Simultaneous Detection of the T790M and L858R Mutations in the EGFR Gene by Oligoribonucleotide Interference-PCR


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
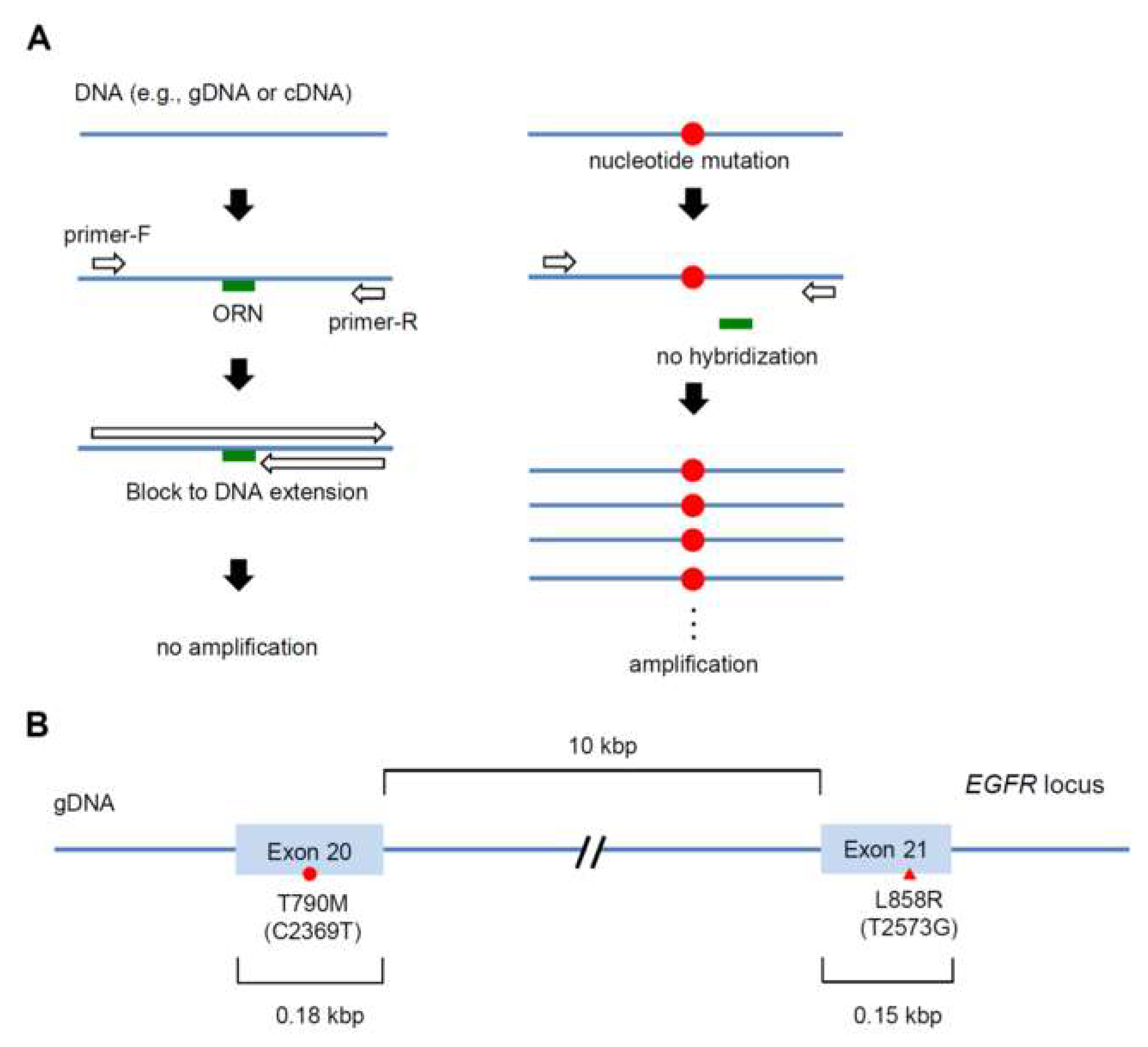
2. Results
2.1. Detection of the Single-Nucleotide Mutation L858R (T2573G) by ORNi-PCR
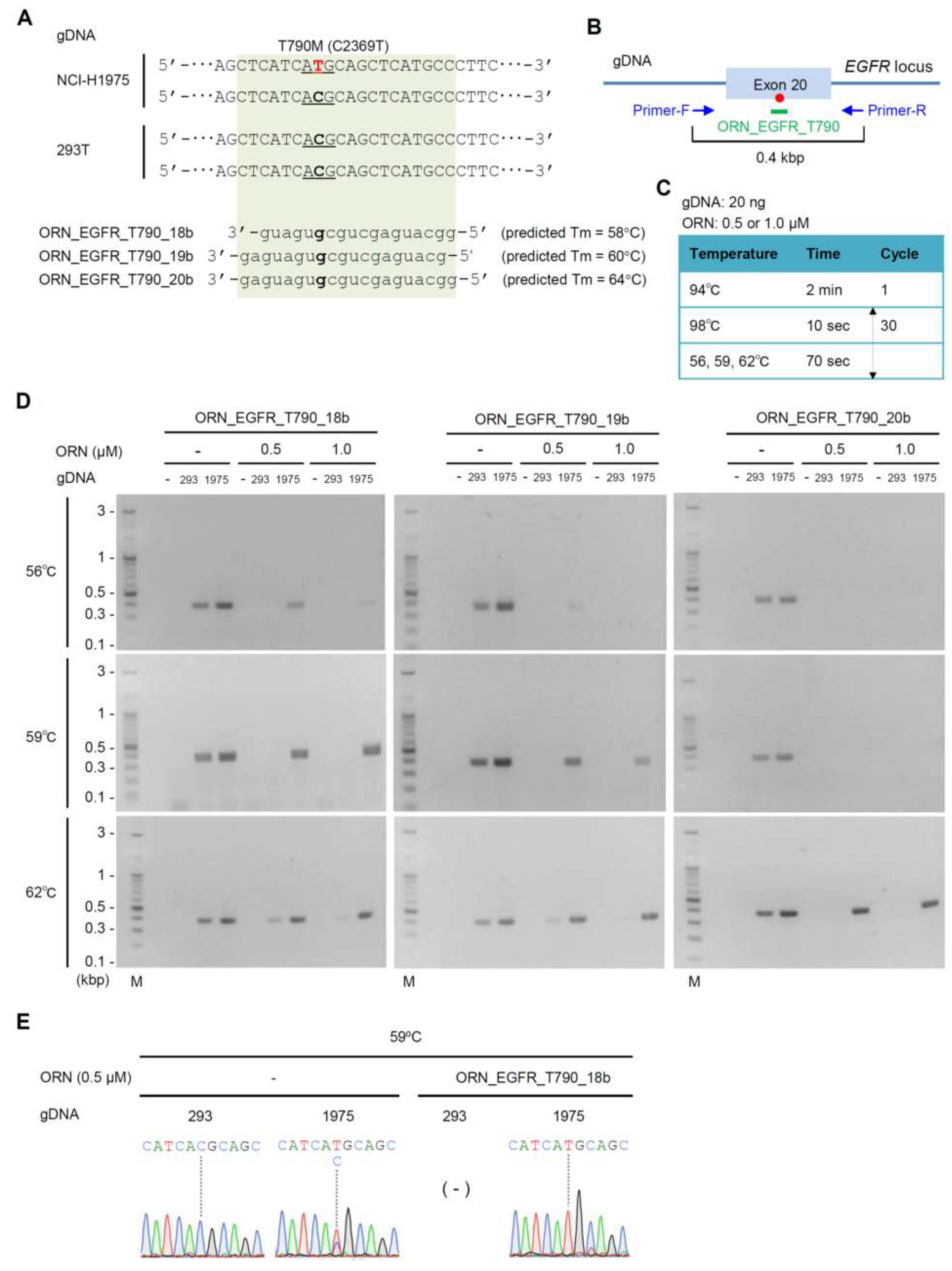
2.2. Detection of the Single-Nucleotide Mutation T790M (C2369T) by ORNi-PCR
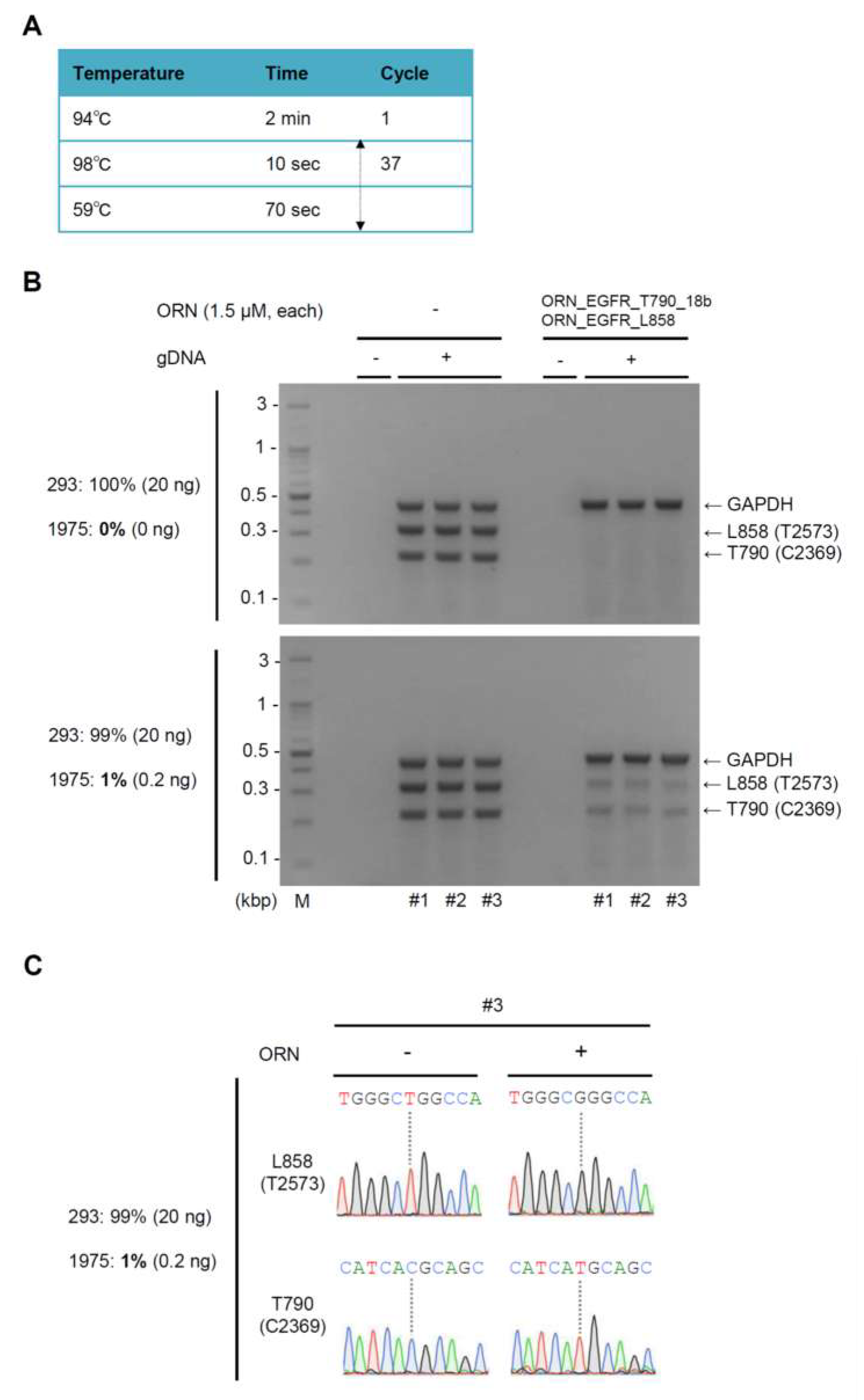
2.3. Simultaneous Detection of the T790M (C2369T) and L858R (T2573G) Mutations by Multiplex ORNi-PCR with gDNA
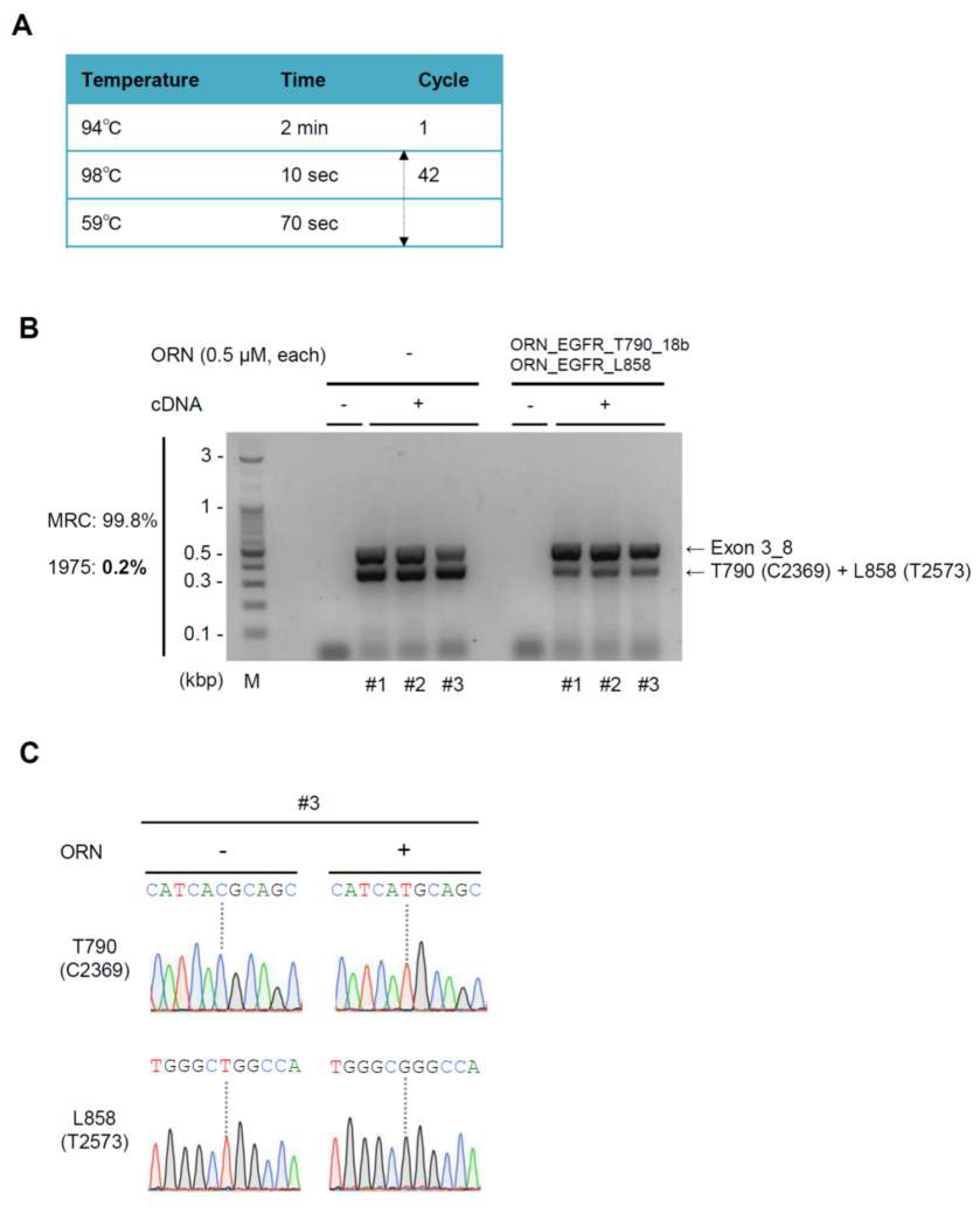
2.4. Simultaneous Detection of the T790M (C2369T) and L858R (T2573G) Mutations by ORNi-PCR with cDNA
2.5. Sensitivity of ORNi-PCR for Simultaneous Detection of Two EGFR Mutations
3. Discussion
4. Materials and Methods
4.1. Oligonucleotides
4.2. Cell Culture
4.3. ORNi-PCR with gDNA
4.4. Electrophoresis and DNA Sequencing Analysis
4.5. RNA Extraction, RT Reactions, and Two-Step ORNi-PCR with cDNA
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EGFR | Epidermal Growth Factor Receptor |
| gDNA | genomic DNA |
| cDNA | complementary DNA |
| PCR | Polymerase Chain Reaction |
| ORNi-PCR | Oligoribonucleotide interference-PCR |
References
- Lee, Y.T.; Tan, Y.J.; Oon, C.E. Molecular targeted therapy: treating cancer with specificity. Eur. J. Pharmacol. 2018, 834, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Krause, D.S.; Van Etten, R.A. Tyrosine kinases as targets for cancer therapy. N. Engl. J. Med. 2005, 353, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic signaling pathways in the cancer genome atlas. Cell 2018, 173, 321–337. [Google Scholar] [CrossRef]
