The Insulin/IGF System in Mammalian Sexual Development and Reproduction

Abstract
1. Introduction
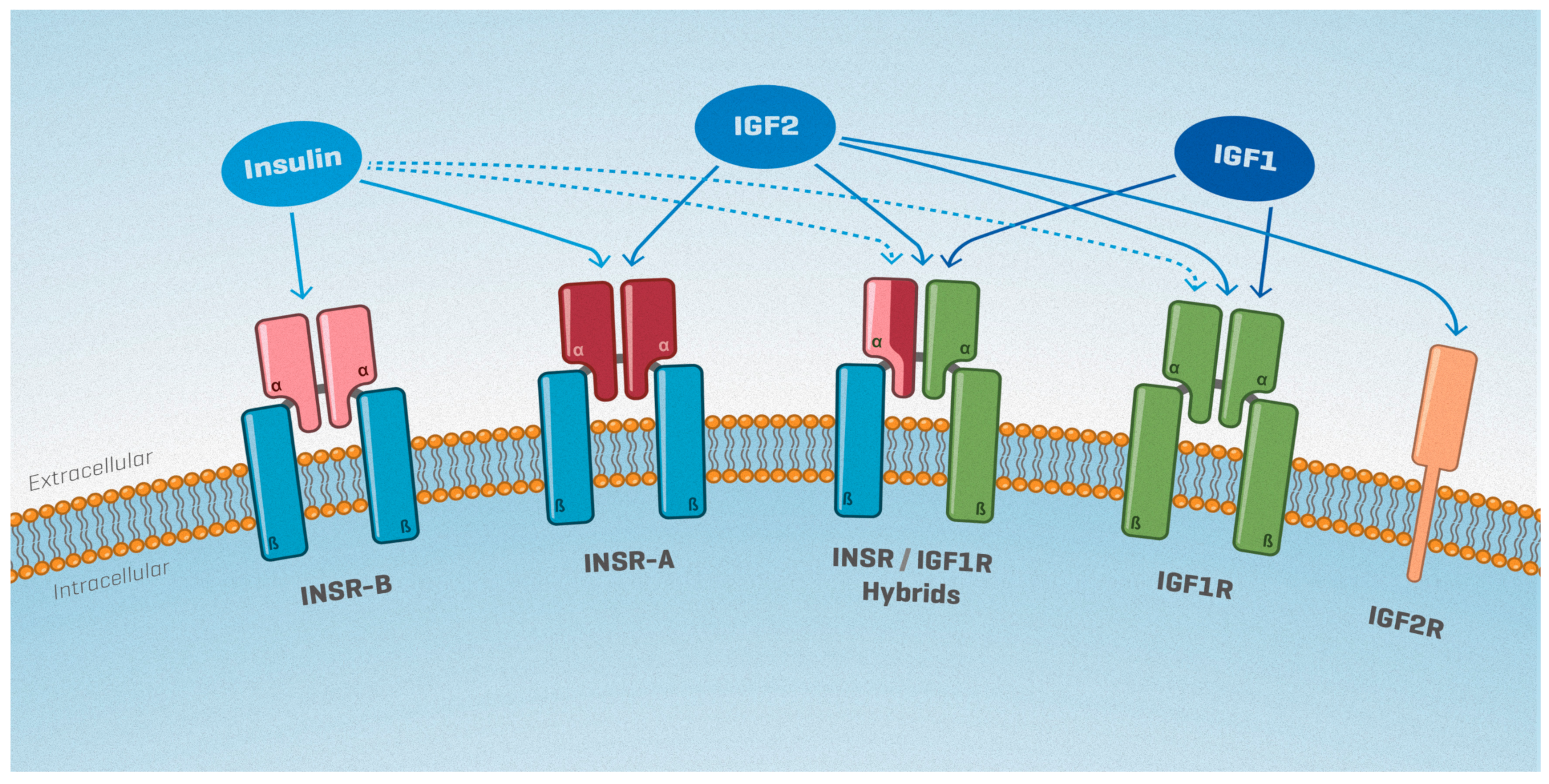
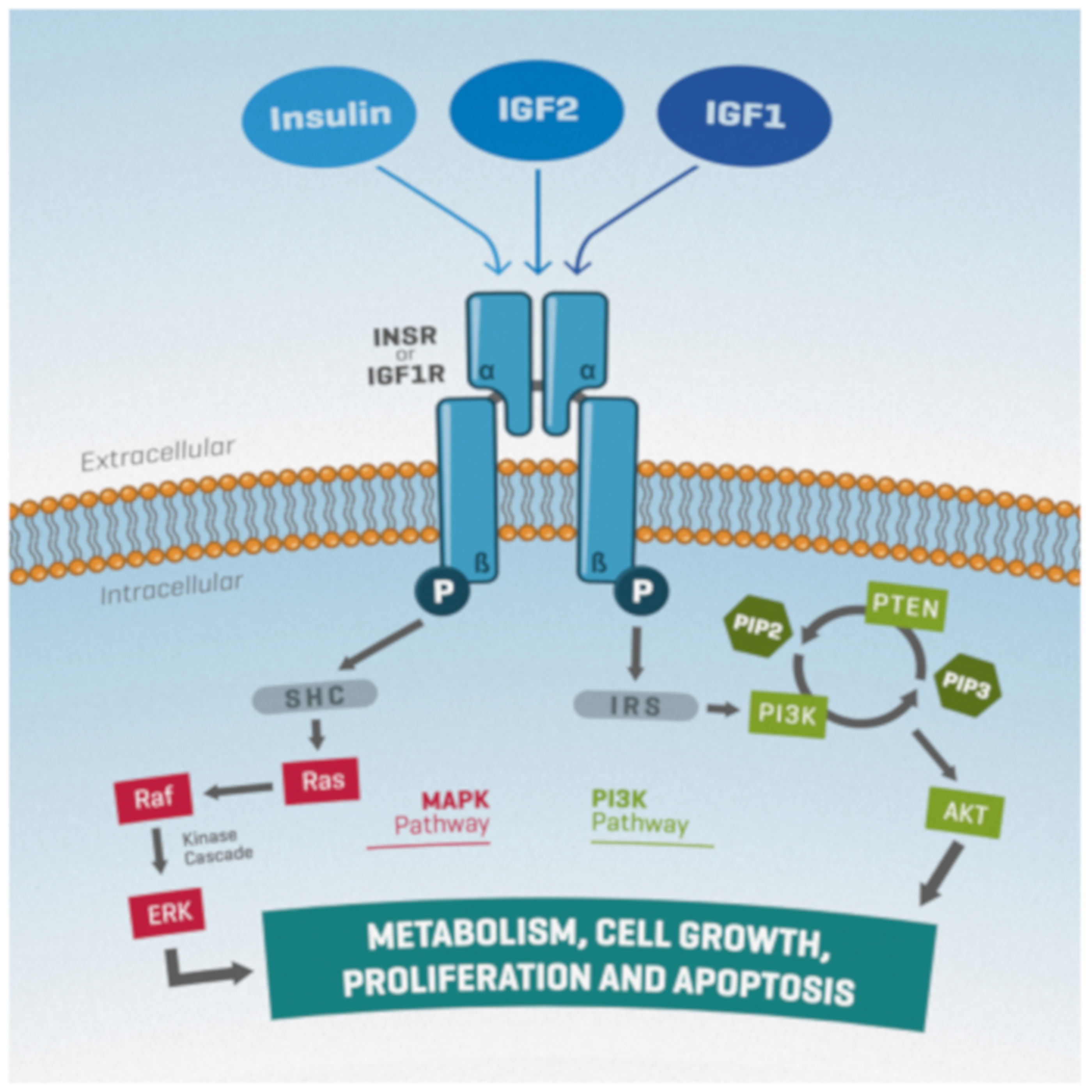
2. The Insulin/IGF System
3. Genital Ridge Formation, Sex Determination and Sex-Specific Gonadal Development
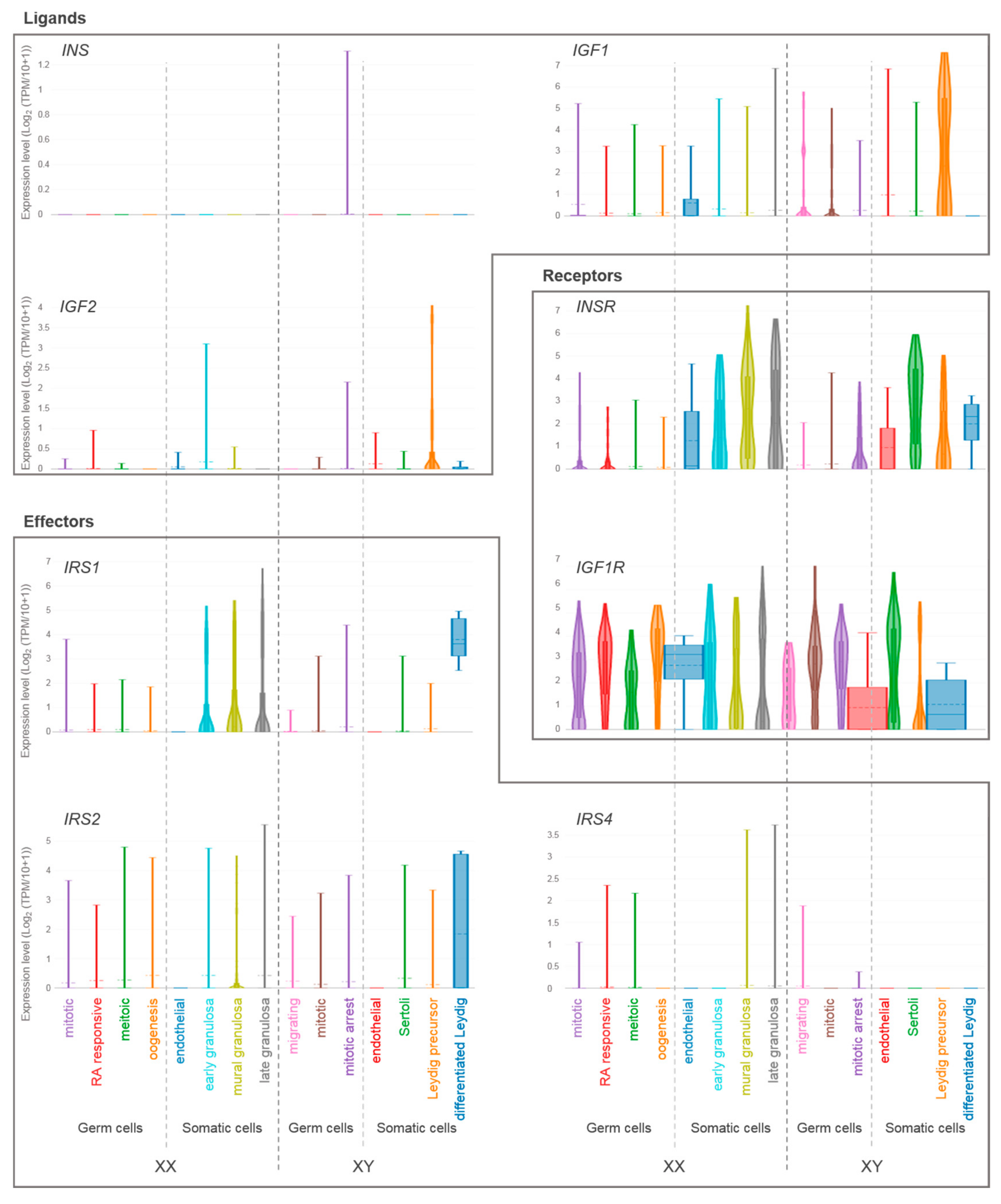
4. Expression of Insulin/IGF Family Members in the Gonads of Mice and Humans
5. Biological Effects of the Insulin/IGF System in Sexual Development and Reproduction
5.1. Differential Contribution of IGF Ligands to Sexual Development and Reproduction
5.2. The Insulin/IGF System and Sex Determination
5.3. The Role of the Insulin/IGF System in Sertoli and Granulosa Cells
5.4. The Role of the Insulin/IGF System in Steroidogenic Lineages: FLCs, ALCs and Adrenal Glands
5.5. Cell-Autonomous Insulin/IGF Signaling Does Not Play an Essential Role in Germ Cell Development and Gamete Production
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | Protein kinase B |
| ALC | Adult Leydig cell |
| ATP | Adenosine triphosphate |
| CTNNB1 | Catenin (cadherin associated protein), beta 1 |
| CYP19A1 | Cytochrome P450, family 19, subfamily a, polypeptide 1 |
| E | Embryonic day |
| ERK | Extracellular signal-regulated kinase |
| FGF9 | Fibroblast growth factor 9 |
| FLC | Fetal Leydig cell |
| FOXL2 | Forkhead box L2 |
| FSH | Follicle stimulating hormone |
| HCG | Human chorionic gonadotropin |
| HPG | Hypothalamic–pituitary–gonadal |
| GC | Germ cell |
| GH | Growth hormone |
| IGFs | Insulin-like growth factors |
| IGF1R | Insulin-like growth factor 1 receptor |
| IGF2R | Insulin-like growth factor 2 receptor |
| IGFBP | Insulin-like growth factor binding protein |
| INS | Insulin |
| INSR | Insulin receptor |
| INSR-A | Insulin receptor isoform A |
| INSR-B | Insulin receptor isoform B |
| IRR | Insulin receptor-related receptor |
| IRS | Insulin receptor substrate |
| JAK | Janus-family tyrosine kinase |
| LC | Leydig cell |
| M6P/IGF2R | Mannose-6-phosphate/IGF2 receptor |
| MAPK | Mitogen-activated protein kinase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NR0B1 | Nuclear receptor subfamily 0 group B member 1 (also known as DAX1) |
| NR5A1 | Nuclear receptor subfamily 5, group A, member 1 (also known as Sf1) |
| P | Postnatal day |
| PGC | Primordial germ cell |
| PI3K | Phosphatidylinositol-3 kinase |
| PIP2 | Phosphatidylinositol 4,5-bisphosphate |
| PIP3 | Phosphatidylinositol 3,4,5-trisphosphate |
| PLC | Progenitor Leydig cell |
| PTEN | Phosphatase and tensin homolog deleted on chromosome 10 |
| RA | Retinoic acid |
| RAF | Rapidly accelerated fibrosarcoma |
| RSPO1 | R-spondin 1 |
| SC | Sertoli cell |
| SHC | Src homology domain-containing |
| siRNA | Small interfering RNA |
| SOX9 | SRY (sex determining region Y)-box 9 |
| SRY | Sex determining region of chromosome Y |
| STAT | Signal transducer and activator of transcription |
| TPM | Transcripts per million |
| WNT4 | Wingless-type MMTV integration site family, member 4 |
References
- Michalakis, K.; Mintziori, G.; Kaprara, A.; Tarlatzis, B.C.; Goulis, D.G. The complex interaction between obesity, metabolic syndrome and reproductive axis: A narrative review. Metabolism 2013, 62, 457–478. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Flatt, T.; Aguilaniu, H. Reproduction, fat metabolism, and life span: What is the connection? Cell Metab. 2013, 17, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Efstratiadis, A. Genetics of mouse growth. Int. J. Dev. Biol. 1998, 42, 955–976. [Google Scholar] [PubMed]
