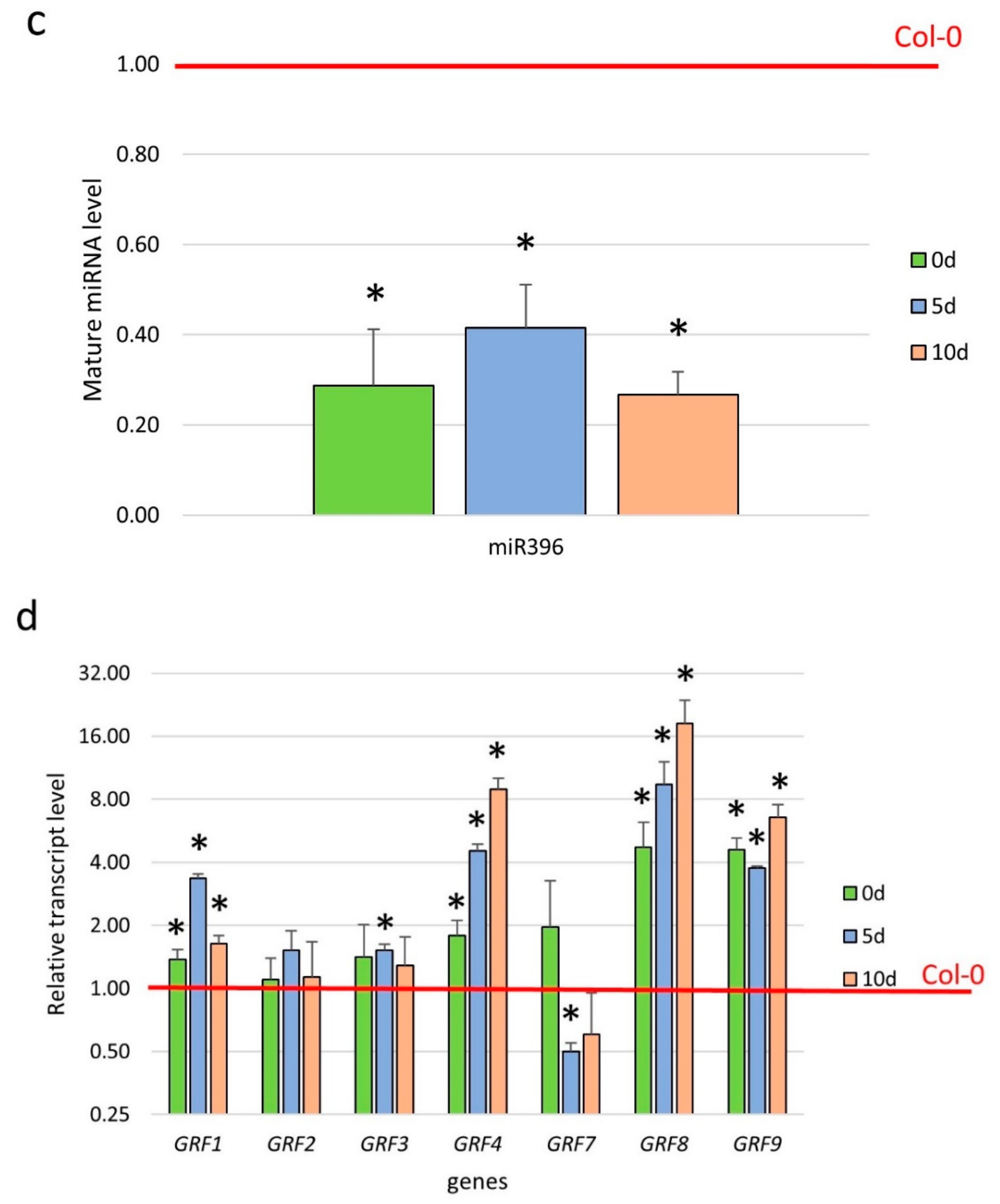
Figure 8.
A model of the miR396-controlled network that is involved in auxin-regulated SE induction in Arabidopsis: 2,4-D treatment increased the accumulation of miR396 in the explants by activating the auxin-responsive MIR396 genes. As a result, the GRF genes (GRF1/4/8/9) were repressed, the PLT genes (PLT1/PLT2/BBM) were activated, and, in turn, the LEC2 gene, which is a positive regulator of the YUC (YUC1 and YUC4) genes that are engaged in auxin biosynthesis during SE, was upregulated [25,32,44,47]. A feedback regulatory loop between PLT and miR396 might also be expected [25].
Figure 8.
A model of the miR396-controlled network that is involved in auxin-regulated SE induction in Arabidopsis: 2,4-D treatment increased the accumulation of miR396 in the explants by activating the auxin-responsive MIR396 genes. As a result, the GRF genes (GRF1/4/8/9) were repressed, the PLT genes (PLT1/PLT2/BBM) were activated, and, in turn, the LEC2 gene, which is a positive regulator of the YUC (YUC1 and YUC4) genes that are engaged in auxin biosynthesis during SE, was upregulated [25,32,44,47]. A feedback regulatory loop between PLT and miR396 might also be expected [25].
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szczygieł-Sommer, A.; Gaj, M.D. The miR396–GRF Regulatory Module Controls the Embryogenic Response in Arabidopsis via an Auxin-Related Pathway. Int. J. Mol. Sci. 2019, 20, 5221. https://doi.org/10.3390/ijms20205221
AMA Style
Szczygieł-Sommer A, Gaj MD. The miR396–GRF Regulatory Module Controls the Embryogenic Response in Arabidopsis via an Auxin-Related Pathway. International Journal of Molecular Sciences. 2019; 20(20):5221. https://doi.org/10.3390/ijms20205221
Chicago/Turabian StyleSzczygieł-Sommer, Aleksandra, and Małgorzata D. Gaj. 2019. "The miR396–GRF Regulatory Module Controls the Embryogenic Response in Arabidopsis via an Auxin-Related Pathway" International Journal of Molecular Sciences 20, no. 20: 5221. https://doi.org/10.3390/ijms20205221
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.