- Paez, J.G.; Jänne, P.A.; Lee, J.C.; Tracy, S.; Greulich, H.; Gabriel, S.; Herman, P.; Kaye, F.J.; Lindeman, N.; Boggon, T.J.; et al. EGFR mutations in lung cancer: correlation with clinical response to gefitinib therapy. Science 2004, 304, 1497–1500. [Google Scholar] [CrossRef]
- Mok, T.S.; Wu, Y.L.; Thongprasert, S.; Yang, C.H.; Chu, D.T.; Saijo, N.; Sunpaweravong, P.; Han, B.; Margono, B.; Ichinose, Y.; et al. Gefitinib or carboplatin-paclitaxel in pulmonary adenocarcinoma. N. Engl. J. Med. 2009, 361, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Maemondo, M.; Inoue, A.; Kobayashi, K.; Sugawara, S.; Oizumi, S.; Isobe, H.; Gemma, A.; Harada, M.; Yoshizawa, H.; Kinoshita, I.; et al. Gefitinib or chemotherapy for non-small-cell lung cancer with mutated EGFR. N. Engl. J. Med. 2010, 362, 2380–2388. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Au, J.S.; Thongprasert, S.; Srinivasan, S.; Tsai, C.M.; Khoa, M.T.; Heeroma, K.; Itoh, Y.; Cornelio, G.; Yang, P.C. A prospective, molecular epidemiology study of EGFR mutations in Asian patients with advanced non-small-cell lung cancer of adenocarcinoma histology (PIONEER). J. Thorac. Oncol. 2014, 9, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Mitsudomi, T. Not all epidermal growth factor receptor mutations in lung cancer are created equal: perspectives for individualized treatment strategy. Cancer Sci. 2016, 107, 1179–1186. [Google Scholar] [CrossRef]