- Kaprara, A.; Huhtaniemi, I.T. The hypothalamus-pituitary-gonad axis: Tales of mice and men. Metabolism 2018, 86, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Agrogiannis, G.D.; Sifakis, S.; Patsouris, E.S.; Konstantinidou, A.E. Insulin-like growth factors in embryonic and fetal growth and skeletal development (Review). Mol. Med. Rep. 2014, 10, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Le Roith, D.; Bondy, C.; Yakar, S.; Liu, J.-L.; Butler, A. The Somatomedin Hypothesis: 2001. Endocr. Rev. 2001, 22, 53–74. [Google Scholar] [CrossRef] [PubMed]
- Lewitt, M.S.; Boyd, G.W. The Role of Insulin-Like Growth Factors and Insulin-Like Growth Factor-Binding Proteins in the Nervous System. Biochem. Insights 2019, 12, 1178626419842176. [Google Scholar] [CrossRef]
- Gallagher, E.J.; LeRoith, D. The proliferating role of insulin and insulin-like growth factors in cancer. Trends Endocrinol. Metab. 2010, 21, 610–618. [Google Scholar] [CrossRef]
- Frasca, F.; Pandini, G.; Scalia, P.; Sciacca, L.; Mineo, R.; Costantino, A.; Goldfine, I.D.; Belfiore, A.; Vigneri, R. Insulin receptor isoform A, a newly recognized, high-affinity insulin-like growth factor II receptor in fetal and cancer cells. Mol. Cell. Biol. 1999, 19, 3278–3288. [Google Scholar] [CrossRef]
- Boone, D.N.; Lee, A.V. Targeting the insulin-like growth factor receptor: Developing biomarkers from gene expression profiling. Crit. Rev. Oncog. 2012, 17, 161–173. [Google Scholar] [CrossRef]
- Baker, J.; Liu, J.P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef]
- Dinchuk, J.E.; Cao, C.; Huang, F.; Reeves, K.A.; Wang, J.; Myers, F.; Cantor, G.H.; Zhou, X.; Attar, R.M.; Gottardis, M.; et al. Insulin receptor (IR) pathway hyperactivity in IGF-IR null cells and suppression of downstream growth signaling using the dual IGF-IR/IR inhibitor, BMS-754807. Endocrinology 2010, 151, 4123–4132. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.P.; Baker, J.; Perkins, A.S.; Robertson, E.J.; Efstratiadis, A. Mice carrying null mutations of the genes encoding insulin-like growth factor I (Igf-1) and type 1 IGF receptor (Igf1r). Cell 1993, 75, 59–72. [Google Scholar] [CrossRef]
- Louvi, A.; Accili, D.; Efstratiadis, A. Growth-promoting interaction of IGF-II with the insulin receptor during mouse embryonic development. Dev. Biol. 1997, 189, 33–48. [Google Scholar] [CrossRef]
- Zhang, H.; Pelzer, A.M.; Kiang, D.T.; Yee, D. Down-regulation of Type I Insulin-like Growth Factor Receptor Increases Sensitivity of Breast Cancer Cells to Insulin. Cancer Res. 2007, 67, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Dahms, N.M.; Kornfeld, S. Mannose 6-phosphate receptors: New twists in the tale. Nat. Rev. Mol. Cell Biol. 2003, 4, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.; Petnga, W.; Macaulay, V.M.; Weyer-Czernilofsky, U.; Bogenrieder, T. Insulin-Like Growth Factor (IGF) Pathway Targeting in Cancer: Role of the IGF Axis and Opportunities for Future Combination Studies. Target. Oncol. 2017, 12, 571–597. [Google Scholar] [CrossRef] [PubMed]
- Himpe, E.; Kooijman, R. Insulin-like growth factor-I receptor signal transduction and the Janus Kinase/Signal Transducer and Activator of Transcription (JAK-STAT) pathway. BioFactors 2009, 35, 76–81. [Google Scholar] [CrossRef]
- Sarfstein, R.; Werner, H. Minireview: Nuclear insulin and insulin-like growth factor-1 receptors: A novel paradigm in signal transduction. Endocrinology 2013, 154, 1672–1679. [Google Scholar] [CrossRef]
