1. Introduction
Perennial woody plants are present across the world and cover about 30% of the land surface [
1]. Trees are a source of numerous economical products, such as timber, fuel, food, resins, and oil, but they also provide a range of ecosystem services including carbon storage, maintenance of wildlife habitats, and soil stabilization [
2]. Unlike herbaceous plants, they usually have a longer life cycle, spanning from decades to centuries. Therefore, they have to withstand numerous challenges throughout their life span, under the form of both biotic and abiotic stress constraints. Abiotic stresses are the main factors affecting plant distribution and primary production [
3]. From this perspective, unravelling stress acclimation and resistance mechanisms could help us to optimize plant primary production [
4].
Basket willow (
Salix viminalis L.) is a fast-growing tree species with a wide Eurasian distribution that can be found in cool, temperate, and boreal climates. It is a pioneer species of riparian environments with a moderate to high metal tolerance [
5,
6]. It is one of the most important species used in basketry and its fast growth makes it amongst the most used species in intensive short-rotation agro-forestry systems in northern temperate regions [
7]. In addition to this, its highly developed root system and important evapotranspiration rate make
S. viminalis highly suitable for phytoremediation, on-site metal phytoextraction and the treatment of wastewater, sewage sludge and landfill leachate [
8,
9]. However, while
S. viminalis is well studied at a physiological level, it is poorly characterized at a molecular level, making breeding programs and clone selection time consuming. For example, field trials to assess
Salix performance in soils contaminated with metals can take up to five years [
10]. In addition, most of the studies focus on the aboveground organs, which represent the most valued parts but not the primary site of exposure to metals. Roots, which are directly exposed to soil and hence high concentrations of metals, act as a metal filter and in most species the bulk of metal ions are sequestered in the roots, limiting their progress into the aboveground organs via the xylem [
11]. In addition, genes influencing the root system architecture are poorly known. Nevertheless, the root system architecture plays a primordial role in plant fitness as it determines the capacity of a plant to access mineral ions and water [
12,
13]. Information about the expression patterns of genes involved in root architecture and stress tolerance at the root level would hasten clone selection and pave the way for the development of trees with improved capacity of metal sequestration [
14].
Gene expression analysis has been applied to unravel biological mechanisms in many organisms. Amongst the techniques available to analyze gene expression, real-time or quantitative polymerase chain reaction (qPCR) has been an inestimable source of data, thanks to its accuracy, specificity, high sensitivity, repeatability, and reproducibility. As such, Minimum Information for the publication of real-time Quantitative PCR Experiments (MIQE) was written in an effort to standardize qPCR practice and reduce the publication of incorrect qPCR data [
15]. These guidelines include considerations on nomenclature, concepts, sample preparation, and normalization. The normalization step involves the comparison of mRNA ratios of genes of interest (GOIs) to those of reference genes (RGs). Reference genes are genes with a stable expression level across the studied conditions and are typically genes involved in basic cellular processes [
16]. For proper gene expression normalization, more than one RG should be used and several algorithms exist to select the most suitable ones [
17]. For the moment, no “universal reference gene” is known, necessitating the description and validation of RGs depending on the species, organs, and conditions to which plants are exposed in a study. However, stability validation of RGs is not systematic in plant science, resulting in poor data normalization, which can lead to inadequate conclusions [
18,
19,
20].
To our knowledge, no systematic study aiming to validate potential RGs has been conducted for S. viminalis. Given the potential of S. viminalis in phytoremediation and energy production, it is essential to identify stably expressed RGs in different organs and across several conditions to foster its selection in future applications. In this work, we studied the expression pattern of 14 candidate reference genes in the roots and leaves of S. viminalis exposed to different conditions. In addition, we investigated the expression stability of these candidate reference genes in roots stele, roots cortex, stem xylem, and bark under control conditions. Using different algorithms and combinations thereof, we subsequently determined the most stable reference genes, produced a consensus list and the optimal combination of RGs, with an emphasis on those that could be used for studies at the root level.
3. Discussion
Currently, qPCR is widely used to perform gene expression analysis [
15]. However, the use of non-stable RGs for data normalization can lead to improper analysis and conclusions [
20]. Therefore, the aim of this study was to identify RGs in
S. viminalis with a stable expression level across various conditions and organs. While validation of appropriate RGs has been done in many plants, most studies focus on herbaceous plants [
30] and no RGs are publicly available for the closely related
S. purpurea, which has been sequenced.
Due to the high potential of S. viminalis in phytoremediation, together with its high primary productivity, we performed RG validation for leaves and roots exposed to various conditions. The study showed that the potential RGs had different expression profiles depending on the organ studied, which highlights the need to perform condition- and organ-specific RG validation before every gene expression experiment.