- Beau-Faller, M.; Prim, N.; Ruppert, A.M.; Nanni-Metéllus, I.; Lacave, R.; Lacroix, L.; Escande, F.; Lizard, S.; Pretet, J.L.; Rouquette, I.; et al. Rare EGFR exon 18 and exon 20 mutations in non-small-cell lung cancer on 10 117 patients: a multicentre observational study by the French ERMETIC-IFCT network. Ann. Oncol. 2014, 25, 126–131. [Google Scholar] [CrossRef]
- Oxnard, G.R. The cellular origins of drug resistance in cancer. Nat. Med. 2016, 22, 232–234. [Google Scholar] [CrossRef]
- Pao, W.; Miller, V.A.; Politi, K.A.; Riely, G.J.; Somwar, R.; Zakowski, M.F.; Kris, M.G.; Varmus, H. Acquired resistance of lung adenocarcinomas to gefitinib or erlotinib is associated with a second mutation in the EGFR kinase domain. PLoS Med. 2005, 2, e73. [Google Scholar] [CrossRef]
- Nguyen, K.S.; Kobayashi, S.; Costa, D.B. Acquired resistance to epidermal growth factor receptor tyrosine kinase inhibitors in non-small-cell lung cancers dependent on the epidermal growth factor receptor pathway. Clin. Lung Cancer 2009, 10, 281–289. [Google Scholar] [CrossRef]
- Mok, T.S.; Wu, Y.-L.; Ahn, M.-J.; Garassino, M.C.; Kim, H.R.; Ramalingam, S.S.; Shepherd, F.A.; He, Y.; Akamatsu, H.; Theelen, W.S.; et al. Osimertinib or platinum-pemetrexed in EGFR T790M-positive lung cancer. N. Engl. J. Med. 2017, 376, 629–640. [Google Scholar] [CrossRef]
- Soria, J.C.; Ohe, Y.; Vansteenkiste, J.; Reungwetwattana, T.; Chewaskulyong, B.; Lee, K.H.; Dechaphunkul, A.; Imamura, F.; Nogami, N.; Kurata, T.; et al. Osimertinib in untreated EGFR-mutated advanced non-small-cell lung cancer. N. Engl. J. Med. 2018, 378, 113–125. [Google Scholar] [CrossRef]