- Nef, S.; Stévant, I.; Greenfield, A. Characterizing the bipotential mammalian gonad. Curr. Top. Dev. Biol. 2019, 134, 167–194. [Google Scholar]
- Stévant, I.; Nef, S. Genetic Control of Gonadal Sex Determination and Development. Trends Genet. 2019, 35, 346–358. [Google Scholar] [CrossRef]
- Morohashi, K. The ontogenesis of the steroidogenic tissues. Genes Cells 1997, 2, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Gubbay, J.; Collignon, J.; Koopman, P.; Capel, B.; Economou, A.; Münsterberg, A.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. A gene mapping to the sex-determining region of the mouse Y chromosome is a member of a novel family of embryonically expressed genes. Nature 1990, 346, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Hacker, A.; Capel, B.; Goodfellow, P.; Lovell-Badge, R. Expression of Sry, the mouse sex determining gene. Development 1995, 121, 1603–1614. [Google Scholar] [PubMed]
- Koopman, P.; Münsterberg, A.; Capel, B.; Vivian, N.; Lovell-Badge, R. Expression of a candidate sex-determining gene during mouse testis differentiation. Nature 1990, 348, 450–452. [Google Scholar] [CrossRef]
- Chassot, A.A.; Gregoire, E.P.; Magliano, M.; Lavery, R.; Chaboissier, M.C. Genetics of Ovarian Differentiation: Rspo1, a Major Player. Sex. Dev. 2008, 2, 219–227. [Google Scholar] [CrossRef]
- Potter, S.J.; Kumar, D.L.; Defalco, T. Origin and Differentiation of Androgen- Producing Cells in the Gonads. Chin. Phys. 2016, 58, 101–134. [Google Scholar]
- Shima, Y.; Morohashi, K. Leydig progenitor cells in fetal testis. Mol. Cell. Endocrinol. 2017, 445, 55–64. [Google Scholar] [CrossRef]
- Rotgers, E.; Jørgensen, A.; Yao, H.H.-C. At the Crossroads of Fate—Somatic Cell Lineage Specification in the Fetal Gonad. Endocr. Rev. 2018, 39, 739–759. [Google Scholar] [CrossRef]
- Agrimson, K.S.; Hogarth, C.A. Germ Cell Commitment to Oogenic Versus Spermatogenic Pathway: The Role of Retinoic Acid. Results Probl. Cell Differ. 2016, 58, 135–166. [Google Scholar]
- Spiller, C.; Bowles, J. Sexually dimorphic germ cell identity in mammals. Curr. Top. Dev. Biol. 2019, 134, 253–288. [Google Scholar] [PubMed]
- Jégou, B. The Sertoli cell. Baillieres. Clin. Endocrinol. Metab. 1992, 6, 273–311. [Google Scholar] [CrossRef]
- Mruk, D.D.; Cheng, C.Y. Sertoli-Sertoli and Sertoli-germ cell interactions and their significance in germ cell movement in the seminiferous epithelium during spermatogenesis. Endocr. Rev. 2004, 25, 747–806. [Google Scholar] [CrossRef] [PubMed]
- Rebourcet, D.; Darbey, A.; Monteiro, A.; Soffientini, U.; Tsai, Y.T.; Handel, I.; Pitetti, J.-L.; Nef, S.; Smith, L.B.; O’Shaughnessy, P.J. Sertoli Cell Number Defines and Predicts Germ and Leydig Cell Population Sizes in the Adult Mouse Testis. Endocrinology 2017, 158, 2955–2969. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, A.J.W.; Kawamura, K.; Cheng, Y.; Fauser, B.C.J.M. Intraovarian control of early folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; Holzenberger, M. Biology of insulin-like growth factors in development. Birth Defects Res. Part C Embryo Today Rev. 2003, 69, 257–271. [Google Scholar] [CrossRef] [PubMed]
- DeChiara, T.M.; Robertson, E.J.; Efstratiadis, A. Parental imprinting of the mouse insulin-like growth factor II gene. Cell 1991, 64, 849–859. [Google Scholar] [CrossRef]
