IL-4 Augments IL-31/IL-31 Receptor Alpha Interaction Leading to Enhanced Ccl 17 and Ccl 22 Production in Dendritic Cells: Implications for Atopic Dermatitis

 , and
, and
Abstract
1. Introduction
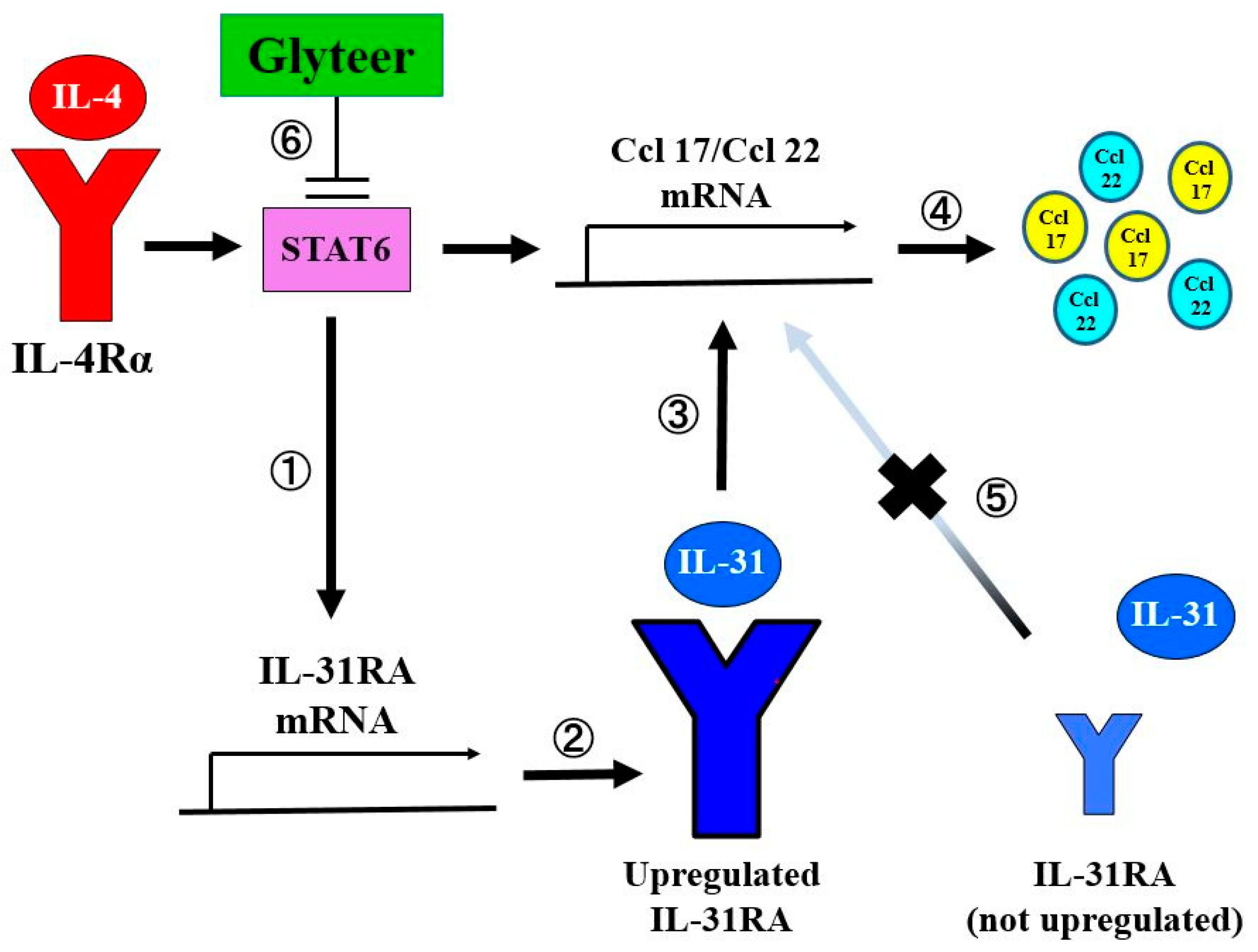
2. Results
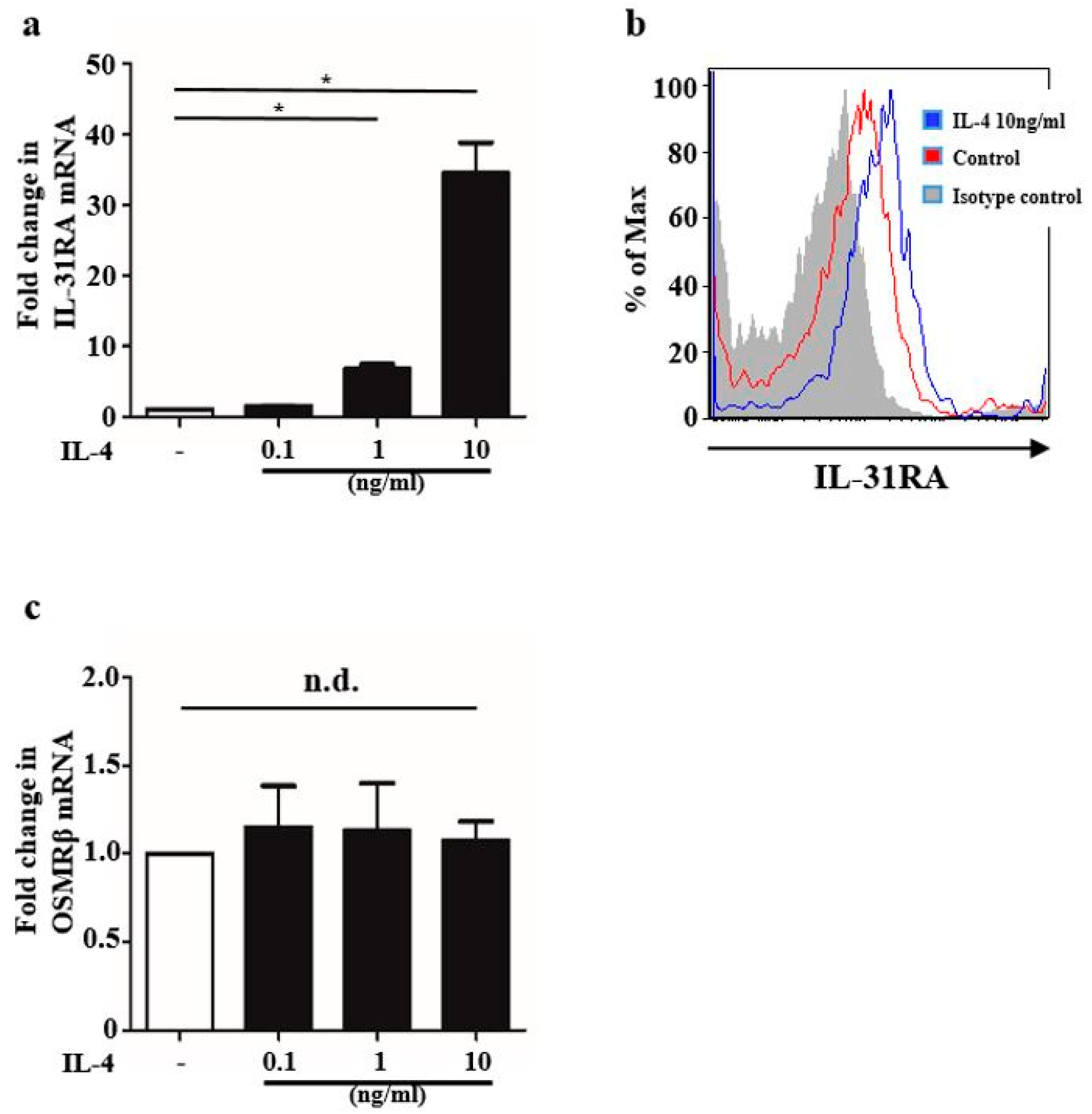
2.1. IL-4 Stimulation Upregulated IL-31RA Expression in BMDCs
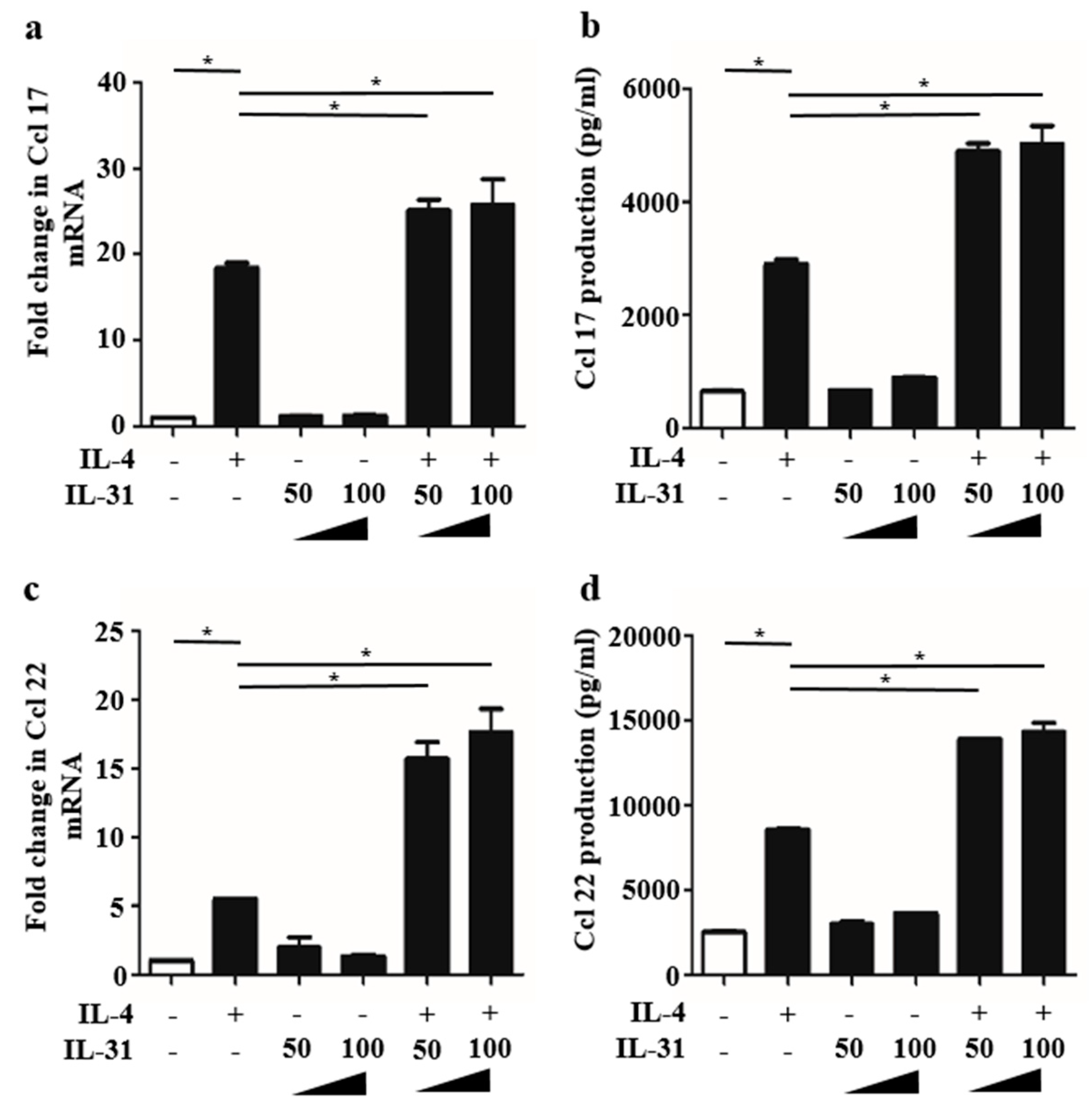
2.2. IL-31 Stimulation Enhanced IL-4-Induced Ccl 17 and Ccl 22 Production in BMDCs
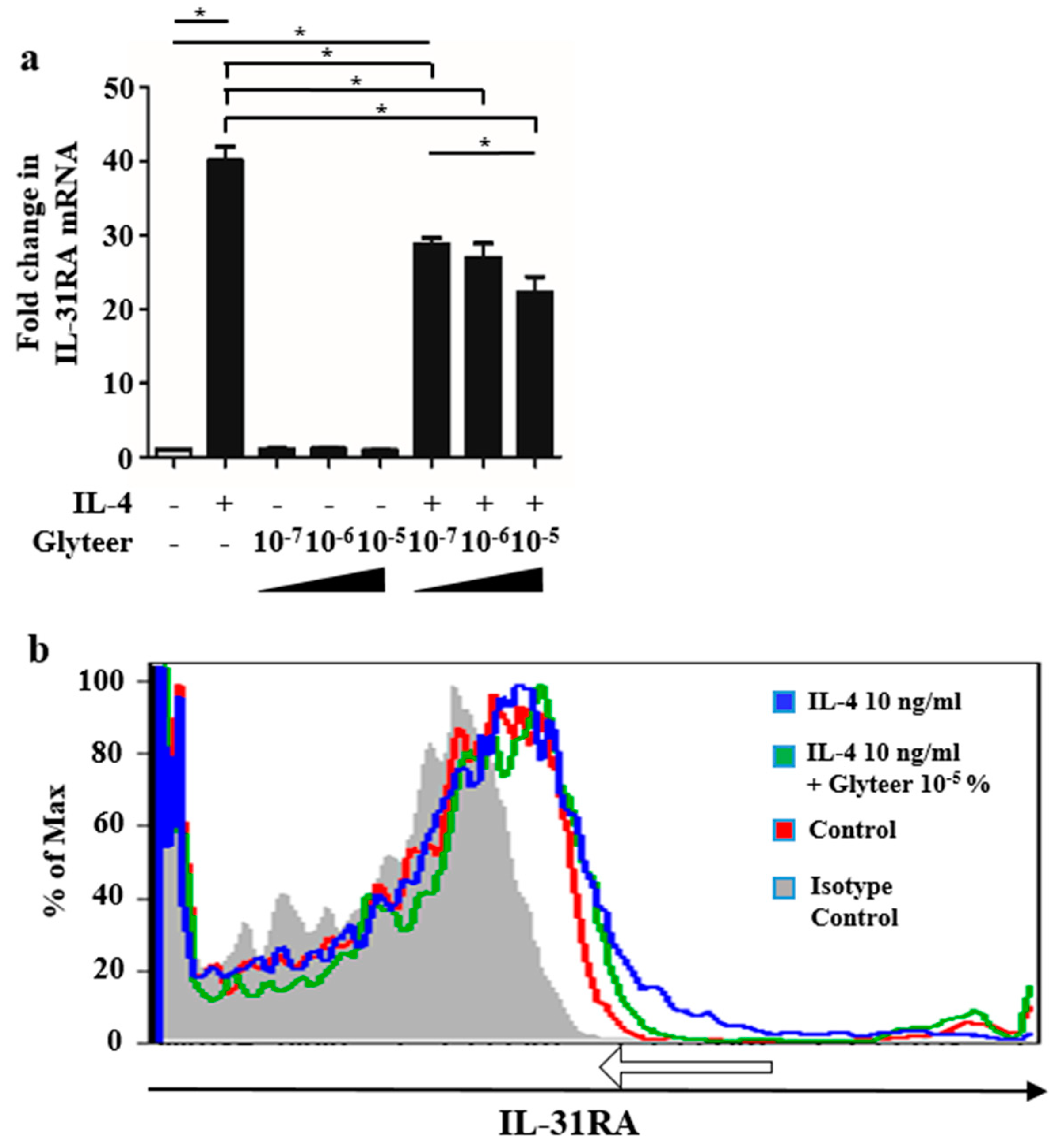
2.3. Glyteer Treatment Inhibited IL-4-Induced IL-31RA Expression in BMDCs
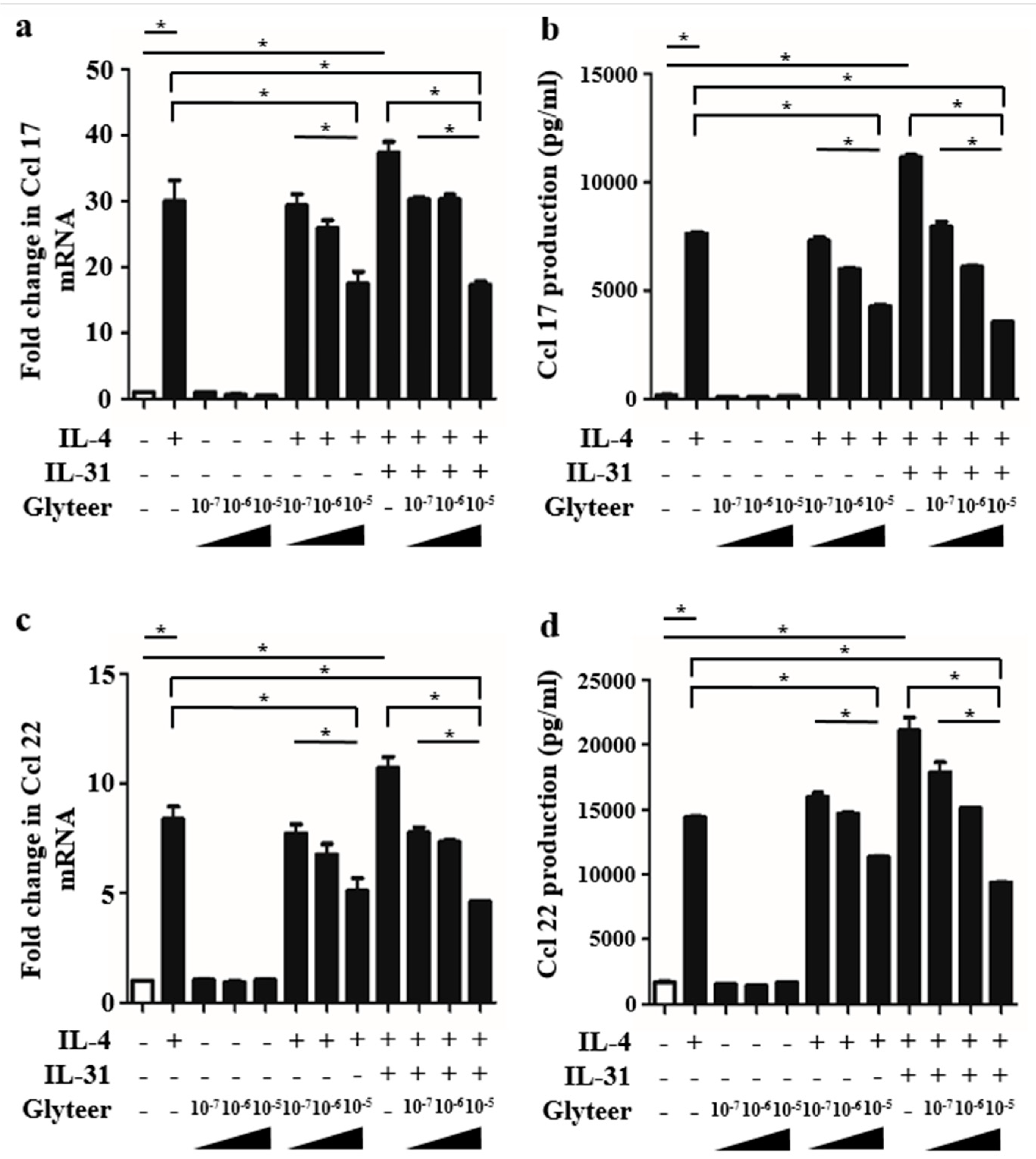
2.4. Glyteer Treatment Inhibited IL-4- and IL-31-Induced Ccl 17 and Ccl 22 Production in BMDCs
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Generation of Bone Marrow-Derived DCs (BMDCs) and Cell Culture
4.3. Quantitative Reverse-Transcription (qRT)-PCR Analysis
4.4. ELISA
4.5. Fluorescence-Activated Cell Sorting (FACS) Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dillon, S.R.; Sprecher, C.; Hammond, A.; Bilsborough, J.; Rosenfeld-Franklin, M.; Presnell, S.R.; Haugen, H.S.; Maurer, M.; Harder, B.; Johnston, J.; et al. Interleukin 31, a Cytokine Produced by Activated T Cells, Induces Dermatitis in Mice. Nat. Immunol. 