Chlorophyll fluorescence was measured and the F
V/F
M ratio was calculated and used as a stress marker. Interestingly, the F
V/F
M ratio of the drought-exposed plants only slightly decreased during the six first days but on the seventh, two of the three plants had lost their leaves. Conversely, the plants exposed to cold showed a drop in F
V/F
M value the first day and subsequently recovered, although the stress was maintained at the same level. According to Lichtenthaler (1998) [
35], stress has a dose-effect relationship i.e., while some stressors have a low effect on plant survival at low “concentration”, higher “concentration” or longer exposure time could lead to significant damage to the plant integrity (exhaustion phase). Conversely, some stresses can induce a temporary distress (alarm phase) which act as a stimulatory signal for the plant and induces its hardening. The F
V/F
M ratio could indicate the alarm and exhaustion phases in the stress response rather than the application of a stressor to the plant.
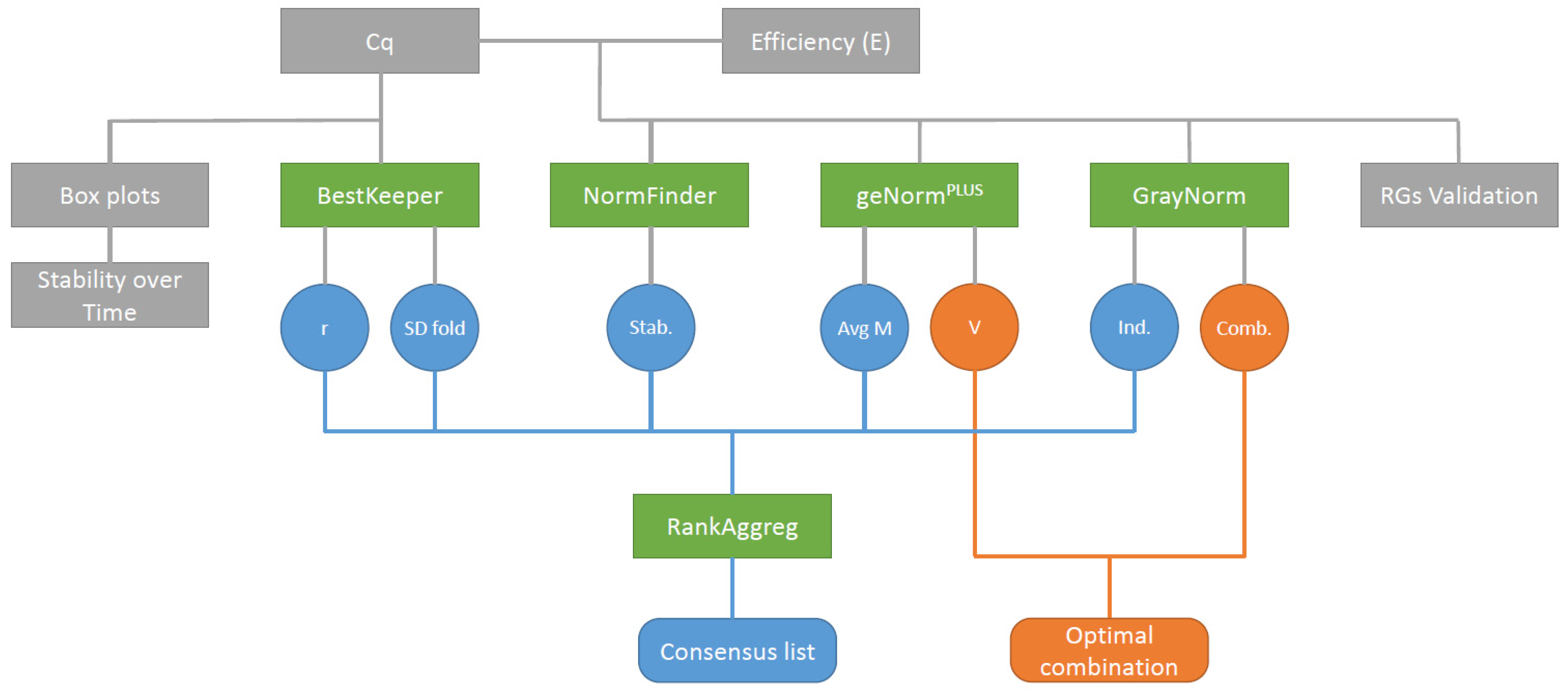
Four different algorithms, geNormPLUS, BestKeeper, NormFinder, and GrayNorm were used to analyze the expression stability of 14 candidate RGs. While the four algorithms produced globally similar rankings, some punctual but important differences could be observed between the algorithms’ outputs. For example, while geNormPLUS, BestKeeper and NormFinder ranked GAPDH amongst the least stable genes across all conditions, GrayNorm ranked this gene among the most stable ones when all conditions were grouped and under heat stress. These kinds of discrepancies arise from the fact that each algorithm is based on specific premises and working hypotheses. However, under most conditions, the same general pattern could be observed across the different algorithms. More specifically, CDC2 and TIP41 always ranked amongst the most stable genes across all conditions; CYP displayed a high expression stability under control and cold conditions but a lower expression stability under the other conditions; ACT, ARI8, and EF1b were globally stable; and GAPDH, β-TUB, and PT1 were the least stable genes. Nevertheless, the variations existing between the outputs of the different algorithms were smoothened by the use of RankAggreg, which allowed us to produce a consensus list.