- Peters, J. The role of genomic imprinting in biology and disease: An expanding view. Nat. Rev. Genet. 2014, 15, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Mann, J.R. Imprinting in the Germ Line. STEM CELLS 2001, 19, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Lui, J.C.; Finkielstain, G.P.; Barnes, K.M.; Baron, J. An imprinted gene network that controls mammalian somatic growth is down-regulated during postnatal growth deceleration in multiple organs. Am. J. Physiol. Integr. Comp. Physiol. 2008, 295, R189–R196. [Google Scholar] [CrossRef]
- Ren, H.; Yin, P.; Duan, C. IGFBP-5 regulates muscle cell differentiation by binding to IGF-II and switching on the IGF-II auto-regulation loop. J. Cell Biol. 2008, 182, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Neirijnck, Y.; Kühne, F.; Mayère, C.; Pavlova, E.; Sararols, P.; Foti, M.; Atanassova, N.; Nef, S. Tumor Suppressor PTEN Regulates Negatively Sertoli Cell Proliferation, Testis Size, and Sperm Production In Vivo. Endocrinology 2019, 160, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Pitetti, J.-L.; Calvel, P.; Zimmermann, C.; Conne, B.; Papaioannou, M.D.; Aubry, F.; Cederroth, C.R.; Urner, F.; Fumel, B.; Crausaz, M.; et al. An essential role for insulin and IGF1 receptors in regulating sertoli cell proliferation, testis size, and FSH action in mice. Mol. Endocrinol. 2013, 27, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Griffeth, R.J.; Carretero, J.; Burks, D.J. Insulin receptor substrate 2 is required for testicular development. PLoS ONE 2013, 8, e62103. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schoeller, E.L.; Albanna, G.; Frolova, A.I.; Moley, K.H. Insulin rescues impaired spermatogenesis via the hypothalamic–pituitary–gonadal axis in Akita diabetic mice and restores male fertility. Diabetes 2012, 61, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dong, J.; Yan, L.; Yong, J.; Liu, X.; Hu, Y.; Fan, X.; Wu, X.; Guo, H.; Wang, X.; et al. Single-Cell RNA-Seq Analysis Maps Development of Human Germline Cells and Gonadal Niche Interactions. Cell Stem Cell 2017, 20, 858–873.e4. [Google Scholar] [CrossRef] [PubMed]
- Darde, T.A.; Sallou, O.; Becker, E.; Evrard, B.; Monjeaud, C.; Le Bras, Y.; Jégou, B.; Collin, O.; Rolland, A.D.; Chalmel, F. The ReproGenomics Viewer: An integrative cross-species toolbox for the reproductive science community. Nucleic Acids Res. 2015, 43, W109–W116. [Google Scholar] [CrossRef]
- Darde, T.A.; Lecluze, E.; Lardenois, A.; Stévant, I.; Alary, N.; Tüttelmann, F.; Collin, O.; Nef, S.; Jégou, B.; Rolland, A.D.; et al. The ReproGenomics Viewer: A multi-omics and cross-species resource compatible with single-cell studies for the reproductive science community. Bioinformatics 2019. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.W.; Gang, G.-T.; Kim, Y.D.; Ahn, R.-S.; Harris, R.A.; Lee, C.-H.; Choi, H.-S. Insulin directly regulates steroidogenesis via induction of the orphan nuclear receptor DAX-1 in testicular Leydig cells. J. Biol. Chem. 2013, 288, 15937–15946. [Google Scholar] [CrossRef]
- Baker, J.; Hardy, M.P.; Zhou, J.; Bondy, C.; Lupu, F.; Bellvé, A.R.; Efstratiadis, A. Effects of an Igf1 gene null mutation on mouse reproduction. Mol. Endocrinol. 1996, 10, 903–918. [Google Scholar]
- Wang, G.-M.; O’Shaughnessy, P.J.; Chubb, C.; Robaire, B.; Hardy, M.P. Effects of Insulin-Like Growth Factor I on Steroidogenic Enzyme Expression Levels in Mouse Leydig Cells. Endocrinology 2003, 144, 5058–5064. [Google Scholar] [CrossRef] [PubMed][Green Version]