2004, 5, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Putheti, P.; Zhou, Q.; Liu, Q.; Gao, W. Structures and Biological Functions of IL-31 and IL-31 Receptors. Cytokine Growth Factor Rev. 2008, 19, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Stott, B.; Lavender, P.; Lehmann, S.; Pennino, D.; Durham, S.; Schmidt-Weber, C.B. Human IL-31 is Induced by IL-4 and Promotes TH2-driven Inflammation. J. Allergy Clin. Immunol. 2013, 132, 446–454.e5. [Google Scholar] [CrossRef] [PubMed]
- Edukulla, R.; Singh, B.; Jegga, A.G.; Sontake, V.; Dillon, S.R.; Madala, S.K. Th2 Cytokines Augment IL-31/IL-31RA Interactions via STAT6-dependent IL-31RA Expression. J. Biol. Chem. 2015, 290, 13510–13520. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Chiba, T.; Tsuji, G.; Ulzii, D.; Kido-Nakahara, M.; Nakahara, T.; Kadono, T. Atopic Dermatitis: Immune Deviation, Barrier Dysfunction, IgE Autoreactivity and New Therapies. Allergol. Int. 2017, 66, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.L.; Bieber, T.; Guttman-Yassky, E.; Beck, L.A.; Blauvelt, A.; Cork, M.J.; Silverberg, J.I.; Deleuran, M.; Kataoka, Y.; Lacour, J.P.; et al. Two Phase 3 Trials of Dupilumab Versus Placebo in Atopic Dermatitis. N. Engl. J. Med. 2016, 375, 2335–2348. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, T.; Hanifin, J.M.; Furue, M.; Pulka, G.; Mlynarczyk, I.; Wollenberg, A.; Galus, R.; Etoh, T.; Mihara, R.; Yoshida, H.; et al. Anti-Interleukin-31 Receptor A Antibody for Atopic Dermatitis. N. Engl. J. Med. 2017, 376, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Cevikbas, F.; Wang, X.; Akiyama, T.; Kempkes, C.; Savinko, T.; Antal, A.; Kukova, G.; Buhl, T.; Ikoma, A.; Buddenkotte, J.; et al. A Sensory Neuron-expressed IL-31 Receptor Mediates T Helper Cell-dependent Itch: Involvement of TRPV1 and TRPA1. J. Allergy Clin. Immunol. 2014, 133, 448–460. [Google Scholar] [CrossRef]
- Kato, A.; Fujii, E.; Watanabe, T.; Takashima, Y.; Matsushita, H.; Furuhashi, T.; Morita, A. Distribution of IL-31 and its Receptor Expressing Cells in Skin of Atopic Dermatitis. J. Dermatol. Sci. 2014, 74, 229–235. [Google Scholar] [CrossRef]