In the consensus list,
CDC2 and
TIP41 were the most stable RGs across all conditions.
CDC2 codes for a cyclin dependent kinase (CDK1/CDKA), which is involved in eukaryotic cell cycle regulation. In association with various cyclins, CDK1/CDKA is known for its role in the control of the transition from one cell cycle phase to another [
36]. It is interesting that its expression remained stable during metal stress, as cadmium has been reported to block the cell cycle at the G2 checkpoint in the roots of
Lactuca sativa [
37].
TIP41 codes for a 41kDa protein interacting with TAP42, which is involved in the regulation of cellular growth in response to the nutrient status and the environment [
38]. It has been shown to be induced by long-term exposure to NaCl [
38], but this was not observed in our study.
CYP codes for a cyclophilin peptidyl prolyl isomerase involved in a wide range of basal functions, such as proper protein folding and regulation of plant growth and development. Some cyclophilin isoforms have been observed to be induced by abiotic stresses [
39], but this was not the case for the cyclophilin isoform used in the present study.
UCEE2 and
ARI8 both code for proteins involved in the ubiquitination process (respectively in ubiquitin conjugation and ubiquitin ligation) [
40]. Proteins involved in ubiquitination are constitutively expressed and have been reported to be reliable RGs in various species and under diverse stresses [
16]. Interestingly, while
ARI8 was relatively stable,
UCEE2 expression stability varied from relatively stable (sixth most stable gene in all conditions grouped) to unstable (thirteenth under control condition).
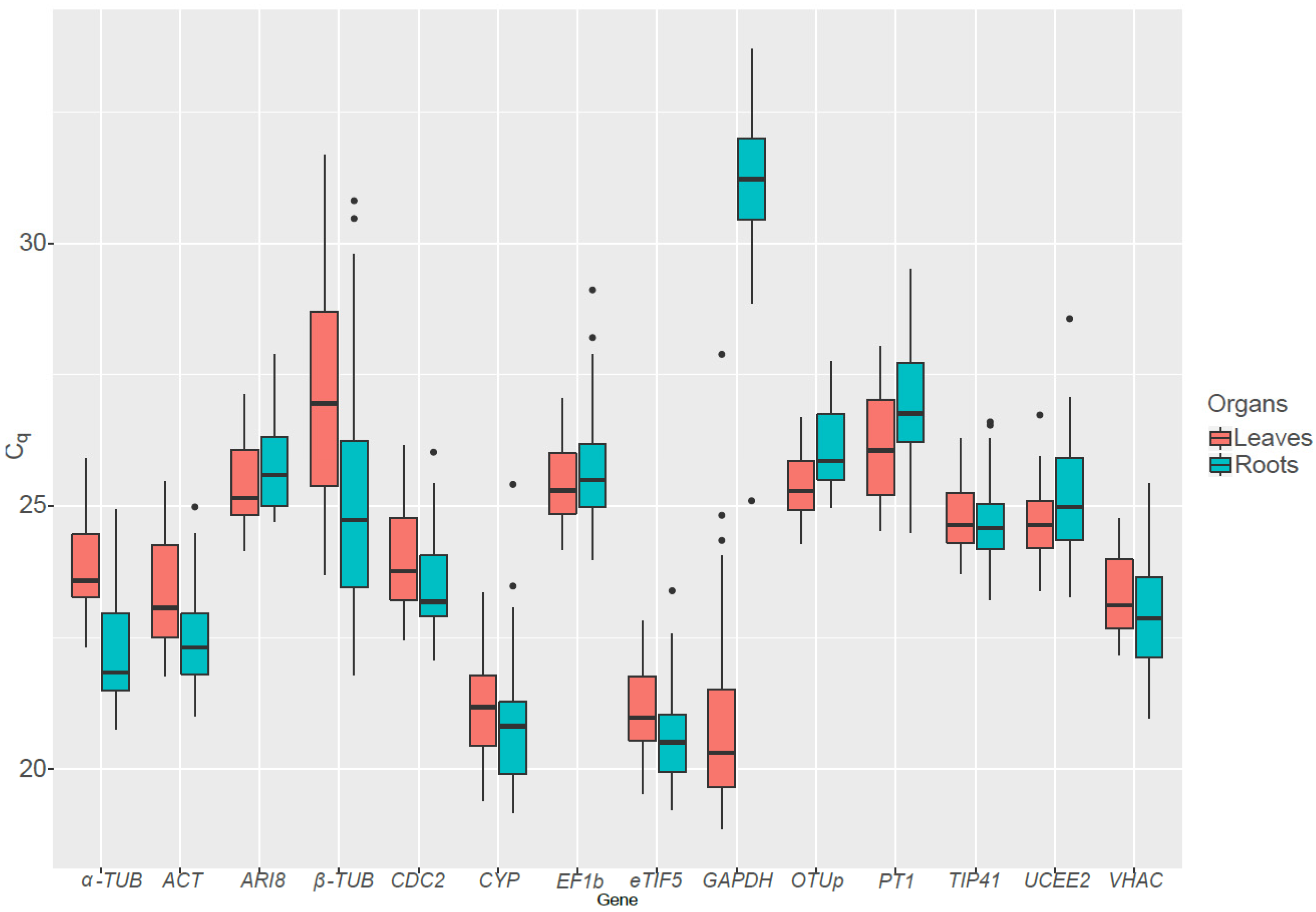