- DeChiara, T.M.; Efstratiadis, A.; Robertson, E.J. A growth-deficiency phenotype in heterozygous mice carrying an insulin-like growth factor II gene disrupted by targeting. Nature 1990, 345, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Nef, S.; Verma-Kurvari, S.; Merenmies, J.; Vassalli, J.-D.; Efstratiadis, A.; Accili, D.; Parada, L.F. Testis determination requires insulin receptor family function in mice. Nature 2003, 426, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Pitetti, J.-L.; Calvel, P.; Romero, Y.; Conne, B.; Truong, V.; Papaioannou, M.D.; Schaad, O.; Docquier, M.; Herrera, P.L.; Wilhelm, D.; et al. Insulin and IGF1 receptors are essential for XX and XY gonadal differentiation and adrenal development in mice. PLoS Genet. 2013, 9, e1003160. [Google Scholar] [CrossRef] [PubMed]
- Neirijnck, Y.; Calvel, P.; Kilcoyne, K.R.; Kühne, F.; Stévant, I.; Griffeth, R.J.; Pitetti, J.-L.; Andric, S.A.; Hu, M.-C.; Pralong, F.; et al. Insulin and IGF1 receptors are essential for the development and steroidogenic function of adult Leydig cells. FASEB J. 2018, 32, 3321–3335. [Google Scholar] [CrossRef]
- Shirneshan, K.; Binder, S.; Böhm, D.; Wolf, S.; Sancken, U.; Meinhardt, A.; Schmid, M.; Engel, W.; Adham, I.M. Directed overexpression of insulin in Leydig cells causes a progressive loss of germ cells. Mol. Cell. Endocrinol. 2008, 295, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Woods, K.A.; Camacho-Hübner, C.; Savage, M.O.; Clark, A.J.L. Intrauterine Growth Retardation and Postnatal Growth Failure Associated with Deletion of the Insulin-Like Growth Factor I Gene. N. Engl. J. Med. 1996, 335, 1363–1367. [Google Scholar] [CrossRef]
- Bonapace, G.; Concolino, D. A novel mutation in a patient with insulin-like growth factor 1 (IGF1) deficiency. J. Medical Genetics 2003, 40, 913–917. [Google Scholar] [CrossRef]
- Walenkamp, M.J.E.; Karperien, M.; Pereira, A.M.; Hilhorst-Hofstee, Y.; Van Doorn, J.; Chen, J.W.; Mohan, S.; Denley, A.; Forbes, B.; Van Duyvenvoorde, H.A.; et al. Homozygous and Heterozygous Expression of a Novel Insulin-Like Growth Factor-I Mutation. J. Clin. Endocrinol. Metab. 2005, 90, 2855–2864. [Google Scholar] [CrossRef]
- Netchine, I.; Azzi, S.; Houang, M.; Seurin, D.; Perin, L.; Ricort, J.-M.; Daubas, C.; Legay, C.; Mester, J.; Herich, R.; et al. Partial Primary Deficiency of Insulin-Like Growth Factor (IGF)-I Activity Associated with IGF1 Mutation Demonstrates Its Critical Role in Growth and Brain Development. J. Clin. Endocrinol. Metab. 2009, 94, 3913–3921. [Google Scholar] [CrossRef]
- Begemann, M.; Zirn, B.; Santen, G.; Wirthgen, E.; Soellner, L.; Büttel, H.-M.; Schweizer, R.; Van Workum, W.; Binder, G.; Eggermann, T. Paternally Inherited IGF2 Mutation and Growth Restriction. N. Engl. J. Med. 2015, 373, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Cannarella, R.; Condorelli, R.A.; La Vignera, S.; Calogero, A.E. Effects of the insulin-like growth factor system on testicular differentiation and function: A review of the literature. Andrology 2018, 6, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Constância, M.; Hemberger, M.; Hughes, J.; Dean, W.; Ferguson-Smith, A.; Fundele, R.; Stewart, F.; Kelsey, G.; Fowden, A.; Sibley, C.; et al. Placental-specific IGF-II is a major modulator of placental and fetal growth. Nature 2002, 417, 945–948. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, R.; Cohen, P. The role of the insulin-like growth factor system in prenatal growth. Mol. Genet. Metab. 2005, 86, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Chernausek, S.D. Update: Consequences of Abnormal Fetal Growth. J. Clin. Endocrinol. Metab. 2012, 97, 689–695. [Google Scholar] [CrossRef]