- Feld, M.; Garcia, R.; Buddenkotte, J.; Katayama, S.; Lewis, K.; Muirhead, G.; Hevezi, P.; Plesser, K.; Schrumpf, H.; Krjutskov, K.; et al. The Pruritus- and TH2-associated Cytokine IL-31 Promotes Growth of Sensory Nerves. J. Allergy Clin. Immunol. 2016, 138, 500–508.e24. [Google Scholar] [CrossRef]
- Yamamura, K.; Uruno, T.; Shiraishi, A.; Tanaka, Y.; Ushijima, M.; Nakahara, T.; Watanabe, M.; Kido-Nakahara, M.; Tsuge, I.; Furue, M.; et al. The Transcription Factor EPAS1 Links DOCK8 Deficiency to Atopic Skin Inflammation via IL-31 Induction. Nat. Commun. 2017, 8, 13946. [Google Scholar] [CrossRef] [PubMed]
- Stutte, S.; Quast, T.; Gerbitzki, N.; Savinko, T.; Novak, N.; Reifenberger, J.; Homey, B.; Kolanus, W.; Alenius, H.; Förster, I. Requirement of CCL17 for CCR7- and CXCR4-dependent Migration of Cutaneous Dendritic Cells. Proc. Natl. Acad. Sci. USA 2010, 107, 8736–8741. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, Y. Thymus and Activation-regulated Chemokine as a Clinical Biomarker in Atopic Dermatitis. J. Dermatol. 