- Pytela, R.; Suzuki, S.; Breuss, J.; Erle, D.J.; Sheppard, D. Polymerase chain reaction cloning with degenerate primers: homology-based identification of adhesion molecules. Methods Enzymol. 1994, 245, 420–451. [Google Scholar]
- Vestheim, H.; Deagle, B.E.; Jarman, S.N. Application of blocking oligonucleotides to improve signal-to-noise ratio in a PCR. Methods Mol. Biol. 2011, 687, 265–274. [Google Scholar] [CrossRef]
- Dominguez, P.L.; Kolodney, M.S. Wild-type blocking polymerase chain reaction for detection of single nucleotide minority mutations from clinical specimens. Oncogene 2005, 24, 6830–6834. [Google Scholar] [CrossRef]
- Nagai, Y.; Miyazawa, H.; Huqun;Tanaka, T.; Udagawa, K.; Kato, M.; Fukuyama, S.; Yokote, A.; Kobayashi, K.; Kanazawa, M.; et al. Genetic heterogeneity of the epidermal growth factor receptor in non-small cell lung cancer cell lines revealed by a rapid and sensitive detection system, the peptide nucleic acid-locked nucleic acid PCR clamp. Cancer Res. 2005, 65, 7276–7282. [Google Scholar] [CrossRef]
- Tanigawa, N.; Fujita, T.; Fujii, H. Oligoribonucleotide (ORN) interference-PCR (ORNi-PCR): a simple method for suppressing PCR amplification of specific DNA sequences using ORNs. PLoS One 2014, 9, e113345. [Google Scholar] [CrossRef]
- Fujita, T.; Yuno, M.; Kitaura, F.; Fujii, H. Detection of genome-edited cells by oligoribonucleotide interference-PCR. DNA Res. 2018, 25, 395–407. [Google Scholar] [CrossRef]