- Aquila, S.; Gentile, M.; Middea, E.; Catalano, S.; Andò, S. Autocrine Regulation of Insulin Secretion in Human Ejaculated Spermatozoa. Endocrinology 2005, 146, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Carpino, A.; Rago, V.; Guido, C.; Casaburi, I.; Aquila, S. Insulin and IR-beta in pig spermatozoa: A role of the hormone in the acquisition of fertilizing ability. Int. J. Androl. 2010, 33, 554–562. [Google Scholar] [CrossRef]
- Dissen, G.A.; Garcia-Rudaz, C.; Tapia, V.; Parada, L.F.; Hsu, S.-Y.T.; Ojeda, S.R. Expression of the Insulin Receptor-Related Receptor Is Induced by the Preovulatory Surge of Luteinizing Hormone in Thecal-Interstitial Cells of the Rat Ovary. Endocrinology 2006, 147, 155–165. [Google Scholar] [CrossRef]
- Zhou, P.; Baumgarten, S.C.; Wu, Y.; Bennett, J.; Winston, N.; Hirshfeld-Cytron, J.; Stocco, C. IGF-I signaling is essential for FSH stimulation of AKT and steroidogenic genes in granulosa cells. Mol. Endocrinol. 2013, 27, 511–523. [Google Scholar] [CrossRef]
- Khan, S.A.; Ndjountche, L.; Pratchard, L.; Spicer, L.J.; Davis, J.S. Follicle-Stimulating Hormone Amplifies Insulin-Like Growth Factor I-Mediated Activation of AKT/Protein Kinase B Signaling in Immature Rat Sertoli Cells. Endocrinology 2002, 143, 2259–2267. [Google Scholar] [CrossRef]
- Rappaport, M.S.; Smith, E.P. Insulin-Like Growth Factor I Inhibits Aromatization Induced by Follicle-Stimulating Hormone in Rat Sertoli Cell Culture1. Biol. Reprod. 1996, 54, 446–452. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Abel, M.H.; Baker, P.J.; Charlton, H.M.; Monteiro, A.; Verhoeven, G.; De Gendt, K.; Guillou, F.; O’Shaughnessy, P.J. Spermatogenesis and Sertoli Cell Activity in Mice Lacking Sertoli Cell Receptors for Follicle-Stimulating Hormone and Androgen. Endocrinology 2008, 149, 3279–3285. [Google Scholar] [CrossRef] [PubMed]
- Baumgarten, S.C.; Convissar, S.M.; Fierro, M.A.; Winston, N.J.; Scoccia, B.; Stocco, C. IGF1R signaling is necessary for FSH-induced activation of AKT and differentiation of human Cumulus granulosa cells. J. Clin. Endocrinol. Metab. 2014, 99, 2995–3004. [Google Scholar] [CrossRef] [PubMed]
- Baumgarten, S.C.; Convissar, S.M.; Zamah, A.M.; Fierro, M.A.; Winston, N.J.; Scoccia, B.; Stocco, C. FSH Regulates IGF-2 Expression in Human Granulosa Cells in an AKT-Dependent Manner. J. Clin. Endocrinol. Metab. 2015, 100, E1046–E1055. [Google Scholar] [CrossRef] [PubMed]
- Baumgarten, S.C.; Armouti, M.; Ko, C.; Stocco, C. IGF1R Expression in Ovarian Granulosa Cells Is Essential for Steroidogenesis, Follicle Survival, and Fertility in Female Mice. Endocrinology 2017, 158, 2309–2318. [Google Scholar] [CrossRef] [PubMed]
- O’Shaughnessy, P.J.; Morris, I.D.; Huhtaniemi, I.; Baker, P.J.; Abel, M.H. Role of androgen and gonadotrophins in the development and function of the Sertoli cells and Leydig cells: Data from mutant and genetically modified mice. Mol. Cell. Endocrinol. 2009, 306, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Hardy, M.P. Development of Leydig Cells in the Insulin-Like Growth Factor-I (IGF-I) Knockout Mouse: Effects of IGF-I Replacement and Gonadotropic Stimulation1. Biol. Reprod. 2004, 70, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.-X.; Lin, H.; Chen, G.-R.; Chen, B.-B.; Lian, Q.-Q.; Hardy, D.O.; Zirkin, B.R.; Ge, R.-S. Deletion of the Igf1 gene: Suppressive effects on adult Leydig cell development. J. Androl. 2010, 31, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.R.; Chandrala, S.P.; King, S.R.; Jo, Y.; Counis, R.; Huhtaniemi, I.T.; Stocco, D.M. Molecular mechanisms of insulin-like growth factor-I mediated regulation of the steroidogenic acute regulatory protein in mouse leydig cells. Mol. Endocrinol. 2006, 20, 362–378. [Google Scholar] [CrossRef]