2014, 41, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Nakamura, K.; Oyama, N.; Kaneko, F.; Tsunemi, Y.; Saeki, H.; Tamaki, K. Macrophage-derived Chemokine (MDC)/CCL22 Produced by Monocyte Derived Dendritic Cells Reflects the Disease Activity in Patients with Atopic Dermatitis. J. Dermatol. Sci. 2006, 44, 93–99, Epub Sep 27, 2006. [Google Scholar] [CrossRef] [PubMed]
- Takemura, M.; Nakahara, T.; Hashimoto-Hachiya, A.; Furue, M.; Tsuji, G. Glyteer, Soybean Tar, Impairs IL-4/Stat6 Signaling in Murine Bone Marrow-Derived Dendritic Cells: The Basis of Its Therapeutic Effect on Atopic Dermatitis. Int. J. Mol. Sci. 2018, 19, 1169. [Google Scholar] [CrossRef] [PubMed]
- Horejs-Hoeck, J.; Schwarz, H.; Lamprecht, S.; Maier, E.; Hainzl, S.; Schmittner, M.; Posselt, G.; Stoecklinger, A.; Hawranek, T.; Duschl, A. Dendritic Cells Activated by IFN-γ/STAT1 Express IL-31 Receptor and Release Proinflammatory Mediators upon IL-31 Treatment. J. Immunol. 2012, 188, 5319–5326. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Mitoma, C.; Hashimoto-Hachiya, A.; Uchi, H.; Takahara, M.