GAPDH, which was amongst the least stable genes, is involved in glycolysis but also in non-metabolic functions during abiotic stress [
41]. It was less expressed in the roots than in the leaves, as it also plays a role in the Calvin-Benson cycle.
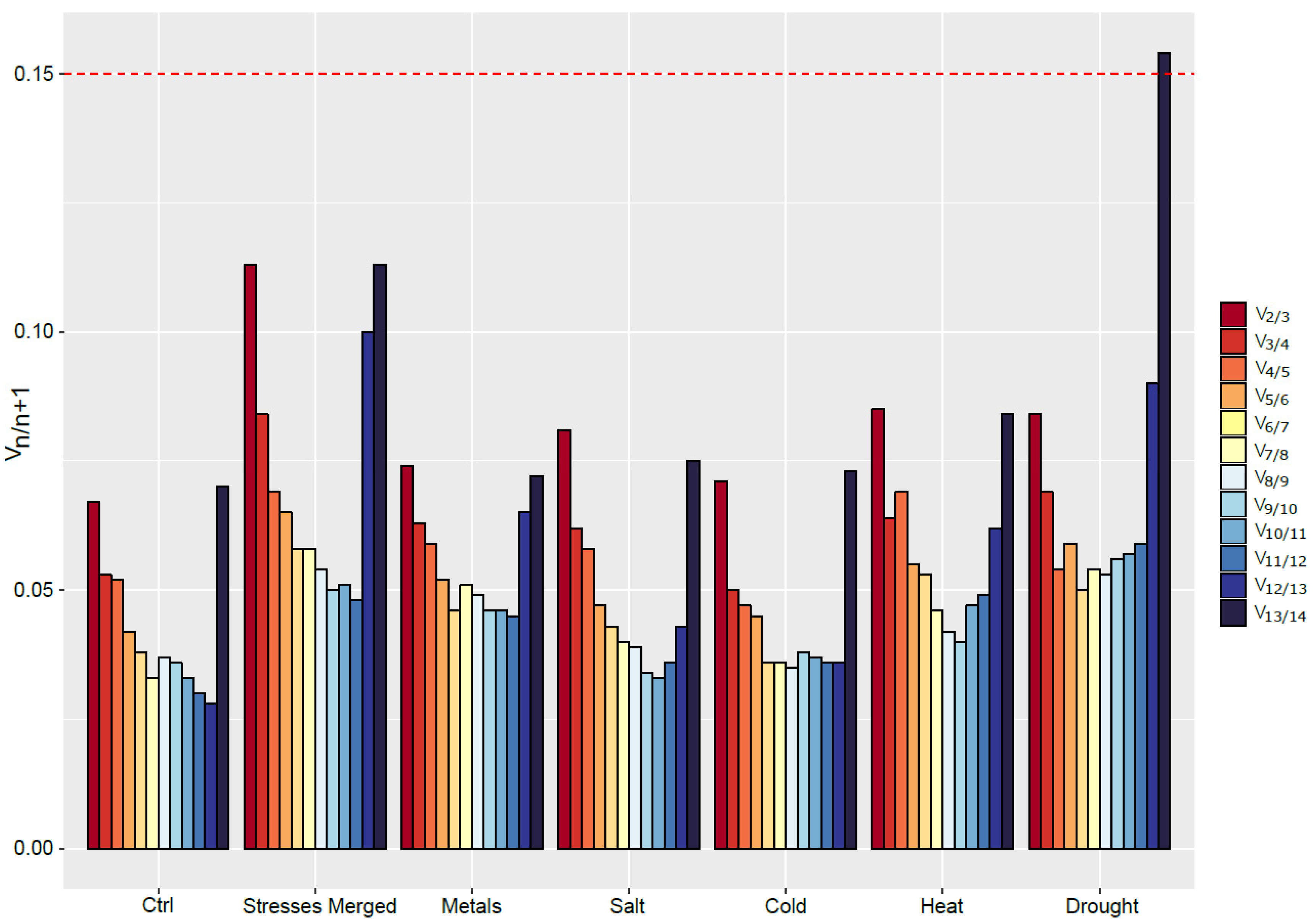
The expression level of the different candidate RGs was globally stable over time in the roots of S. viminalis under control condition. The same pattern could be observed for every RGs except GAPDH in the roots of plants exposed to cold and heat. No clear trend could be observed in the roots exposed to salt stress as some genes showed increasing expression level and other decreasing. Under drought conditions, a significant increase in the Cqs value could be observed for most genes, except for GAPDH, which displays opposite trend. Curiously, candidate RGs that showed different trends than the other RGs (for example CYP, β-TUB and GAPDH in the roots under heat stress) were ranked as less stable by the algorithms. This is due to the fact that ideal RGs expression patterns are expected to be highly correlated between them.
Interestingly,
GAPDH and
β-TUB, which are both widely used as reference genes in plants, were ranked as the least stable genes under most conditions. This is worrisome, as standardization with non-stable RGs can lead to misinterpretations of data, all the more since often only one RG is used. For example, let us consider an experiment in which we want to investigate the effect of metal exposure on the expression level of some GOI in the roots of
S. viminalis. If a GOI is standardized with
GAPDH, which has a maximum variation of 4.04 C
q between control roots and roots exposed to metals, there could potentially be a [(amplification factor)