- Buaas, F.W.; Gardiner, J.R.; Clayton, S.; Val, P.; Swain, A. In vivo evidence for the crucial role of SF1 in steroid-producing cells of the testis, ovary and adrenal gland. Development 2012, 139, 4561–4570. [Google Scholar] [CrossRef]
- Luo, X.; Ikeda, Y.; Parker, K.L. A cell-specific nuclear receptor is essential for adrenal and gonadal development and sexual differentiation. Cell 1994, 77, 481–490. [Google Scholar] [CrossRef]
- Sirianni, R.; Chimento, A.; Malivindi, R.; Mazzitelli, I.; Andò, S.; Pezzi, V. Insulin-like growth factor-I, regulating aromatase expression through steroidogenic factor 1, supports estrogen-dependent tumor Leydig cell proliferation. Cancer Res. 2007, 67, 8368–8377. [Google Scholar] [CrossRef] [PubMed]
- Fujieda, K.; Tajima, T. Molecular Basis of Adrenal Insufficiency. Pediatr. Res. 2005, 57, 62R–69R. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sakaguchi, M.; Dominko, T.; Yamauchi, N.; Leibfried-Rutledge, M.L.; Nagai, T.; First, N.L. Possible mechanism for acceleration of meiotic progression of bovine follicular oocytes by growth factors in vitro. Reproduction 2002, 123, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Stefanello, J.R.; Barreta, M.H.; Porciuncula, P.M.; Arruda, J.N.; Oliveira, J.F.; Oliveira, M.A.; Gonçalves, P.B. Effect of angiotensin II with follicle cells and insulin-like growth factor-I or insulin on bovine oocyte maturation and embryo development. Theriogenology 2006, 66, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Augustin, R.; Pocar, P.; Wrenzycki, C.; Niemann, H.; Fischer, B. Mitogenic and anti-apoptotic activity of insulin on bovine embryos produced in vitro. Reproduction 2003, 126, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Pitetti, J.L.; Torre, D.; Conne, B.; Papaioannou, M.D.; Cederroth, C.R.; Xuan, S.; Kahn, R.; Parada, L.F.; Vassalli, J.D.; Efstratiadis, A.; et al. Insulin Receptor and IGF1R Are Not Required for Oocyte Growth, Differentiation, and Maturation in Mice. Sex. Dev. 2009, 3, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Schuller, A.G.P.; Conover, C.A.; Pintar, J.E. Insulin-like growth factor (IGF) binding protein-4 is both a positive and negative regulator of IGF activity in vivo. Mol. Endocrinol. 2008, 22, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Hoang, B.; Schuller, A.G.P.; Cominski, T.P.; Hsu, M.-S.; Wood, T.L.; Pintar, J.E. Delayed Mammary Gland Involution in Mice with Mutation of the Insulin-Like Growth Factor Binding Protein 5 Gene. Endocrinology 2007, 148, 2138–2147. [Google Scholar] [CrossRef]
- Bøtkjær, J.A.; Pors, S.E.; Petersen, T.S.; Kristensen, S.G.; Jeppesen, J.V.; Oxvig, C.; Andersen, C.Y. Transcription profile of the insulin-like growth factor signaling pathway during human ovarian follicular development. J. Assist. Reprod. Genet. 2019, 36, 889–903. [Google Scholar] [CrossRef]