- Fujita, T.; Yuno, M.; Kitaura, F.; Fujii, H. A refined two-step oligoribonucleotide interference-PCR method for precise discrimination of nucleotide differences. Sci. Rep. 2018, 8, 17195. [Google Scholar] [CrossRef]
- Zhang, S.R.; Xu, Y.S.; Jin, E.; Zhu, L.C.; Xia, B.; Chen, X.F.; Li, F.Z.; Ma, S.L. Capilliposide from Lysimachia capillipes inhibits AKT activation and restores gefitinib sensitivity in human non-small cell lung cancer cells with acquired gefitinib resistance. Acta Pharmacol. Sin. 2017, 38, 100–109. [Google Scholar] [CrossRef]
- Jacobs, J.P.; Jones, C.M.; Baille, J.P. Characteristics of a human diploid cell designated MRC-5. Nature 1970, 227, 168–170. [Google Scholar] [CrossRef]
- Li, J.; Deng, H.; Hu, M.; Fang, Y.; Vaughn, A.; Cai, X.; Xu, L.; Wan, W.; Li, Z.; Chen, S.; et al. Inhibition of non-small cell lung cancer (NSCLC) growth by a novel small molecular inhibitor of EGFR. Oncotarget 2015, 6, 6749–6761. [Google Scholar] [CrossRef]
- Sutani, A.; Nagai, Y.; Udagawa, K.; Uchida, Y.; Koyama, N.; Murayama, Y.; Tanaka, T.; Miyazawa, H.; Nagata, M.; Kanazawa, M.; et al. Gefitinib for non-small-cell lung cancer patients with epidermal growth factor receptor gene mutations screened by peptide nucleic acid-locked nucleic acid PCR clamp. Br. J. Cancer 2006, 95, 1483–1489. [Google Scholar] [CrossRef]
- Goto, K.; Satouchi, M.; Ishii, G.; Nishio, K.; Hagiwara, K.; Mitsudomi, T.; Whiteley, J.; Donald, E.; McCormack, R.; Todo, T. An evaluation study of EGFR mutation tests utilized for non-small-cell lung cancer in the diagnostic setting. Ann. Oncol. 2012, 23, 2914–2919. [Google Scholar] [CrossRef]