maximum variation = 1.88
4.04 =] 12.81-fold difference in GOI quantification compared to the situation where we use an ideal RG, which would have no variation across samples and conditions. However, as mentioned previously, perfect RGs do not exist, and even
TIP41, which was ranked as the most stable gene under metal exposure, displayed a maximum variation of 2.24-fold. As such, the use of multiple RGs is recommended, as it cancels out the variation in expression observed for individual RG [
17,
26].
The optimum combination of genes given by geNorm
PLUS and GrayNorm were not the same. This was expected, as the two algorithms have a different approach to determine the best combination of RGs. Indeed, geNorm
PLUS combines genes based on their individual stability i.e., it groups together the
n most stable genes, as such the addition of a new RG (
n+1) does not significantly affect the normalization factor. On the contrary, GrayNorm groups genes based on the stability of their NF. This can lead to the proposed use of genes with poor individual stability but which, grouped together, have a higher stability. In other words, geNorm
PLUS computes the combination of the best RGs while GrayNorm calculates the best combination of RGs. Nevertheless, it is clear from both the geNorm
PLUS and GrayNorm output that the optimal number of genes to use for gene expression normalization depends on the condition and the organ that is targeted, as it was already observed in
Arabidopsis thaliana [
42],
Petunia hybrid [
43],
Populus euphratica [
30], and
Salix psammophila [
22] amongst others. Nevertheless, a minimum of three stable genes should be sufficient for data normalization in most studies [
28].