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T.; Furue, M. Antioxidant Soybean Tar Glyteer Rescues T-helper-mediated Downregulation of Filaggrin Expression via Aryl Hydrocarbon Receptor. J. Dermatol. 2015, 42, 171–180. [Google Scholar] [CrossRef]
- Dambacher, J.; Beigel, F.; Seiderer, J.; Haller, D.; Göke, B.; Auernhammer, C.J.; Brand, S. Interleukin 31 Mediates MAP Kinase and STAT1/3 Activation in Intestinal Epithelial Cells and its Expression is Upregulated in Inflammatory Bowel Disease. Gut 2007, 56, 1257–1265. [Google Scholar] [CrossRef]
- Kasraie, S.; Niebuhr, M.; Werfel, T. Interleukin (IL)-31 Activates Signal Transducer and Activator of Transcription (STAT)-1, STAT-5, and Extracellular Signal-regulated Kinase 1/2 and Down-regulates IL-12p40 Production in Activated Human Macrophages. Allergy 2013, 68, 739–747. [Google Scholar] [CrossRef]
- Kwon, D.J.; Bae, Y.S.; Ju, S.M.; Goh, A.R.; Youn, G.S.; Choi, S.Y.; Park, J. Casuarinin Suppresses TARC/CCL17 and MDC/CCL22 Production via Blockade of NF-κB and STAT1 Activation in HaCaT Cells. Biochem. Biophys. Res. Commun. 2012, 417, 1254–1259. [Google Scholar] [CrossRef]