- Inoue, Y.; Shiihara, J.; Miyazawa, H.; Ohta, H.; Higo, M.; Nagai, Y.; Kobayashi, K.; Saijo, Y.; Tsuchida, M.; Nakayama, M.; et al. A highly specific and sensitive massive parallel sequencer-based test for somatic mutations in non-small cell lung cancer. PLoS ONE 2017, 12, e0176525. [Google Scholar] [CrossRef]
- Tanaka, T.; Matsuoka, M.; Sutani, A.; Gemma, A.; Maemondo, M.; Inoue, A.; Okinaga, S.; Nagashima, M.; Oizumi, S.; Uematsu, K.; et al. Frequency of and variables associated with the EGFR mutation and its subtypes. Int. J. Cancer 2010, 126, 651–655. [Google Scholar] [CrossRef]
- Yamada, N.; Oizumi, S.; Asahina, H.; Shinagawa, N.; Kikuchi, E.; Kikuchi, J.; Sakakibara-Konishi, J.; Tanaka, T.; Kobayashi, K.; Hagiwara, K.; et al. The peptide nucleic acid-locked nucleic acid polymerase chain reaction clamp-based test for epidermal growth factor receptor mutations in bronchoscopic cytological specimens of non-small cell lung cancer. Oncology 2012, 82, 341–346. [Google Scholar] [CrossRef]
- Shakeel, S.; Karim, S.; Ali, A. Peptide nucleic acid (PNA)—a review. J. Chem. Technol. Biotechnol. 2006, 81, 892–899. [Google Scholar] [CrossRef]
- Seyama, T.; Ito, T.; Hayashi, T.; Mizuno, T.; Nakamura, N.; Akiyama, M. A novel blocker-PCR method for detection of rare mutant alleles in the presence of an excess amount of normal DNA. Nucleic Acids Res. 1992, 20, 2493–2496. [Google Scholar] [CrossRef]