- Baba, T.; Otake, H.; Sato, T.; Miyabayashi, K.; Shishido, Y.; Wang, C.-Y.; Shima, Y.; Kimura, H.; Yagi, M.; Ishihara, Y.; et al. Glycolytic genes are targets of the nuclear receptor Ad4BP/SF-1. Nat. Commun. 2014, 5, 3634. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.H.; Cheng, J. FSH and testosterone signaling in Sertoli cells. Reproduction 2005, 130, 15–28. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ins2/INS | Igf1/IGF1 | Igf2/IGF2 | Insr/INSR | Igf1r/IGF1R | Irs1/IRS1 | Irs2/IRS2 | |||
|---|---|---|---|---|---|---|---|---|---|
| Mouse XY | Gene expression 1 | Interstitial progenitors | - | +++ | +++ | - | +++ | - | + |
| Leydig cells | - | + | + | + | +++ | - | + | ||
| Sertoli cells | - | - | - | - | +++ | - | +++ | ||
| Germ cells | - | - | + | - | +++ | - | + | ||
| Endothelial cells | - | + | +++ | - | +++ | - | - | ||
| Phenotypes | - Insulin-injected and HFD mouse models: reduced steroidogenic enzyme gene expression and steroidogenesis [49] | - Constitutive KO: steroidogenic failure, PLC markers upregulation, reduced testis size and sperm count [50,51] | - Constitutive KO: no testicular defects [52] | - Constitutive double KO: reduced proliferation rates of somatic progenitors, male-to-female sex reversal [53,54] | - Constitutive KO: no testicular defects [44] | - Constitutive KO: reduced testis size and sperm count [44] | |||
| - LC specific-KO: adult Leydig cells (ALCs) maturation defects, cell-autonomous steroidogenic failure, non-cell autonomous PLCs (progenitor Leydig cells) enrichment, reduced testis size and sperm count. Fetal LCs (FLCs) not affected [55] | |||||||||
| - LC specific-KI: age-dependent germ cell degeneration [56] | - Sertoli (SC) specific-KO: reduced follicle stimulating hormone (FSH)-dependant SC proliferation, testis size and sperm count [43] acting through IGF/PTEN/PI3K pathway [42] | ||||||||
| - GC specific-KO: no testicular defects [43] | |||||||||
| Human XY | Gene expression 1 | Interstitial progenitors | - | +++ | + | +++ | +++ | - | - |
| Leydig cells | - | - | - | +++ | +++ | +++ | +++ | ||
| Sertoli cells | - | - | - | +++ | +++ | - | - | ||
| Germ cells | - | + | - | + | +++ | - | - | ||
| Endothelial cells | - | - | - | +++ | +++ | - | - | ||
| Conditions | - Homozygous mutation: reproductive system not affected [57,58,59,60] | - Paternally inherited IGF2 mutation: ambiguous genitalia, penoscrotal hypospadia, unilateral cryptorchidism, hypogonadism [61] | |||||||
| - Positive correlation between IGF1 and testicular volume [62] | |||||||||
| - IGF1 administration to Laron syndrome patients: increased testis size [62] | |||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neirijnck, Y.; Papaioannou, M.D.; Nef, S. The Insulin/IGF System in Mammalian Sexual Development and Reproduction. Int. J. Mol. Sci. 2019, 20, 4440. https://doi.org/10.3390/ijms20184440
Neirijnck Y, Papaioannou MD, Nef S. The Insulin/IGF System in Mammalian Sexual Development and Reproduction. International Journal of Molecular Sciences. 2019; 20(18):4440. https://doi.org/10.3390/ijms20184440
Chicago/Turabian StyleNeirijnck, Yasmine, Marilena D. Papaioannou, and Serge Nef. 2019. "The Insulin/IGF System in Mammalian Sexual Development and Reproduction" International Journal of Molecular Sciences 20, no. 18: 4440. https://doi.org/10.3390/ijms20184440
APA StyleNeirijnck, Y., Papaioannou, M. D., & Nef, S. (2019). The Insulin/IGF System in Mammalian Sexual Development and Reproduction. International Journal of Molecular Sciences, 20(18), 4440. https://doi.org/10.3390/ijms20184440