- Yano, C.; Saeki, H.; Komine, M.; Kagami, S.; Tsunemi, Y.; Ohtsuki, M.; Nakagawa, H. Mechanism of Macrophage-Derived Chemokine/CCL22 Production by HaCaT Keratinocytes. Ann. Dermatol. 2015, 27, 152–156. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5’ to 3’) | |
|---|---|---|
| β-actin | forward | GGCTGTATTCCCCTCCATCG |
| reverse | CCAGTTGGTAACAATGCCATGT | |
| Ccl17 | forward | AGGTCACTTCAGATGCTGCTC |
| reverse | ACTCTCGGCCTACATTGGTG | |
| Ccl22 | forward | GACACCTGACGAGGACACA |
| reverse | GCAGAGGGTGACGGATGTAG | |
| IL-31RA | forward | CGATTGTTGTGGAAGAAGGCAA |
| reverse | TACTGCTGGGTGGTGATGTTG | |
| OSMRβ | forward | CAGGCGGGTAATCAGACCAATG |
| reverse | CATGAGTAAGGGCTGGGACA | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miake, S.; Tsuji, G.; Takemura, M.; Hashimoto-Hachiya, A.; Vu, Y.H.; Furue, M.; Nakahara, T. IL-4 Augments IL-31/IL-31 Receptor Alpha Interaction Leading to Enhanced Ccl 17 and Ccl 22 Production in Dendritic Cells: Implications for Atopic Dermatitis. Int. J. Mol. Sci. 2019, 20, 4053. https://doi.org/10.3390/ijms20164053