- Hao, W.; Fan, L.; Chen, Q.; Chen, X.; Zhang, S.; Lan, K.; Lu, J.; Zhang, C. Modified proofreading PCR for detection of point mutations, insertions and deletions using a ddNTP-blocked primer. PLoS One 2015, 10, e0123468. [Google Scholar] [CrossRef]
- Shibata, H.; Tahira, T.; Hayashi, K. RNA-primed PCR. Genome Res. 1995, 5, 400–403. [Google Scholar] [CrossRef]
- Miyazawa, H.; Tanaka, T.; Nagai, Y.; Matsuoka, M.; Huqun;Sutani, A.; Udagawa, K.; Zhang, J.; Hirama, T.; Murayama, Y.; Koyama, N.; et al. Peptide nucleic acid-locked nucleic acid polymerase chain reaction clamp-based detection test for gefitinib-refractory T790M epidermal growth factor receptor mutation. Cancer Sci. 2008, 99, 595–600. [Google Scholar] [CrossRef]
- Watanabe, M.; Hashida, S.; Yamamoto, H.; Matsubara, T.; Ohtsuka, T.; Suzawa, K.; Maki, Y.; Soh, J.; Asano, H.; Tsukuda, K.; et al. Estimation of age-related DNA degradation from formalin-fixed and paraffin-embedded tissue according to the extraction methods. Exp. Ther. Med. 2017, 14, 2683–2688. [Google Scholar] [CrossRef]
- Suzuki, M.; Makinoshima, H.; Matsumoto, S.; Suzuki, A.; Mimaki, S.; Matsushima, K.; Yoh, K.; Goto, K.; Suzuki, Y.; Ishii, G.; et al. Identification of a lung adenocarcinoma cell line with CCDC6-RET fusion gene and the effect of RET inhibitors in vitro and in vivo. Cancer Sci. 2013, 104, 896–903. [Google Scholar] [CrossRef]
- Ohtsuka, K.; Ohnishi, H.; Fujiwara, M.; Kishino, T.; Matsushima, S.; Furuyashiki, G.; Takei, H.; Koshiishi, Y.; Goya, T.; Watanabe, T. Abnormalities of epidermal growth factor receptor in lung squamous-cell carcinomas, adenosquamous carcinomas, and large-cell carcinomas: tyrosine kinase domain mutations are not rare in tumors with an adenocarcinoma component. Cancer 2007, 109, 741–750. [Google Scholar] [CrossRef]