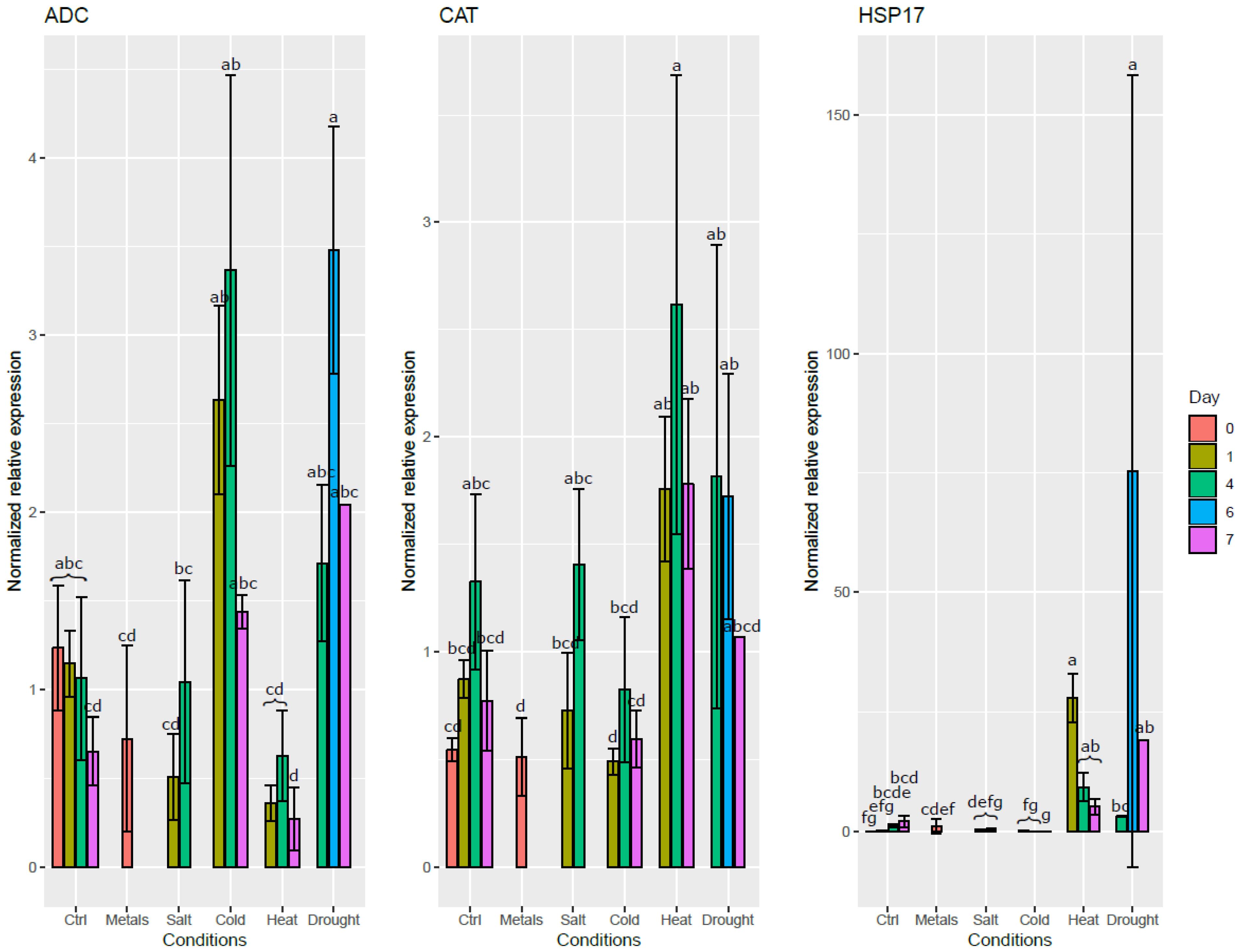
For the validation phase, we used three genes known to change in expression during exposure to abiotic stresses. In general, the stress-responsive genes displayed the same response patterns in both roots and leaves. However, some differences were observed between both organs. For example, while metals did not induce ADC in the roots, its expression level increased in the leaves. Similarly, while cold did not significantly altered CAT expression in the roots, it was significantly down-regulated in the cold-exposed leaves.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}