Miake S, Tsuji G, Takemura M, Hashimoto-Hachiya A, Vu YH, Furue M, Nakahara T. IL-4 Augments IL-31/IL-31 Receptor Alpha Interaction Leading to Enhanced Ccl 17 and Ccl 22 Production in Dendritic Cells: Implications for Atopic Dermatitis. International Journal of Molecular Sciences. 2019; 20(16):4053. https://doi.org/10.3390/ijms20164053
Chicago/Turabian StyleMiake, Sho, Gaku Tsuji, Masaki Takemura, Akiko Hashimoto-Hachiya, Yen Hai Vu, Masutaka Furue, and Takeshi Nakahara. 2019. "IL-4 Augments IL-31/IL-31 Receptor Alpha Interaction Leading to Enhanced Ccl 17 and Ccl 22 Production in Dendritic Cells: Implications for Atopic Dermatitis" International Journal of Molecular Sciences 20, no. 16: 4053. https://doi.org/10.3390/ijms20164053
APA StyleMiake, S., Tsuji, G., Takemura, M., Hashimoto-Hachiya, A., Vu, Y. H., Furue, M., & Nakahara, T. (2019). IL-4 Augments IL-31/IL-31 Receptor Alpha Interaction Leading to Enhanced Ccl 17 and Ccl 22 Production in Dendritic Cells: Implications for Atopic Dermatitis. International Journal of Molecular Sciences, 20(16), 4053. https://doi.org/10.3390/ijms20164053