- Chan, D.L.H.; Segelov, E.; Wong, R.S.; Smith, A.; Herbertson, R.A.; Li, B.T.; Tebbutt, N.; Price, T.; Pavlakis, N. Epidermal growth factor receptor (EGFR) inhibitors for metastatic colorectal cancer. Cochrane Database Syst. Rev. 2017, 6, CD007047. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baba, K.; Fujita, T.; Tasaka, S.; Fujii, H. Simultaneous Detection of the T790M and L858R Mutations in the EGFR Gene by Oligoribonucleotide Interference-PCR. Int. J. Mol. Sci. 2019, 20, 4020. https://doi.org/10.3390/ijms20164020
Baba K, Fujita T, Tasaka S, Fujii H. Simultaneous Detection of the T790M and L858R Mutations in the EGFR Gene by Oligoribonucleotide Interference-PCR. International Journal of Molecular Sciences. 2019; 20(16):4020. https://doi.org/10.3390/ijms20164020
Chicago/Turabian StyleBaba, Keisuke, Toshitsugu Fujita, Sadatomo Tasaka, and Hodaka Fujii. 2019. "Simultaneous Detection of the T790M and L858R Mutations in the EGFR Gene by Oligoribonucleotide Interference-PCR" International Journal of Molecular Sciences 20, no. 16: 4020. https://doi.org/10.3390/ijms20164020
APA StyleBaba, K., Fujita, T., Tasaka, S., & Fujii, H. (2019). Simultaneous Detection of the T790M and L858R Mutations in the EGFR Gene by Oligoribonucleotide Interference-PCR. International Journal of Molecular Sciences, 20(16), 4020. https://doi.org/10.3390/ijms20164020